Introduction
The incidence of diabetes mellitus (DM), a complex
metabolic disorder associated with defective insulin secretion and
activity, has been increasing worldwide over the past 20 years
(1,2). According to the World Health
Organization estimation, ~7% of the world's adult population is
diabetic and the diabetic population is likely to increase to ≥300
million by the year 2025 (3). Type 1
DM (T1DM) is an organ-specific autoimmune disease associated with
failure to distinguish self- from non-self-antigens (4). It is caused by T cell-mediated
destruction of insulin-producing pancreatic β-cells (4). The incidence of T1DM is increasing
steadily by 3% annually and has a concordance rate of 40–60% for
monozygotic twins (4). As such,
environmental and genetic factors may contribute to disease onset
(5–7). Currently, insulin therapy is the
primary treatment for T1DM. However, tight glycemic control is
difficult to achieve in a number of patients, leading to long-term
vascular damage associated with kidney failure, heart disease,
retinopathy and neuropathy (8).
Recent advances in pancreatic islet transplantation and partial or
whole pancreas transplantation represent alternate treatment
options for T1DM (9). However, due
to the limited number of organs available for transplant, this
approach is not widely used (9).
Since β-cell damage is crucial to the development of T1DM,
treatments that are able to prevent β-cell damage may slow disease
progression.
Pancreatic β-cell damage is known to be mediated by
the immune response (10–12). Previous studies have suggested that
inflammatory cytokines and immune cell infiltration activate
oxidative and endoplasmic reticulum (ER) stress and damage β-cell
viability (10–16). In the early stages of disease, the
infiltration of inflammatory cells promotes the release of
cytokines, including interleukin-1β (IL-1β), tumor necrosis
factor-α (TNF-α) and interferon-γ (IFN-γ) (15–16).
IL-1β, alone or in combination with TNF-α or IFN-γ, upregulates the
expression of inducible nitric oxide synthase (iNOS) and promotes
the generation of nitric oxide (NO) in pancreatic islets (17,18).
Excessive NO production leads to dysfunctions of mitochondrial
metabolism, protein modification and DNA cleavage, which may
contribute to the impairment of insulin secretion and triggering
β-cell death (19).
Considering the inflammatory nature of T1DM, it is
plausible that anti-inflammatory agents may have potential as
anti-DM drugs. Icariin is a naturally occurring flavonoid isolated
from traditional Chinese medicinal herbs of the Epimedium
genus (20). The compound has been
revealed to have anti-inflammatory, antidepressant, male
reproductive, antineoplastic, bone-healing and neuroprotective
effects (20). Early in vivo
and in vitro studies revealed that icariin acts as a natural
anti-inflammatory drug via multiple mechanisms targeting
pro-inflammatory cytokines (TNF-α and IL-6), inflammatory mediators
(NO) and adhesion molecules (CD11b) (21,22). Xu
et al (23) reported that
icariin activates the phosphoinositide 3-kinase (PI3K)/protein
kinase B (Akt) signaling pathway to ameliorate lipopolysaccharide
(LPS)-induced acute inflammatory responses. The known
anti-inflammatory effects of icariin suggest that it may inhibit
inflammation-induced β-cell death. The aim of the present study was
to use rat pancreatic β-cell lines as an in vitro model to
investigate the role of icariin. The results suggest that icariin
inhibits cytokine-induced NF-κB activation and prevents β-cell
death.
Materials and methods
Cell culture
Rat pancreatic β-cell RINm5F cells were obtained
from ATCC (Manassas, VA, USA). Cells were cultured in RPMI-1640
medium (Hyclone; GE Healthcare Life Sciences, Logan, UT, USA) with
10% (v/v) heat-inactivated fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), 2 mM glutamine
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), 1% non-essential
amino acids (Sigma-Aldrich; Merck KGaA), 100 U/ml streptomycin and
100 U/ml penicillin (Sigma-Aldrich; Merck KGaA) at 37°C in an
atmosphere containing 5% CO2. Icariin was purchased from
Sigma-Aldrich (Merck KGaA). Rat IL-1β and IFN-γ proteins were
obtained from R&D Systems (Minneapolis, MN, USA).
MTT assay
MTT (Sigma-Aldrich; Merck KGaA) was used to
determine cell viability according to the manufacturer's protocols.
Briefly, 5 ml MTT solvent (Beyotime Institute of Biotechnology,
Haimen, China) was used to dissolve 25 mg MTT to form an MTT
solution at 5 mg/ml. A total of 10 µ1 MTT solution was added to
each well and incubated for 4 h at 37°C in an incubator.
Subsequently, 100 µl formazan solution (Beyotime Institute of
Biotechnology) was added for 4 h at 37°C. The optical density of
viable cells was measured using a microplate reader (BMG Labtech
GmbH, Ortenburg, Germany) at a wavelength of 570 nm.
NO measurement
Biologically synthesized NO is quickly oxidized to
form nitrite and nitrate in aqueous solutions (19). Therefore, detecting the nitrite
concentration in cell-free culture supernatants using a
colorimetric assay may be indicative of NO generation. In brief,
RINm5F cells (5×106) or 30 islets were treated with the
5 or 10 µM concentrations of icariin for 3 h, prior to being
treated with IL-1β (1 U/ml) and IFN-γ (100 U/ml) for 24 h.
