Introduction
Cervical cancer (CC) is the fourth most common
cancer affecting women worldwide, with an annual mortality of
~270,000 (1,2). Standard therapies for CC, including
radiotherapy, chemotherapy and surgical intervention, are often
less effective for patients in the advanced stages of disease
(3). The mechanisms of CC remain
unclear and gaining a better understanding of tumor progression and
identifying novel biomarkers for the disease is of great
importance.
MicroRNAs (miRNAs or miRs) are a group of small
non-coding RNAs (~22–25 nucleotides in length) that negatively
regulate their target genes via binding to complementary sequences
of the 3′-untranslated region (3′-UTR) (4). A number of studies have revealed that
miRNAs serve important roles in the development of CC and are
associated with tumor suppressing and oncogenic activities
(5). miR-218 is a miRNA that has a
tumor-suppressing effect in a number of cancers (6–8). The
precursors of miR-218 are mir-218-1 and mir-218-2, which are
located in the Slit2 and Slit3 introns and can be expressed
together with Slit2 and Slit3 (6–7,9). Expression of the miR-218 precursors is
downregulated in several human malignant tumors and has a
significant tumor suppressive effect (7,10–12).
miR-218 has been reported to be downregulated in
tumor tissues and sera of patients with CC (12). Additionally, miR-218 downregulation
is associated with tumor invasion (13). A number of studies have reported that
the expression of miR-218, which serves a tumor-suppressive role in
CC, is affected by human papillomavirus (HPV) infection (14–19). A
previous study on HPV, a major carcinogenic risk factor for CC,
reported that the expression of miR-218 and its host gene tumor
suppressor gene SLIT2 was lower in HPV-16 and −18 positive cells
compared with normal cervical tissue and HPV negative cells
(20). It was suggested that the
expression of miR-218 in CC tissues was lower compared with normal
cervical tissues and that there are certain strains of HPV that are
associated with a greater risk of CC, may result in further
downregulation (20). miR-218 is
downregulated in a number of malignant tumors, including lung
cancer, breast cancer, thyroid cancer, mesenteric adenocarcinoma,
colon cancer, gastric cancer, prostate cancer, glioblastoma and
bladder cancer (21). Low miR-218
expression was demonstrated to be associated with tumor cell
invasion and migration, and poor prognosis (7,22,23).
Furthermore, miR-218 expression is lower in malignant tumors
compared with benign tumors (24).
Low miR-218 expression is associated with tumor invasion, migration
and poor prognosis (7,21–24).
Together, these studies suggest that miR-218 serves an important
role in the development of tumors. Although these studies link
miR-218 with tumor metastasis, the specific mechanisms of miR-218
in tumors remain unclear and require further study. The aim of the
present study was to investigate the role and mechanism of miR-218
in the metastasis of CC.
Gli3 serves a key and complex role in the occurrence
and development of tumors, participating in multi-gene, multi-step
regulatory processes (25). Bai
et al (25) reported that
Gli3 full-length protein (Gli3-FL) was found in the cytoplasm and
Gli3 terminal removal protein (Gli3-TR) was found in the nucleus of
tumor epithelial cells in small cell lung cancer. Gli3-FL and
Gli3-TR are highly expressed in lung cancer; Gli3-FL is not
associated with clinic pathological parameters and survival rate,
while Gli3-TR is associated with lymph node metastasis and overall
survival. Gli3-TR, together with tumor differentiation and disease
stage, can be used as independent prognostic indicators in patients
with lung cancer.
In the present study, Gli3 was confirmed as a
potential target gene of miR-218 via a series of experiments. The
regulatory mechanisms of miR-218 and Gli3 in CC were also
investigated and it was demonstrated that miR-218 regulates the
proliferation and apoptosis of CC cells by targeting to Gli3 in
vitro. These results provide a theoretical basis for miR-218 as
a target for CC progression and treatment
Materials and methods
Clinical samples
A total of 112 CC tissues and corresponding adjacent
normal tissues were obtained from patients with cervical cancer
(age range, 25–55 years old), who had not undergone radiotherapy or
chemotherapy, in the First Affiliated Hospital of Bengbu Medical
College (Bengbu, China) from 2014 to 2016. All tissue samples were
immediately frozen in liquid nitrogen and stored at −80°C until
use. All participants provided signed statements of informed
consent according to the principles of the Declaration of Helsinki.
The study protocol was approved by the Ethics Committee of the
First Affiliated Hospital of Bengbu Medical College.
Cell lines and transfection
A human cervical epithelial cell line (HCvEpCs) and
Hela, SiHa and C33A CC cell lines were obtained from the American
Type Culture Collection (Manassas, VA, USA). Cells were grown in
Dulbecco's Modified Eagle's medium (DMEM) supplemented with 10%
heat-inactivated fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C in an atmosphere
containing 5% CO2. SiHa cells were seeded in 96-well
plates at a density of 2×105 cells/well and grown to 70%
confluence. Cells were then transfected with negative control miRNA
(miR-NC), miR-218 mimics or miR-218 inhibitor using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Subsequently, the transfected cells were
incubated for a further 72 h. The miR-218 sequences transfected
were 5′-TGCATGGTTAGATCAAGCACAAGGG-3′ and
5′-CTCGTCTTATTTCCGTGACTGTTTT-3′.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was performed to measure the expression of
miR-218 in CC cell lines and frozen tissue specimens. miRNAs were
isolated using TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.) and the concentration of total RNA was measured.
Reverse transcription of the extracted RNA was completed by the
two-step method using a SuperScript™ RT-PCR System with Platinum™
Taq DNA Polymerase kit (Fermentas; Thermo Fisher Scientific,
Inc.). The reaction conditions were as follows: 70°C for 10 min,
4°C for 2 min, 42°C for 60 min, 70°C for 10 min and put temporary
on hold at −80°C. The expression of miR-218, Ki67 and Gli3 was
measured using the TaqMan miRNA assay kit (Invitrogen; Thermo
Fisher Scientific, Inc.). The PCR thermocycling conditions were as
follows: 97°C for 5 min, followed by 35 cycles at 95°C for 30 sec,
65°C for 30 sec and 73°C for 1 min, 73°C for 10 min, 4°C forever.
PCR primers used were as follows: Gli3, forward
5′-TGGTTACATGGAGCCCCACTA-3′ and revers
5′-GAATCGGAGATGGATCGTAATGG-3′; KI67, forward
5′-GCCTGCTCGACCCTACAGA-3′ and reverse 5′-GCTTGTCAACTGCGGTTGC-3′;
cleaved-caspase-3, forward 5′-AGAGGGGATCGTTGTAGAAGTC-3′ and reverse
5′-ACAGTCCAGTTCTGTACCACG-3′; cleaved-poly ADP-ribose polymerase
(PARP), forward 5′-TGGAAAAGTCCCACACTGGTA-3′ and reverse
5′-AAGCTCAGAGAACCCATCCAC-3′; cyclin B1, forward
5′-AATAAGGCGAAGATCAACATGGC-3′ and reverse
5′-TTTGTTACCAATGTCCCCAAGAG-3′; cyclin D1, forward
5′-CAATGACCCCGCACGATTTC-3′ and reverse 5′-CATGGAGGGCGGATTGGAA-3′;
and GAPDH, forward 5′-CTGGGCTACACTGAGCACC-3′ and reverse
5′-AAGTGGTCGTTGAGGGCAATG-3′. The relative expression of miR-218,
Ki67, Gli3, cleaved-caspase-3, cleaved-PARP, cyclin B1 and cyclin
D1 was calculated. The 2−ΔΔCq method was used for
relative quantification (26). U6
was used as an endogenous control for miRNA quantification and
GAPDH was used as an internal reference.
