Introduction
Cardiovascular disease (CVD) is the main cause of
mortality in patients with chronic kidney disease (CKD), with a
10–20-fold higher risk compared to the age and sex-matched general
population (1). One of the
potentially life-threatening cardiovascular (CV) consequences of
CKD are vascular calcifications (VCs), which contribute to the high
prevalence of CV morbidity and mortality in CKD (2–4).
Furthermore, risk factors for the VC of CKD patients include
abnormal phosphorus and calcium metabolism, bone disorders,
phenotype transformation of vascular smooth muscle cells (VSMCs)
and deficiency of endogenous calcification inhibitors including
Klotho, matrix Gla protein (MGP), pyrophosphate and fetuin-A
(5–7).
It has previously been reported that the
pathogenesis of VC is similar to bone formation (8–10). One
of the inhibitors of VC is the vitamin K-dependent protein (VKDPs),
MGP, which regulates ectopic mineralization. Vitamin K functions as
a cofactor in the enzymatic reaction that converts specific
glutamate residues in MGP into g-carboxyglutamate ones, which are
needed for its ability to inhibit calcification (11–13).
Furthermore, vitamin K has the same role in the process of
converting growth arrest-specific-6, which can also prevent VC, to
their biologically active forms.
Several previous studies have shown that a
suboptimal vitamin K status is common in patients with CKD
(14–16). Insufficient levels of vitamin K
result in VKDPs that are functionally inactive and could therefore
represent a modifiable risk factor for VC in CKD patients.
Additionally, vitamin K is a fat-soluble compound and consists of
two natural forms, vitamins K1 and K2. Both vitamins function as
cofactors for the enzyme γ-glutamylcarboxylase. Menaquinone (MK)-4
is a member of the vitamin K2 family, and there is data of the
Rotterdam study (17) hypothesizing
that vitamin K2 is of greater importance than K1 for vascular
health. Additionally, MK-4 deficiency is a predictor of aortic
calcification (18). To the best of
our knowledge, there are no previous studies that evaluate the
effect of MK-4 on the VSMCs in a β-glycerophosphate (β-GP)-induced
VC model and the target genes in the regulation of VSMCs
differentiation. Therefore, the primary aim of the present study
was to investigate how MK-4 inhibited the VC and to characterize
the underlying mechanisms.
Materials and methods
Cell culture of VSMCs
Rat VSMCs were isolated from the thoracic aorta
tunica media of adult male Sprague Dawley rats (Experimental Animal
Center of Hebei Medical University, Hebei, China) adopting the
method of explant culture as previously described (19). Briefly, the rats were anesthetized by
400 mg/kg chloral hydrate and sacrificed via cervical dislocation.
The thoracic aorta was then removed in gnotobasis. The thoracic
aorta was then cut into 1–2 mm2 pieces following the
washing off of any residual blood. The tissue pieces were then
cultured in dishes with Dulbecco's modified Eagle's medium (DMEM;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 15% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 4.5 g glucose, 100 µg/ml streptomycin and 100
U/ml penicillin in a 5% CO2 incubator at 37°C. Cells
that migrated from tissue pieces were collected when they reached
confluence. Furthermore, the cells were cultured in DMEM
replenished with 15% FBS, and the medium was changed twice per
week. VSMCs were identified by positive staining of α-smooth muscle
actin (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and used for
all experiments between generations 3–4.
The current study conformed to the Guide for Care
and Use of Laboratory Animals as adopted and promulgated by the
United National Institutes of Health. All experimental protocols
were approved by the Review Committee for the Use of Animal
Subjects of Hebei Medical University (Shijiazhuang, China).
Calcification assays
In order to induce calcification, the VSMCs were
incubated with a calcifying medium, which consisted of growth
medium supplemented with 10 mM β-GP (Sigma-Aldrich; Merck KGaA). In
some experiments, the cells were exposed to calcifying medium
accreted with 10, 25 and 50 µM of MK-4 (Sigma-Aldrich; Merck KGaA).
After incubation for 10 days, the cells were washed twice with
phosphate-buffered saline and fixed with 95% ethanol. Next, the
cells were incubated with 0.2% Alizarin red [(pH 8.3); Beijing
Solarbio Science & Technology Co., Ltd., Beijing, China].
Subsequent to washing, an LH50A inverted phase contrast microscope
(Olympus Corporation, Tokyo, Japan) was used for visualizing cells
and capturing images to record the incidence of induced
calcification. NIS-Element F3.0 software was used to capture the
images (Olympus Corporation). Subsequently, calcium deposited in
the extracellular matrix was treated with 0.6 M HCl for 24 h at
37°C, and the content of calcium in the supernatant was measured
with the o-cresolphthalein complexone method using a calcium assay
kit (BioSino Biotechnology & Science, Inc., Beijing, China).
This was then normalized relative to the protein concentration of
the same culture.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was extracted from VSMCs by reference
methods using TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) in accordance with the manufacturer's instructions, and
quantified by an UV absorbance of 260–280 nm. In total, 2 µg RNA
was used for synthesizing cDNA with the RT-for-PCR kit (Clontech
Laboratories, Inc., Mountainview, CA, USA) with oligo (dT) primers
as recommended in the protocol provided. cDNA was used as regular
RT-PCR template. Furthermore, the internal control was the
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene. All of the
primers used are listed in Table I.