Subsequently, 100 µl aliquots of culture supernatant were incubated
at room temperature for 5 min with 100 µl modified Griess reagent
in a 1:1 mixture of 1% sulfanilamide in 30% acetic acid and 0.1%
N-(1-naphthyl) ethylenediamine dihydrochloride in 60% acetic acid
(Beyotime Institute of Biotechnology). The absorbance was measured
at 540 nm. The NO concentration was calculated from the linear
standard curve of serial dilutions of sodium nitrite in a working
medium.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cultured cells using
TRIzol reagent (Thermo Fisher Scientific, Inc.). The primer for
iNOS was synthesized based on the following previously published
sequences (24): Forward,
5′-GAATCTTGGAGCGAGTTGTGG-3′ and reverse,
5′-AGGAAGTAGGTGAGGGCTTGG-3′. First-strand cDNA was obtained using
Super M-MLV Reverse transcriptase (BioTeke Corporation, Beijing,
China). Reverse transcription was performed at 42°C for 15 min and
72°C for 2 min according to the manufacturer's protocols. PCR was
performed using SYBR-Green master mix (Beijing Solarbio Science
& Technology Co., Ltd., Beijing, China). The following
thermocycling conditions were used: Predenaturation at 95°C for 30
sec followed by 40 cycles of amplification at 95°C for 5 sec and
annealing and extension at 60°C for 30 sec. GADPH was used to
normalize iNOS mRNA expression. GAPDH forward,
5′-GATGACCTTGCCCACAGCCT-3′ and reverse, 5′-ATCTCTGCCCCCTCTGCTGA-3′.
The 2−∆∆Cq method was used to quantify data (24). ABI Prism 7000 software (Applied
Biosystems; Thermo Fisher Scientific, Inc.) was used to analyze
data.
Western blotting
Following treatment, proteins were extracted from
RINm5F cells using a Nuclear and Cytoplasmic Protein Extraction kit
(cat. no. P0027; Beyotime Institute of Biotechnology). Protein
concentrations were determined using an Enhanced BCA Protein Assay
kit (cat. no. P0010S; Beyotime Institute of Biotechnology). A total
of 20 µg/lane was separated by 12% SDS-PAGE and transferred to
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA). The membranes were blocked using Blocking Buffer (cat. no.
P0023B; Beyotime Institute of Biotechnology) for 2 h at room
temperature. Proteins were probed using specific primary antibodies
at 4°C overnight, followed by incubation with secondary antibodies
at room temperature for 1 h. Specific primary antibodies against
pro-caspase-3 (ab44976; 1:500), cleaved caspase-3 (ab13847; 1:500)
and cleaved poly ADP-ribose polymerase (PARP; ab32064; 1:2,000)
were purchased from Abcam (Cambridge, UK). Secondary antibodies
against β-actin (ab8227; 1:2,000) and Larmin A (ab26300; 1:1,000)
used in this study were horseradish peroxidase (HRP) conjugated
goat anti-rabbit IgG or anti-mouse IgG-HRP (Beyotime Institute of
Biotechnology). β-actin and Larmin A were used as internal controls
to normalize results. Signals were monitored using a
chemiluminescent substrate (KPL, Inc., Gaithersburg, MD, USA).
Following electrophoresis, gray values were analyzed using Quantity
One v4.4.0.36 software (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Caspase-3 activity
The activity of caspase-3 was conducted using a
commercial ELISA kit (cat. no. HC079; Shanghai Gefan Biotechnology
Co., Ltd., Shanghai, China) according to the manufacturer's
protocols. In brief, cells (1×106) were resuspended in
50 µl lysis buffer (Shanghai Gefan Biotechnology Co., Ltd.) and
incubated for 1 h in an ice bath. The supernatant was collected
following centrifugation for 10 min at 800 × g at room temperature,
following which a colorimetric reagent was added and incubated for
4 h at 37°C. The colorimetric product was monitored using an ELISA
reader at a wavelength of 405 nm.
Apoptosis detection using flow
cytometry
A total of 1×106 cells were washed with
PBS and resuspended in binding buffer containing Annexin V-APC and
propidium iodide, and incubated at 20–25°C for 10–20 min (Beyotime
Institute of Biotechnology). The samples were analyzed using a
FACScan flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
The percentage of apoptotic cells in a 10,000-cell cohort was
determined using flow cytometry.
NF-κB P65 activity
Following treatment, nuclear extracts were isolated
using the Nuclear Extract kit according to the manufacturer's
protocols (Active Motif, Carlsbad, CA, USA; cat. no. 40010). The
activity of NF-κB p65 was assessed using an ELISA kit (cat. no.
40596; Active Motif).
Statistical analysis
Values are presented as the mean ± standard
deviation. Statistical comparisons between cell lines were
performed using one-way analysis of variance, followed by Dunnett's
t-test. GraphPad Prism 7.03 software (GraphPad Software Inc., La
Jolla, CA, USA) was used to analyze experimental data and a
P<0.05 was considered to indicate a statistically significant
difference.
