Introduction
Resveratrol (3,5,4′-trihydroxy-trans-stilbene), a
stilbenoid belonging to the stilbene family, was first reported by
Takaoka in 1939 (1). It is a natural
polyphenolic phytoalexin produced by a number of plants, including
Arachis hypogaea Linn. and Vitis vinifera Linn., in response to
fungal infection (2). Sources of
resveratrol include food, such as the skin of grapes and red wine
(3,4). Resveratrol is a natural compound
indicated to have beneficial health effects, particularly
anticancer effects in humans (5,6). In
1997, Jang et al (5)
determined its chemopreventive properties, including antioxidant,
antimutagen, anti-inflammatory and anti-progression activity in a
number of disease models, including cancer. Subsequently, numerous
studies revealed that resveratrol exhibits multiple anticancer
effects, preventing tumor formation in different cancer types,
including leukemia, breast cancer, skin tumor, colorectal cancer
and liver cancer (6–12).
Gastric cancer was the fifth most common cancer
globally, with almost 951,000 new cases occurring (6.8% of the
total), causing an estimated 723,000 cancer-associated mortalities
in 2012, becoming the third leading cause of cancer-associated
mortality (13). Of gastric cancer
cases, >70% were estimated to occur in developing countries and
half of the total new cases occurred in China in 2012 (13). The estimated mortality rates are
notably high in Eastern Asia (14.0/100,000 in males and 9.8/100,000
in females), but low in Northern America (2.8/100,000 in males and
1.5/100,000) (13). In the clinic,
surgery, chemotherapy and radiotherapy are the primary treatment
options for gastric cancer (14–16).
Resveratrol is a polyphenol compound used in traditional Chinese
medicine and has beneficial effects as a cancer chemopreventive
agent in humans (5–7); however, there are limited studies
focused on the action of resveratrol regarding the treatment and
prevention of gastric cancer, and the anticancer mechanism of
resveratrol remains unclear.
In the present study, the effects of resveratrol on
gastric cancer cell line BGC823, the underlying mechanisms of the
involvement of resveratrol and the role of metastasis-associated
lung adenocarcinoma transcript 1 (MALAT1) in
epithelial-to-mesenchymal transition were investigated.
Materials and methods
Cell culture
Human gastric cancer cell lines SGC7901 and BGC823
were purchased from Cell-Land Biotech Co., Ltd. (Hangzhou, China;
www.cell-land.com). Non-malignant gastric
epithelium cell line GES1 was obtained from Cell Bank of Type
Culture Collection of Chinese Academy of Sciences (Shanghai,
China). All cells were cultured in RPMI-1640 medium supplemented
with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and maintained at 37°C in an atmosphere
containing 5% CO2 with saturated humidity in a humidified cell
incubator (Thermo Fisher Scientific, Inc.). The cells were
collected during their logarithmic phase and stored at −80°C for
further study.
RNA interference
MALAT1 siRNA and negative control siRNA (siNC) were
obtained from Guangzhou RiboBio Co., Ltd. (Guangzhou, China). The
following sequences were used in the present study: siRNA-1, sense,
5′-GCAAAUGAAAGCUACCAAU-3′ and antisense 5′-AUUGGUAGCUUUCAUUUGC-3′;
siRNA-2, sense, 5′-CUAGAAUCCUAAAGGCAAA-3′ and antisense,
5′-UUUGCCUUUAGGAUUCUAG-3′; and siNC, sense,
5′-UUCUCCGAACGUGUCACGU-3′ and anti-sense,
5′-ACGUGACACGUUCGGAGAA-3′. A total of 2 ml BGC823 cells (8×104
ells/ml) were plated onto 6 well plates and grown overnight at 37°C
in an atmosphere containing 5% CO2 in a humidified cell incubator.
Cell transfections were performed with the Lipofectamine RNAiMAX
Reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. The final siRNA oligonucleotide
concentration was 20 pM. Following 24, 48, 72 or 96 h of
incubation, the transfected cells were harvested to be used in
other experiments. Cell transfected with siNC were used as the
control.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from cells using
TRIzol® reagent and an RNA Fast Mini kit (cat. no.
