Introduction
Atherosclerosis is a main pathological basis of
cardiovascular and cerebrovascular disease, such as coronary heart
disease, ischemic cerebrovascular disease and thromboembolic
disease (1). Atherosclerosis is a
large and middle artery disease, a major cause of heart disease and
stroke, and a chronic inflammatory disease induced by large amounts
of lipid and cholesterol accumulated in the large and medium-sized
arteries, and accompanied by fibrous plaque formation (2). Epidemiological studies (3,4) have
demonstrated that certain environmental and genetic factors are
associated with the occurrence and development of atherosclerosis.
The main genetic factors include high density lipoprotein
reduction, increased low density lipoprotein and very low density
lipoprotein, elevated blood pressure and elevated blood sugar
(5). In addition, there are certain
major environmental factors, including a high-fat diet, smoking,
lack of exercise, antioxidant levels and infectious disease
(2).
Vascular endothelial cells not only constitute a
semi-selective permeability barrier between blood and tissue fluid,
but also synthesize and secrete a variety of bioactive substances.
Their structural integrity and barrier function have a central role
in maintaining cardiovascular homeostasis (6). It has been widely recognized that
damaged blood vessel endothelium leads to endothelial dysfunction,
further expression and release of various bioactive substances and
adhesion molecules (2). These
changes ultimately lead to the formation of atherosclerosis, which
is a chronic inflammatory process (1). Vascular endothelial dysfunction is an
early manifestation of atherosclerosis (7,8),
promoting lipid deposits, monocytes under the endothelium and the
formation of foam cells.
The maintenance of the normal barrier function of
vascular endothelium is associated with the dynamic balance between
tight junctions (TJs), adherence junctions and cytoskeleton
contraction (6). Cell contraction is
a common way to increase barrier permeability. Cell contraction is
mainly affected by actin and myosin, and endothelial permeability
is increased when actin and myosin produce a greater contraction
force (6). Myosin II includes a
heavy chain and a light chain, and myosin light chain (MLC) is
divided into the basic light chain and regulatory light chain. It
is the regulatory light chain that serves a regulatory role in
myosin activity (9). MLC kinase
(MLCK), dependent on Ca2+/calmodulin (CaM) binding
protein kinase, is a member of the immunoglobulin superfamily and
is activated by the regulatory light chain in MLC. When activated
by certain factors, MLCK could induce phosphorylation of MLC and
vascular endothelial cell cytoskeleton rearrangement (10). This induces the following effects:
Concentric contraction of endothelial cells, the destruction of TJs
between cells and vascular barrier dysfunction (11). Finally, the endothelial permeability
increases, a large number of lipid components infiltrate and
deposit under the endothelium which then affects vascular
endothelial barrier function (12).
Glycyrrhiza uralensis Fisch is mainly from
the West, including leguminous plants Glycyrrhiza uralensis,
Ural Glycyrrhiza uralensis, Glycyrrhiza glabra and dry root and
rhizome (13). Glabridin is
extracted from Guangguo licorice, insoluble in water and has
antioxidant, anti-inflammatory (14)
and anti-atherosclerosis (15)
properties, which provide rationale for its use as a treatment of
cardiovascular disease.
In the present study, the association between the
effect of Glabridin on atherosclerosis, and the mitogen-activated
protein kinase (MAPK) signaling pathway and myosin light chain
phosphatase-dependent MLC phosphorylation was investigated. The
experiment was conducted by establishing an animal model of
atherosclerosis, and analyzing blood lipid changes, and observing
changes in vascular morphology, MLCK expression, and intimal
permeability.
Materials and methods
Reagents
Glabridin was purchased from DC Chemicals (Shanghai,
China). Cholesterol was obtained from China Pharmaceutical (Group)
Shanghai Chemical Reagent Company (Shanghai, China). Normal rabbit
feed was obtained from Experimental Animal Center of Anhui Medical
University (Hefei, China). The Triglyceride assay kit (cat. no.
10018) and the Total Cholesterol assay kit (cat. no. 10028) were
purchased from Zhejiang Dongou Biochemical Co., Ltd. (Zhejiang,
China).
Animals and groups
A total of 18 3-month-old, male New Zealand white
rabbits (2.4±0.5 kg) were purchased from Shandong Qingdao Kang
group (Qingdao, China). Rabbits were housed at room temperature
with a humidity of 50–60% and were subjected to a 12 h light/dark
cycle, with free access to food and water. The rabbits were
randomly divided into 3 groups and fed as follows: Control group
(n=6), basic diet for 12 weeks; HF group (n=6), high fat diet for
12 weeks; and Glabridin treatment group (n=6), high fat diet with 2
mg/kg/day Glabridin via oral catheter from weeks 6–12. The
Glabridin dose was selected on the basis of a preliminary
experiment according to a previous study (16). High fat diet consisted of 1%
cholesterol and 5% lard prepared in rabbit food. The cholesterol
concentration was determined in accordance with previous literature
(17).
For the determination of total cholesterol (TCH) and
triglyceride (TG), the rabbits were fasted for 1 day. The rabbits
were then anesthetized with 3% sodium pentobarbital (30 mg/kg;
Sigma-Aldrich; Merck KgaA, Darmstadt, Germany), and GES6 color
ultrasonic diagnostic apparatus was used to observe the abdominal
aorta. Blood samples were taken from the abdominal aorta and
following standing for 2 h at 37°C, the blood was centrifuged for
10 min at room temperature at 1,006 × g. The supernatant was
removed and stored at −80°C.
A previous study was referenced to understand the
structure of rabbit aortas (18).
Following sacrifice, rabbit aortas were removed and divided into
three parts. Each part had the same number of rabbit aortas from
the control, treatment and high fat group. For part one, one
abdominal aorta was randomly selected from each of the three
groups. These aortas were fixed in 10% formalin at room temperature
for 8 h, and embedded in paraffin for the immunohistochemical and
morphological analysis. Another part was embedded in optimum
cutting temperature compound (OCT), immediately at −80°C prior to
freezing, preparing for permeability analysis and oil red O
staining. The third part of abdominal aortas was frozen immediately
at −80°C for western blot analysis. All experiments were approved
by the Ethics Committee of Anhui Medical University.
Detection of serum lipids
The enzyme coupling colorimetric method was utilized
to detect the TCH and TG in model rabbits. All protocols were
performed in strict accordance with the assay kits and ELISA
instructions.
Endothelial function assay
The endothelial function assay was performed as
described previously (17). Rabbits
were anesthetized using 3% sodium pentobarbital (30 mg/kg), and the
abdomens were shaved. The abdominal aorta was observed using a GES6
color ultrasonic diagnostic apparatus and 13-Mhz ultrasound probe.