Results
Icariin prevents cytokine-induced loss
of cell viability
To assess the therapeutic potential of icariin in
rat pancreatic β cells, the viability of cultured RINm5F cells was
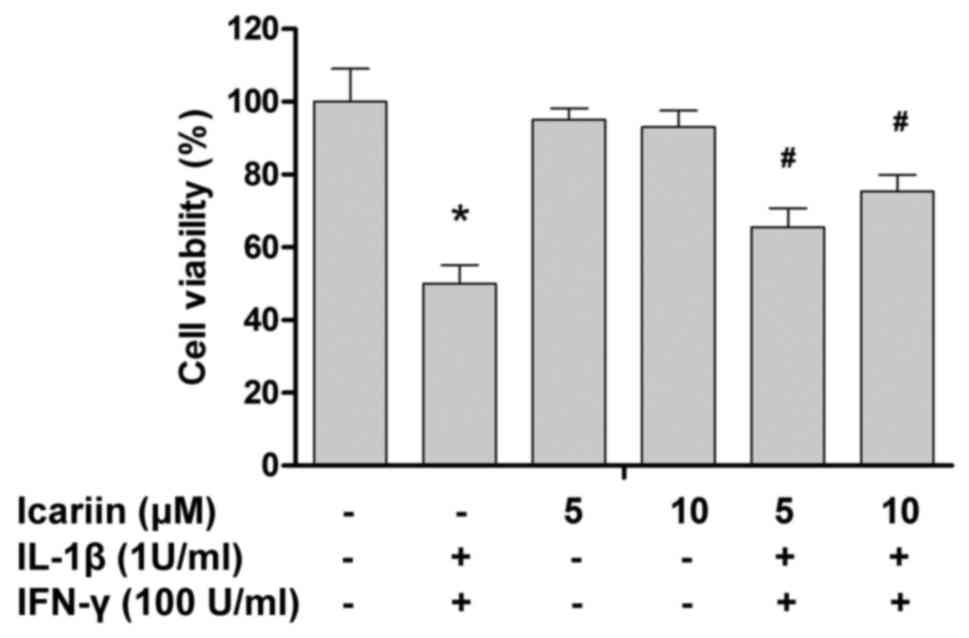
initially examined. As presented in Fig.
1, treatment with icariin up to 10 µM did not result in a
significant loss of cell viability. Next, whether icariin protected
RINm5F cells from cytokine toxicity was investigated. Treatment
with cytokines IL-1β and IFN-γ significantly reduced the cell
viability to 49.9±5.2% of the control value (Fig. 1). Pretreatment with icariin
significantly abrogated the cytotoxic effects of cytokines on
RINm5F cells in a concentration-dependent manner.
Icariin prevents cytokine-induced NO
production
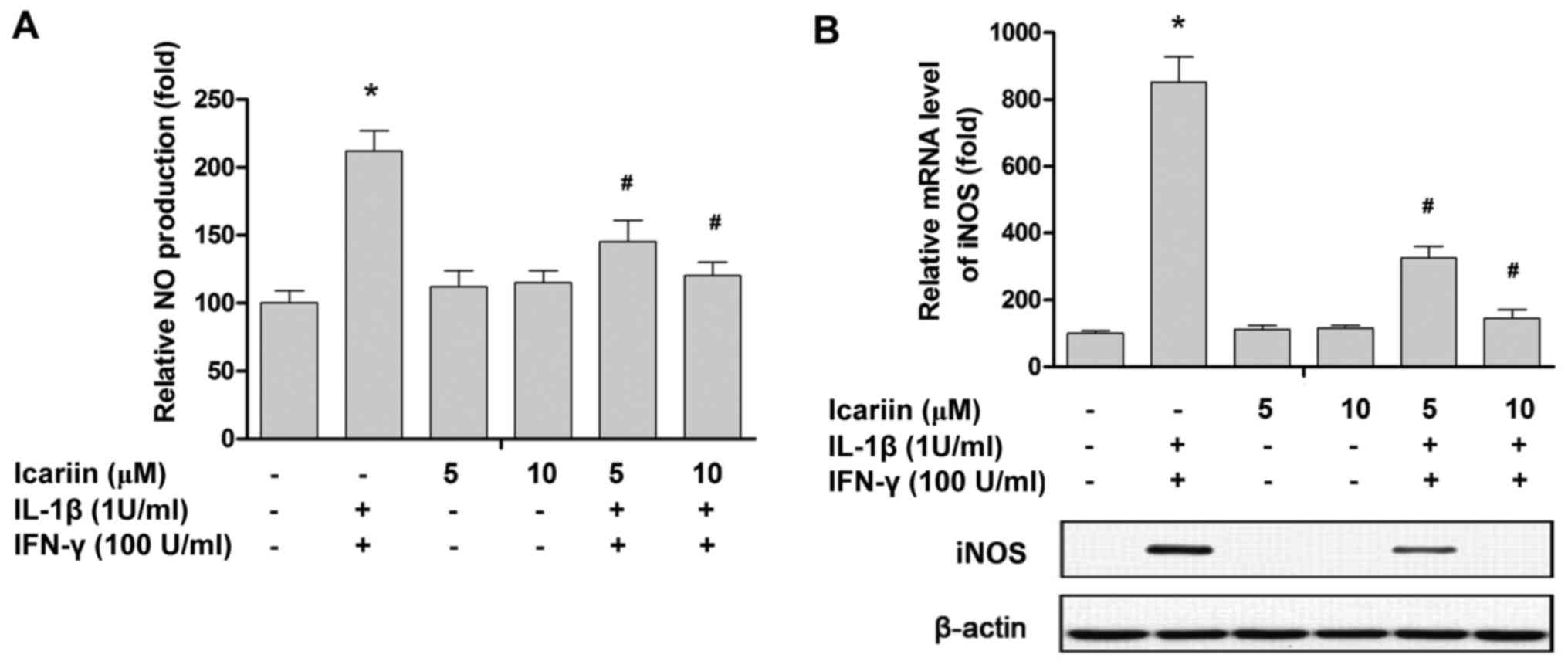
NO production was significantly increased following
24 h treatment with cytokines (Fig.
