Introduction
Astragalus membranaceus (Fisch.)
Bunge is a plant used in traditional Chinese
medicine (TCM), with known ‘Qi tonifying’ adaptogenic effects, as
documented in the Chinese Materia Medica. Astragalus
membranaceus (Fisch.) Bunge has been used in TCM for over 2,000
years and is still being use in various herbal preparations. It is
known to enhance immune functions; protect the liver; act as a
diuretics; and have anti-aging, anti-stress, antihypertensive, and
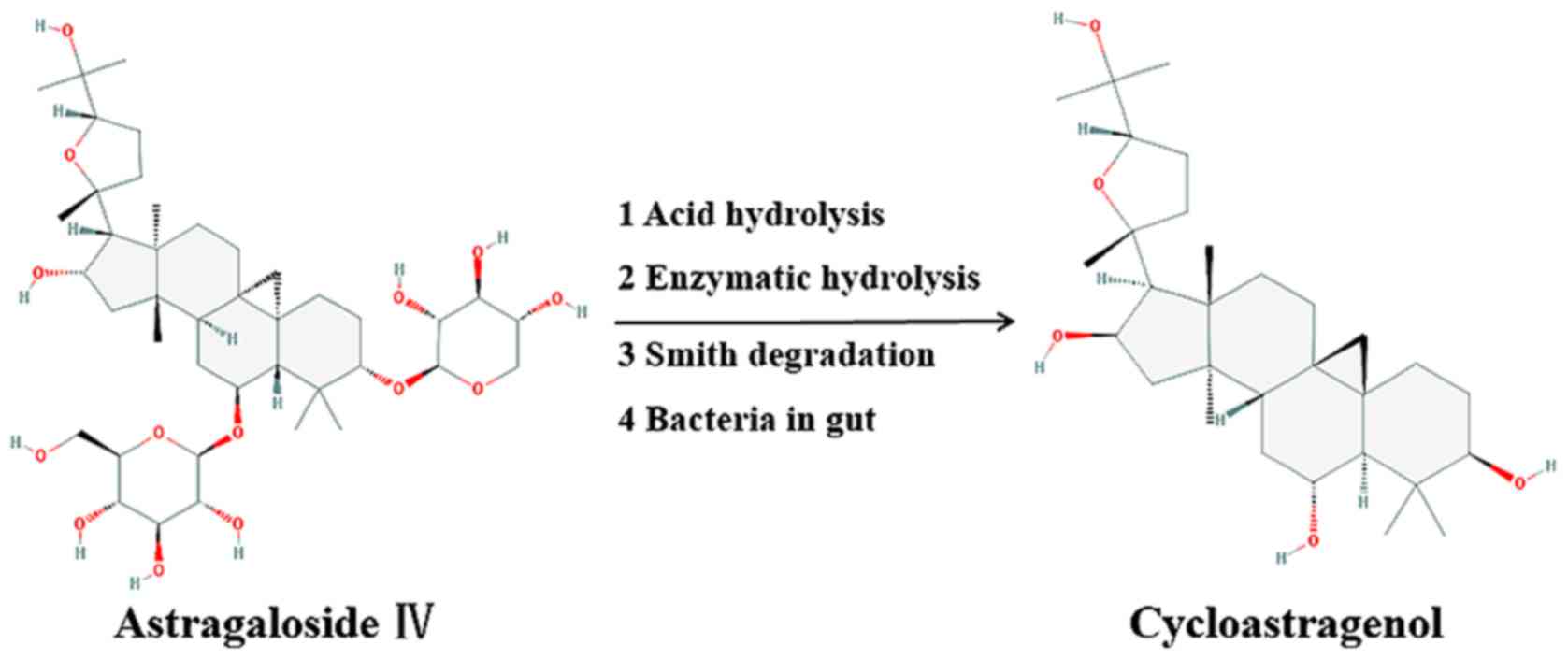
extensive antibacterial properties (1). Astragaloside IV (AG) is the main
compound found in Astragalus membranaceus (Fisch.) Bunge
while cycloastragenol (CAG), an algycone of AG, is a triterpenoid
saponin compound obtained from AG hydrolysis products (Fig. 1) (2).
The Geron Corporation in cooperation with The Hong
Kong University of Science and Technology screened natural
compounds from Astragalus membranaceus extracts. CAG was
identified as an anti-aging compound that promotes telomerase
activity and wound closure (3). CAG
is currently the only compound known to activate telomerase in
humans (4), making it a great
prospect for development. This study reviews the efficacy,
pharmacokinetics studies, and adverse reactions of CAG, and
provides a basis for further study into CAG.
Pharmacodynamics
Efficacy
Efficacy base on pre-clinical
studies
Telomerase activation
Telomeres are located at the ends of linear
chromosomes capped by nucleoprotein structures that consist of
tandem repeats of hexameric sequences (TTAGGG in vertebrates) bound
by a dedicated set of proteins. Telomeres shorten with every
mitotic event. Telomerase is a ribonucleoprotein complex that
lengthen telomeres and fundamentally consists of catalytic reverse
transcriptase enzymes (TERT) and a telomerase RNA component (TERC).