The probe was placed 0.5–1.0 cm below the renal artery to obtain a
longitudinal axis view of the abdominal aorta. The basal diastolic
diameter (D0) of the abdominal aorta was measured
initially, followed by the maximum diameter of the end diastolic
(DX) of the abdominal aorta after the left hind limb was
compressed for 5 min. During the process, the probe position was
maintained. Maximum diastolic rate of abdominal aorta in each
rabbit (%) was calculated as follows:
(DX-D0)/D0 ×100%.
Permeability assay
The permeability assay using the surface
biotinylation technique was performed as described by Wang et
al (19) with modifications
specifically for the aortic intima. The abdominal aorta canals were
filled with a Sulfo-NHS-LC-Biotin solution (Pierce; Thermo Fisher
Scientific, Inc.) in HBS at 1 mg/ml for 30 min at room temperature.
Aortas were then rinsed with PBS, embedded in OCT, and
cryosectioned. Subsequently, the 6-µm frozen sections were
incubated in 5% skimmed milk powder at 4°C overnight. Following
washing 3 times with PBS, the sections were incubated at 4°C
overnight with Rhodamine 600 Avidin D (XRITC-avidin; 1:200; cat.
no. A-2005; Shanghai Haoran Biological Technology Co., Ltd.,
Shanghai, China). The samples were then examined by fluorescence
microscopy (magnification, ×400). Images were captured, and the
JD-801 Pathological Image Analysis system 4.0 (Jiangsu JEDA
Science-Technology Development Co., Ltd., Jiangsu, China) was used
to measure the integral optical density values.
Histological examination
Abdominal aorta specimens were dehydrated and
embedded in paraffin, sectioned (4 µm) and stained with hematoxylin
and eosin as previously reported (20). Extracellular matrix deposition was
assessed using Masson trichrome staining, via the sequential
addition of Bouin (at 25°C for 24 h), Weigert (at 25°C for 5 min),
and Biebrich (at 25°C for 5 min) solutions (OriGene Technologies,
Inc., Rockville, MD, USA) to sections, in accordance with the
manufacturer's protocol provided by the supplier of Bouin, Weigert
and Biebrich solutions.
Lipid and fat content was evaluated by oil red O
staining. Frozen sections were fixed with 10% formalin for 30 min
at room temperature. Following rinsing with 60% isopropyl alcohol
for 5 min, filtered oil red O working solution was used to stain
the aorta sections (4 µm) for 5 min at room temperature. Sections
were rinsed again with distilled water to reduce the background,
and imaged using light microscopy (magnification, ×100). Images
were captured, and the JD-801 Pathological Image Analysis system
was used to measure the integral optical density values. Image-Pro
Plus software 6.0 (Media Cybernetics, Inc., Rockville, MD, USA) was
used to measure the area ratio. The area ratio (%) was calculated
as follows: Plaque area/total arterial area ×100%.
Immunohistochemistry analysis
The abdominal aorta tissues sections were
deparaffinized, rehydrated, then fixed with a methanol −0.3%
H2O2 solution at room temperature for 20 min.
Following high compression heating at 60°C, the abdominal aorta was
immersed in xylene (I) for 5 min, xylene (II) for 5 min, 100%
ethanol for 5 min, 95% ethanol for 2 min, 90% ethanol for 2 min,
85% ethanol for 2 min, 80% ethanol for 2 min, soaked in 75% ethanol
for 2 min and washed using distilled water. Antigens were unmasked
by cooling for 30 min at room temperature. Sections (4 µm) were
then blocked using PBS containing 0.05% Tween-20 and 1% BSA
(Sigma-Aldrich; Merck KGaA) at room temperature for 30 min. Primary
antibodies against MLCK (1:1,000; M7905; Sigma-Aldrich; Merck KGaA)
were incubated with the tissue sections at 4°C overnight;
subsequently, sequential incubation was performed with biotinylated
secondary antibodies (1:1,000; cat. no. 211-032-171; AmyJet
Scientific Inc., Wuhan, China) and horseradish streptavidin
(Cusabio Technology LLC, Houston, TX, USA) for 30 min at 37°C.
Finally, the samples were incubated with diaminobenzidine together
at room temperature for 2 min, and hematoxylin was used to
counterstain the nuclei. Positive stained cells appeared brown in
color. Images were captured using light microscopy (magnification,
×400), and the JD-801 Pathological Image Analysis system was used
to measure the integral optical density values.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the aorta using the
Trizol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). mRNA
was then reverse transcribed into cDNA using the RT-qPCR reverse
transcription kit (Takara Biotechnology Co., Ltd., Dalian, China)
in accordance with the manufacturer's protocol. RNA expression was
determined using RT-qPCR with the following primers: MLCK forward,
5′-TCTTGGGAGAGGATGACGAA-3′ and reverse, 5′-GTGTGGCAGAGGGTGAGAAG-3′;
GADPH forward, 5′-TCACCACCTTCTTGATGTCG-3′ and reverse,
5′-TGAACGGGAAACTCACTGG-3′. The thermocycling conditions were as
follows: Initial denaturation at 95°C for 15 min; 40 cycles of
denaturation at 95°C for 10 sec, annealing at 60°C for 30 sec and
extention at 72°C for 30 sec. The reaction mixture contained the
following materials: 2X Power SYBR Green Master Mix (Applied
Biosystems; Thermo Fisher Scientific, Inc.), cDNA template, 10
pmol/µl of each primer and sterile water. Using GraphPad Prism
software version 5.01 (GraphPad Software, Inc., La Jolla, CA, USA),
relative gene expression was quantified according to the
comparative Cq method (21). With
respect to the expression levels of GAPDH, all results were
normalized. A reverse transcription kit was purchased from, and
primers of MLCK and GAPDH were obtained from Shanghai Shenggong
Biology Engineering Technology Service, Ltd. (Shanghai, China).
Western blotting
The levels of MLCK, phosphorylated (p)-c-Jun
N-terminal kinase (JNK), p-extracellular signal regulated kinase
(ERK) and p-p38 were measured by western blotting. The following
antibodies ERK (cat. no. sc-135900), p-38 (cat. no. sc-7149), JNK
(cat. no. sc-7345), MLCK (cat. no. sc-365352), p-JNK (cat. no.
sc-6254), p-ERK (cat. no. sc-7976), β-actin (cat. no. sc-47778) and
p-p38 (cat. no. sc-7975-R) were purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). All were used at a dilution
of 1:1,000. The experimental procedures were described in a
previous study (22). Levels of
β-actin were used to normalize the relative protein expression
levels. Experiments were repeated three times independently.