2A). However, the cytokine-induced NO production was
effectively inhibited by treatment with 10 µM icariin (Fig. 2A). To investigate the underlying
mechanisms responsible for the effects of icariin, RT-qPCR and
western blotting were performed to measure the expression of iNOS
at the mRNA and protein level, respectively. Treatment with IL-1β
and IFN-γ significantly increased the expression of iNOS, while
icariin treatment significantly ameliorated this increase at the
mRNA and protein level (Fig. 2).
Icariin prevents cytokine-induced
apoptosis
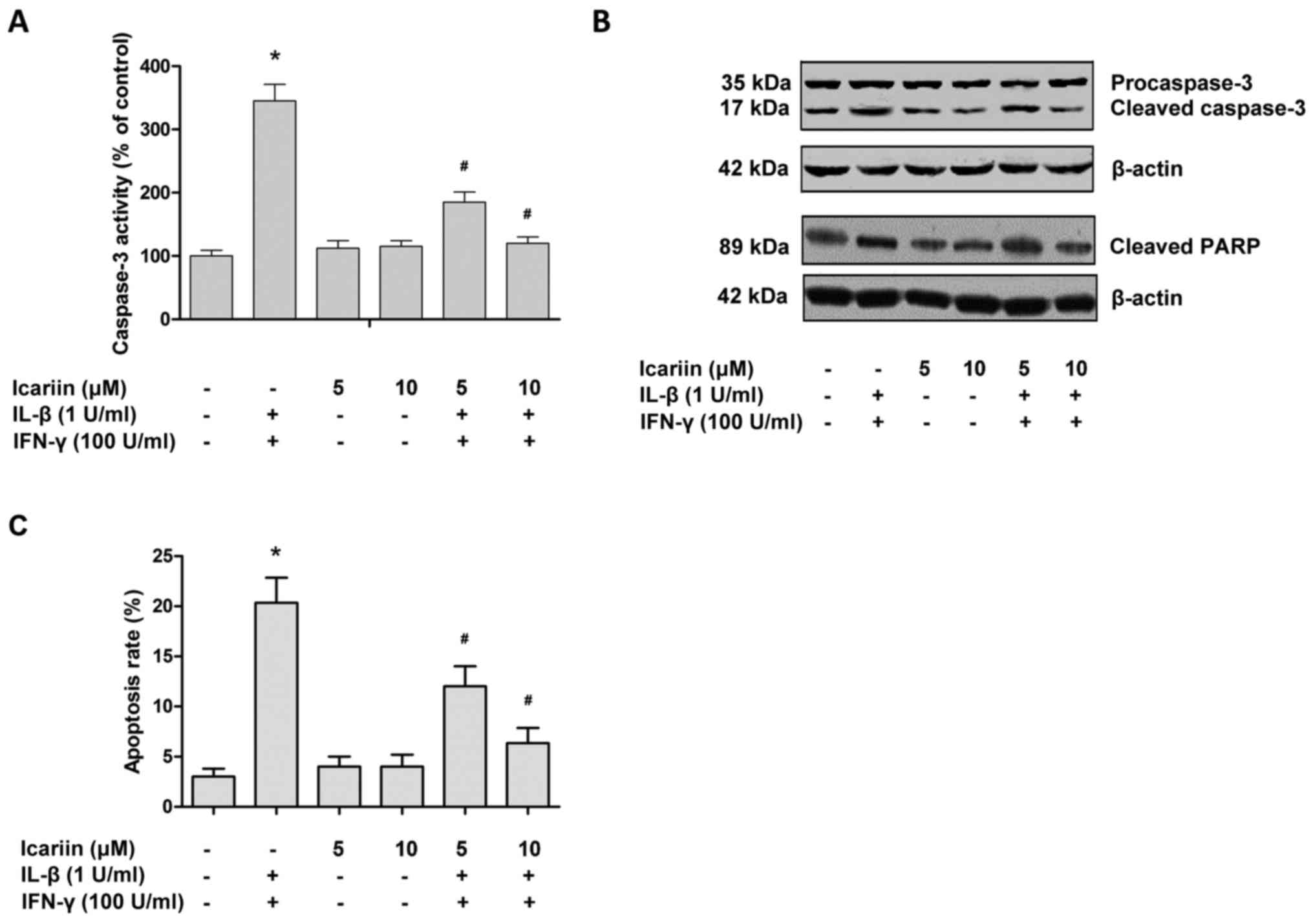
Cytokines are able to promote β-cell death through
apoptosis and necrosis (25).
Caspase-3 serves a pivotal role in the apoptotic signaling pathway,
and so the activation status of caspase-3 was assessed in the
present study. Treatment with IL-1β and IFN-γ increased the
activity of caspase-3 and cell apoptosis in RINm5F cells, while
icariin effectively reversed these effects (Fig. 3). The activation of apoptotic
signaling was also confirmed by western blotting (Fig. 3B). Cleaved caspase-3 is the main
marker of cell apoptosis (26), and
so its expression was assessed. As presented in Fig. 3B, IL-1β and IFN-γ were able to
activate caspase-3 and increase the cleavage of PARP in RINm5F
cells, while treatment with icariin reduced cleaved caspase-3 and
cleaved PARP levels in cytokine-stimulated cells.