PCR was performed using the PCR Master Mix kit (Promega
Corporation, Madison, WI, USA) according to the manusfacturer's
protocol and purified prior to sequencing. The PCR conditions were
as follows: Initial incubation for 2 min at 95°C; 35 cycles of 30
sec at 95°C for denaturation; 30 sec at 55°C for annealing; 45 sec
at 72°C for extension; and a final extension at 72°C for 5 min.
Cycle sequencing was performed with the Dye Terminator Cycle
Sequencing Ready Reaction kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.). For regular RT-PCR, the PCR products were
detected by a 1.5% agarose gel with electrophoresis and visualized
by staining with ethidium bromide. Furthermore, the reaction of
each sample was repeated once for quality control. The optical
density of the band was measured using a Gel Documentation System
(CST Biological Reagents Co., Ltd., Shanghai, China) and the final
data is expressed as the mRNA level relative to that of GAPDH.
 | Table I.Primers used in reverse transcription
quantitative polymerase chain reaction. |
Table I.
Primers used in reverse transcription
quantitative polymerase chain reaction.
|
| Primer |
|---|
|
|
|
|---|
| Gene | Forward | Reverse |
|---|
| Runx2 |
5′-CCGCACGACAACCGCACCAT-3′ |
5′-CGCTCCGGCCCACAAATCTC-3′ |
| BMP-2 |
5′-ACTCGAAATTCCCCGTGACC-3′ |
5′-CCACTTCCACCACGAATCCA-3′ |
| SMAD1 |
5′-GGTGACTGGGAACGGATCG-3′ |
5′-TGGTCTTCGGTTCGGAAAGG-3′ |
| SMAD7 |
5′-CCTCGGAAGTCAAGAGGCTG-3′ |
5′-CCTCGGAAGTCAAGAGGCTG-3′ |
| GAPDH |
5′-CAAGGTCATCCATGACAACTTTG-3′ |
5′-GTCCACCACCCTGTTGCTGTAG-3′ |
Western blot analysis
Total protein was collected from the VSMCs, and the
concentrations were measured with the bicinchoninic acid protein
assay kit (Sigma-Aldrich; Merck KGaA). Equivalent protein (50 µg)
from VSMCs were separated by 10% SDS-PAGE and electrotransferred
onto a polyvinylidene fluoride membrane (Amersham; GE Healthcare
Life Sciences, Chalfont, UK). Non-specific protein binding was
blocked with 5% non-fat dry milk in Tris-buffered saline and
Tween-20 [TBS-T; (20 mmol/l Tris-HCl (pH 7.6), 150 mmol/l NaCl and
0.02% Tween-20; Invitrogen, Carlsbad, CA, USA] by incubating the
membrane for 1 h at room temperature with agitation. Anti-mouse
runt-related transcription factor 2 (Runx2) primary monoclonal
antibody (cat. no. ab76956; Abcam, Cambridge, UK) was incubated
with the membrane at a 1:500 dilution in TBS-T and at 4°C overnight
with agitation. The secondary antibody (horesradish perocidase
conjugated Goat anti-Mouse Immunoglobulin G; cat. no. 074-1806;
KPL, Inc., Gaithersburg, MD, USA) was diluted in TBS-T at 1:2,000
dilution and applied to the membrane, and the reaction was
incubated at room temperature for 1 h with agitation. Between each
of the three proceeding steps (primary antibody, secondary antibody
and visualization) the membrane was washed 3 times with TBS-T for
15 min at room temperature. The membrane was then immediately
visualized and analyzed using the enhanced chemiluminescence
detection system (Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
according to the manufacturer's instructions. Furthermore, β-actin
was used as the endogenous control and the experiments were
repeated three times.
Statistical analysis
Results are presented as the mean ± standard
deviation, and SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA)
was used for the analysis of variance and Dunnett's test. P<0.05
was considered to indicate a statistically significant
difference.