GK3016; Generay Biotech Co., Ltd., Shanghai, China). cDNA was
synthesized using a RevertAid First Strand cDNA synthesis kit (cat.
no. K1622; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. RT-qPCR was performed using a CFX connect
Real-Time PCR system with SsoAdvance Universal SYBR®
Green Supermix (cat. no. 172-5274; both Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) in the following cycling conditions: 95°C
denaturation for 3 min; 40 cycles of denaturation at 95°C for 15
sec, annealing at 59°C for 30 sec and extension at 72°C for 30 sec.
GAPDH was used as a reference gene and all reactions were performed
in triplicate. The following primers were used in the current
study: Long non-coding RNA (lncRNA) MALAT1 (Gene ID, 378938;
www.ncbi.nlm.nih.gov), forward,
5′-ATACCTAACCAGGCATAAC-3′, and reverse, 5′-GTAGACCAACTAAGCGAAT-3′;
GAPDH (Gene ID: 2597), forward, 5′-CGGATTTGGTCGTATTG-3′; and
reverse, 5′-GAAGATGGTGATGGGATT-3′; E-cadherin (Gene ID, 999),
forward, 5′-CGCATTGCCACATACAC-3′, and reverse,
5′-CCTTCCATGACAGACCC-3′; and vimentin (Gene ID, 7431), forward,
5′-TTGAACGCAAAGTGGAATC-3′, and reverse, 5′-AGGTCAGGCTTGGAAACA-3′.
The primers were synthesized by Invitrogen (Thermo Fisher
Scientific, Inc.). Relative levels of each RNA were calculated
using Bio-Rad CFX Manager software (version 3.1; Bio-Rad
Laboratories, Inc.) (17).
Cell viability assay
A Cell Counting Kit-8 (CCK-8) assay was performed to
determine the viability of the gastric cancer cells according to
the manufacturer's protocol (CCK-8; Dojindo Molecular Technologies,
Inc., Kumamoto, Japan). Firstly, 100 µl cell suspensions (3×104
cells/well) were seeded in a 96-well plate and pre-incubated for 24
h at 37°C in a humidified incubator. Secondly, a series of various
concentrations of resveratrol (0, 5, 10, 25, 50, 100, 200 and 400
µM; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) were added to
the plate (100 µl/well) and were then pre-incubated for various
periods of time (24, 48 and 72 h) at 37°C. In another experiment to
assess the effect of small interfering (siRNA)-mediated MALAT1
knockdown on cell viability, cells were transfected with siRNA-1
for 24, 48, 72 and 96 h, respectively. Subsequently, 10 µl CCK-8
solution was added to each well followed by incubation at 37°C for
1 h with protection from light. All experiments were performed in
triplicate. The absorbance was measured at a wavelength of 450 nm
using a microplate reader (Bio-Rad Laboratories, Inc.) and
presented as the mean optical density (OD). The percentage of
viable cells=OD of the treatment group/OD of the control group ×
100%.
Cell invasion and migration assay
BGC823 cells (3×105 cells/well) were cultured in a
humidified incubator in RPMI-1640 medium with 10% FBS at 37°C in an
atmosphere containing 5% CO2 for 24 h. A total of 200 µM
resveratrol was added to the cells, followed by incubation for 48 h
at 37°C. The treated cells were digested with 0.02% EDTA
supplemented with 0.25% trypsin and collected for trypan blue
staining to count the cells at 37°C. For invasion analysis, 100 µl
Matrigel (BD Biosciences, San Jose, CA, USA) was firstly added to
the bottom of the Transwell chamber (8 µm) prior to BGC823 cells
being seeded. RPMI-1640 medium with 10% FBS were plated in the
lower chamber. Subsequently, 2×104 cells were placed into the upper
chamber with RPMI-1640 medium and were allowed to invade for 24 h.
Following this, the Transwell chambers were removed, and
non-invading cells were removed from the upper chamber with a
cotton swab. The upper chamber was then washed with PBS.