Icariin suppresses the
cytokine-induced activation of NF-κB
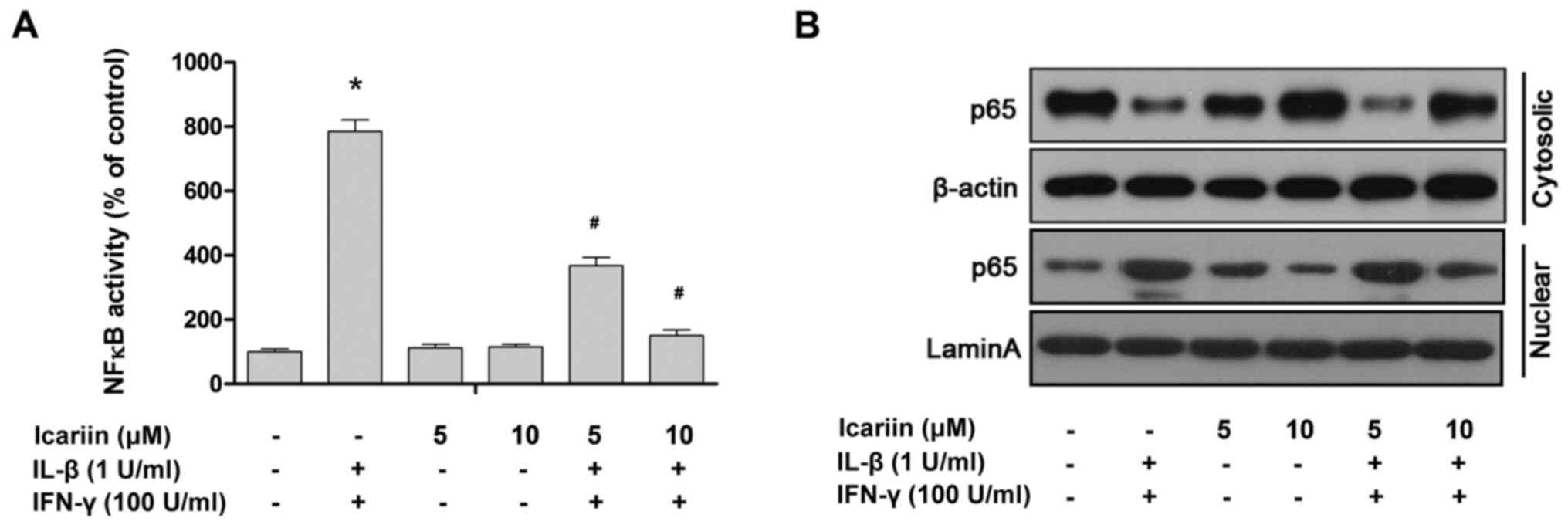
NF-κB is a key transcription factor that induces
iNOS and regulates subsequent NO production (27). The results of a previous study by our
group demonstrated that NF-κB was activated by cytokines or
oxidative stress (28). Based on
this, it was investigated whether icariin affects the
cytokine-induced activation and translocation of NF-κB from the
cytosol to the nucleus in RINm5F cells. NF-κB and the nuclear
translocation of p65, a key subunit of the NF-κB complex, were
significantly promoted by treatment with IL-1β and IFN-γ compared
with the control (Fig. 4). In
contrast, icariin pretreatment markedly suppressed the
cytokine-stimulated activation and nuclear translocation of NF-κB.
In summary, these data suggest that icariin may downregulate iNOS
expression via inhibiting the cytokine-stimulated activation of
NF-κB.
Discussion
Icariin is a biologically active flavonoid with a
favorable therapeutic profile in metabolic syndrome (29,30).
Notably, icariin has been reported to ameliorate
streptozocin-induced rat diabetic retinopathy and nephropathy
(31,32). A previous study demonstrated that
icariin could serve as a peroxisome proliferator-activated receptor
α agonist, which activates gene expression associated with lipid
metabolism in the liver to contribute towards diabetes management
(33). In the present study, it was
revealed that icariin is able to prevent cytokine-induced β-cell
death, which is an important cause of T1DM.
Inflammation is the primary cause of T1DM as well as
a direct cause of a number of diabetic complications (34). An acute, intense inflammatory
response triggers T1DM through lymphocyte-mediated destruction of
pancreatic β cells. A chronic state of low-grade inflammation
persists within in the body, which is periodically exacerbated by
hyperglycemic fluctuations (34–35).
Increased inflammation markers (35), immune activation (36) and oxidative stress have been recorded
in patients with T1DM (37,38). It has therefore been hypothesized
that anti-inflammatory agents may be an effective clinical
treatment for patients with T1DM. A number of in vivo and
in vitro studies have confirmed the anti-inflammatory effect
of icariin, including in the brain, heart, bones and airways
(22,23,39–50). The
present study demonstrated that icariin prevents viability loss in
rat pancreatic β cells, as well as suppressing cytokine-induced NO
production and apoptosis activation. These results suggest that
icariin may interfere with the inflammatory response and resulting
pancreatic β cell death during T1DM.
Furthermore, a key factor in cytokine-induced
pancreatic β-cell damage is NF-κB. In vivo studies of
transgenic mice revealed that NF-κB inhibition is a protective
mechanism against cytokine-induced apoptosis in pancreatic β-cells
(28). In addition, the use of
A20-overexpressing islets to abrogate NF-κB signaling during islet
transplantation reduces the number of islets required to achieve
euglycemia in diabetic recipients (51). Therefore, suppression of the NF-κB
pathway may also be a novel strategy for delaying the progression
of T1DM. The regulatory role of icariin on NF-κB has been reported,
however it may vary between different cell types (20). Xu and Huang (52) demonstrated that icariin could
increase the expression of endothelial NOS in human endothelial
cells, which was implicated in the activation of NF-κB (53). In contrast, icariin was able to
abrogate the effects of LPS on neuroinflammation, lung
inflammation, osteoclast differentiation and bone resorption via
decreasing NF-κB activity (23,49,54,55). It
has also been reported that icariin inhibits NF-κB activity in a
wide range of cancerous cells (56–59). The
results of the present study demonstrated that icariin suppresses
the cytokine-induced activation of NF-κB in rat pancreatic β cells.