Western blotting images were analyzed using the quantity one image
processing system 4.62 (Bio-Rad Laboratories, Hercules, CA,
USA).
Statistical analysis
All statistical analyses were carried out using SPSS
software, version 16.0 (SPSS, Inc., Chicago, IL, USA). All data are
expressed as the mean ± standard deviations. Comparisons between
groups were carried out using one-way analysis of variance followed
by Student-Newman-Keuls method. P<0.05 was considered to
indicate a statistically significant difference.
Results
Blood lipid analysis
Following 12 weeks of treatment, the TCH and TG in
the high fat model group were significantly higher than those in
the normal control group. Compared with the high-fat model group,
Glabridin significantly reduced the serum TCH level, and the level
of TG was markedly reduced (Table
I).
 | Table I.The effect of high fat diet and
Glabridin on TCH and TG of atherosclerosis model rabbits. |
Table I.
The effect of high fat diet and
Glabridin on TCH and TG of atherosclerosis model rabbits.
| Level | Control | High fat diet | Glabridin |
|---|
| TCH (mmol/l) | 1.03±0.38 |
68.56±4.78a |
52.90±9.3b |
| TG (mmol/l) | 0.40±0.21 |
9.16±2.57a | 6.45±2.68 |
Morphological characteristics
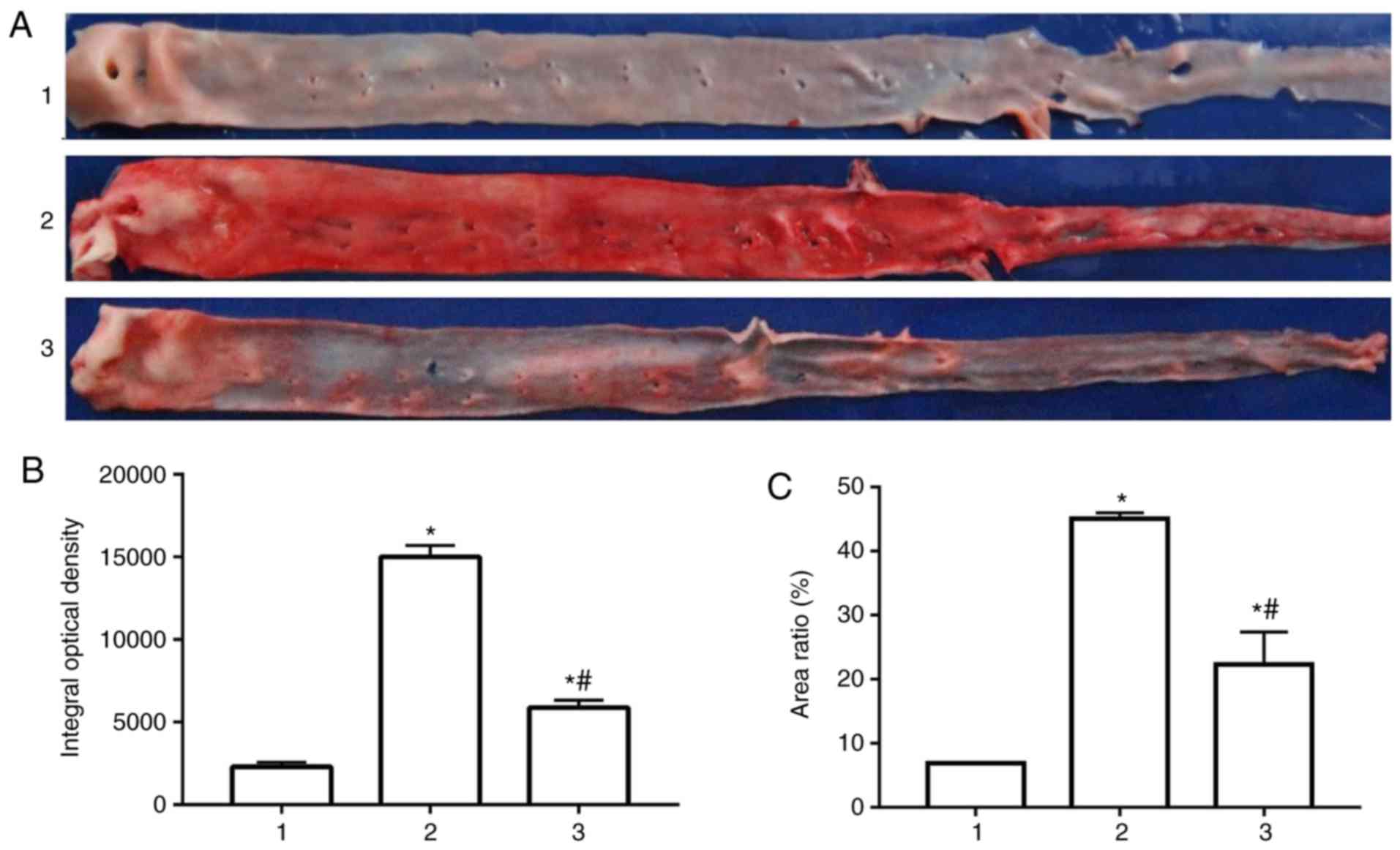
Following 12 weeks of high-fat diet intervention,
the rabbit abdominal aorta was observed by oil red O staining. As
presented in Fig. 1A,
atherosclerotic plaques were observed in the rabbit aorta of the
high fat model group and Glabridin group in comparison with the
normal control group. Plaques of the rabbit aorta in the high fat
model group were more obvious than in the normal control group and
Glabridin treatment group. Compared with the high-fat model group,
the plaques of the high-fat group were decreased following
treatment with Glabridin. The results of the integrated optical
density and plaque area ratio also illustrate these findings
(Fig. 1B and C).