Cell proliferation analysis
A Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) assay was performed to assess
the proliferative ability of SiHa cells. At 24, 48 and 72 h
following transfection, 2×103 cells/well were seeded
into 96-well plates and 10 µl of CCK-8 solution was added to assess
cell viability according to the manufacturer's protocol. The
optical density (OD) was measured at 450 nm using a plate
reader.
Apoptosis analysis
To elucidate the mechanisms of miR-218 in CC cells,
a flow cytometry assay was performed to measure apoptosis.
Harvested cells were incubated with Annexin V-phycoerythrin
(PE)/7-amino-actinomycin D (7-AAD; KGA1015; Nanjing KeyGen Biotech
Co., Ltd., Nanjing, China) for 15 min at room temperature and
samples were analyzed using FACSCalibur flow cytometry (BD
Biosciences, Franklin Lakes, NJ, USA).
Cell cycle analysis
Cell cycle distribution was assessed using flow
cytometry. Following transfection, SiHa cells were harvested by
trypsinization and washed twice with ice-cold PBS, following which
they were fixed with 70% ethanol at 4°C overnight. Fixed cells were
rehydrated in PBS and stained with Annexin V-PE/7-AAD for 15 min at
room temperature in the dark and analyzed using FACSCalibur.
Subsequently, the percentage of cells in the G1, S and G2 phase was
measured using PV ELITE software (version 18.0; Integraph
Corporation, Madison, AL, USA).
Dual luciferase reporter assay
A luciferase reporter assay was performed using a
Dual-Luciferase Reporter Assay reagent (Promega, Madison, WI, USA)
according to the manufacturer's instructions. A total of
3.5×104 SiHa cells/well were seeded in 24-well plates
overnight and pGL3-Gli3-3′UTR-wild type (WT) or
pGL3-Gli3-3′UTR-mutant (mut) and miR-218 or miR-control vectors
were co-transfected into the cells using Lipofectamine®
2000. Three independent experiments were performed and data are
presented as the mean ± standard deviation.
Bioinformatics analysis
Bioinformatics analysis with TargetScan (version
6.2; http://genes.mit.edu/targetscan) was
conducted to predict its target gene.
Western blotting
Western blotting was performed to measure the
expression of Gli3, cleaved-caspase-3 and cleaved-PARP proteins.
SiHa cells were homogenized in lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) and the total protein was extracted
by centrifuged at 16,000 × g at 4°C for 5 min. The proteins (20
µg/lane) were quantified according to the protocol of the BCA kit
(cat. no. AR0146; Wuhan Boshide Biological Engineering Co. Ltd.,
Wuhan, China), separated by 10% SDS-PAGE and transferred onto a
polyvinylidene fluoride membrane. Membranes were subsequently
incubated with primary antibodies against Gli3 (1:1,000, PA5-19822;
Invitrogen, Thermo Fisher Scientific, Inc.), cleaved-caspase-3
(1:500; ab49822) cleaved-PARP (1:1,000; ab32064), cyclin B1 (1:500;
ab72), cyclin D1 (1:10,000; ab134175; all Abcam, Cambridge, UK) and
GAPDH (1:1,000 dilution; MA5-15738; Invitrogen; Thermo Fisher
Scientific, Inc.) for 1 h at room temperature. GAPDH was the
internal control. Membranes were then incubated with horseradish
peroxidase-conjugated anti-mouse (1:1,000; ab131368) and
anti-rabbit (1:1,000; ab191866; Abcam) antibodies at room
temperature for 2 h. Finally, membranes were incubated with BeyoECL
Plus (Beyotime Institute of Biotechnology) and the detected using
Chemi Doc™ XRS+ imaging system (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Statistical analysis
All statistical analyses were performed using
GraphPad Prism 6.0 software (GraphPad Software, Inc., La Jolla, CA,
USA). Differences between two groups were analyzed using a
Student's t-test. Differences between multiple groups were assessed
using one-way analysis of variance followed by the Dunnett's
post-hoc test. The correlation between miR-218 expression and other
genes mRNA expression levels was examined using Pearson's
correlation analysis. Each experiment was repeated ≥3 times. Values
were presented as the mean ± standard deviation and P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-218 expression is negatively
correlated with Ki67 and Gli3 in CC tissues
The expression of miR-218 was decreased in the CC
cell lines and tissues compared with the HCvEpCs and normal
tissues, respectively (Fig. 1A and
B). Ki67 and Gli3 were also downregulated in CC tissues
compared with normal tissues (Fig.
1C). The correlation between miR-218 and Ki67 and Gli3
expression was assessed using Pearson's correlation analysis and
the results revealed that miR-218 was negatively correlated with
Ki67 and Gli3 (Fig. 1D).