A key function of telomeres is to protect chromosomal ends from
fusion and degradation by capping chromosomal ends. Cells recognize
critically short telomeres as DNA damage; therefore, the shortest
telomere, rather than average telomere length, is critical for cell
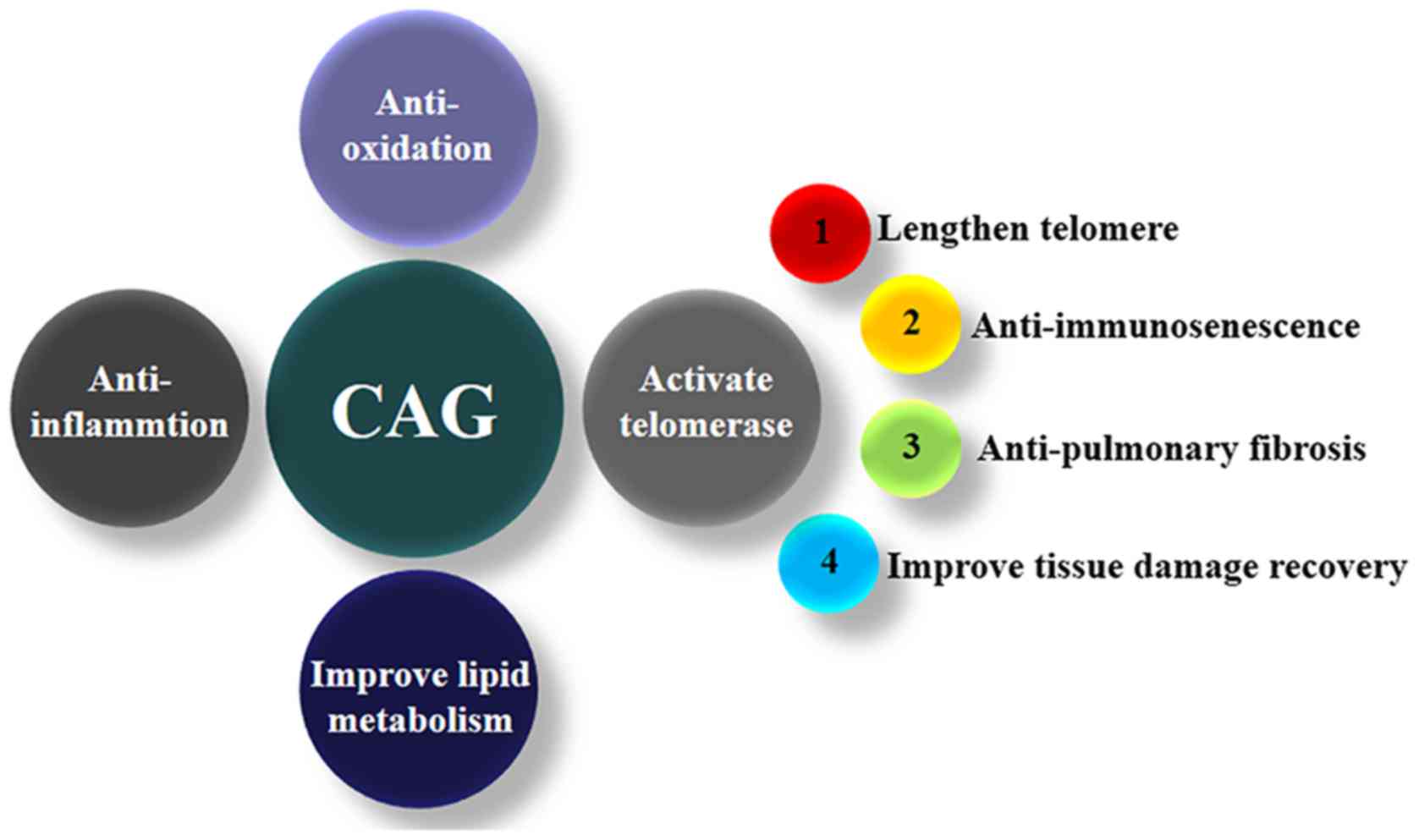
viability and chromosomal stability (5). CAG activates telomerase, lengthens
telomeres, and exerts a variety of pharmacological effects as
introduced below (Fig. 2).
Telomerase activation and lengthening
of telomeres
Several studies have shown that CAG activates
telomerase both in vitro and in vivo. CAG activates
telomerase and lengthens telomeres in a telomerase-dependent way
in vitro; therefore, CAG decreases the percentage of
critically short telomeres and DNA damage in the cell. CAG could
activate telomerase in various cell types, including hematopoietic
progenitors and mouse embryonic fibroblast haplo-insufficient for
the TERC (MEF Terc+/−) cells. In vivo,
dietary supplementation of CAG increases TERT expression in tissues
such as bone marrow, lungs, heart, brain, and liver. Further, CAG
rescued short telomeres in 1- and 2-year old female mice. However,
data have shown that dietary supplementation with CAG does not
impact the mean or maximum longevity of female mice (6,7).
Therefore, a necessary relation between telomerase activity and
longevity does not likely exist.
The above study led to another question: How does
CAG activate telomerase? Telomerase activity in human embryonic
kidney HEK293 fibroblasts increases upon treatment with CAG and is
not meditated via common secondary messenger pathways, including
Ca2+, inositol trisphosphate (IP3), cyclic adenosine
monophosphate (cAMP), and protein kinase B (Akt). However, CAG
induced the phosphorylation of extracellular signal-regulated
kinase (ERK) in many cell lines, including HEK293 and HEK-neo
keratinocytes, as well as cells from lung, brain, mammary,
endothelial, and hematopoietic origins. Further, proto-oncogene
tyrosine-protein kinase (c-Src), ERK kinase (MEK), and epidermal
growth factor receptors are involved in CAG-induced ERK
phosphorylation. CAG may activate telomerase and other cellular
effects by activating the Src/MEK/ERK pathway (8). Our previous research showed that CAG
induces expression of Janus kinase 2 (JAK2) and signal transducer
and activator of transcription 5b (STAT5b) and enhances
phosphorylation of STAT5b following an increase in TERT expression.
Further, expression of molecules downstream from the JAK/STAT
signaling pathway was also increased, suggesting that CAG activates
telomerase through the JAK/STAT signaling pathway (9). In addition, CAG-mediated activation of
telomerase is associated with the cAMP response element-binding
protein (CREB) (3). The above
preliminary studies of the mechanisms associated with CAG
demonstrate that it activates telomerase and is related to the
CREB, MAPK, and JAK/STAT pathways; however, further investigation
is needed to comprehensively understand the mechanisms.
Telomerase activation and
anti-immunosenescence
A prevailing view is that the immune system
gradually ages during the process of senescence. Weak immune
systems lead to an increased susceptibility to infections, which
accelerates the process of senescence. The immune system consists
of immune organs, immune cells, and active immune substances and is
associated with senescence (10).
Immune cells include T lymphocytes, B lymphocytes, and phagocytes,
in which the number of lymphocytes is related to immune function.