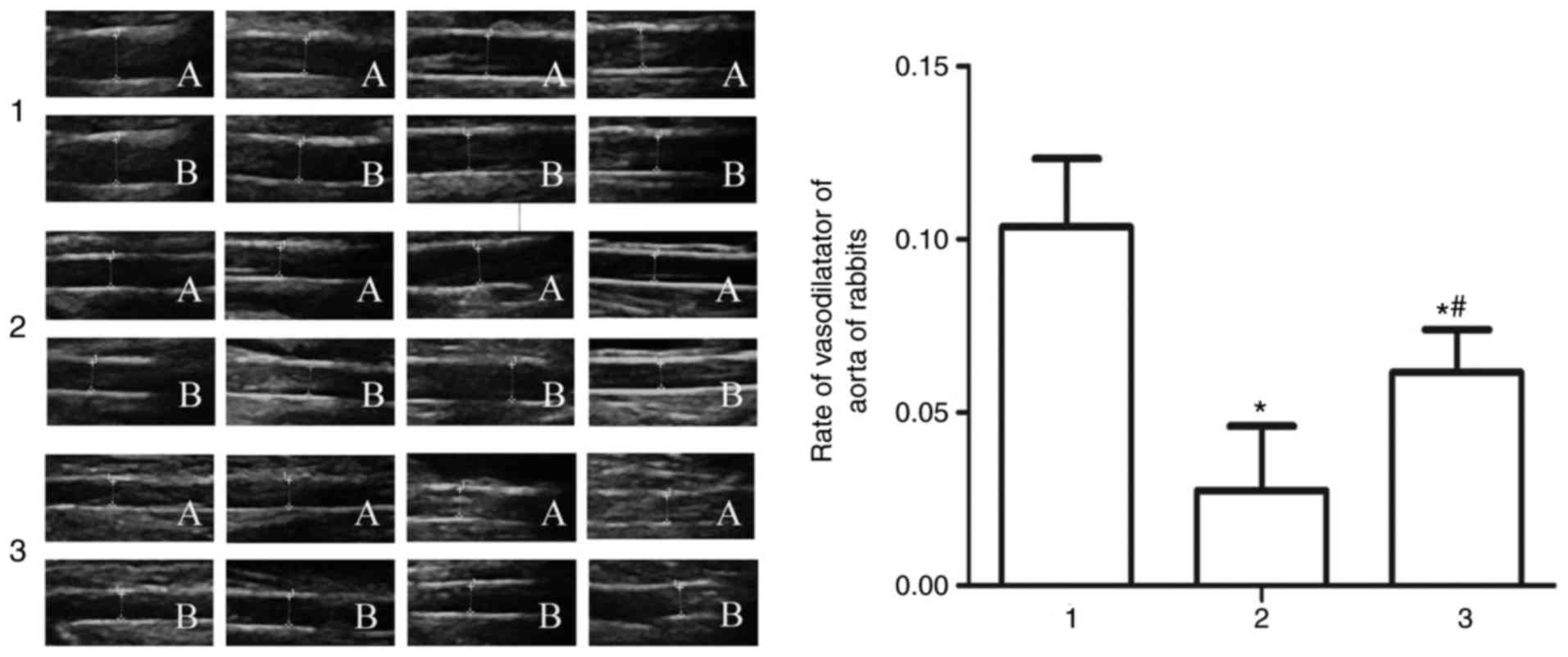
Endothelial function
The effects of Glabridin on vascular endothelial
function were evaluated. Following pressurization for 5 min, aortic
vasodilatation function in the high-fat model group was poorer than
the normal control group. The results were as follows: Normal
group. The fundamental maximum diameters of the abdominal aortas
were 2.65, 2.25, 2.42 and 2.25 mm; the maximum diameter of the
abdominal aortas following pressuring left hind limbs for 5 min
were 2.90, 2.43, 2.71 and 2.51 mm, which were increased by 9.43,
8.00, 11.98 and 11.56%, respectively, compared with the fundamental
maximum diameter. Model group. The fundamental maximum diameters of
the abdominal aortas were 2.07, 2.06, 2.59 and 1.81 mm; the maximum
diameter of the abdominal aortas following pressuring left hind
limbs for 5 min were 2.11, 2.10, 2.63 and 1.91 mm, which were
increased by 1.93, 1.94, 1.54 and 5.52%, respectively, compared
with the fundamental maximum diameter. Glabridin group. The
fundamental maximum diameters of the abdominal aortas were 1.77,
1.96, 1.92 and 1.96 mm; the maximum diameter of the abdominal
aortas following pressuring left hind limbs for 5 min were 1.86,
2.10, 2.06 and 2.06 mm, which were increased by 9.00, 7.14, 7.29
and 5.10%, respectively, compared with the fundamental maximum
diameter. Compared with the high fat model group, the aortic
vasodilatation rate was significantly increased in the Glabridin
group (Fig. 2).
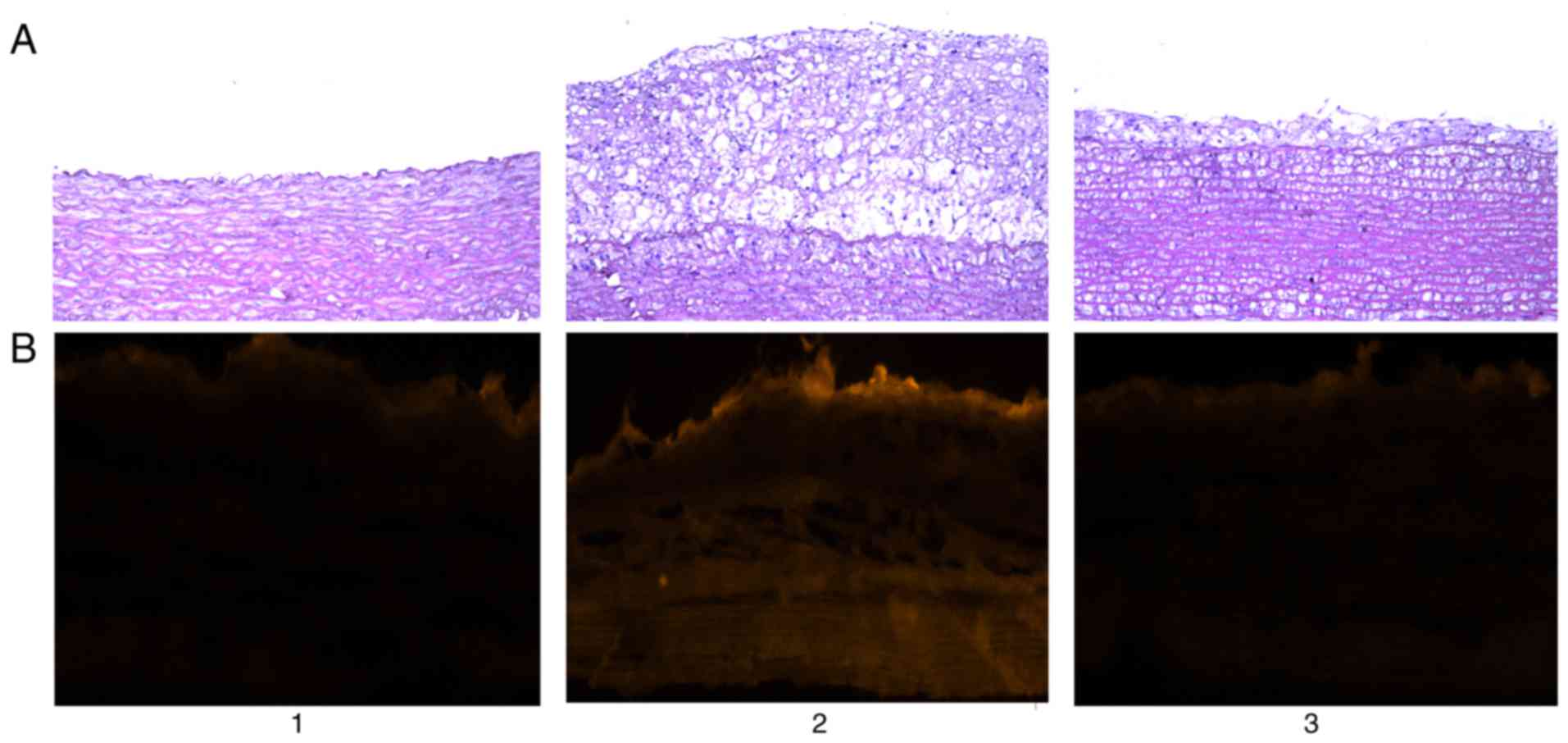
Endometrial permeability
In the normal control group, the structure of aortic
intima was intact, and the endothelial cells were closely connected
and arranged neatly. Compared with the normal control group, a
large number of foam cells were observed in the aorta intima in the
high-fat model group, and the aortic intima markedly thickened.