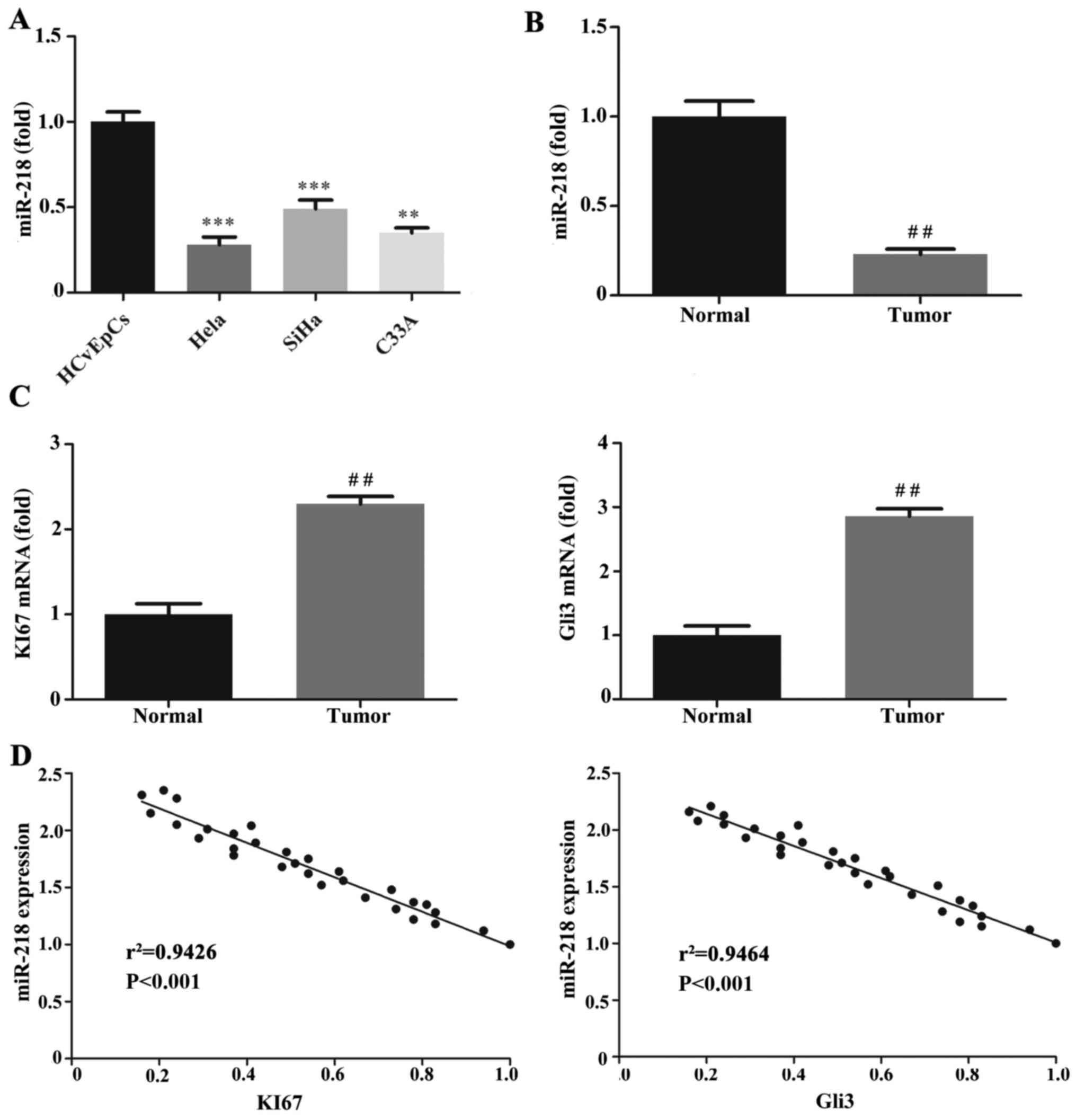 | Figure 1.Expression of miR-218, KI67 and Gli3
mRNA. RT-qPCR was performed to quantify miR-218 expression in (A)
HCvEpCs, Hela, SiHa and C33A cells and (B) CC tumor and normal
cervical tissues. (C) RT-qPCR was performed to measure KI67 and
Gli3 mRNA expression in CC tumor and normal tissues. (D) Pearson's
correlation analysis of miR-218 expression with KI67 or Gli3 mRNA
in 112 cervical cancer tissues. KI67, r2=0.9426; Gli3,
r2=0.9464. **P<0.01, ***P<0.001 vs. HCvEpCs
##P<0.01 vs. Normal. miR, microRNA; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; HCvEpCs,
normal human cervical cells; CC, cervical cancer. |
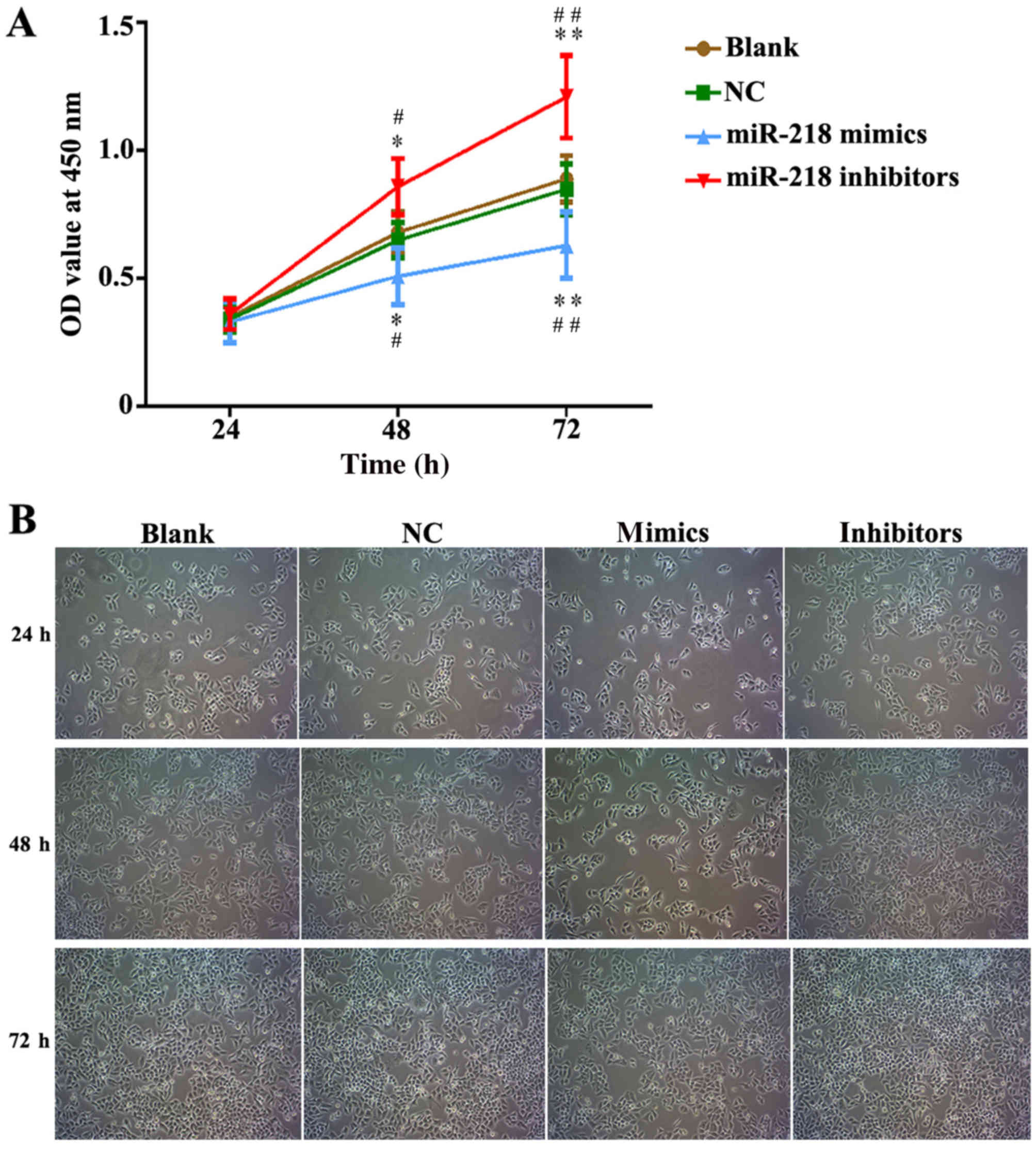
miR-218 suppresses CC proliferation,
cell cycle progression and apoptosis in vitro
In order to explore the potential role of miR-218 in
the pathogenesis of CC, SiHa cells were transfected with miR-218
mimics, NC vectors and inhibitors. The results of a cell viability
assay revealed that miR-218 overexpression dramatically suppressed
the proliferation of SiHa cells compared with the miR-NC group,
while proliferation was significantly suppressed in the inhibitors
group (Fig. 2A). Cell morphology was
affected as presented in Fig. 2B;
the number of cell invasions decreased in miR-218 mimics group and
increased in the miR-218 inhibitors group compared with the NC
group. Furthermore, the results of flow cytometry revealed that the
percentage of cells in the G0/G1 phase was
increased in the miR-218 mimics group and decreased in the miR-218
inhibitors group (Fig. 3A).