Results
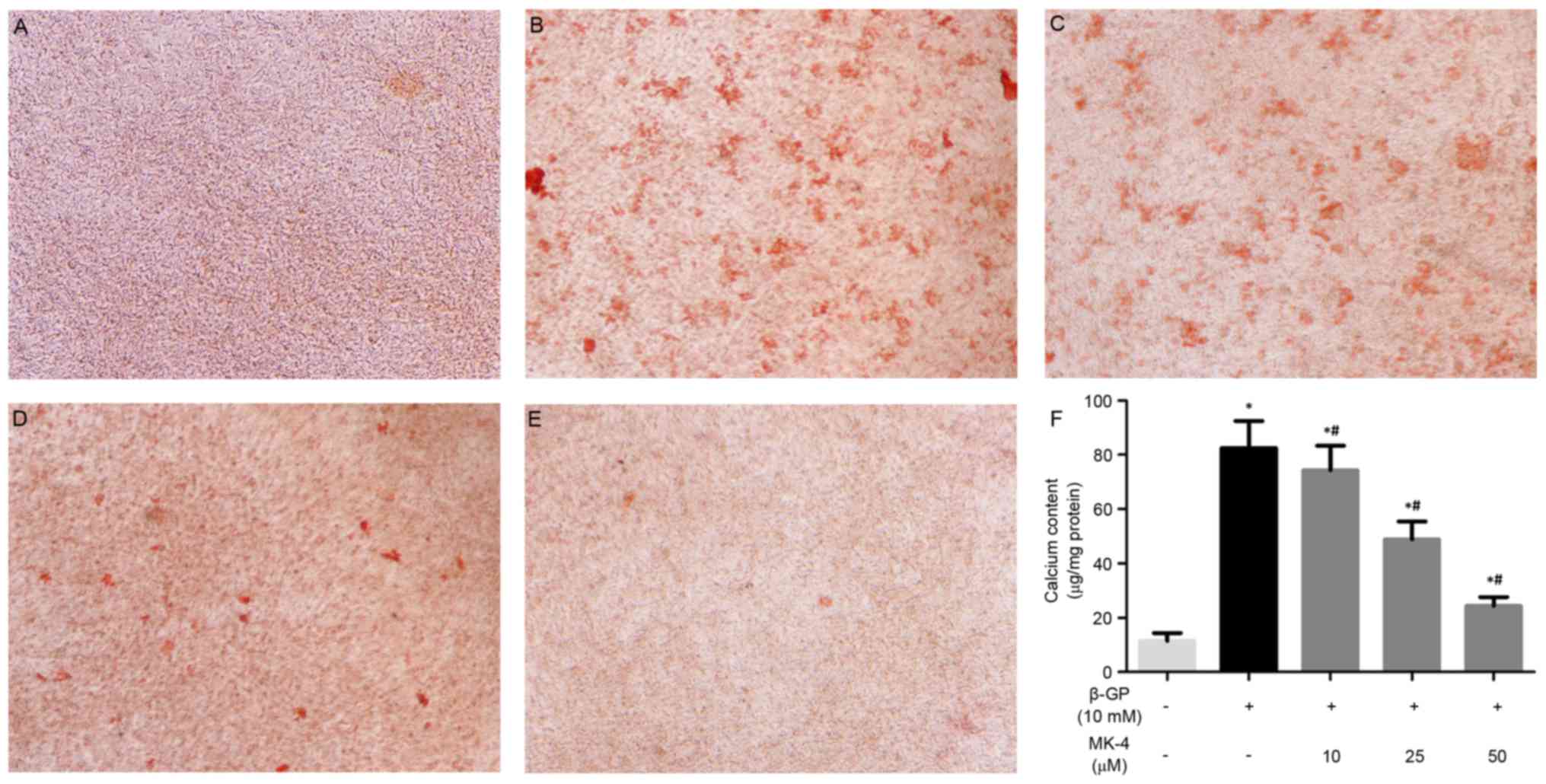
MK-4 attenuates calcification in VSMCs
induced by β-GP
The present study investigated the effects of MK-4
on the calcification of VSMCs induced by β-GP as previously
described (20). MK-4 significantly
inhibited calcium deposition of VSMCs induced by β-GP at 10 days
and in a dose-dependent manner (Fig.
1A-E). In agreement, the quantitative analysis indicated that
the calcium content was noticeably reduced in VSMCs maintained in
the β-GP + MK-4 medium compared with β-GP medium (Fig. 1F). These results suggest that MK-4
functions in suppressing the calcification of VSMCs induced by
β-GP.
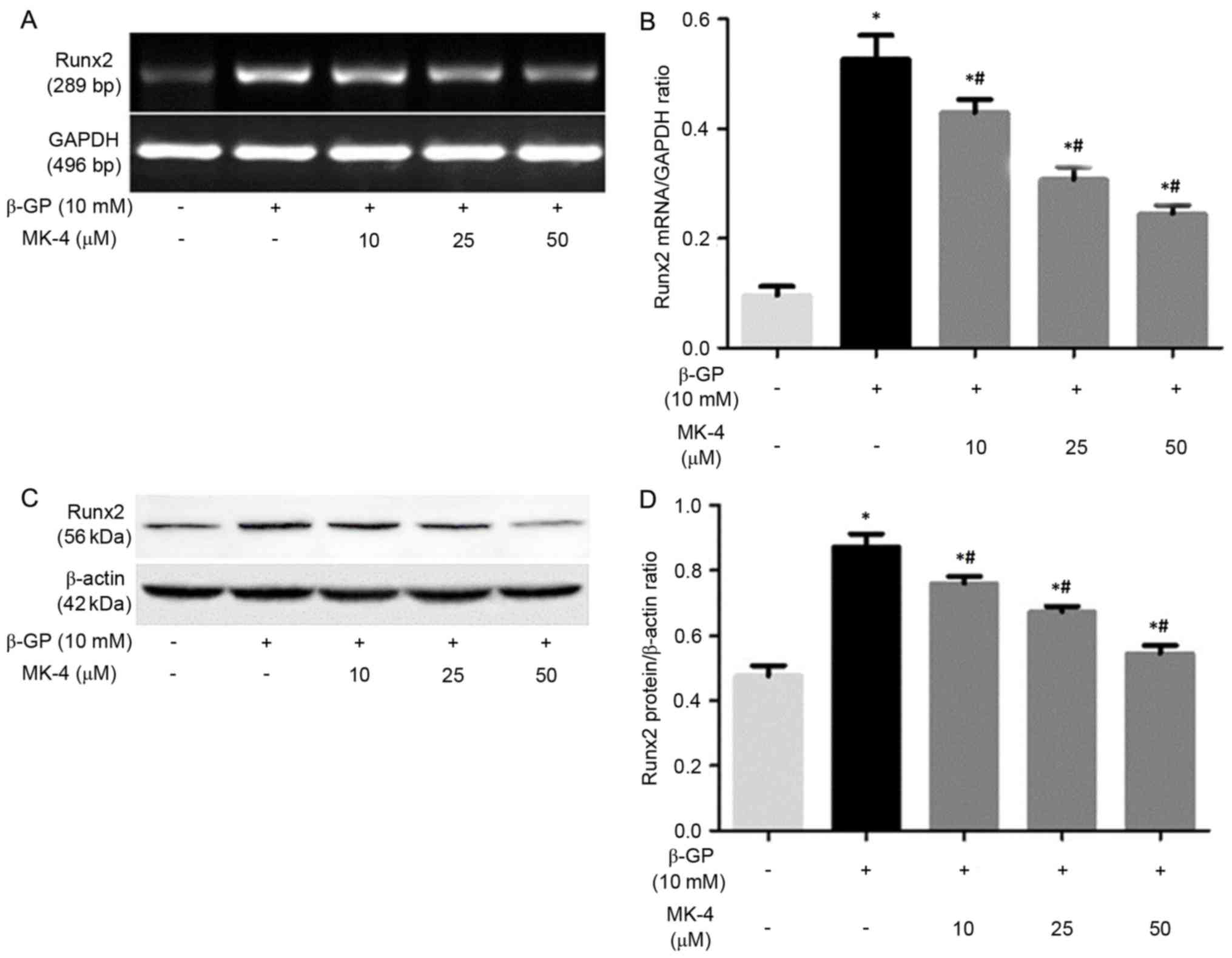
MK-4 inhibits Runx2 expression of
VSMCs induced by β-GP
It has been demonstrated that VSMCs enhanced