However, in the Glabridin treatment group, both foam cells and
aortic intima were markedly ameliorated. As presented in Fig. 3, fluorescence microscopy indicated
that in the normal control group there was no fluorescent dye
penetration, and in the high fat model group rabbit aortic
permeability increased markedly, with fluorescent dye penetration
throughout almost all layers of the wall. Following administration
of Glabridin, a degree of permeability remained. However, the
degree of penetration of fluorescent dyes was decreased in
comparison with the high fat model group (Fig. 3).
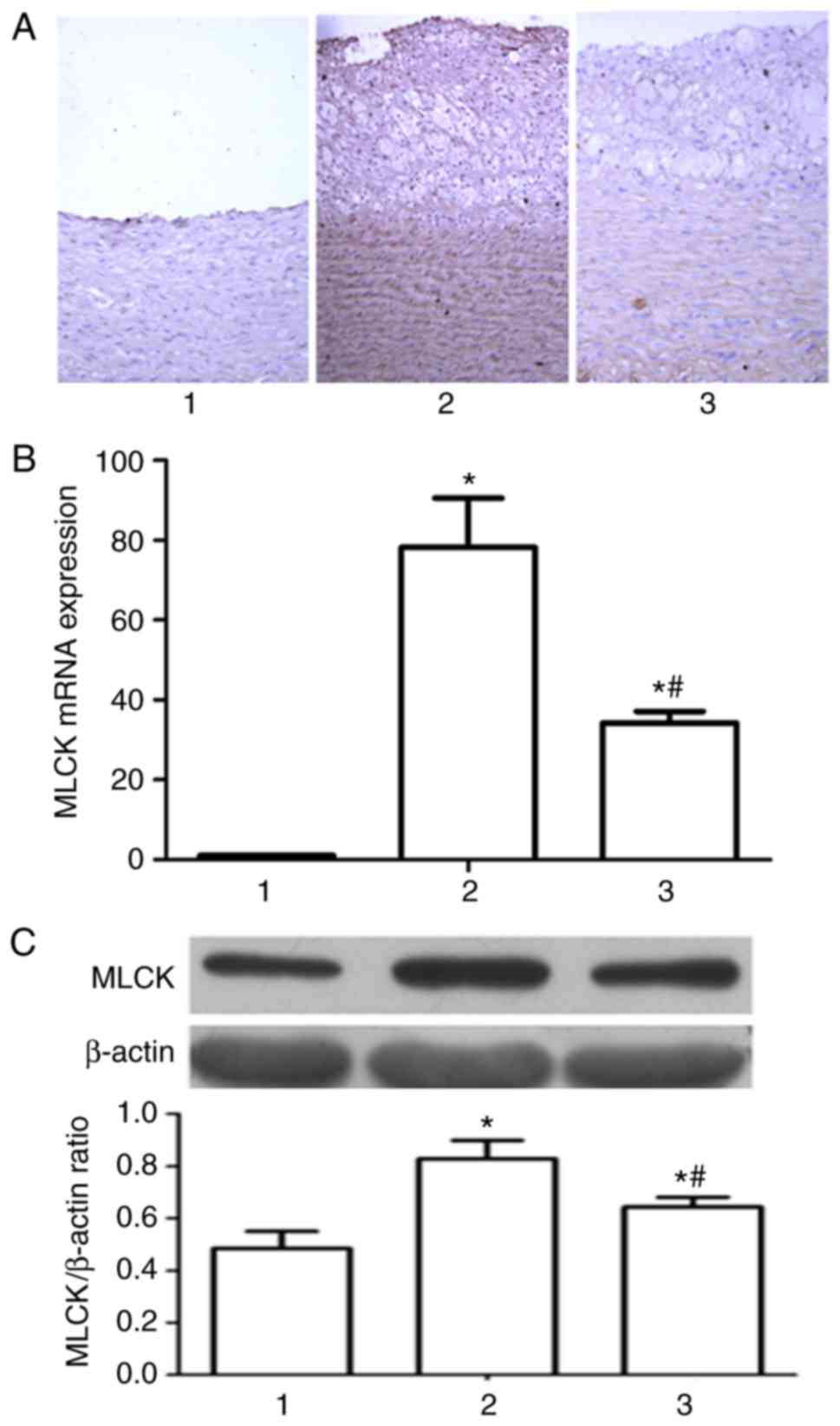
Measurement of relative MLCK
expression levels
As observed by immunohistochemical methods (Fig. 4A), following 12 weeks of diet
intervention, high MLCK expression levels were observed in the
arterial wall in the high fat model group, in the aortic
intima-medias and smooth muscle layers, with a marked increase in
comparison with the normal control group. In the Glabridin
treatment group, this expression was markedly reduced, compared
with the high fat diet group. The transcription level of MLCK was
analyzed by RT-qPCR through. As presented in Fig. 4B, Glabridin significantly
downregulated the transcription level of MLCK mRNA in comparison
with the high fat diet groups, which was in accordance with the
results of western blotting. Western blotting (Fig. 4C) demonstrated that following 12
weeks of high-fat diet intervention, compared with the normal
control group, the expression level of MLCK protein increased,
which was then downregulated by Glabridin.