Furthermore, the rate of apoptosis was increased in the miR-218
mimics group, while apoptosis was decreased in the inhibitors group
(Fig. 3B). The quantified flow
cytometry results demonstrated that a significantly greater number
of cells treated with the inhibitor were in the G1 phase
compared with the NC and Blank groups (Fig. 3C) and that apoptosis was
significantly increased in the mimc group and decreased in the
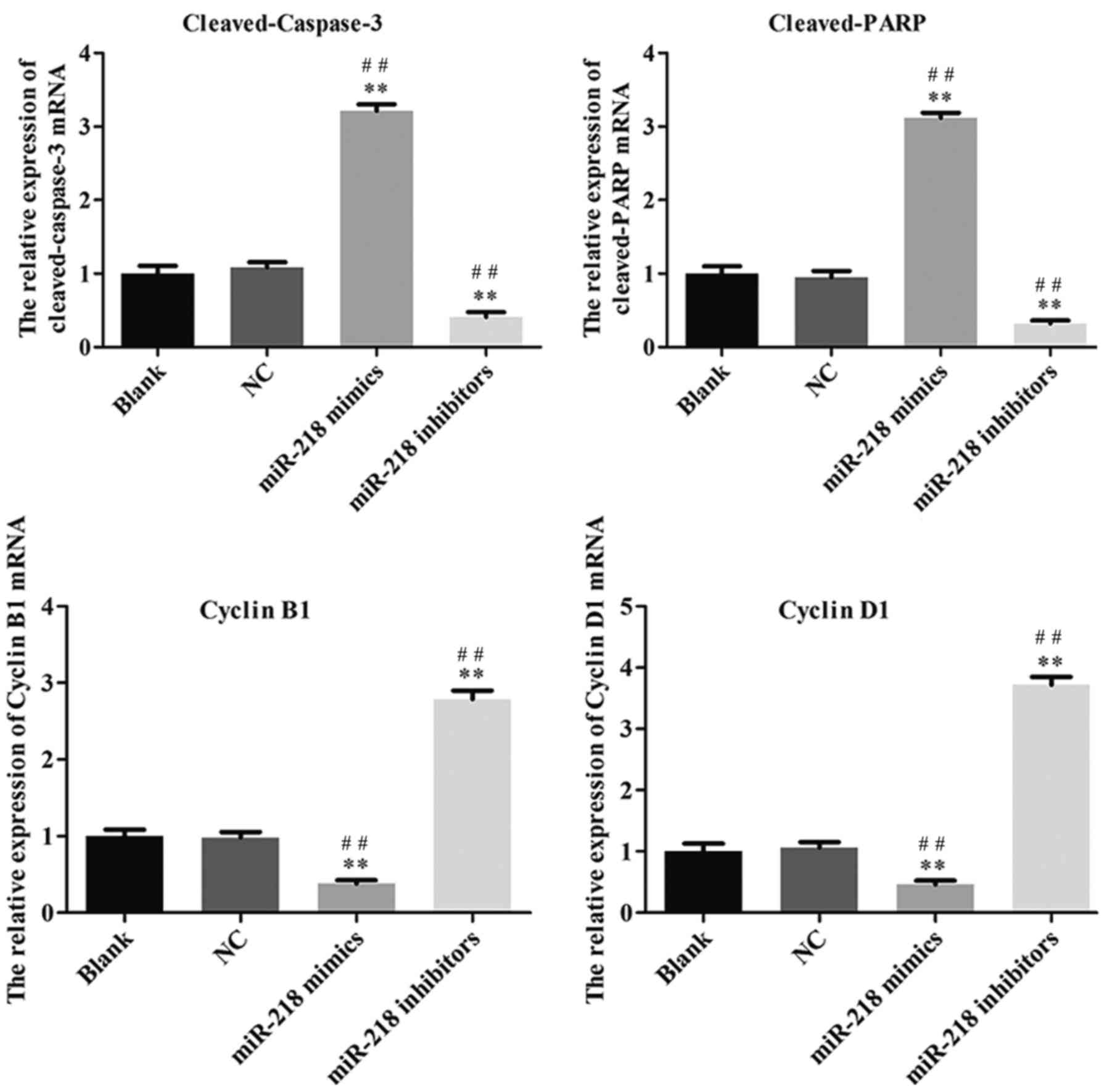
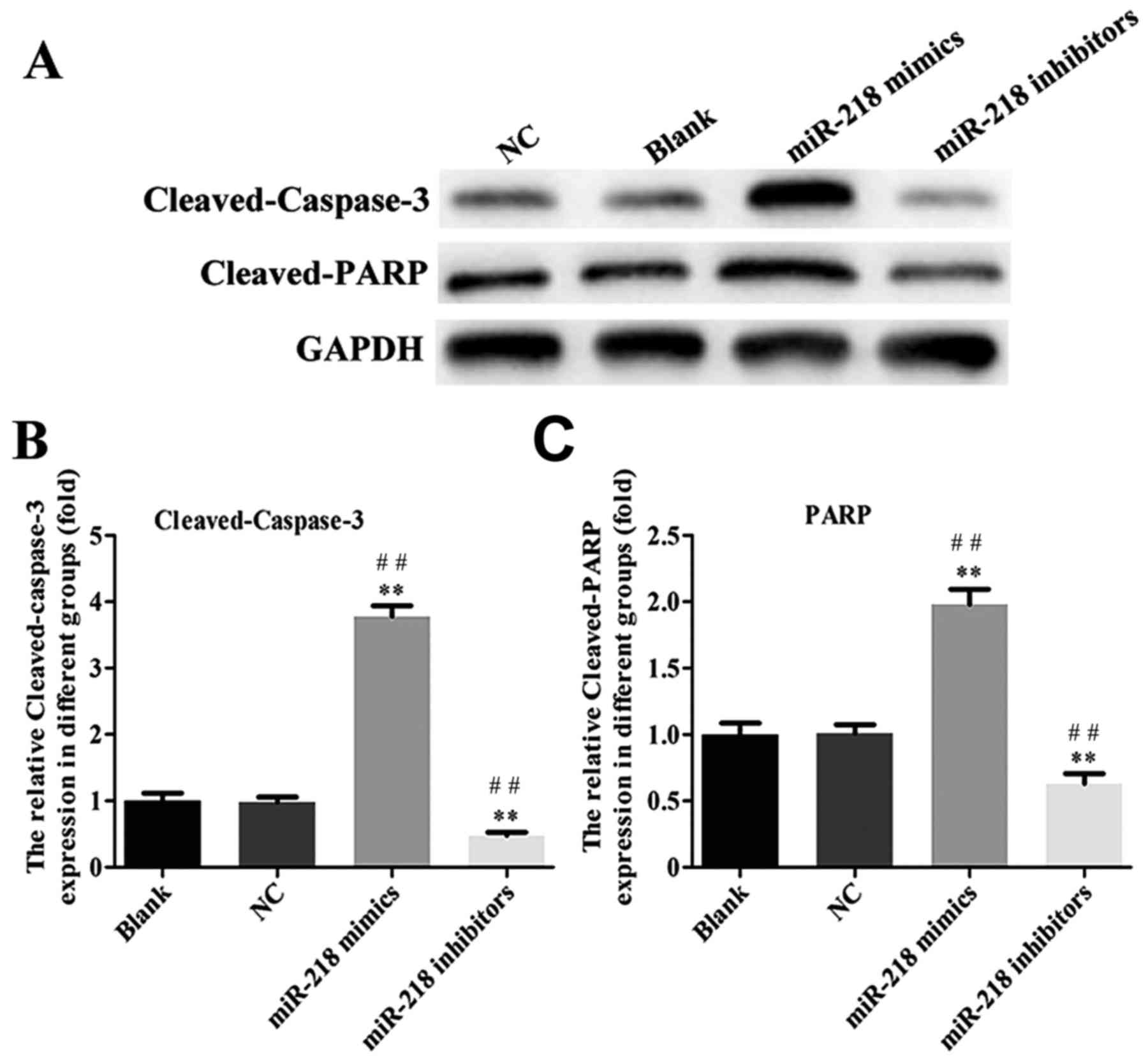