Subsequently, cells of the upper chamber were fixed and stained
with 0.1% crystal violet staining solution (Sigma-Aldrich; Merck
KGaA) for 30 min at 37°C. The cells were counted and imaged under
an ECLIPSE Ti-S microscope (Nikon Corporation, Tokyo, Japan) at a
magnification of ×100.
For the cell migration assay, cells (5×104
cells/well) were plated in the upper chamber with RPMI-1640 medium,
whereas the bottom chamber was filled with 600 µl RPMI-1640 medium
supplemented with 10% FBS. Following incubation for 16 h, the cells
were stained with crystal violet staining solution for 30 min at
37°C (Sigma-Aldrich; Merck KGaA), counted and imaged under an
ECLIPSE Ti-S microscope. The cells without resveratrol (Mock) were
used as the control group.
Western blot analysis
Cell lysates were prepared for western blot analysis
as previously described (7). The
samples were analyzed with a standard bicinchoninic acid (BCA)
assay for protein concentration using a BCA Protein Assay kit
(Beyotime Institute of Biotechnology, Shanghai, China). Total
protein (30 µg) was loaded per lane, separated by SDS-PAGE on a
12.5% gel and then all protein samples were transferred to 0.45-µm
Immobilon® polyvinylidene difluoride (PVDF) membranes
(EMD Millipore, Billerica, MA, USA). For western blots, the PVDF
membrane was blocked with TBS-T buffer containing 5% (w/v) nonfat
dry milk, slowly agitated on a shaker and sealed at room
temperature for 2 h. The primary antibodies [dilution, 1:1,000
(v/v)] used in the present study included E-cadherin (cat. no.
3195), vimentin (cat. no. 5741) and GAPDH (cat. no. 5174), which
were purchased from Cell Signaling Technology, Inc. (Danvers, MA,
USA). A horseradish peroxidase-conjugated secondary antibody [goat
anti-rabbit immunoglobulin G; cat. no. BL003A; 1:2,000 (v/v);
Biosharp, Inc., Hefei, China] was incubated at room temperature for