CAG enhanced both CD8(+) T lymphocytes and CD4(+) T lymphocytes,
telomerase activity, and proliferative capacity in vitro
(11,12). CD8(+) T lymphocytes from human
immunodeficiency virus (HIV)-infected human donors treated with CAG
showed an increase in telomerase activity, modest retardation of
telomere attrition, increased proliferative potential of CD8(+) T
lymphocytes, and enhanced cytokine/chemokine production, thus
enhancing the antiviral potential. CAG-induced increases in
telomerase activity are blocked by MAKP and ERK inhibitors;
therefore, it is likely that CAG-associated enhanced antiviral
functions are mediated through the ERK/MAPK pathway (13). In addition to T lymphocyte
proliferation in HIV-infected human donors, CAG enhances normal
human T lymphocyte proliferation.
An Astragalus membranaceus root extract (HTA;
not a single purified compound) has also been shown to be a
telomerase activator. However, CAG is a more effective telomerase
activator, increasing telomerase activity and the proliferative
potential of human CD4 and CD8 T cells more than that of HTA.
Further, CAG has been shown to enhance the anti-viral ability of
cells and has the potential to ameliorate acquired immunodeficiency
syndrome (AIDS); however, the anti-viral properties of CAG have not
yet been proven in animal models.
Telomerase activation and
anti-pulmonary fibrosis
Within the past decade, numerous diseases or
maladies have been associated with mutations in telomerase and/or
are known to accelerate telomere loss, including AIDS, congenital
dyskeratosis, aplastic anemia, and idiopathic pulmonary fibrosis
(IPF). CAG, a telomerase activator, has the possibility to treat
these diseases or maladies; to this end, several studies have been
conducted. CAG dose-dependently hindered bleomycin-induced fibrosis
in mTERT heterozygous mice (TRET+/−mice). However, CAG did not
alter the inflammatory response to bleomycin-induced fibrosis,
suggesting that the underlying mechanisms are not related to the
anti-inflammatory properties of CAG. Prevention against fibrosis
and suppression of senescence in cells are dependent on telomerase
activation. Further, data suggest that the mechanisms associated
with the protective effects of CAG against bleomycin-induced lung
fibrosis involve specific types of lung cells rather than all cell
types in the lung (7). The
mechanisms by which CAG works in specific cell types are likely
related to the basic cellular proliferative ability; however, this
issue has yet to be resolved.
Telomerase activation and recovery of
damaged tissue
Telomere length is proportional to cell
proliferation at the cellular level; that is, telomerase activation
and elongation of telomeres can increase the proliferative ability
of cells. The present study indicates that this hypothesis is
correct. CAG can improve skin fitness, wound healing, and fur
recovery in elderly mice and is a more remarkable agent for wound
healing in both in vitro and in vivo assays than that
of the other three main saponins from Astragalus species,
including AG, cyclocephaloside I (CCI), and cyclocanthoside E
(CCE). Of these compounds, CAG is the most effective at inducing
human keratinocyte proliferation and migration at a concentration
of 1 ng/ml. Further, a 5% preparation of CAG is the most effective
for in vivo wound healing (14). CAG stimulates telomerase activity in
human keratinocytes, which may contribute to the wound healing
effects associated with CAG.
In addition to its wound healing properties, CAG
promotes recovery from brain injuries, as shown in both cell and
animal experiments. Neural stem cells are important for recovery
from brain injuries such as hypoxic-ischemic brain injury. CAG
increases proliferation and improves survival in neural stem cells
and counters the effects of oxygen-glucose deprivation injury in
vitro (15). Further, CAG
reduces neuron apoptosis, improves nerve functional scoring, and
reduces cerebral infarction volume in a cerebral ischemia
reperfusion injury rat model in vivo. These functions are
related to increasing expression of TERT (16,17).
Furthermore, CAG promotes the renewal capacity of flight feathers
in captive zebra finches, which are nonmammalian animals (18). In addition, CAG attenuates
depression-like behavior in experimental mice. Telomere elongation
and activation of telomerase activity are proportional to the
tissue telomere-dependent recovery of cells and are potential
treatment targets and indicators of tissue recovery.
Improved lipid metabolism
Lipids store energy; however, when lipids accumulate
too much, it is harmful for health. CAG could ameliorate various
biomarkers associated with lipid metabolism. CAG has been shown to
reduce cytoplasmic lipid droplets in 3T3-L1 adipocytes at a low
dose. At high doses, CAG prevents differentiation of 3T3-L1
preadipocytes. Further, it was observed that CAG dose-dependently
stimulates calcium influx in 3T3-L1 preadipocytes. Since elevated
intracellular calcium plays a vital role in suppressing adipocyte
differentiation, CAG may mediate the balance in lipid metabolism by
activating calcium influx in 3T3-L1 preadipocytes (19).
Farnesoid X receptor (FXR) is a potential drug
target for the treatment of non-alcoholic fatty liver disease
(NAFLD), and CAG is a direct activator of FXR. Animal studies have
shown that CAG reduces high-fat diet-induced lipid accumulation in
the liver, accompanied by lowered blood glucose levels, serum
triglyceride levels, and hepatic bile acids. CAG also ameliorates
hepatic steatosis in methionine- and choline-deficient l-amino acid
diet (MCD)-induced non-alcoholic steatohepatitis (NASH) mice. These
results indicate that CAG ameliorates NAFLD via enhancement of the
FXR signaling pathway (20). In
addition, CAG increases TERT, c-Myc, and c-Jun expression in the
liver and ameliorates liver lipid accumulation as well as glucose
tolerance in 2-year old mice. CAG-ameliorated lipid metabolism may
occur via multiple mechanisms, and CAG may be a multi-target
monomer that binds with various receptors and exerts a variety of
functions.
Anti-oxidative properties
The oxidative stress hypothesis is a generally
accepted mechanism of senescence. Reactive oxygen species (ROS),
including superoxide (O2−), hydrogen peroxide
(H2O2), and the hydroxyl radical (-OH) damage cell
functions and are a primary cause of disease and cell senescence.