It is likely that icariin exerts cell-specific regulatory effects
and only suppresses high levels of NF-κB activity in tumor cells or
cells stimulated by inflammatory cytokines.
The results of the present study demonstrate that
icariin abrogates the pro-apoptotic effect of cytokines and
significantly suppresses NF-κB activation in rat pancreatic
β-cells. Despite being used extensively as a model for the human
pancreas, the physiology of rat pancreatic β cells does not
perfectly mimic that of primary cells (60). The RINm5F cells used in the present
study have limitations in terms of glucose sensitivity, transport
and phosphorylation (60,61). Therefore, experiments utilizing human
pancreatic cells and in vivo analysis are required to
confirm these findings. Nevertheless, the results of the present
study suggest that icariin may have potential as a therapeutic
agent against T1DM.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81573911).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
SZ and J-YY conceived the study, acquired data,
interpreted the results and drafted the manuscript. JG made
substantial contributions to the experiments and data analysis.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Philippe J and Raccah D: Treating type 2
diabetes: How safe are current therapeutic agents? Int J Clin
Pract. 63:321–332. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Akkati S, Sam KG and Tungha G: Emergence
of promising therapies in diabetes mellitus. J Clin Pharmacol.
51:796–804. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
King H, Aubert RE and Herman WH: Global
burden of diabetes, 1995–2025: Prevalence, numerical estimates and
projections. Diabetes Care. 21:1414–1431. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
American Diabetes Association, : Diagnosis
and classification of diabetes mellitus. Diabetes Care. 31 Suppl
1:S55–S60. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Patterson CC, Gyurus E, Rosenbauer J,
Cinek O, Neu A, Schober E, Parslow RC, Joner G, Svensson J, Castell
C, et al: Trends in childhood type 1 diabetes incidence in Europe
during 1989–2008: Evidence of non-uniformity over time in rates of
increase. Diabetologia. 55:2142–2147. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Patterson CC, Dahlquist GG, Gyurus E,
Green A and Soltész G; EURODIAB Study Group, : Incidence trends for
childhood type 1 diabetes in Europe during 1989–2003 and predicted
new cases 2005–20: a multicentre prospective registration study.
Lancet. 373:2027–2033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Redondo MJ, Jeffrey J, Fain PR, Eisenbarth
GS and Orban T: Concordance for islet autoimmunity among
monozygotic twins. N Engl J Med. 359:2849–2850. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Davidson MH: Cardiovascular risk factors
in a patient with diabetes mellitus and coronary artery disease:
Therapeutic approaches to improve outcomes: Perspectives of a
preventive cardiologist. Am J Cardiol. 110 9 Suppl:43B–49B. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Robertson RP: Islet transplantation a
decade later and strategies for filling a half-full glass.
Diabetes. 59:1285–1291. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ehses JA, Perren A, Eppler E, Ribaux P,
Pospisilik JA, Maor-Cahn R, Gueripel X, Ellingsgaard H, Schneider
MK, Biollaz G, et al: Increased number of islet-associated
macrophages in type 2 diabetes. Diabetes. 56:2356–2370. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Eizirik DL, Sammeth M, Bouckenooghe T,
Bottu G, Sisino G, Igoillo-Esteve M, Ortis F, Santin I, Colli ML,
Barthson J, et al: The human pancreatic islet transcriptome:
expression of candidate genes for type 1 diabetes and the impact of
pro-inflammatory cytokines. PLoS Genet. 8:e10025522012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wellen KE and Hotamisligil GS:
Inflammation, stress and diabetes. J Clin Invest. 115:1111–1119.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Arif S, Moore F, Marks K, Bouckenooghe T,
Dayan CM, Planas R, Vives-Pi M, Powrie J, Tree T, Marchetti P, et
al: Peripheral and islet interleukin-17 pathway activation
characterizes human autoimmune diabetes and promotes
cytokine-mediated beta-cell death. Diabetes. 60:2112–2119. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Atkinson MA, Eisenbarth GS and Michels AW:
Type 1 diabetes. Lancet. 383:69–82. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Baldwin AC, Green CD, Olson LK, Moxley MA
and Corbett JA: A role for aberrant protein palmitoylation in
FFA-induced ER stress and beta-cell death. Am J Physiol Endocrinol
Metab. 302:E1390–E1398. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Imai Y, Dobrian AD, Weaver JR, Butcher MJ,
Cole BK, Galkina EV, Morris MA, Taylor-Fishwick DA and Nadler JL:
Interaction between cytokines and inflammatory cells in islet
dysfunction, insulin resistance and vascular disease. Diabetes Obes
Metab. 15 Suppl 3:S117–S129. 2013. View Article : Google Scholar
|
|
17
|
Cnop M, Welsh N, Jonas JC, Jörns A, Lenzen
S and Eizirik DL: Mechanisms of pancreatic beta-cell death in type
1 and type 2 diabetes: many differences, few similarities.
Diabetes. 54 Suppl 2:S97–S107. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Eizirik DL, Colli ML and Ortis F: The role
of inflammation in insulitis and beta-cell loss in type 1 diabetes.
Nat Rev Endocrinol. 5:219–226. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Corbett JA and McDaniel ML: Does nitric
oxide mediate autoimmune destruction of beta-cells? Possible
therapeutic interventions in IDDM. Diabetes. 41:897–903. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen Y, Huang JH, Ning Y and Shen ZY:
Icariin and its pharmaceutical efficacy: research progress of
molecular mechanism. Zhong Xi Yi Jie He Xue Bao. 9:1179–1184.