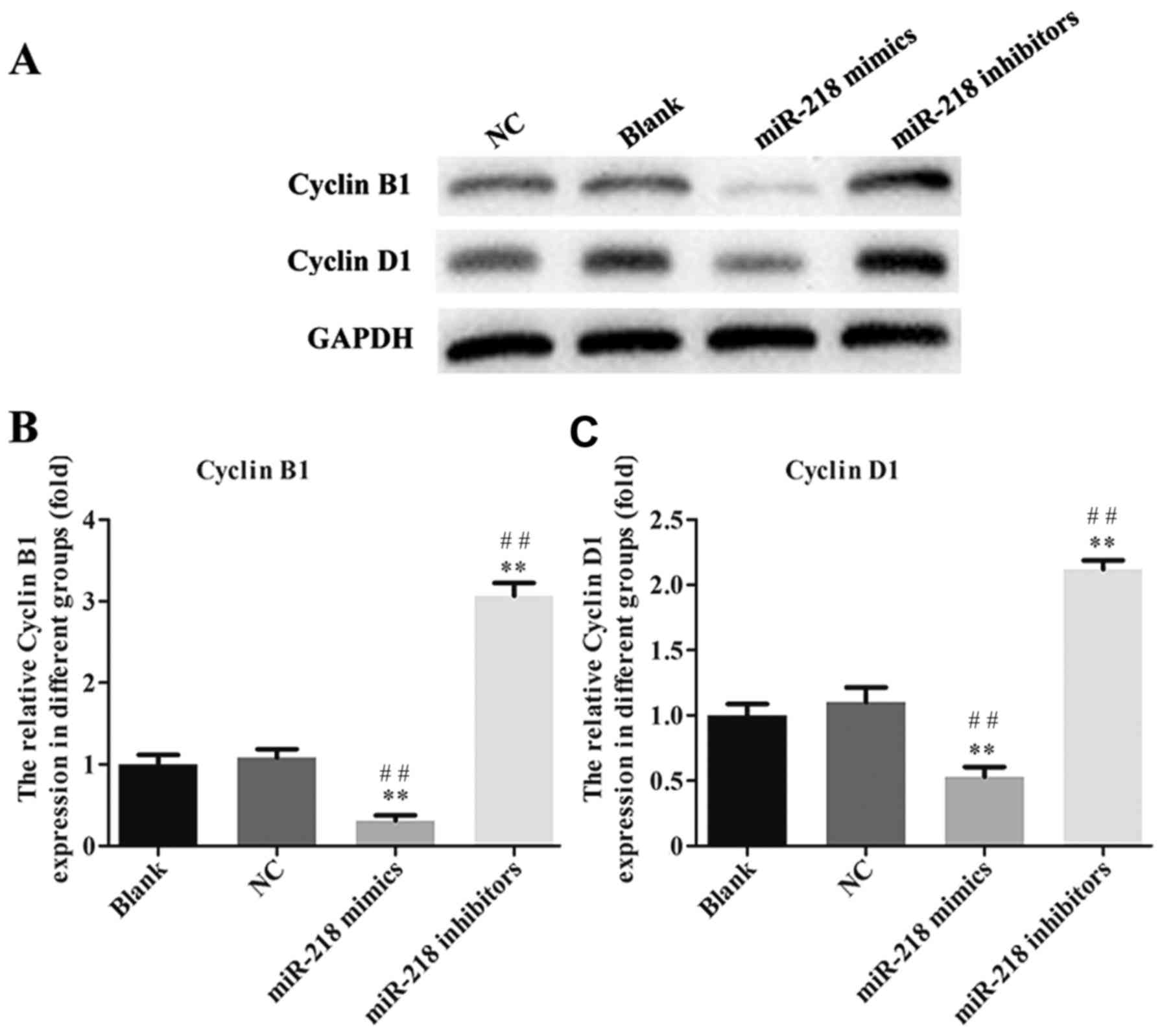
inhibitor group compared with the NC and Blank groups (Fig. 3D). The expression of
cleaved-caspase-3, cleaved-PARP, cyclin and cyclin D1 was assessed
in transfected SiHa cells. PCR results confirmed that the
expression of cleaved-caspase-3 and cleaved-PARP was increased
while the expression of cyclin B1 and cyclin D1 was decreased in
the miR-218-mimics group at the mRNA (Fig. 4) and protein (Figs. 5 and 6) level. The opposite was observed in the
miR-218 inhibitors group (Figs.
4–6).