In a d-galactose-induced senescence mouse model, CAG treatment
up-regulated total antioxidant capacity (T-AOC) and superoxide
dismutase (T-SOD) activity, increased hydroxyproline (HYP) stores,
and down-regulated malondialdehyde (MDA) in the skin, heart, and
liver. Further, CAG enhanced the antioxidant capacity in a
non-dose-dependent manner (21). The
anti-oxidant effects of CAG may be related to the hydroxyl found in
the chemical structure of CAG. Since oxidative stress is a main
reason for telomere attrition (22),
the telomere-protective effects of CAG may be related to both its
antioxidant and telomerase activation properties.
Anti-inflammatory properties
Inflammation is a very common and important basic
pathological process. Body surface trauma, infections, common organ
diseases, and frequently-occurring diseases (such as pneumonia,
hepatitis, and nephritis) are all related to inflammation. ROS
generation was suppressed in human umbilical vein EA.hy926 cells
exposed to CAG in the setting of endoplasmic reticulum (ER) stress.
Further, CAG attenuates phosphorylation of inositol-requiring
enzyme 1 (IRE1)-α, suppresses thioredoxin-interacting protein/NLR
family pyrin domain containing 3 (TXNIP/NLRP3) inflammasome
activation, inhibits mitochondrial cell death, and reduces
apoptosis in endothelial cells. Further, CAG enhances
5′AMP-activated protein kinase (AMPK) phosphorylation, an effect
that is diminished by AMPK inhibitors, indicating a potential role
for AMPK in the anti-inflammatory properties of CAG (23).
We discussed the role of CAG in the normal immune
system above; however, CAG also plays a role in the activated
immune system. The effects of CAG in a concanavalin (Con) A
pan-activated lymphocyte model were reported by Sun et al
(24). CD69 (an early-activated
marker) and CD25 (a middle-activated marker), both of which are
expressed on the surface of lymphocytes, are inhibited in
lymphocytes pretreated with CAG and are activated via Con A
treatment in vitro. Further, pretreatment with CAG blocks
Con A-induced lymphocyte mitogenesis, induces cell-cycle arrest in
the G0/G1-phase, and downregulates cytokine expression. In
addition, Ca2+ release from intracellular ER storage and
Ca2+ influx from the extracellular environment are
inhibited in Con A-induced activated T-cells treated with CAG;
these changes may be related to the anti-inflammatory properties of
CAG (24). CAG could activate the
immune system and inhibit activated immunity similar to that of
feedback inhibition. It is possible that the structure of CAG
resembles substances found in our body, and it exerts its functions
in a similar way.
Efficacy base on clinical trial
The Patton protocol-1 provided 37 subjects with a
comprehensive dietary supplement pack containing CAG for 12 months.
This clinical trial concluded that CAG lengthens critically short
telomeres and remodels the relative proportion of circulating
leukocytes in cytomegalovirus-positive [CMV(+)] subjects toward a
more ‘youthful’ profile, as seen in CMV(−) subjects (25). Furthermore, the supplement pack
improved biomarkers of metabolic, bone, and cardiovascular health,
such as fasting glucose, insulin, cholesterol, blood pressure, and
bone mineral density (BMD). These effects are mostly attributable
to CAG (26). A study reported by
Harley et al (26), reported
an increase in BMD in naturally aging rats; clinical trials have
further verified this function. It is noteworthy that the Patton
protocol-1 did not establish a control before initiating the study,
which may have affected the trial results.
A random clinical trial conducted in 117 relatively
healthy CMV (+) subjects 53–87 years old verified that TA-65 can
significantly lengthen telomeres (4). Other TCM extracts, such as
Cynomorium songaricum polysaccharides and Epimedium
brevicornu flavonoids, can also activate telomerase or protect
telomeres; however, CAG is the only compound that has been shown to
activate telomerase in humans to date. Further, a randomized
placebo-controlled study showed that CAG significantly improves the
macular function of treatment subjects (27).
CAG activates human telomerase and has various
positive functions. It is still unknown whether these different
functions are related to each other and to CAG dosages and whether
CAG functions differently in different groups of people.
Adverse reactions
Critically short telomeres will lead to cell
senescence and apoptosis, and numerous diseases or maladies are
related to short telomeres. Telomerase activators could attenuate
this process. However, telomeres are elongated via the telomerase
enzyme in more than 80% of tumors. As such, activated telomerase is
a hallmark of cancer (28). As CAG
is a telomerase activator, it is vital to understand the adverse
reactions associated with CAG treatment regimens. No
treatment-related mortalities were observed in rats ingesting 0,
40, 80, or 150 mg/kg/day CAG via oral gavage for 91 consecutive
days in a subchronic study with a 4-week recovery period; further,
no adverse effects were observed. In male and female rats, the oral
no-observed-adverse-effect level (NOAEL) of CAG was >150
mg/kg/day. In addition, CAG did not induce any toxic or genotoxic
effect according to the result of bacterial reverse mutation assay,
in vitro chromosome aberration assay and in vivo
erythrocyte micronucleus assay (29). Administration of CAG for 4 months in
adult female mice (25 mg/kg/day) did not increase the incidence of
cancer. However, CAG competitively inhibited
UDP-glucuronosyltransferase (UGT) 1A8 and noncompetitively
inhibited UGT2B7. Herb-drug interactions between CAG and drugs that
are mainly metabolized by UGT1A8 or UGT2B7 might occur in
vivo when the plasma concentration of CAG is above 0.034 and
20.98 µM, respectively (30).
Furthermore, the Patton protocol-1, a commercial health maintenance
program that provided 7000 person-years of use with a comprehensive
dietary supplement pack containing CAG over 5 years, reported no
adverse events or effects attributed to administration of the
dietary supplement pack (26).
According to current research results, CAG is
relatively safe within a certain dose range and has no serious
adverse reactions; CAG was determined to be generally recognized as
safe (GRAS) by an independent expert panel of the Food and Drug
Administration (FDA) on the 19th of November, 2014. CAG has been
used as a medical food (31).
However, telomerase activation has been observed in 80% of human
tumors, suggesting that telomerase plays a nonnegligible role in
tumor development (32). Thus, the
potential adverse reactions of CAG should be noted. More studies
regarding adverse reactions of CAG are necessary to fully
understand how to use CAG rationally.