mineralization under calcifying conditions by phenotype
transformation of VSMCs, which is a strictly regulated cellular
process that is similar to bone formation (21). Thus, the present study assessed the
effects of β-GP on the expression of Runx2 in the presence or
absence of MK-4 medium. It was revealed that the expression of
Runx2 mRNA and protein in the VSMCs incubated with the β-GP + MK-4
medium evidently decreased, with the effect remaining enhanced in a
dose-dependent manner (Fig. 2).
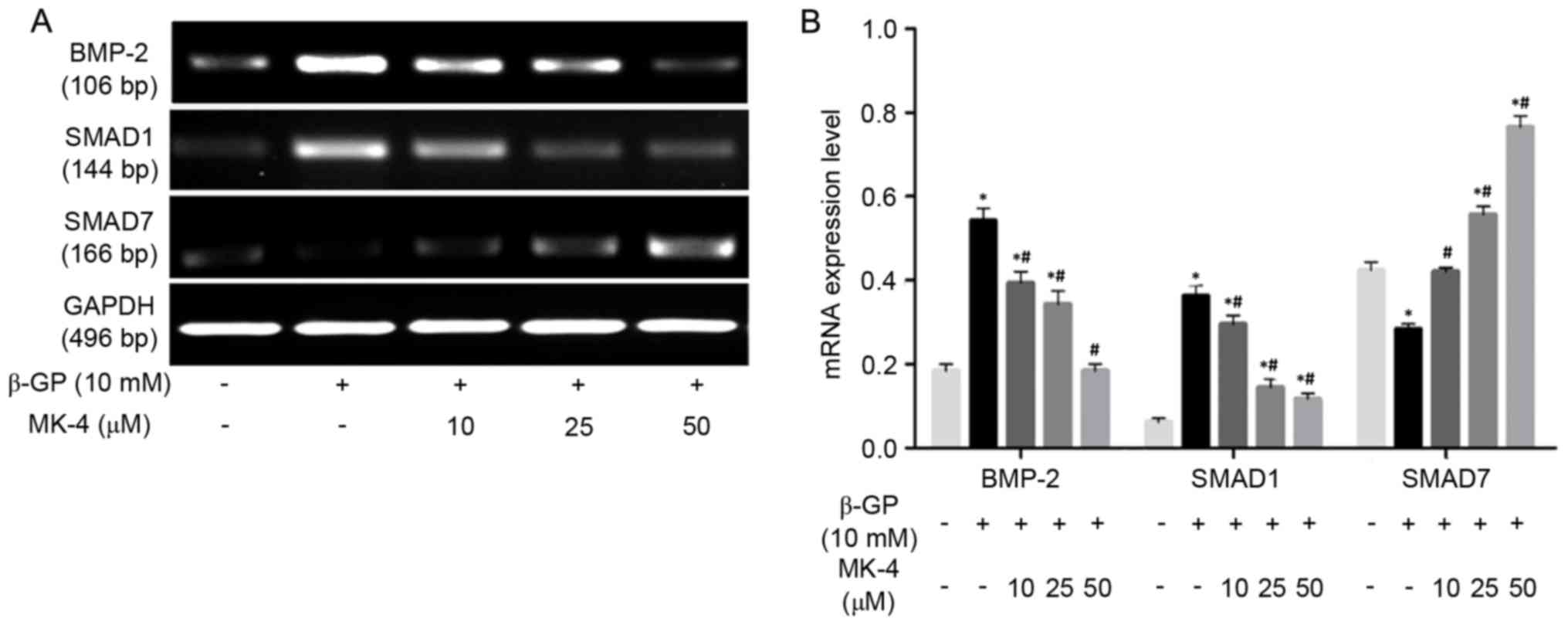
Effects of MK-4 on bone morphogenetic
protein-2 (BMP-2), SMAD1, SMAD7 mRNA expression of VSMCs induced by
β-GP
RT-PCR was performed to detect the mRNA expression
of BMP-2, SMAD1, SMAD7 in VSMCs induced by β-GP in the presence or