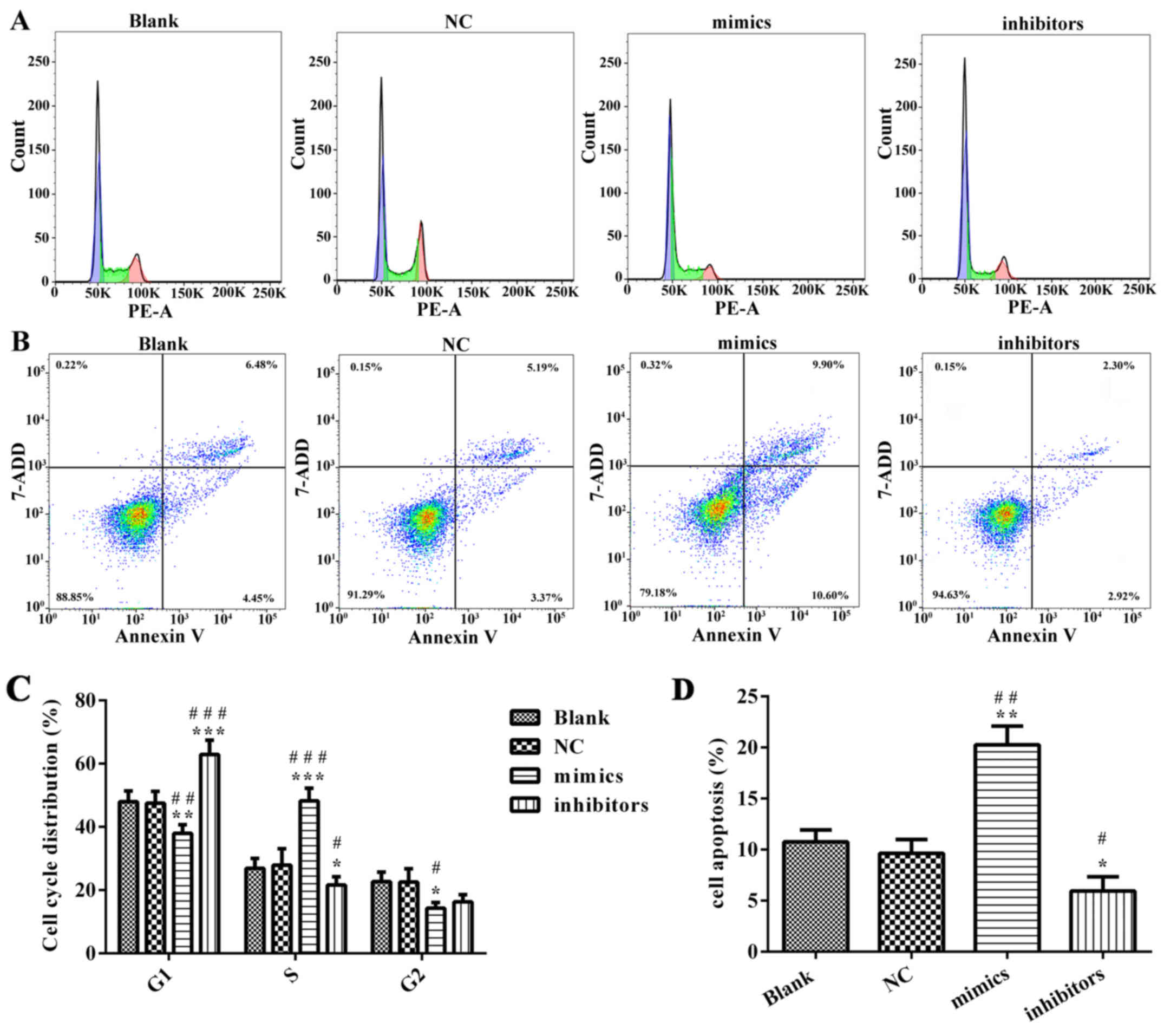 | Figure 3.miR-218 suppresses cervical cancer
cell cycle progression and apoptosis in vitro. (A) After
SiHa cells were transfected with miR-218 inhibitors or mimics, flow
cytometry was performed to test the cell cycle progression. (B)
Apoptosis was evaluated using flow cytometry. (C) Quantification of
cell cycle distribution, including G1-phase, G0-phase and S-phase.
(D) Quantification of cell apoptosis. *P<0.05, **P<0.01,
***P<0.001 vs. NC. #P<0.05,
##P<0.01, ###P<0.001 vs. Blank. miR,
microRNA; NC, negative control; PE, phycoerythrin; 7-AAD,
7-aminonactinomycin D. |
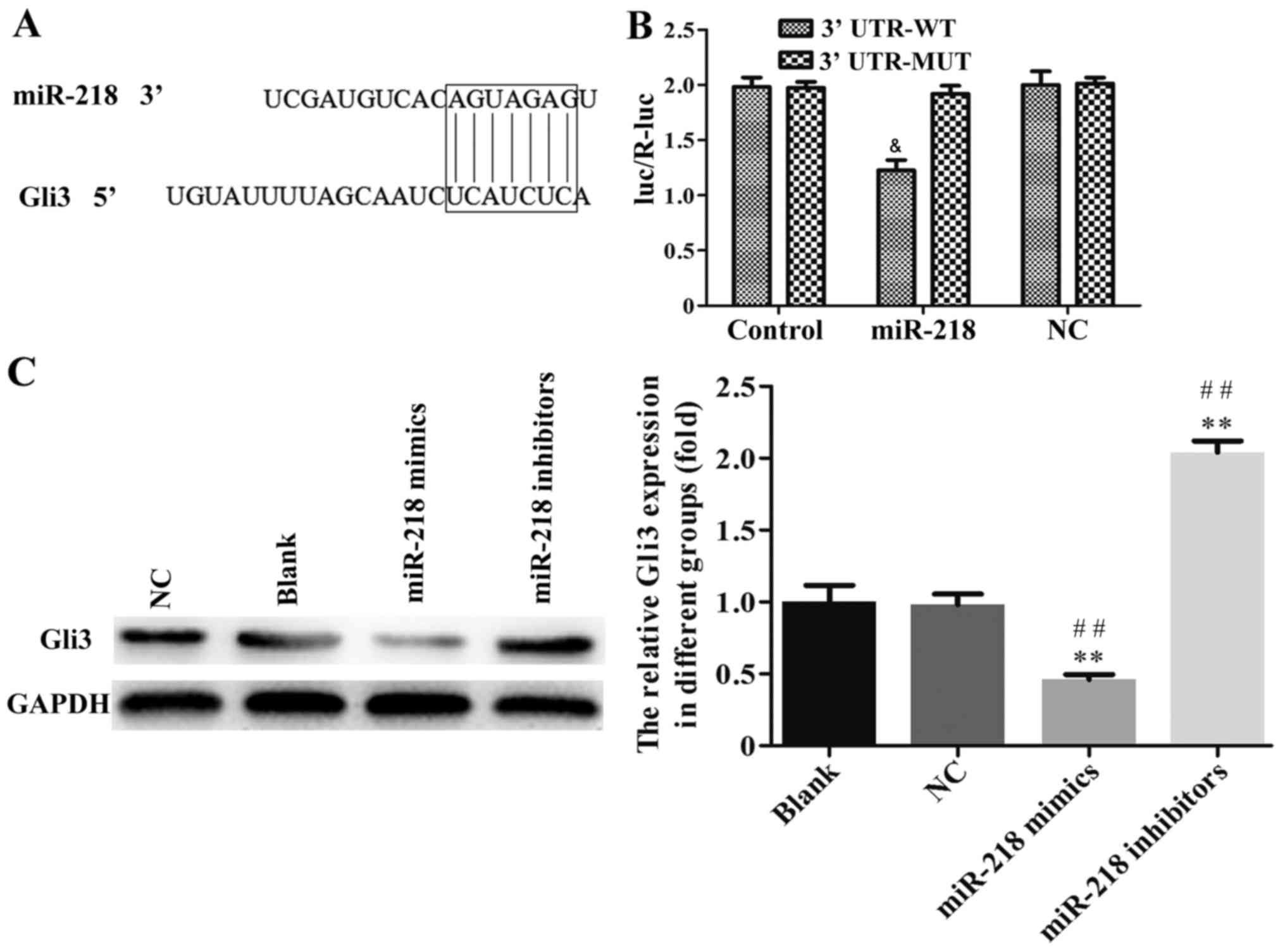
Gli3 is an important target of miR-218
in CC cells
To further investigate the underlying mechanism of
miR-218 in the progression of CC, TargetScan 6.2 was used to
predict its target gene. As shown in the Fig. 7A, Gli3 may be a target of miR-218.
The results of the luciferase activity assay indicated that miR-218
overexpression suppressed the activity of Gli3 WT-3′-UTR in SiHa
cells compared with NC group (Fig.