Pharmacokinetics
CAG is efficiently absorbed through the intestinal
epithelium by passive diffusion, as shown by a study that
investigated the intestinal absorption and metabolism of CAG using
an in vitro models composed of human small intestinal mucosa
(Caco-2 model) and liver microsomes. However, metabolites were
found in the apical and basolateral sides of Caco-2 monolayers,
suggesting that first-pass intestinal metabolism of CAG might occur
upon passage through the intestinal epithelium. After incubation
for 30 min, CAG underwent extensive metabolism in rat (17.4%) and
human (8.2%) liver microsomes (33).
An comparison of CAG metabolizing ability of different rat tissues
showed that the metabolic capacity of heart, spleen, lung, and
kidney was lower than that of the liver in vivo (34). Further, a significant inhibitory
effect on the cytochrome P450 3A4 (CYP3A4) subunit, as
well as significant induction of the CYP2E1 subunit were observed
after administration of CAG via continuous gavage (35). These results indicate that although
CAG is efficiently absorbed through the intestinal epithelium, its
oral bioavailability CAG would be limited by extensive first-pass
hepatic metabolism.
Discussion
CAG naming
In the process of searching documents, we found that
CAG (CAS number: 78574-94-4) is noted by diverse names in different
documents. CAG is named
(2aR,3R,4S,5aS,5bS,7S,7aR,9S,11aR,12aS)-3-((2R,5S)-5-
(2-hydroxypropan-2-yl)-2-methyltetrahydrofuran-2-yl)-2a,5a,8,8-tetramethylhexadecahydrocyclopenta[a]cyclopropa[e]phenanthrene-4,7,9-triol,
other synonyms include CAG, CA, astramembrangenin,
cyclogalegigenin, GRN510, and TA-65. TA-65 was named by the T.A
Science Corporation as a nutrition and health care product. A
document reported that GRN510 is a novel and proprietary chemical
entity derived from GRN665/TAT2. However, GRN510 is considered
equivalent to CAG in PubChem (an open chemistry database);
therefore, we considered GRN510 as equivalent to CAG in this
article. For simplicity, we termed cycloastragenol as CAG in this
report.
Advantages and disadvantages
associated with CAG
CAG has multiple pharmacological effects, including
activation of telomerase, improved lipid metabolism,
anti-oxidation, and anti-inflammatory properties. Clinical trials
have proven components involved in CAG functions. No serious
adverse effects are associated with CAG within a certain dose
range. However, the carcinogenic potential of CAG is the main
factor preventing the use of CAG clinically. Clarification of the
quantitative effects and efficacy associated with CAG in different
individuals is needed.
Unknown information regarding CAG
While there is a large body of literature regarding
CAG, there are still significant gaps in knowledge.
CAG stimulates calcium influx or inhibits calcium
influx depending on the cell type and pathological state. Further,
CAG has different pharmacological effects at diverse concentrations
in 3T3-L1 preadipocytes. The underlying mechanisms regarding these
effects have not yet been reported.
CAG has extensive pharmacological effects; however,
the detailed underlying mechanisms for most of these effects remain
unclear. A summary of CAG-associated effects and the corresponding
mechanisms are show in Table I.
Mechanisms underlying the extensive pharmacological effects
associated with CAG are complex. We attempted to establish a
hypothetical mechanism for the effects of CAG based on previous
results (Fig. 3).
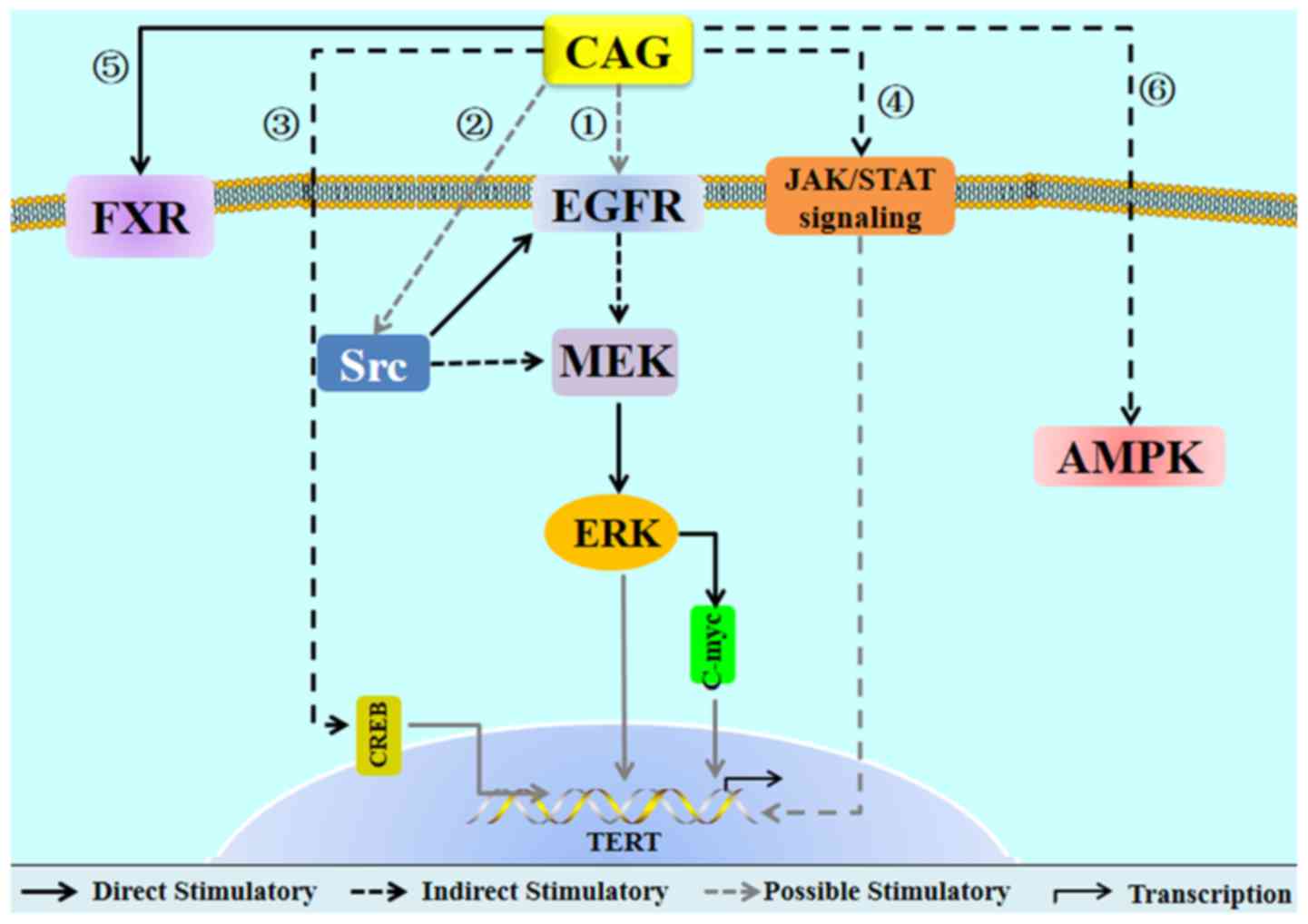 | Figure 3.Hypothetical mechanisms associated
with CAG. Arrow heads represent stimulatory modifications. Solid
lines are direct stimulatory modifications based on the literature.