Measurement of reactive signal pathway
activity level
Western blotting revealed that the levels of
phosphorylation of JNK, ERK and p38 were significantly enhanced in
the high fat group, compared with the normal control group.
However, the phosphorylation level of JNK, ERK, p38 in MAPK
signaling pathway was significantly downregulated by Glabridin, in
comparison with the high fat diet group (Fig. 5).
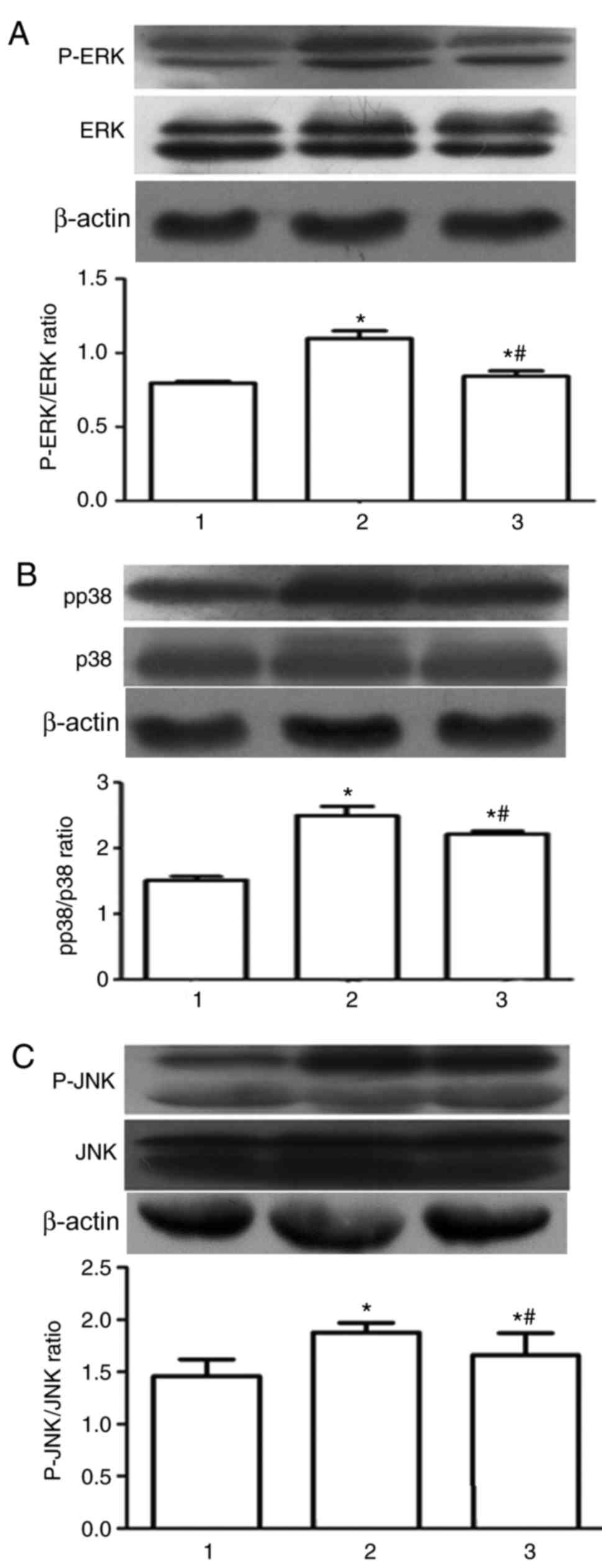 | Figure 5.The effects of Glabridin on the
activation of MAPK Signaling pathway. Phosphorylation of (A) ERK,
(B) p38 and (C) JNK indicated by western blot images and bar graphs
of protein quantification. *P<0.05 vs. 1; #P<0.05
vs. 2. 1, normal group; 2, model group; 3, Glabridin group; MLCK,
myosin light chain kinase; MAPK, mitogen-activated protein kinase;
MLC, myosin light chain; ERK, extracellular signal-regulated
kinase; JNK, c-Jun N-terminal kinase; p, phosphorylated. |
Discussion
Up to now, the study of atherosclerosis is not
comprehensive. The changes in the different stages of
atherosclerosis can be regarded as different stages of chronic
inflammation. Various cellular responses are similar to the
inflammatory response, which serves as the body's response to
injury (23,24). The dysfunction of endothelial
structure is the initial stage of atherosclerosis, followed by a
series of inflammatory reactions, a large number of inflammatory
factors released and promoting the further development of
atherosclerosis (25). Endothelial
cells serve an important role in regulating cardiovascular
functions and systemic homeostasis and in regulating
pathophysiological processes such as inflammation and immunity
(26). Clinically, there is no
effective drug for prevention and treatment.
New Zealand rabbits were used to establish an
atherosclerosis model as the rabbit diet is sensitive to high
cholesterol, and they typically have low levels of cholesterol in
their blood plasma (27), which
significantly increases following the administration of a
high-cholesterol diet. Food-induced lesions are located in the
aorta, and inflammatory cell infiltration, increased permeability
of intima and a large number of macrophages differentiating into
foam cells, are similar to those observed in humans (28).
The rabbit models of atherosclerosis were
established through diet intervention for 12 weeks. The TCH and TG
in the high fat model group were significantly higher than those in
the normal control group, compared with the high-fat model group.
Glabridin significantly reduced the serum TCH level and markedly
reduced the level of TG.
Following diet intervention for 12 weeks, the
expression of MLCK in the high fat group was significantly higher
than the normal control group. Following Glabridin treatment, this
was ameliorated, suggesting that Glabridin served a role in
downregulating the expression of MLCK; as observed by
immunohistochemistry and western blotting. The decrease of MLCK
expression contributed to inhibiting the phosphorylation level of
MLC, ultimately downregulating the vascular permeability. Increased
vascular permeability is also an important factor in promoting
atherosclerosis (29). In the
present study, the permeability of arterial intima was detected by
macromolecular fluorescent dye penetration. It was demonstrated
that there was almost no osmotic dye in the aorta of rabbits in the
normal group. Vascular permeability was increased in the high-fat
group, and the dye almost immersed into the whole layer. However,
this phenomenon was markedly ameliorated by Glabridin, which
indicated that Glabridin could ameliorate vascular permeability.