2011.(In Chinese). View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu JF, Dong JC and Xu CQ: Effects of
icariin on inflammation model stimulated by lipopolysaccharide in
vitro and in vivo. Zhongguo Zhong Xi Yi Jie He Za Zhi. 29:330–334.
2009.(In Chinese). PubMed/NCBI
|
|
22
|
Liu MH, Sun JS, Tsai SW, Sheu SY and Chen
MH: Icariin protects murine chondrocytes from
lipopolysaccharide-induced inflammatory responses and extracellular
matrix degradation. Nutr Res. 30:57–65. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu CQ, Liu BJ, Wu JF, Xu YC, Duan XH, Cao
YX and Dong JC: Icariin attenuates LPS-induced acute inflammatory
responses: involvement of PI3K/Akt and NF-kappaB signaling pathway.
Eur J Pharmacol. 642:146–153. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bae UJ, Lee da Y, Song MY, Lee SM, Park
JW, Ryu JH and Park BH: A prenylated flavan from Broussonetia
kazinoki prevents cytokine-induced beta-cell death through
suppression of nuclear factor-kB activity. Biol Pharm Bull.
34:1026–1031. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Saldeen J: Cytokines induce both necrosis
and apoptosis via a common Bcl-2-inhibitable pathway in rat
insulin-producing cells. Endocrinology. 141:2003–2010. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tewari M, Quan LT, O'Rourke K, Desnoyers
S, Zeng Z, Beidler DR, Poirier GG, Salvesen GS and Dixit VM:
Yama/CPP32 beta, a mammalian homolog of CED-3, is a
CrmA-inhibitable protease that cleaves the death substrate poly
(ADP-ribose) polymerase. Cell. 81:801–809. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Heimberg H, Heremans Y, Jobin C, Leemans
R, Cardozo AK, Darville M and Eizirik DL: Inhibition of
cytokine-induced NF-kappaB activation by adenovirus-mediated
expression of a NF-kappaB super-repressor prevents beta-cell
apoptosis. Diabetes. 50:2219–2224. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Eldor R, Yeffet A, Baum K, Doviner V, Amar
D, Ben-Neriah Y, Christofori G, Peled A, Carel JC, Boitard C, et
al: Conditional and specific NF-kappaB blockade protects pancreatic
beta cells from diabetogenic agents. Proc Natl Acad Sci USA.
103:5072–5077. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gong Y, Shi J, Xie GY, Liu HR and Qi MY:
Amelioration of icariin for the epididymis impairment induced by
streptozocin (STZ) in rats. Zhongguo Ying Yong Sheng Li Xue Za Zhi.
29:47–50. 2013.(In Chinese). PubMed/NCBI
|
|
30
|
Zhang WP, Bai XJ, Zheng XP, Xie XL and
Yuan ZY: Icariin attenuates the enhanced prothrombotic state in
atherosclerotic rabbits independently of its lipid-lowering
effects. Planta Med. 79:731–736. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xin H, Zhou F, Liu T, Li GY, Liu J, Gao
ZZ, Bai GY, Lu H and Xin ZC: Icariin ameliorates
streptozotocin-induced diabetic retinopathy in vitro and in vivo.
Int J Mol Sci. 13:866–878. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qi MY, Kai C, Liu HR, Su YH and Yu SQ:
Protective effect of Icariin on the early stage of experimental
diabetic nephropathy induced by streptozotocin via modulating
transforming growth factor beta1 and type IV collagen expression in
rats. J Ethnopharmacol. 138:731–736. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lu YF, Xu YY, Jin F, Wu Q, Shi JS and Liu
J: Icariin is a PPARα activator inducing lipid metabolic gene
expression in mice. Molecules. 19:18179–18191. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tran B, Oliver S, Rosa J and Galassetti P:
Aspects of inflammation and oxidative stress in pediatric obesity
and type 1 diabetes: An overview of ten years of studies. Exp
Diabetes Res. 2012:6836802012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Devaraj S, Dasu MR, Rockwood J, Winter W,
Griffen SC and Jialal I: Increased toll-like receptor (TLR) 2 and
TLR4 expression in monocytes from patients with type 1 diabetes:
Further evidence of a proinflammatory state. J Clin Endocrinol
Metab. 93:578–583. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Devaraj S, Glaser N, Griffen S,
Wang-Polagruto J, Miguelino E and Jialal I: Increased monocytic
activity and biomarkers of inflammation in patients with type 1
diabetes. Diabetes. 55:774–779. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yamagishi S: Advanced glycation end
products and receptor-oxidative stress system in diabetic vascular
complications. Ther Apher Dial. 13:534–539. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Maritim AC, Sanders RA and Watkins JB III:
Diabetes, oxidative stress and antioxidants: A review. J Biochem
Mol Toxicol. 17:24–38. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen Y, Sun T, Wu J, Kalionis B, Zhang C,
Yuan D, Huang J, Cai W, Fang H and Xia S: Icariin intervenes in
cardiac inflammaging through upregulation of SIRT6 enzyme activity
and inhibition of the NF-Kappa B pathway. Biomed Res Int.