Dashed lines indicate stimulatory modifications; grey dashed lines
indicate possible stimulatory modifications. CAG may activate
telomerase through pathway 1, 2, 3 and 4, and then exert various
effects. Through pathway 5, CAG directly stimulates the FXR to
improve hepatitis. Through pathway 6, CAG indirectly stimulates
AMPK to improve inflammation. CAG, cycloastragenol; FXR, farnesoid
X receptor; AMPK, 5′adenosine monophosphate-activated protein
kinase; EGFR, epidermal growth factor receptor; ERK, extracellular
signal-regulated kinase; MEK, mitogen-activated protein kinase
kinase; JAK, Janus kinase; STAT, signal transducer and activator of
transcription; TERT, telomerase reverse transcriptase. |
 | Table I.CAG pharmacological effects and
mechanisms. |
Table I.
CAG pharmacological effects and
mechanisms.
| Author, year | Object of
study | Effect of CAG | Mechanism | Tool or method of
studying mechanism | (Refs.) |
|---|
| Fauce et al,
2008 | CD8(+) T
lymphocytes from HIV-infected human donors | Enhanced antiviral
functions | Increase telomerase
activity | Telomerase template
antagonist-GRN163L | (13) |
| Fauce et al,
2008 | CD8(+) T
lymphocytes from HIV-infected human donors | Increase telomerase
activity | ERK/MAPK
pathway | MAKP inhibitor, ERK
inhibitor | (13) |
| Molgora et
al, 2013 | Healthy human CD4
and CD8 T cells | Increase telomerase
activity | MAPK pathway | MAKP
inhibitor(PD98059) | (11) |
| Zhao et al,
2015 | Endothelial
cell | Ameliorated
endothelial inflammation and reduced cell apoptosis | AMPK pathway | AMPK inhibitor | (23) |
| Sun et al,
2017 | Activated
lymphocytes |
Anti-inflammation | Inhibited
Ca2+ overload | Flow cytometry | (24) |
| Bernardes de Jesus
et al, 2011 | MEF Terc+/− | Lengthened
telomeres, reduced critically short telomeres and DNA damage | Increase telomerase
activity | Gene knockout | (6) |
| Yung et al,
2012 | Human embryonic
kidney HEK293 fibroblasts | Increased
telomerase activity | Src/MEK/ERK
pathway | Selective
inhibitors and dominant negative mutants | (8) |
| Le Saux et
al, 2013 | TRET+/− mice | Inhibited fibrosis
and prevented senescent cell accumulation | Increased
telomerase activity | Telomerase
inhibitor-GRN163L | (7) |
| Wang et al,
2014 | 3T3-L1
preadipocytes | Reduced cytoplasmic
lipid droplets | Stimulated calcium
influx | Calcium
mobilization assay | (19) |
| Gu et al,
2017 | High-fat diet
mice | Improved hepatic
steatosis | Activated farnesoid
X receptor signaling | PCR, WB, Molecular
docking, gene knockout | (20) |
| Bernardes de Jesus
et al, 2011 | 2-year old
mice | Improved hepatic
lipid accumulation | Increased
telomerase activity via c-Myc and c-Jun | PCR, IHC | (6) |
| Ip et al,
2014 | Human neonatal
keratinocytes | Improved wound
healing | Increased
telomerase activity | RQ-TRAP assay | (3) |
| Meng et al,
2017; Yi et al, 2015; Gao, 2015 | Hypoxic-ischemic
brain injury | Countered
hypoxic-ischemic brain injury | Increased
telomerase activity | Telomerase
inhibitor | (15–17) |
| Reichert et
al, 2014 | Zebra finches | Improved flight
feather renewal capacity | Increased telomere
length | qPCR | (18) |
| Cao et al,
2012 | D-galactose-induced
senescent mouse model | Enhanced
antioxidant capacity | CAG
anti-oxidant | Corresponding
detection method | (21) |
| Salvador et
al, 2016 | Relatively healthy
cytomegalovirus-positive subjects | Lengthened
telomeres | – | Oral CAG | (4) |
| Dow and Harley,
2016 | Patients | Improved macular
function | – | Oral CAG | (26) |
| Ip et al,
2014 | PC12 cells and
primary neurons and Bcl2 expression | Induced telomerase
activity and CREB activation followed by tert | CAG function
related to CREB activation | Blockade of CREB
expression via RNA | (3) |
CAG could increase BMD and improve glucose
metabolism in naturally aged rats. There is no literature
describing whether CAG has anti-osteoporosis or anti-diabetic
effects in corresponding animal models.