Normal endothelial cells express a certain amount of MLCK, and the
level of MLCK expression was increased following 12 weeks of
high-fat diet intervention. The expression of MLCK promoted the
phosphorylation of MLC, inducing vascular endothelial cell
cytoskeleton rearrangement, resulting in vascular endothelial cells
in concentric contraction, TJs between the endothelial cells were
damaged, cell gaps increased, and finally, the permeability of
endothelial cells increased. Increased endothelial cell
permeability promotes lipid, monocytes and other depositions in the
subcutaneous tissue, and contributes to the formation of foam cells
and atherosclerotic plaque, leading to decreased vascular
elasticity and diastolic function. The results of the abdominal
aorta diastolic function measured by percutaneous ultrasonography
were also consistent with the above results.
Considerable evidence has suggested that MAPK is
associated with atherosclerosis (30–32).
MAPK belongs is a serine/threonine protein kinase, and there are 4
MAPK pathways known at present, namely, ERK1,2;
JNK/stress-activated protein kinase; p38 MAPK; and big mitogen
activated protein kinase, in which ERK/MAPK is an important
pathway. The four pathways activate the cascade reaction, that is,
the kinase of MAPK kinase to the MAPK kinase to MAPK (30). The MAPK signaling pathway includes
the activation of p38/MAPK, ERK/MAPK and JNK/MAPK, and is
associated with the induction of inflammatory factors (33). The present study suggested that the
phosphorylation levels of p38, ERK and JNK were increased in the
high fat group, but decreased by Glabridin. This suggests that
Glabridin may inhibit formation of foam cells via downregulating
the phosphorylation of p38, ERK and JNK. This indicated that
protein kinase inhibitors could be used as potential therapeutic
agents for the prevention and treatment of vascular barrier
dysfunction and endothelial permeability.
MAP kinase (ERK1 and ERK2) affects movement
mechanism of cells through phosphorylation, and enhances MLCK
activity, leading to MLC phosphorylation, cell junction disruption
and cell gap formation (34).
Inhibition of MAPK activity results in decreased MLCK function,
decreased MLC phosphorylation and decreased cell migration in the
extracellular matrix. However, increased activity of MAPK kinase
leads to activation of MAPK, resulting in phosphorylation of MLCK
and MLC, and enhanced cell migration (35). MAPK is able to phosphorylate MLCK
directly, increasing its ability to phosphorylate MLC, which
promotes cytoskeleton contraction necessary for cell movement
(35,36). MLCK, encoded by the MLCK 1–3 gene
located on the human chromosome 3,20,16 (12), contains multiple phosphorylation
sites of MAP kinase. Previous studies have demonstrated that MLCK,
a key regulator of cell movement and contraction, is a MAPK
substrate (34,37). Notably, ERK1 and ERK2 can directly
phosphorylate MLCK in vitro, leading to enhanced
phosphorylation of MLC (34). This
phosphorylation was associated with increased sensitivity to
Ca2+/CaM. In addition to the MAPK signaling pathway, MLC
serves an important role in myosin function through
Ca2+/phospholipid dependent protein kinase II, protein
kinase C and cAMP dependent protein kinase (38,39).
In conclusion, the present study demonstrated that
Glabridin has effects on the regulation of lipid metabolism
disorder, endothelial dysfunction and increased permeability. By
downregulating the phosphorylation levels of p38, ERK and JNK,
Glabridin inhibits the expression of MLCK and the activity of MLC,
MLCK and the formation of foam cells. Therefore, Glabridin may be a
potential treatment for atherosclerosis and serve a specific role
in the prevention and treatment of increased vascular
permeability.
Acknowledgements
Not applicable.
Funding
The present study was supported by National Natural
Science Foundation of China (grant nos. 81570419 and 81270372).
Availability of data and materials
The datasets used and/or analyzed during the present
study may be obtained at the reasonable request of the respective
authors without prejudice to the confidentiality of the
participants.
Authors' contributions
HZ designed and organized the study. GW and GS
performed the permeability assay, histological examination,
immunohistochemistry analysis, reverse transcription-quantitative
polymerase chain reaction and Western blotting. YW and PY detected
serum lipids. XW assayed endothelial function. BZ analyzed the data
and wrote the manuscript.
Ethics approval and consent to
participate
The present protocols were approved by the Ethics
Committee of Anhui Medical University (Hefei, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hansson GK: Inflammation, atherosclerosis,
and coronary artery disease. N Engl J Med. 352:1685–1695. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lusis AJ: Atherosclerosis. Nature.
407:233–241. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kronenberg F, Kronenberg MF, Kiechl S,
Trenkwalder E, Santer P, Oberhollenzer F, Egger G, Utermann G and
Willeit J: Role of lipoprotein(a) and apolipoprotein(a) phenotype
in atherogenesis. Circulation. 100:1154–1160. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Assmann G, Cullen P, Jossa F, Lewis B and
Mancini M: Coronary heart disease: Reducing the risk: The
scientific background to primary and secondary prevention of
coronary heart disease. A worldwide view. International Task force
for the Prevention of Coronary Heart disease. Arterioscl Thromb
Vasc Biol. 19:1819–1824. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lusis AJ, Mar R and Pajukanta P: Genetics
of atherosclerosis. Annu Rev Genomics Hum Genet. 5:189–218. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Williams IL, Wheatcroft SB, Shah AM and
Kearney MT: Obesity, atherosclerosis and the vascular endothelium:
Mechanisms of reduced nitric oxide bioavailability in obese humans.
Int J Obes Relat Metab Disord. 26:754–764. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Daiber A, Steven S, Weber A, Shuvaev VV,
Muzykantov VR, Laher I, Li H, Lamas S and Münzel T: Targeting
vascular (endothelial) dysfunction. Br J Pharmacol. 174:1591–1619.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hamlat-Khennaf N, Neggazi S, Ayari H,
Feugier P, Bricca G, Aouichat-Bouguerra S and Beylot M:
Inflammation in the perivascular adipose tissue and
atherosclerosis. C R Biol. 340:156–163. 2017.(In French).
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Quiros M and Nusrat A: RhoGTPases,
actomyosin signaling and regulation of the epithelial apical
junctional complex. Semin Cell Dev Biol. 36:194–203. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Petrache I, Birukov K, Zaiman AL, Crow MT,
Deng H, Wadgaonkar R, Romer LH and Garcia JG: Caspase-dependent
cleavage of myosin light chain kinase (MLCK) is involved in
TNF-alpha-mediated bovine pulmonary endothelial cell apoptosis.