7B). The association between miR-218 and Gli3 expression was
further confirmed by western blotting and it was demonstrated that
the expression of Gli3 protein was significantly downregulated in
the miR-218 mimics group and upregulated in the miR-218 inhibitor
group compared with the NC group (Fig.
7C).
Discussion
CC is one of the most prevalent and deadly
malignancies in women worldwide (27,28). A
World Health Organization report revealed that >270,000 women
succumbed to CC in 2014 and >85% of cases occur in less
developed regions (28). The
morbidity and mortality of CC are high (29) and the age of onset is a decreasing
(30,31). In the past few decades, great
progress has been made researching treatments for CC (32,33).
However, there remains a challenge to improve the survival of CC
patients so far.
miRNAs have been reported to serve a role in the
initiation, progression and metastasis of CC via regulating cancer
cell proliferation, apoptosis, cell cycle arrest, migration and
invasion (34,35). As a member of the miRNA family,
miR-218 is correlated with tumor progression and poor prognosis
(36,37). It has been reported that miR-218
binds to its target mRNA in order to inhibit or promote the
formation and development of tumors (38). In previous studies, miR-218
downregulation was identified in the tumor tissues and sera of
patients with CC (39). However, it
remains unclear whether miR-218 expression is downregulated in CC
cells.
The aim of the present study was to investigate the
pivotal roles of miR-218 in the progression of CC. The expression
of miR-218 in CC tissues and cells was examined and it was
demonstrated that miR-218 was significantly decreased in CC tumor
tissues compared with adjacent normal tissues. These results are
consistent with those observed in CC and normal cervical cell
lines. The expression of Ki67 mRNA in CC tissues was demonstrated
to be increased and a negative correlation was observed between
Ki67 and miR-218. The present study also revealed that, compared
with adjacent tissues, Ki67 expression was upregulated in CC
tissues, suggesting that miR-218 was negatively correlated with the
proliferation marker Ki67. Following transfection with miR-218
mimics, the proliferation ability of CC cells was suppressed,
suggesting that miR-218 inhibits the proliferation of CC cells and
its downregulation serves a role in the development and progression
of CC. Gli3 was predicted and confirmed as a gene target of miR-218
and its expression in CC tissues and transfected CC cell lines was
assessed. A previous study indicated that the downregulated
expression of Glli3 attenuates pancreatic cancer cell activity and
proliferation (40). Gli3 is
expressed in a subset of colon cancers and associated with the
degree of tumor differentiation (40). Gli3 signaling significantly enhances
the tumorigenicity of colon cancer (41). Kang et al (42) reported that, compared with normal
tissues, Gli3 is upregulated in colon cancer tissues and Gli3
downregulation inhibits cancer cell proliferation. The
downregulated of Gli3 expression was also reported to enhance the
sensitivity of colon cancer cells to 5-FU (42). Together, these results suggest that
the role of Gli3 in tumor tissue is complex. In the present study,
Gli3 mRNA and protein expression decreased dramatically and was
inversely correlated with levels of miR-218 in CC tissues. It was
demonstrated that miR-218 could inhibit the proliferation,
apoptosis and cell cycle progression of CC cells via suppressing
Gli3 expression. The present study revealed that miR-218 mimic
transfection in CC cells promotes apoptosis and enhances caspase-3
activity. These results suggest that miR-218 is able to inhibit
cell growth and regulate tumor progression by enhancing caspase-3
activity and promoting apoptosis in CC. Similarly, it was also
demonstrated that miR-218, which is downregulated in CC, inhibits
cell growth by blocking the transition from G1 to S phase. These
results provide a novel insight into the pathogenesis and potential
treatment targets of CC and may have clinical implications.
In summary, miR-218 overexpression in CC cells and
tissues can inhibit cancer progression by blocking transition from
G0/G1 to S phase, promoting apoptosis,
increasing the activity of proteins associated with cell apoptosis
and decreasing the expression of cell cycle-associated proteins.
The results of the present study suggest that miR-218 serves an
important role in the pathogenesis of CC and provide a theoretical
basis for the development of targeted therapy for CC.
Acknowledgements
The authors would like to thank the Department of
Gynecology, First Affiliated Hospital of Bengbu Medical
College.
Funding
Not applicable.
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ contributed to writing the manuscript, study
conception and design and the acquisition and analysis of data. SL
contributed to study conception and design as well as revising and
approving the final version of the manuscript. YL drafted the
manuscript and analyzed data. HL collected and interpreted data and
revised the final manuscript. YZ contributed to study conception,
data analysis and interpretation and drafting the manuscript. QZ
contributed to study conception and design, acquisition of data and
drafting the manuscript. All authors have read and approved the
final version of this manuscript to be published.
Ethics approval and consent to
participate
All experimental protocols were performed in
accordance with the principles of the Declaration of Helsinki and
were approved by the Clinical Research Ethics Committee of the
First Affiliated Hospital of Bengbu Medical College (Anhui, China).
Prior to enrolment, written informed consent was obtained from all
patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lahue BJ, Baginska E, Li SS and Parisi M:
Health technology assessment on cervical cancer screening
2000–2014. Int J Technol Assess Health Care. 31:171–180. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Subramanian S, Trogdon J, Ekwueme DU,
Gardner JG, Whitmire JT and Rao C: Cost of cervical cancer
treatment: Implications for providing coverage to low-income women
under the Medicaid expansion for cancer care. Womens Health Issues.
20:400–405. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Arbyn M, Castellsague X, de Sanjose S,
Bruni L, Saraiya M, Bray F and Ferlay J: Worldwide burden of
cervical cancer in 2008. Ann Oncol. 22:2675–2686. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gómez-Gómez Y, Organista-Nava J and
Gariglio P: Deregulation of the miRNAs expression in cervical
cancer: Human papillomavirus implications. Biomed Res Int 2013.
4070522013.
|
|
6
|
Fish JE, Wythe JD, Xiao T, Bruneau BG,
Stainier DY, Srivastava D and Woo S: A Slit/miR-218/Robo regulatory
loop is required during heart tube formation in zebrafish.
Development. 138:1409–1419. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Alajez NM, Lenarduzzi M, Ito E, Hui AB,
Shi W, Bruce J, Yue S, Huang SH, Xu W, Waldron J, et al: miR-218
suppresses nasopharyngeal cancer progression through downregulation
of surviving and SLIT2-ROBO1 pathway. Cancer Res. 71:2381–2391.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Small EM, Sutherland LB, Rajagopalan KN,
Wang S and Olson EN: MicroRNA-218 regulates vascular patterning by
modulation of Slit-Robo signaling. Circ Res. 107:1336–1344. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao H, Anand AR and Ganju RK: Slit2-Robo4
pathway modulates lipopolysaccharide-induced endothelial
inflammation and its expression is dysregulated during endotoxemia.
J Immunol. 192:385–393. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dickinson RE, Dallol A, Bieche I, Krex D,
Morton D, Maher ER and Latif F: Epigenetic inactivation of SLIT3
and SLIT1 genes in human cancers. Br J Cancer. 91:2071–2078. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Narayan G, Goparaju C, Arias-Pulido H,
Kaufmann AM, Schneider A, Dürst M, Mansukhani M, Pothuri B and
Murty VV: Promoter hypermethylation-mediated inactivation of
multiple Slit-Robo pathway genes in cervical cancer progression.