Conclusions and prospective
In this review, the current state of CAG research is
detailed and elucidated, and the efficacy, pharmacokinetics, and
adverse reactions of CAG are summarized. According to the present
research results, CAG has extensive pharmacological effects,
including telomerase activation, telomere elongation,
anti-inflammation, anti-oxidation, anti-viral, anti-pulmonary
fibrosis, anti-ischemic and hypoxic injury, and anti-lipid
accumulation properties. Studies also suggest that CAG improves
liver metabolic homeostasis, promotes wound healing, promotes
feather growth, and improves certain health-span indicators both in
humans and animals. However, more attention and further studies are
needed to evaluate whether CAG has potential adverse reactions, and
studies examining the detailed underlying mechanisms associated
with CAG are required. In addition, further multicenter clinical
studies are required for specific diseases such as steatohepatitis,
AIDS, pulmonary fibroses, wound healing etc.
Prospective
We anticipate that researchers will conduct more
studies on CAG to evaluate its efficacy and adverse reactions, as
well as understand how to use CAG rationally to enhance its effects
and reduce adverse reactions so that CAG can be used for the
clinical treatment of corresponding diseases. We believe that
development of these clinical uses will benefit people, which is
the purpose of this article.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from
National Natural Science Foundation of China (grant nos. 81102450
and 81673814), the Open Fund Project of Key Laboratory of Guangdong
Province (grant no. 4CX16010G), the Science and Technology Plan of
Guangdong Province (grant nos. 2016ZC0178 and 2016A020215148) and
Open Foundation of Guangdong Key Laboratory for Research and
Development of Natural Drugs (grant no. TRYW201603).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LZ performed the literature review and mapping. YL
and YYg made suggestions and edited the manuscript. YYu wrote the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CAG
|
cycloastragenol
|
|
TCM
|
traditional Chinese medicine
|
|
TERT
|
catalytic reverse transcriptase
enzymes
|
|
TERC
|
telomerase RNA component
|
|
IPE
|
inositol trisphosphate
|
|
cAMP
|
cyclic adenosine monophosphate
|
|
Akt
|
protein kinase B
|
|
c-Src
|
proto-oncogene tyrosine-protein
kinase
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
MEK
|
ERK kinase
|
|
JAK2
|
Janus kinase 2
|
|
STAT5b
|
signal transducer and activator of
transcription 5b
|
|
HIV
|
human immunodeficiency virus
|
|
IPF
|
idiopathic pulmonary fibrosis
|
|
AIDS
|
acquired immunodeficiency syndrome
|
|
AG
|
astragaloside IV
|
|
CCI
|
cyclocephaloside I
|
|
CCE
|
cyclocanthoside E
|
|
FXR
|
Farnesoid X receptor
|
|
NAFLD
|
non-alcoholic fatty liver disease
|
|
MCD
|
methionine- and choline-deficient
L-amino acid diet
|
|
NASH
|
non-alcoholic steatohepatitis
|
|
ROS
|
reactive oxygen species
|
|
T-AOC
|
total antioxidant capacity
|
|
T-SOD
|
total superoxide dismutase
|
|
HYP
|
hydroxyproline
|
|
MDA
|
malondialdehyde
|
|
ER
|
endoplasmic reticulum
|
|
IRE1
|
inositol-requiring enzyme 1
|
|
TXNIP
|
thioredoxin-interacting protein
|
|
NLRP3
|
NLR family pyrin domain containing
3
|
|
AMPK
|
5′AMP-activated protein kinase
|
|
BMD
|
bone mineral density
|
|
NOAEL
|
no-observed-adverse-effect level
|
|
UGT
|
UDP-glucuronosyltransferase
|
|
GRAS
|
generally recognized as safe
|
References
|
1
|
Shen CY, Jiang JG, Yang L, Wang DW and Zhu
W: Anti-ageing active ingredients from herbs and nutraceuticals
used in traditional Chinese medicine: Pharmacological mechanisms
and implications for drug discovery. Br J Pharmacol. 174:1395–1425.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang S, Lu J, Yan J and Zhang H: Research
progress of preparation technology and pharmacological effect of
cycloastragenol. Chin J New Drugs. 16:1872–1875. 2016.
|
|
3
|
Ip FC, Ng YP, An HJ, Dai Y, Pang HH, Hu
YQ, Chin AC, Harley CB, Wong YH and Ip NY: Cycloastragenol is a
potent telomerase activator in neuronal cells: Implications for
depression management. Neurosignals. 22:52–63. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Salvador L, Singaravelu G, Harley CB, Flom
P, Suram A and Raffaele JM: A natural product telomerase activator
lengthens telomeres in humans: A randomized, double blind and
placebo controlled study. Rejuvenation Res. 19:478–484. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hemann MT, Strong MA, Hao LY and Greider
CW: The shortest telomere, not average telomere length, is critical
for cell viability and chromosome stability. Cell. 107:67–77. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bernardes de Jesus B, Schneeberger K, Vera
E, Tejera A, Harley CB and Blasco MA: The telomerase activator
TA-65 elongates short telomeres and increases health span of
adult/old mice without increasing cancer incidence. Aging Cell.
10:604–621. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Le Saux CJ, Davy P, Brampton C, Ahuja SS,
Fauce S, Shivshankar P, Nguyen H, Ramaseshan M, Tressler R, Pirot
Z, et al: A novel telomerase activator suppresses lung damage in a
murine model of idiopathic pulmonary fibrosis. PLoS One.
8:e584232013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yung LY, Lam WS, Ho MK, Hu Y, Ip FC, Pang
H, Chin AC, Harley CB, Ip NY and Wong YH: Astragaloside IV and
cycloastragenol stimulate the phosphorylation of extracellular
signal-regulated protein kinase in multiple cell types. Planta Med.
78:115–121. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li J: Astragaloside IV and cycloastragenol
attenuate osteoblastic injury and aging elicited by oxidative
stress. 2017.
|
|
10
|
Bektas A, Schurman SH, Sen R and Ferrucci
L: Human T cell immunosenescence and inflammation in aging. J
Leukoc Biol. 102:977–988. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Molgora B, Bateman R, Sweeney G, Finger D,
Dimler T, Effros RB and Valenzuela HF: Functional assessment of
pharmacological telomerase activators in human T cells. Cells.