FASEB J. 17:407–416. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Miao W, Wu X, Wang K, Wang W, Wang Y, Li
Z, Liu J, Li L and Peng L: Sodium butyrate promotes reassembly of
tight junctions in caco-2 monolayers involving inhibition of
MLCK/MLC2 pathway and phosphorylation of PKCβ2. Int J Mol Sci.
17:1696–1708. 2016. View Article : Google Scholar
|
|
12
|
Stull JT, Kamm KE and Vandenboom R: Myosin
light chain kinase and the role of myosin light chain
phosphorylation in skeletal muscle. Arch Biochem Biophys.
510:120–128. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brasier AR: The nuclear
factor-kappaB-interleukin-6 signalling pathway mediating vascular
inflammation. Cardiovasc Res. 86:211–218. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yokota T, Nishio H, Kubota Y and Mizoguchi
M: The inhibitory effect of glabridin from licorice extracts on
melanogenesis and inflammation. Pigment Cell Res. 11:355–361. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Simmler C, Pauli GF and Chen SN:
Phytochemistry and biological properties of glabridin. Fitoterapia.
90:160–184. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cui YM, Ao MZ, Li W and Yu LJ: Effect of
Glabridin from Glycyrrhiza glabra on Learning and Memory in Mice.
Planta Med. 74:377–380. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hu ZP, Fang XL, Fang N, Wang XB, Qian HY,
Cao Z, Cheng Y, Wang BN and Wang Y: Melatonin ameliorates vascular
endothelial dysfunction, inflammation, and atherosclerosis by
suppressing the TLR4/NF-κB system in high-fat-fed rabbits. J Pineal
Res. 55:388–398. 2013.PubMed/NCBI
|
|
18
|
Zhang C, Zheng H, Yu Q, Yang P, Li Y,
Cheng F, Fan J and Liu E: A practical method for quantifying
atherosclerotic lesions in rabbits. J Comp Pathol. 142:122–128.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang H, Zhu HQ, Feng W, Zhou Q, Gui SY and
Yuan W: MicroRNA-1 prevents high-fat diet-induced endothelial
permeability in apoE knock-out mice. Mol Cell Biochem. 378:153–159.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou B, Pan Y, Hu Z, Wang X, Han J, Zhou
Q, Zhai Z and Wang Y: All-trans-retinoic acid ameliorated high fat
diet-induced atherosclerosis in rabbits by inhibiting platelet
activation and inflammation. J Biomed Biotechnol. 2012:2596932012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Agil A, Navarro-Alarcón M, Ruiz R,
Abuhamadah S, El-Mir MY and Vázquez GF: Beneficial effects of
melatonin on obesity and lipid profile in young Zucker diabetic
fatty rats. J Pineal Res. 50:207–212. 2011.PubMed/NCBI
|
|
23
|
Liuzzo G: Atherosclerosis: An inflammatory
disease. Rays. 26:221–230. 2001.PubMed/NCBI
|
|
24
|
Fatkhullina AR, Peshkova IO and Koltsova
EK: The role of cytokines in the development of atherosclerosis.
Biochemistry (Mosc). 81:1358–1370. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Libby P: Inflammation in atherosclerosis.
Arterioscler Thromb Vasc Biol. 32:2045–2051. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Corre I, Paris F and Huot J: The p38
pathway, a major pleiotropic cascade that transduces stress and
metastatic signals in endothelial cells. Oncotarget. 8:55684–55714.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yanni AE: The laboratory rabbit: An animal
model of atherosclerosis research. Lab Anim. 38:246–256. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bocan TM, Mueller SB, Mazur MJ, Uhlendorf
PD, Brown EQ and Kieft KA: The relationship between the degree of
dietary-induced hypercholesterolemia in the rabbit and
atherosclerotic lesion formation. Atherosclerosis. 102:9–22. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Urbano RL, Furia C, Basehore S and Clyne
AM: Stiff substrates increase inflammation-induced endothelial
monolayer tension and permeability. Biophys J. 113:645–655. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bryk D, Olejarz W and Zapolska-Downar D:
Mitogen-activated protein kinases in atherosclerosis. Postepy Hig
Med Dosw (Online). 68:10–22. 2014.(In Polish). View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hao XZ and Fan HM: Identification of
miRNAs as atherosclerosis biomarkers and functional role of miR-126
in atherosclerosis progression through MAPK signalling pathway. Eur
Rev Med Pharmacol Sci. 21:2725–2733. 2017.PubMed/NCBI
|
|
32
|
Wagner EF and Nebreda AR: Signal
integration by JNK and p38 MAPK pathways in cancer development. Nat
Rev Cancer. 9:537–549. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Harding A, Cortez-Toledo E, Magner NL,
Beegle JR, Coleal-Bergum DP, Hao D, Wang A, Nolta JA and Zhou P:
Highly efficient differentiation of endothelial cells from
pluripotent stem cells requires the MAPK and the PI3K pathways.
Stem Cells. 35:909–919. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zuo L, Yang X, Lu M, Hu R, Zhu H, Zhang S,
Zhou Q, Chen F, Gui S and Wang Y: All-trans retinoic acid inhibits
human colorectal cancer cells RKO migration via downregulating
myosin light chain kinase expression through MAPK signaling
pathway. Nutr Cancer. 68:1225–1233. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sellers JR and Adelstein RS: Regulation of
contractile activity. Enzymes. Tamanoi F: 18. 3rd. Academic press;
New York, NY: pp. pp381–418. 1987, View Article : Google Scholar
|
|
36
|
Tan JL, Ravid S and Spudich JA: Control of
nonmuscle myosins by phosphorylation. Annu Rev Biochem. 61:721–759.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang B, Yan Y, Zhou J, Zhou Q, Gui S and
Wang Y: A novel all-trans retinoid acid derivatives inhibits the
migration of breast cancer cell lines MDA-MB-231 via myosin light
chain kinase involving p38-MAPK pathway. Biomed Pharmacother.
67:357–362. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ringvold HC and Khalil RA: Protein kinase
C as regulator of vascular smooth muscle function and potential
target in vascular disorders. Adv Pharmacol. 78:203–301. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yu T, Wang Y, Qian D, Sun X, Tang Y, Shen
X and Lin L: Advanced glycation end products impair Ca2+
mobilization and sensitization in colonic smooth muscle cells via
the CAMP/PKA pathway. Cell Physiol Biochem. 43:1571–1587. 2017.
View Article : Google Scholar : PubMed/NCBI
|