2 h. GAPDH was used as the control. The signals were visualized
with a Clarity™ Western ECL Substrate (cat. no. 170-5060; Bio-Rad
Laboratories, Inc.) and detected with a ChemiDoc™ Touch Imaging
system (version 5.2; Bio-Rad Laboratories, Inc.).
Statistical analysis
All data were analyzed using SPSS 20.0 software (IBM
Corp., Armonk, NY, USA). Data are presented as the mean ± standard
deviation. The mean values of the control and treatment groups were
compared using one-way analysis of variance and Fisher's least
significant difference tests. P<0.05 was considered to indicate
a statistically significant difference.
Results
Resveratrol inhibits viability of
human gastric cancer cells
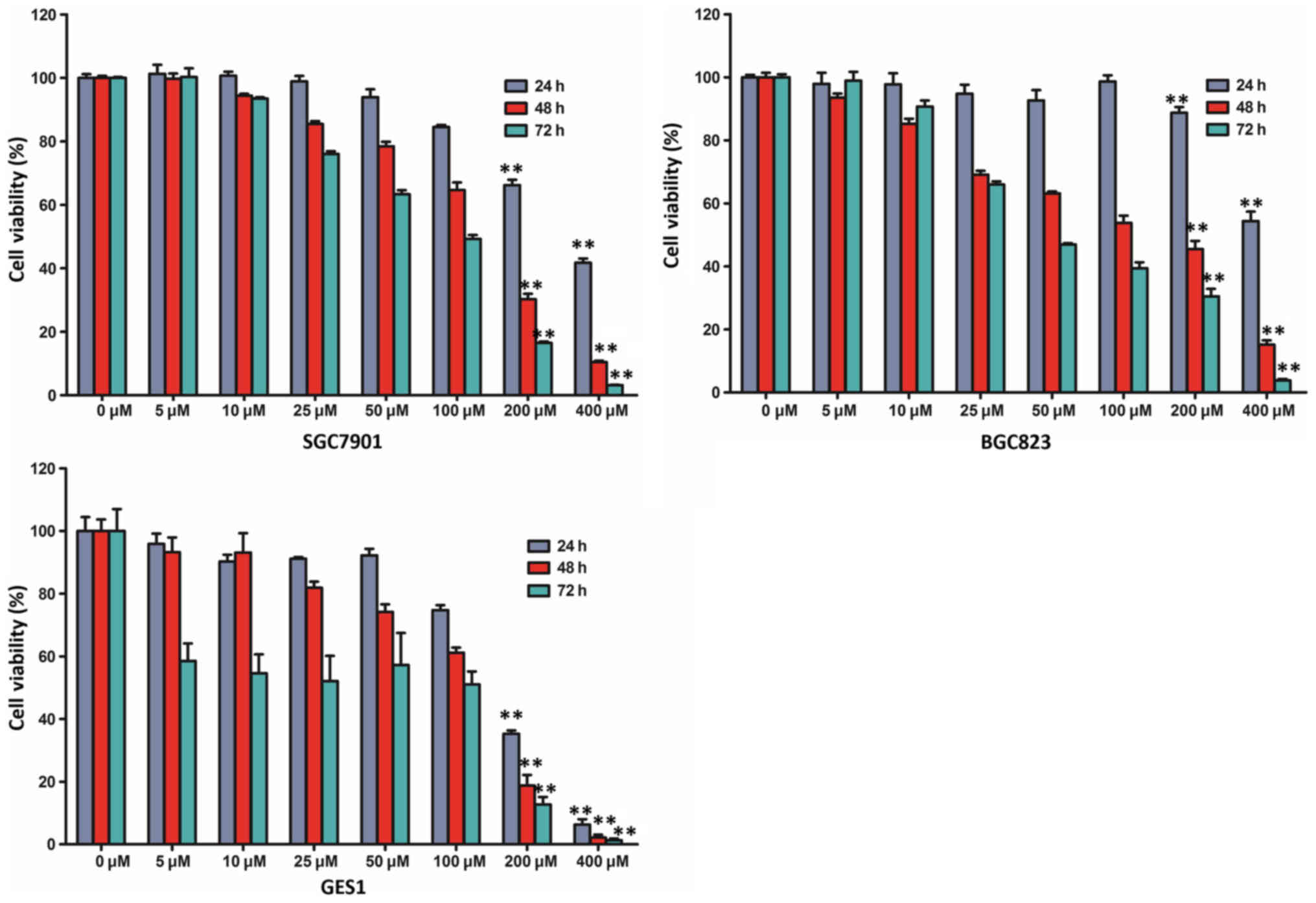
In order to assess the effect of resveratrol on cell
viability, a cell viability assay was performed using two human
gastric cancer cell lines (SGC7901 and BGC823) and a non-malignant
gastric epithelium cell line (GES1). BGC823, SGC7901 and GES1 cells
were treated with various doses of resveratrol (0, 5, 10, 25, 50,
100, 200 and 400 µM) and examined with a CCK-8 assay. The viability
of the three cell lines was markedly reduced following treatment
with resveratrol in a time- and dose-dependent manner (Fig. 1). The CCK-8 assay indicated that the
viability of BGC823 and SGC7901 cells incubated with 200 or 400 µM
resveratrol for 48 and 72 h was markedly lower than those incubated
for 24 h. In addition, compared with the untreated control group,
treatment with 200 µM resveratrol for 48 h reduced the viability of
BGC823 cells by >50% (45.48±2.61%; P<0.01). The half maximal
inhibitory concentration (IC50 value) was closest to 200 µM
following treatment with resveratrol for 48 h, and, therefore, 200
µM resveratrol administration was selected for further study,
according to the cell viability assay.