absence of MK-4. The results revealed that MK-4 significantly
reduced BMP-2 and SMAD1 mRNA expression in a dose-dependent manner,
while MK-4 markedly increased the expression of SMAD7 mRNA in a
dose-dependent manner (Fig. 3). In
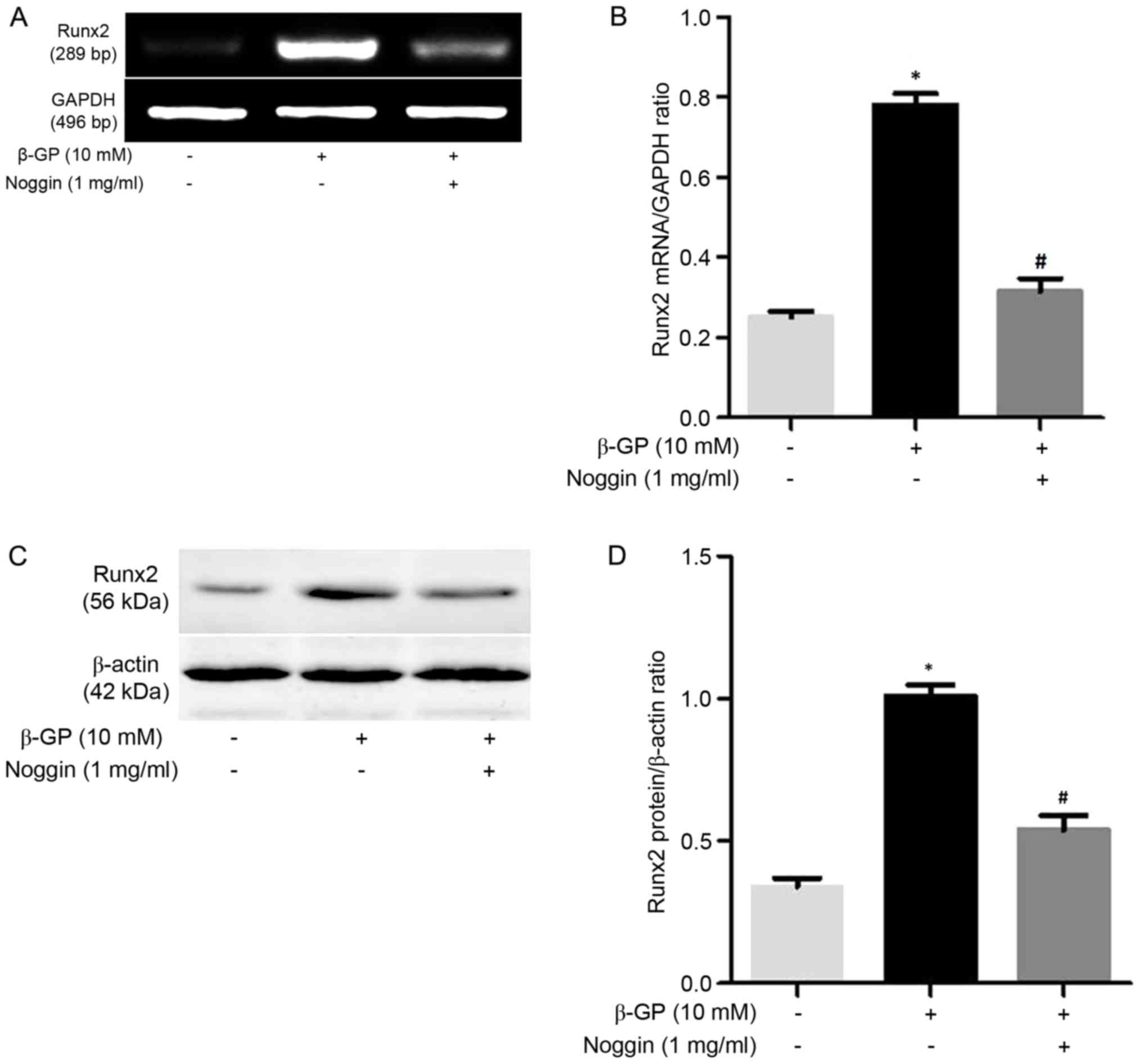
addition, VSMCs were also treated with β-GP in the presence or
absence of 1 mg/ml Noggin, which prevents BMP-2 binding to their
receptors. As expected, Runx2 expression was upregulated in VSMCs
maintained within β-GP medium compared to the control medium, while
the mRNA and protein expression of Runx2 was significantly
downregulated (P<0.05; Fig. 4) by
noggin. These results indicated that MK-4 partly functions in the
modulation of the BMP-2 signaling pathway to inhibit Runx2
expression of VSMCs induced by β-GP.