2:57–66. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Verotta L, Guerrini M, El-Sebakhy NA,
Assad AM, Toaima SM, Radwan MM, Luo YD and Pezzuto JM: Cycloartane
and oleanane saponins from egyptian Astragalus spp. as modulators
of lymphocyte proliferation. Planta Med. 68:986–994. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fauce SR, Jamieson BD, Chin AC, Mitsuyasu
RT, Parish ST, Ng HL, Kitchen CM, Yang OO, Harley CB and Effros RB:
Telomerase-based pharmacologic enhancement of antiviral function of
human CD8+ T lymphocytes. J Immunol. 181:7400–7406. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sevimli-Gur C, Onbasilar I, Atilla P, Genç
R, Cakar N, Deliloğlu-Gürhan I and Bedir E: In vitro growth
stimulatory and in vivo wound healing studies on cycloartane-type
saponins of Astragalus genus. J Ethnopharmacol. 134:844–850. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Meng JJ, Li SP, Zhao FY, Tong Y, Mu DZ and
Qu Y: Effect of telomerase activation on biological behavior of
neural stem cells in hypoxic-ischemic rats. Zhongguo Dang Dai Er Ke
Za Zhi. 19:229–36. 2017.(In Chinese). PubMed/NCBI
|
|
16
|
Yi Z and Gao W: Protective effects of
cycloastragenol on focal cerebral ischemia reperfusion injury in
rats. Journa Cheng Medical Collage. 1–188. 2015.
|
|
17
|
Gao W: Pfect of cycloastragenol on
neuronal apoptosis and TERT expression after cerebral ischemia
reperfusion in rats. Chin J Pathophysiology. 10:18582015.
|
|
18
|
Reichert S, Bize P, Arrive M, Zahn S,
Massemin S and Criscuolo F: Experimental increase in telomere
length leads to faster feather regeneration. Exp Gerontol.
52:36–38. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang S, Zhai C, Liu Q, Wang X, Ren Z,
Zhang Y, Zhang Y, Wu Q, Sun S, Li S and Qiao Y: Cycloastragenol, a
triterpene aglycone derived from Radix astragali, suppresses the
accumulation of cytoplasmic lipid droplet in 3T3-L1 adipocytes.
Biochem Biophysical Res Commun. 450:306–311. 2014. View Article : Google Scholar
|
|
20
|
Gu M, Zhang S, Zhao Y, Huang J, Wang Y, Li
Y, Fan S, Yang L, Ji G, Tong Q and Huang C: Cycloastragenol
improves hepatic steatosis by activating farnesoid X receptor
signalling. Pharmacol Res. 121:22–32. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cao Y, Li W, Wei L, Liu X, Li Z and He W:
Anti-aging effect of cyclogreitol on D-galactose induced aging
mice. Chin J Exp Tradit Med Formulae. 1–211. 2012.
|
|
22
|
Richter T and von Zglinicki T: A
continuous correlation between oxidative stress and telomere
shortening in fibroblasts. Exp Gerontol. 42:1039–1042. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao Y, Li Q, Zhao W, Li J, Sun Y, Liu K,
Liu B and Zhang N: Astragaloside IV and cycloastragenol are equally
effective in inhibition of endoplasmic reticulum stress-associated
TXNIP/NLRP3 inflammasome activation in the endothelium. J
Ethnopharmacol. 169:210–218. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun C, Jiang M, Zhang L, Yang J, Zhang G,
Du B, Ren Y, Li X and Yao J: Cycloastragenol mediates activation
and proliferation suppression in concanavalin A-induced mouse
lymphocyte pan-activation model. Immunopharmacol Immunotoxicol.
39:131–139. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harley CB, Liu W, Blasco M, Vera E,
Andrews WH, Briggs LA and Raffaele JM: A natural product telomerase
activator as part of a health maintenance program. Rejuvenation
Res. 14:45–56. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Harley CB, Liu W, Flom PL and Raffaele JM:
A natural product telomerase activator as part of a health
maintenance program: metabolic and cardiovascular response.
Rejuvenation Res. 16:386–395. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dow CT and Harley CB: Evaluation of an
oral telomerase activator for early age-related macular
degeneration-a pilot study. Clin Ophthalmol. 10:243–249. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Aschacher T, Wolf B, Enzmann F, Kienzl P,
Messner B, Sampl S, Svoboda M, Mechtcheriakova D, Holzmann K and
Bergmann M: LINE-1 induces hTERT and ensures telomere maintenance
in tumour cell lines. Oncogene. 35:94–104. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Szabo NJ: Dietary safety of
cycloastragenol from Astragalus spp: Subchronic toxicity and
genotoxicity studies. Food Chem Toxicol. 64:322–334. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ran R, Zhang C, Li R, Chen B, Zhang W,
Zhao Z, Fu Z, Du Z, Du X, Yang X and Fang Z: Evaluation and
comparison of the inhibition effect of astragaloside IV and
aglycone cycloastragenol on various UDP-Glucuronosyltransferase
(UGT) isoforms. Molecules. 21:E16162016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
T.A.: Sciences®:
TA-65® Receives Generally Recognized as Safe Status.
https://www.tasciences.com/ta-65-receives-generally-recognized-safe-status/November
19–2014
|
|
32
|
Shay JW and Bacchetti S: A survey of
telomerase activity in human cancer. Eur J Cancer. 33:787–791.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhu J, Lee S, Ho MK, Hu Y, Pang H, Ip FC,
Chin AC, Harley CB, Ip NY and Wong YH: In vitro intestinal
absorption and first-pass intestinal and hepatic metabolism of
cycloastragenol, a potent small molecule telomerase activator. Drug
Metab Pharmacokinet. 25:477–486. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu X: In vitro and in vivo metabolism
research of astragaloside iv and cycloastragenol (unpublished PhD
thesis). Beijing University of Chinese Medicine. 2013, PubMed/NCBI
|
|
35
|
Wei B, Ye J, Bi B, Sun G, Yang X and Zhang
Y: Effect of continuous intragastric administration of
cycloastragenol on liver p450 enzymes in rats. Chin J New Drugs.
1–479. 2014.
|