Mol Cancer. 5:162006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dallol A, Morton D, Maher ER and Latif F:
SLIT2 axon guidance molecule is frequently inactivated in
colorectal cancer and suppresses growth of colorectal carcinoma
cells. Cancer Res. 63:1054–1058. 2003.PubMed/NCBI
|
|
13
|
Xin SY, Feng XS, Zhou LQ, Sun JJ, Gao XL
and Yao GL: Reduced expression of circulating microRNA-218 in
gastric cancer and correlation with tumor invasion and prognosis.
World J Gastroenterol. 20:6906–6911. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ding H, Wu YL, Wang YX and Zhu FF:
Characterization of the MicroRNA expression profile of cervical
squamous cell carcinoma metastases. Asian Pac J Cancer Prev.
15:1675–1679. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zheng ZM and Wang X: Regulation of
cellular miRNA expression by human papillomaviruses. Biochim
Biophys Acta. 1809:668–677. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rao Q, Shen Q, Zhou H, Peng Y, Li J and
Lin Z: Aberrant microRNA expression in human cervical carcinomas.
Med Oncol. 29:1242–1248. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yamamoto N, Kinoshita T, Nohata N, Itesako
T, Yoshino H, Enokida H, Nakagawa M, Shozu M and Seki N: Tumor
suppressive microRNA-218 inhibits cancer cell migration and
invasion by targeting focal adhesion pathways in cervical squamous
cell carcinoma. Int J Oncol. 42:1523–1532. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee H, Kim KR, Cho NH, Hong SR, Jeong H,
Kwon SY, Park KH, An HJ, Kim TH, Kim I, et al: MicroRNA expression
profiling and Notch1 and Notch2 expression in minimal deviation
adenocarcinoma of uterine cervix. World J Surg Oncol. 12:3342014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Banno K, Iida M, Yanokura M, Kisu I, Iwata
T, Tominaga E, Tanaka K and Aoki D: MicroRNA in cervical cancer:
OncomiRs and tumor suppressor miRs in diagnosis and treatment.
ScientificWorldJournal. 2014:1780752014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Martinez I, Gardiner AS, Board KF, Monzon
FA, Edwards RP and Khan SA: Human papillomavirus type 16 reduces
the expression of microRNA-218 in cervical carcinoma cells.
Oncogene. 27:2575–2582. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Uesugi A, Kozaki K, Tsuruta T, Furuta M,
Morita K, Imoto I, Omura K and Inazawa J: The tumor suppressive
microRNA miR-218 targets the mTOR component rictor and inhibits AKT
phosphorylation in oral cancer. Cancer Res. 71:5765–5778. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
He H, Di Y, Liang M, Yang F, Yao L, Hao S,
Li J, Jiang Y, Jin C and Fu D: The microRNA-218 and ROBO-1
signaling axis correlates with the lymphatic metastasis of
pancreatic cancer. Oncol Rep. 30:651–658. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang M, Liu R, Sheng J, Liao J, Wang Y,
Pan E, Guo W, Pu Y and Yin L: Differential expression profiles of
microRNAs as potential biomarkers for the early diagnosis of
esophageal squamous cell carcinoma. Oncol Rep. 29:169–176. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tu K, Li C, Zheng X, Yang W, Yao Y and Liu
Q: Prognostic significance of miR-218 in human hepatocellular
carcinoma and its role in cell growth. Oncol Rep. 32:1571–1577.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bai XY, Lin JY, Zhang XC, Xie Z, Yan HH,
Chen ZH, Xu CR, An SJ, Sheng GM and Wu YL: High expression of
truncated GLI3 is associated with poor overall survival in patients
with non-small cell lung cancer. Cancer Biomark. 13:37–47. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-delta delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F:
Cancerincidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dappa E, Elger T, Hasenburg A, Düber C,
Battista MJ and Hötker AM: The value of advanced MRI techniques in
the assessment of cervical cancer: A review. Insights Imaging.
8:471–481. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang HY, Kim G, Cho H, Kim S, Lee D, Park
S, Park KH and Lee H: Diagnostic performance of HPV E6/E7, hTERT,
and Ki67 mRNA RT-qPCR assays on formalin-fixed paraffin-embedded
cervical tissue specimens from women with cervical cancer. Exp Mol
Pathol. 98:510–516. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Arbyn M, Castellsagué X, de Sanjosé S,
Bruni L, Saraiya M, Bray F and Ferlay J: Worldwide burden of
cervical cancer in 2008. Ann Oncol. 22:2675–2686. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Luo Q, Zhang S, Wei H, Pang X and Zhang H:
Roles of Foxp3 in the occurrence and development of cervical
cancer. Int J Clin Exp Pathol. 8:8717–8730. 2015.PubMed/NCBI
|
|
32
|
Burd EM: Human papillomavirus and cervical
cancer. Clin Microbiol Rev. 16:1–17. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nour NM: Cervical cancer: A preventable
death. Rev Obstet Gynecol. 2:240–244. 2009.PubMed/NCBI
|
|
34
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:1–866. 2006. View Article : Google Scholar
|
|
35
|
Kogo R, How C, Chaudary N, Bruce J, Shi W,
Hill RP, Zahedi P, Yip KW and Liu FF: The microRNA-218~survivin
axis regulates migration, invasion, and lymph node metastasis in
cervical cancer. Oncotarget. 6:1090–1100. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yu J, Wang Y, Dong R, Huang X, Ding S and
Qiu H: Circulating MicroRNA-218 was reduced in cervical cancer and
correlated with tumor invasion. J Cancer Res Clin Oncol.
138:671–674. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yuan W, Xiaoyun H, Haifeng Q, Jing L,
Weixu H, Ruofan D, Jinjin Y and Zongji S: MicroRNA-218 enhances the
radiosensitivity of human cervical cancer via promoting radiation
induced apoptosis. Int J Med Sci. 11:691–696. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hartono D, Lioe B, Zhang Y, Li B and Yu J:
Impacts of particulate matter (PM2.5) on the behavior of freshwater
snail Parafossarulus striatulus. Sci Rep. 7:6442017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tang BB, Liu SY, Zhang YU, Wei LQ, Mao XL,
Wang J, Li LI and Lu ZX: microRNA-218 expression and its
association with the clinicopathological characteristics of
patients with cervical cancer. Exp Ther Med. 10:269–274. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Steg A, Amm HM, Novak Z, Frost AR and
Johnson MR: Gli3 mediates cell survival and sensitivity to
cyclopamine in pancreaticcancer. Cancer Biol Ther. 10:893–902.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Iwasaki H, Nakano K, Shinkai K, Kunisawa
Y, Hirahashi M, Oda Y, Onishi H and Katano M: Hedgehog Gli3
activator signal augments tumorigenicity of colorectal cancer via
upregulation of adherence-related genes. Cancer Sci. 104:328–336.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kang HN, Oh SC, Kim JS and Yoo YA:
Abrogation of Gli3 expression suppresses the growth of colon
cancercells via activation of p53. Exp Cell Res. 318:539–549. 2012.
View Article : Google Scholar : PubMed/NCBI
|