Resveratrol inhibits migration and
invasion of BGC823 cells
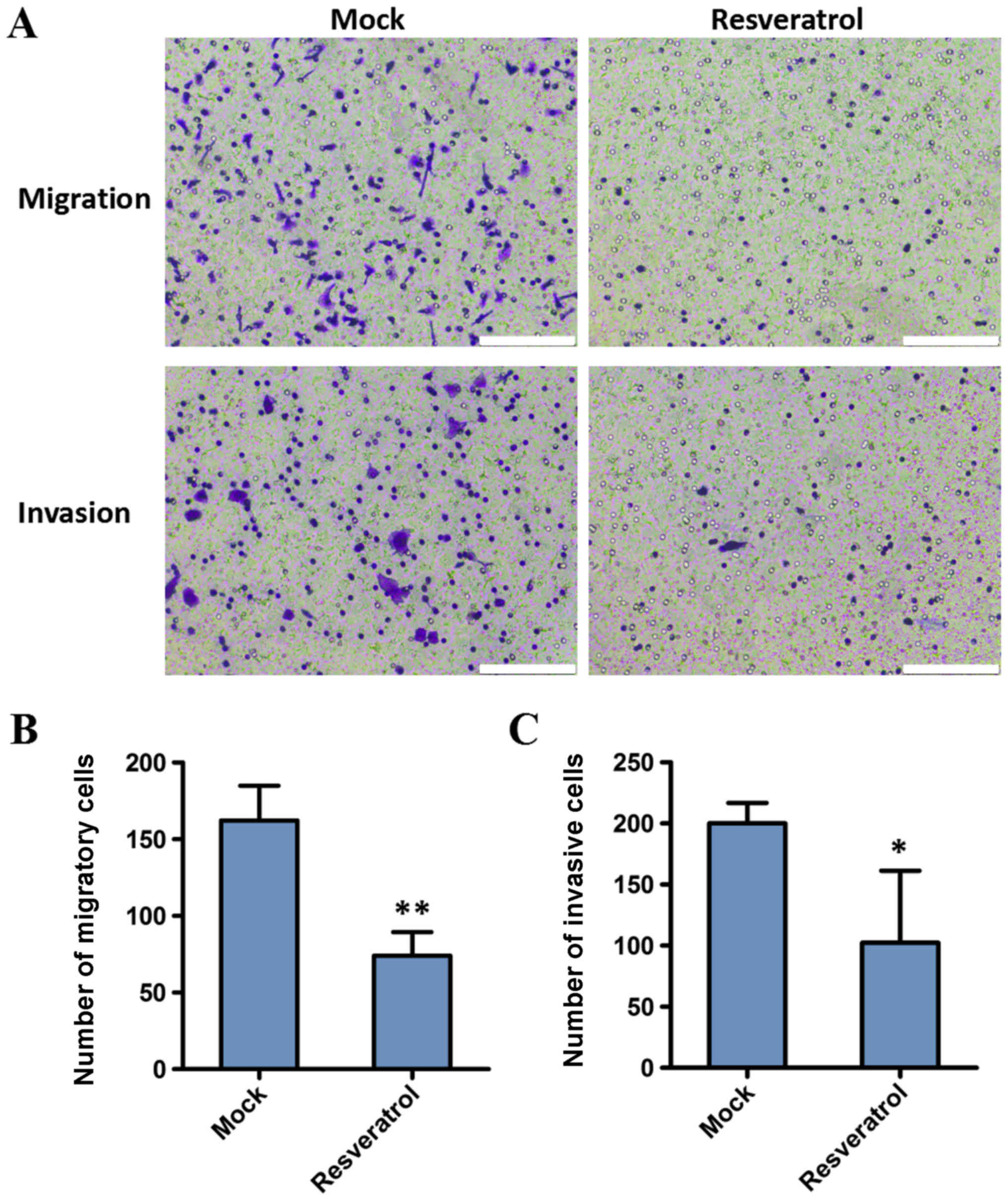
The present study investigated the effect of
resveratrol on the migration and invasion capacities of BGC823
cells. Cell invasion and migration assays were conducted following
treatment with 200 µM resveratrol for 48 h. The results
demonstrated that the migration and invasion decreased in BGC823
cells following administration of resveratrol, compared with the
control (162.25±22.60 vs. 74.00±15.38, P<0.01; and 200.00±16.70
vs. 102.33±58.97, P<0.05, respectively). These results indicated
that the invasion and migration abilities of gastric cancer cells
were significantly inhibited by resveratrol (Fig. 2).