Discussion
VC is known to be a high risk factor for mortality
in CVD, particularly in patients with CKD (22,23).
Although numerous studies have demonstrated that a number of
factors are involved in regulating VC, an effective therapy has yet
to be found. Accumulated evidence manifested that VSMCs to
osteoblast-like cell transdifferentiation (VOT) is important in
promoting VC (10,24). Furthermore, the change in a variety
of key molecules of progression of VOT may promote the occurrence
and development of VC. Pharmacological manipulation of signaling
pathways or key molecules to inhibit VOT has been sufficient in the
treatment of VC (21). Furthermore,
a number of studies have indicated that MK-4 was associated with VC
(25–27). The present study investigated the
effects of MK-4 on the β-GP-induced calcification processes in rat
VSMCs. As predicted, it was identified that adding MK-4 to the
medium significantly inhibited calcification induced by β-GP.
Furthermore, the results confirmed that MK-4 is important in the
prevention of the β-GP-induced calcification of VSMCs, and may
prove useful in the treatment of VC.
However, the mechanisms by which MK-4 inhibited the
VC remains unclear. Previous studies have shown that VC is
cell-mediated and inactive process, which may result from the
osteogenic differentiation of VSMCs and subsequent extracellular
matrix mineralization (28–30). Runx2, which is a central
transcription factor, is necessary for osteogenic matrix gene
expression, bone formation, VSMC calcification and differentiation
(22,31,32).
Furthermore, the increasing phosphate concentration in vitro
results in upregulation of Runx2 expression and downregulation of
smooth muscle-specific gene expression (10). Furthermore, Runx2 has been shown to
be expressed in the calcified vascular lesion of CKD patients
(33). Thus, Runx2 has been regarded
as the earliest and most specific osteogenic marker of
differentiation for the promoting calcification. In the present
study, the expression of Runx2 mRNA and protein was evidently
upregulated following treatment with β-GP. However, the upregulated
expression of Runx2 mRNA and protein were inhibited in cells that
were co-cultured in a medium with different concentrations of MK-4,
with the effect remaining enhanced in a dose-dependent manner.
These results suggested that MK-4 partly reduced VC by reversing
the transdifferentiation of VSMCs in calcifying conditions in a
dose-dependent manner.
The BMPs are members of the transforming growth
factor-β superfamily, and they are important in VC and bone
formation (34,35). Additionally, the BMP subfamily can be
further subdivided into several subgroups, including BMP-2/4,
BMP-5/6/7/8, BMP-5/6/7 and BMP-9/10 (36–38). A
number of studies have shown that BMP signaling is a key regulator
of vascular disease (39–41). In order to confirm the effect of BMP
signaling in β-GP-induced osteochondrocytic phenotypic change, the
effect of Noggin on the Runx2 expression of β-GP-treated VSMCs was
investigated. It was shown that Noggin inhibited the β-GP-induced
Runx2 expression, suggesting that the BMP pathway is involved in
β-GP-induced osteochondrocytic differentiation of rat VSMCs. Next,
the present study examined whether the BMP pathway is regulated by
MK-4 in VSMCs transdifferentiation induced by β-GP. The results
revealed two things. Firstly, MK-4 inhibited the β-GP-induced
overexpression of BMP-2 and SMAD1, one of the receptor-regulated
SMADs (R-SMADs) that form heteromeric complexes with SMAD4 and then
these heteromeric R-SMAD/SMAD4 complexes translocate into the
nucleus. Secondly, it also increased the β-GP-regulated
down-expression of SMAD7, one of the inhibitory SMADs that
antagonize BMP receptor-initiated SMAD signaling by mediating the
degradation of receptors and R-SMADs, and suggesting that the
inhibitory effect of MK-4 on VSMCs transdifferentiation is
mediated, at least in part, through abrogating β-GP-induced
activation of the BMP-2 signaling pathway.
In conclusion, MK-4 was capable of validly reducing
calcification induced by β-GP in rat VSMCs. Furthermore, MK-4
inhibited the transdifferentiation of VSMCs into osteoblast-like
cells by suppressing the expression of Runx2 in a dose-dependent
manner. In addition, the downregulated expression levels of the
inhibitors BMP-2 and SMAD1 and upregulated expression of the
promoter SMAD7 caused by MK-4 in a dose-dependent manner was
observed. These observations reveal that MK-4 reduces
mineralization by the regulation of the signaling pathway of BMP-2
in order to attenuate the expression of Runx2. Furthermore, the
present study may help illuminate the function of MK-4 in the
calcification of VSMCs induced by β-GP.
Acknowledgements
Not applicable.
Funding
The present study was supported by the project of
the Hebei Natural Science Fund (grant no. H2012206157), the project
of the Hebei major medical Science (grant no. GL2011-51) and the
project of Hebei Science and Technology Planning (grant no.
16397733D)
Availability of data and materials
All data are fully available without
restriction.