2015:8959762015.PubMed/NCBI
|
|
40
|
Zhou H, Yuan Y, Liu Y, Ni J, Deng W, Bian
ZY, Dai J and Tang QZ: Icariin protects H9c2 cardiomyocytes from
lipopolysaccharide-induced injury via inhibition of the reactive
oxygen species-dependent c-Jun N-terminal kinases/nuclear factor-kB
pathway. Mol Med Rep. 11:4327–4332. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wei Y, Liu B, Sun J, Lv Y, Luo Q, Liu F
and Dong J: Regulation of Th17/Treg function contributes to the
attenuation of chronic airway inflammation by icariin in
ovalbumin-induced murine asthma model. Immunobiology. 220:789–797.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shen R, Deng W, Li C and Zeng G: A natural
flavonoid glucoside icariin inhibits Th1 and Th17 cell
differentiation and ameliorates experimental autoimmune
encephalomyelitis. Int Immunopharmacol. 24:224–231. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chi L, Gao W, Shu X and Lu X: A natural
flavonoid glucoside, icariin, regulates Th17 and alleviates
rheumatoid arthritis in a murine model. Mediators Inflamm.
2014:3920622014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li L, Sun J, Xu C, Zhang H, Wu J, Liu B
and Dong J: Icariin ameliorates cigarette smoke induced
inflammatory responses via suppression of NF-kB and modulation of
GR in vivo and in vitro. PLoS One. 9:e1023452014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cui J, Zhu M, Zhu S, Wang G, Xu Y and Geng
D: Inhibitory effect of icariin on Ti-induced inflammatory
osteoclastogenesis. J Surg Res. 192:447–453. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tao F, Qian C, Guo W, Luo Q, Xu Q and Sun
Y: Inhibition of Th1/Th17 responses via suppression of STAT1 and
STAT3 activation contributes to the amelioration of murine
experimental colitis by a natural flavonoid glucoside icariin.
Biochem Pharmacol. 85:798–807. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhou J, Wu J, Chen X, Fortenbery N,
Eksioglu E, Kodumudi KN, Pk EB, Dong J, Djeu JY and Wei S: Icariin
and its derivative, ICT, exert anti-inflammatory, anti-tumor
effects and modulate myeloid derived suppressive cells (MDSCs)
functions. Int Immunopharmacol. 11:890–898. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen SR, Xu XZ, Wang YH, Chen JW, Xu SW,
Gu LQ and Liu PQ: Icariin derivative inhibits inflammation through
suppression of p38 mitogen-activated protein kinase and nuclear
factor-kappaB pathways. Biol Pharm Bull. 33:1307–1313. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hsieh TP, Sheu SY, Sun JS and Chen MH:
Icariin inhibits osteoclast differentiation and bone resorption by
suppression of MAPKs/NF-kB regulated HIF-1α and PGE (2) synthesis.
Phytomedicine. 18:176–185. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Guo J, Li F, Wu Q, Lu Y and Shi J:
Protective effects of icariin on brain dysfunction induced by
lipopolysaccharide in rats. Phytomedicine. 17:950–955. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Grey ST, Longo C, Shukri T, Patel VI,
Csizmadia E, Daniel S, Arvelo MB, Tchipashvili V and Ferran C:
Genetic engineering of a suboptimal islet graft with A20 preserves
beta cell mass and function. J Immunol. 170:6250–6256. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xu HB and Huang ZQ: Icariin enhances
endothelial nitric-oxide synthase expression on human endothelial
cells in vitro. Vascul Pharmacol. 47:18–24. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wo Y, Zhu D, Yu Y and Lou Y: Involvement
of NF-kappaB and AP-1 activation in icariin promoted cardiac
differentiation of mouse embryonic stem cells. Eur J Pharmacol.
586:59–66. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zeng KW, Fu H, Liu GX and Wang XM: Icariin
attenuates lipopolysaccharide-induced microglial activation and
resultant death of neurons by inhibiting TAK1/IKK/NF-kappaB and
JNK/p38 MAPK pathways. Int Immunopharmacol. 10:668–678. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wu W, Pan C, Yu H, Gong H and Wang Y:
Heparanase expression in gallbladder carcinoma and its correlation
to prognosis. J Gastroenterol Hepatol. 23:491–497. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yang L, Wang Y, Guo H and Guo M:
Synergistic anti-cancer effects of icariin and temozolomide in
glioblastoma. Cell Biochem Biophys. 71:1379–1385. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Han H, Xu B, Hou P, Jiang C, Liu L, Tang
M, Yang X, Zhang Y and Liu Y: Icaritin sensitizes human
glioblastoma cells to TRAIL-induced apoptosis. Cell Biochem
Biophys. 72:533–542. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang Y, Wei Y, Zhu Z, Gong W, Liu X, Hou
Q, Sun Y, Chai J, Zou L and Zhou T: Icariin enhances
radiosensitivity of colorectal cancer cells by suppressing NF-kB
activity. Cell Biochem Biophys. 69:303–310. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Shi DB, Li XX, Zheng HT, Li DW, Cai GX,
Peng JJ, Gu WL, Guan ZQ, Xu Y and Cai SJ: Icariin-mediated
inhibition of NF-kB activity enhances the in vitro and in vivo
antitumour effect of 5-fluorouracil in colorectal cancer. Cell
Biochem Biophys. 69:523–530. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Gazdar AF, Chick WL, Oie HK, Sims HL, King
DL, Weir GC and Lauris V: Continuous, clonal, insulin- and
somatostatin-secreting cell lines established from a transplantable
rat islet cell tumor. Proc Natl Acad Sci USA. 77:3519–3523. 1980.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Praz GA, Halban PA, Wollheim CB, Blondel
B, Strauss AJ and Renold AE: Regulation of immunoreactive-insulin
release from a rat cell line (RINm5F). Biochem J;. 210:345–352.
1983. View Article : Google Scholar : PubMed/NCBI
|