Authors' contributions
LC, JX, and JZ designed the present study and wrote
the manuscript. MZ and SZ performed the statistical analysis. VSMC
cell culture, calcification assays, reverse
transcription-quantitative polymerase chain reaction and western
blotting were performed by SZ and YB.
Ethics approval and consent to
participate
The current study conformed to the Guide for Care
and Use of Laboratory Animals as adopted and promulgated by the
United National Institutes of Health. All experimental protocols
were approved by the Review Committee for the Use of Animal
Subjects of Hebei Medical University (Shijiazhuang, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declae that they have no competing
interests.
References
|
1
|
Tonelli M, Wiebe N, Culleton B, House A,
Rabbat C, Fok M, McAlister F and Garg AX: Chronic kidney disease
and mortality risk: A systematic review. J Am Soc Nephrol.
17:2034–2047. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tentori F, Blayney MJ, Albert JM,
Gillespie BW, Kerr PG, Bommer J, Young EW, Akizawa T, Akiba T,
Pisoni RL, et al: Mortality risk for dialysis patients with
different levels of serum calcium, phosphorus, and PTH: The
dialysis outcomes and practice patterns study (DOPPS). Am J Kidney
Dis. 52:519–530. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bolasco P: Effects of the use of
Non-calcium phosphate binders in the control and outcome of
vascular calcifications: A review of clinical trials on CKD
patients. Int J Nephrol. 2011:7584502011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Adragão T: Evaluation of vascular
calcifications in CKD patients. Int J Artif Organs. 32:81–86. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Giachelli CM: The emerging role of
phosphate in vascular calcification. Kidney Int. 75:890–897. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lau WL and Ix JH: Clinical detection, risk
factors, and cardiovascular consequences of medial arterial
calcification: A pattern of vascular injury associated with
aberrant mineral metabolism. Semin Nephrol. 33:93–105. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Olauson H and Larsson TE: FGF23 and Klotho
in chronic kidney disease. Curr Opin Nephrol Hypertens. 22:397–404.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li X, Yang HY and Giachelli CM: Role of
the sodium-dependent phosphate cotransporter, Pit-1, in vascular
smooth muscle cell calcification. Circ Res. 98:905–912. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Olszak IT, Poznansky MC, Evans RH, Olson
D, Kos C, Pollak MR, Brown EM and Scadden DT: Extracellular calcium
elicits a chemokinetic response from monocytes in vitro and in
vivo. J Clin Invest. 105:1299–1305. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Steitz SA, Speer MY, Curinga G, Yang HY,
Haynes P, Aebersold R, Schinke T, Karsenty G and Giachelli CM:
Smooth muscle cell phenotypic transition associated with
calcification: Upregulation of Cbfa1 and downregulation of smooth
muscle lineage markers. Circ Res. 89:1147–1154. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schurgers LJ, Uitto J and Reutelingsperger
CP: Vitamin K-dependent carboxylation of matrix Gla-protein: A
crucial switch to control ectopic mineralization. Trends Mol Med.
19:217–226. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wallin R, Schurgers LJ and Loeser RF:
Biosynthesis of the vitamin K-dependent matrix Gla protein (MGP) in
chondrocytes: A fetuin-MGP protein complex is assembled in vesicles
shed from normal but not from osteoarthritic chondrocytes.
Osteoarthritis Cartilage. 18:1096–1103. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cranenburg EC, VAN Spaendonck-Zwarts KY,
Bonafe L, Crettol Mittaz L, Rödiger LA, Dikkers FG, VAN Essen AJ,
Superti-Furga A, Alexandrakis E, Vermeer C, et al: Circulating
matrix γ-carboxyglutamate protein (MGP) species are refractory to
vitamin K treatment in a new case of Keutel syndrome. J Thromb
Haemost. 9:1225–1235. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Holden RM, Morton AR, Garland JS, Pavlov
A, Day AG and Booth SL: Vitamins K and D status in stages 3–5
chronic kidney disease. Clin J Am Soc Nephrol. 5:590–597. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Holden RM, Iliescu E, Morton AR and Booth
SL: Vitamin K status of Canadian peritoneal dialysis patients.
Perit Dial Int. 28:415–418. 2008.PubMed/NCBI
|
|
16
|
Westenfeld R, Krueger T, Schlieper G,
Cranenburg EC, Magdeleyns EJ, Heidenreich S, Holzmann S, Vermeer C,
Jahnen-Dechent W, Ketteler M, et al: Effect of vitamin K2
supplementation on functional vitamin K deficiency in hemodialysis
patients: A randomized trial. Am J Kidney Dis. 59:186–195. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Geleijnse JM, Vermeer C, Grobbee DE,
Schurgers LJ, Knapen MH, van der Meer IM, Hofman A and Witteman JC:
Dietary intake of menaquinone is associated with a reduced risk of
coronary heart disease: The Rotterdam Study. J Nutr. 134:3100–3105.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Koitaya N, Sekiguchi M, Tousen Y, Nishide
Y, Morita A, Yamauchi J, Gando Y, Miyachi M, Aoki M, Komatsu M, et
al: Low-dose vitamin K2 (MK-4) supplementation for 12 months
improves bone metabolism and prevents forearm bone loss in
postmenopausal Japanese women. J Bone Miner Metab. 32:142–150.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Campbell JH and Campbell GR: Culture
techniques and their applications to studies of vascular smooth
muscle. Clin Sci (Lond). 85:501–513. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chiba Y, Sakai H, Wachi H, Sugitani H,
Seyama Y and Misawa M: Upregulation of rhoA mRNA in bronchial
smooth muscle of antigen-induced airway hyperresponsive rats. J
Smooth Muscle Res. 39:221–228. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang S, Xu J, Feng Y, Zhang J, Cui L,
Zhang H and Bai Y: Extracellular acidosis suppresses calcification
of vascular smooth muscle cells by inhibiting calcium influx via
L-type calcium channels. Clin Exp Hypertens. 40:370–377. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu J, Bai Y, Jin J, Zhang J, Zhang S, Cui
L and Zhang H: Magnesium modulates the expression levels of
calcification-associated factors to inhibit calcification in a
time-dependent manner. Exp Ther Med. 9:1028–1034. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cherbuin N: Higher dietary intakes of
potassium, calcium and magnesium are associated with a reduced risk
of developing vascular dementia. Evid Based Ment Health. 16:262013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chaabane C, Heizmann CW and
Bochaton-Piallat ML: Extracellular S100A4 induces smooth muscle
cell phenotypic transition mediated by RAGE. Biochim Biophys Acta.
1853:2144–2157. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Spronk HM, Soute BA, Schurgers LJ,
Thijssen HH, De Mey JG and Vermeer C: Tissue-specific utilization
of menaquinone-4 results in the prevention of arterial
calcification in warfarin-treated rats. J Vasc Res. 40:531–537.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wahlqvist ML, Tanaka K and Tzeng BH:
Clinical decision-making for vitamin K-1 and K-2 deficiency and
coronary artery calcification with warfarin therapy: Are diet,
factor Xa inhibitors or both the answer? Asia Pac J Clin Nutr.
22:492–496. 2013.PubMed/NCBI
|
|
27
|
Koitaya N, Ezaki J, Nishimuta M, Yamauchi
J, Hashizume E, Morishita K, Miyachi M, Sasaki S and Ishimi Y:
Effect of low dose vitamin K2 (MK-4) supplementation on bio-indices
in postmenopausal Japanese women. J Nutr Sci Vitaminol (Tokyo).
55:15–21. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kerage D, Brindley DN and Hemmings DG:
Review: Novel insights into the regulation of vascular tone by
sphingosine 1-phosphate. Placenta. 35 Suppl:S86–S92. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang W, Li C, Pang L, Shi C, Guo F, Chen
A, Cao X and Wan M: Mesenchymal stem cells recruited by active TGFβ
contribute to osteogenic vascular calcification. Stem Cells Dev.
23:1392–1404. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Danilevicius CF, Lopes JB and Pereira RM:
Bone metabolism and vascular calcification. Braz J Med Biol Res.
40:435–442. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin ME, Chen T, Leaf EM, Speer MY and
Giachelli CM: Runx2 expression in smooth muscle cells is required
for arterial medial calcification in mice. Am J Pathol.
185:1958–1969. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li M, Wu P, Shao J, Ke Z, Li D and Wu J:
Losartan inhibits vascular calcification by suppressing the BMP2
and Runx2 expression in rats in vivo. Cardiovasc Toxicol.
16:172–181. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bai Y, Zhang J, Xu J, Cui L, Zhang H and
Zhang S: Alteration of type I collagen in the radial artery of
patients with end-stage renal disease. Am J Med Sci. 349:292–297.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lieberman JR, Daluiski A and Einhorn TA:
The role of growth factors in the repair of bone. Biology and
clinical applications. J Bone Joint Surg Am. 84-A:1032–1044. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sun SX, Guo HH, Zhang J, Yu B, Sun KN and
Jin QH: BMP-2 and titanium particles synergistically activate
osteoclast formation. Braz J Med Biol Res. 47:461–469. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Miyazono K, Kamiya Y and Morikawa M: Bone
morphogenetic protein receptors and signal transduction. J Biochem.
147:35–51. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kawabata M, Imamura T and Miyazono K:
Signal transduction by bone morphogenetic proteins. Cytokine Growth
Factor Rev. 9:49–61. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sato Simoes AY, Bub GL and Campos AH:
BMP-2 and −4 produced by vascular smooth muscle cells from
atherosclerotic lesions induce monocyte chemotaxis through direct
BMPRII activation. Atherosclerosis. 235:45–55. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yao Y, Bennett BJ, Wang X, Rosenfeld ME,
Giachelli C, Lusis AJ and Boström KI: Inhibition of bone
morphogenetic proteins protects against atherosclerosis and
vascular calcification. Circ Res. 107:485–494. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cai J, Pardali E, Sánchez-Duffhues G and
Ten Dijke P: BMP signaling in vascular diseases. FEBS Lett.
586:1993–2002. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shao JS, Aly ZA, Lai CF, Cheng SL, Cai J,
Huang E, Behrmann A and Towler DA: Vascular Bmp Msx2 Wnt signaling
and oxidative stress in arterial calcification. Ann N Y Acad Sci.
1117:40–50. 2007. View Article : Google Scholar : PubMed/NCBI
|