Resveratrol suppresses
epithelial-to-mesenchymal transition
Since epithelial-to-mesenchymal transition is an
important process facilitating tumor invasion and metastasis
(18,19), the effect of resveratrol on
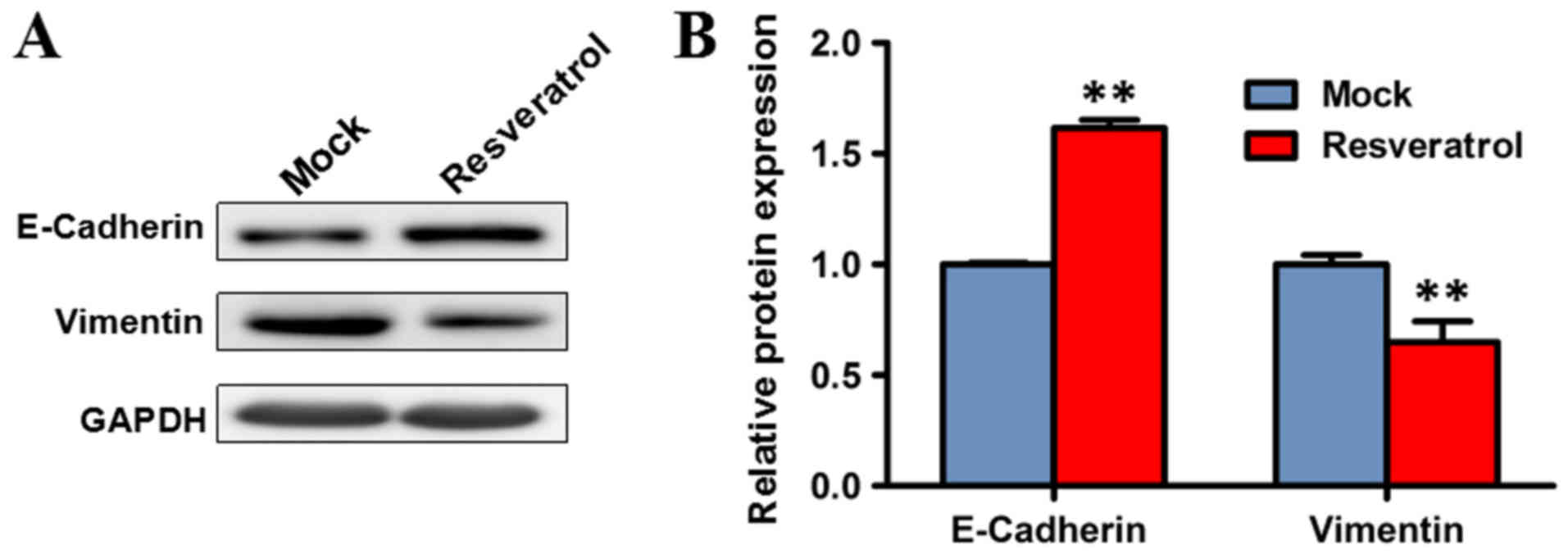
epithelial-to-mesenchymal was investigated. Western blot analysis
was performed to evaluate the expression levels of markers of
epithelial-to-mesenchymal transition, including E-cadherin and
vimentin, following treatment with resveratrol. The expression
level of vimentin significantly decreased in BGC823 cells treated
with 200 µM resveratrol for 48 h, compared with the mock group
(P<0.01), while the protein expression of E-cadherin was
significantly increased (P<0.01). These results indicated that
resveratrol may suppress epithelial-to-mesenchymal transition
(Fig. 3).
Resveratrol inhibits the expression of
MALAT1
MALAT1 was reported as a prognostic biomarker of
gastric cancer, and had significantly higher expression in gastric
cancer tissues of patients with distant metastasis compared with
adjacent normal tissues of those patients and gastric cancer
patients without distant metastasis (20). To determine whether resveratrol
exhibits any prognostic significance in gastric cancer cells
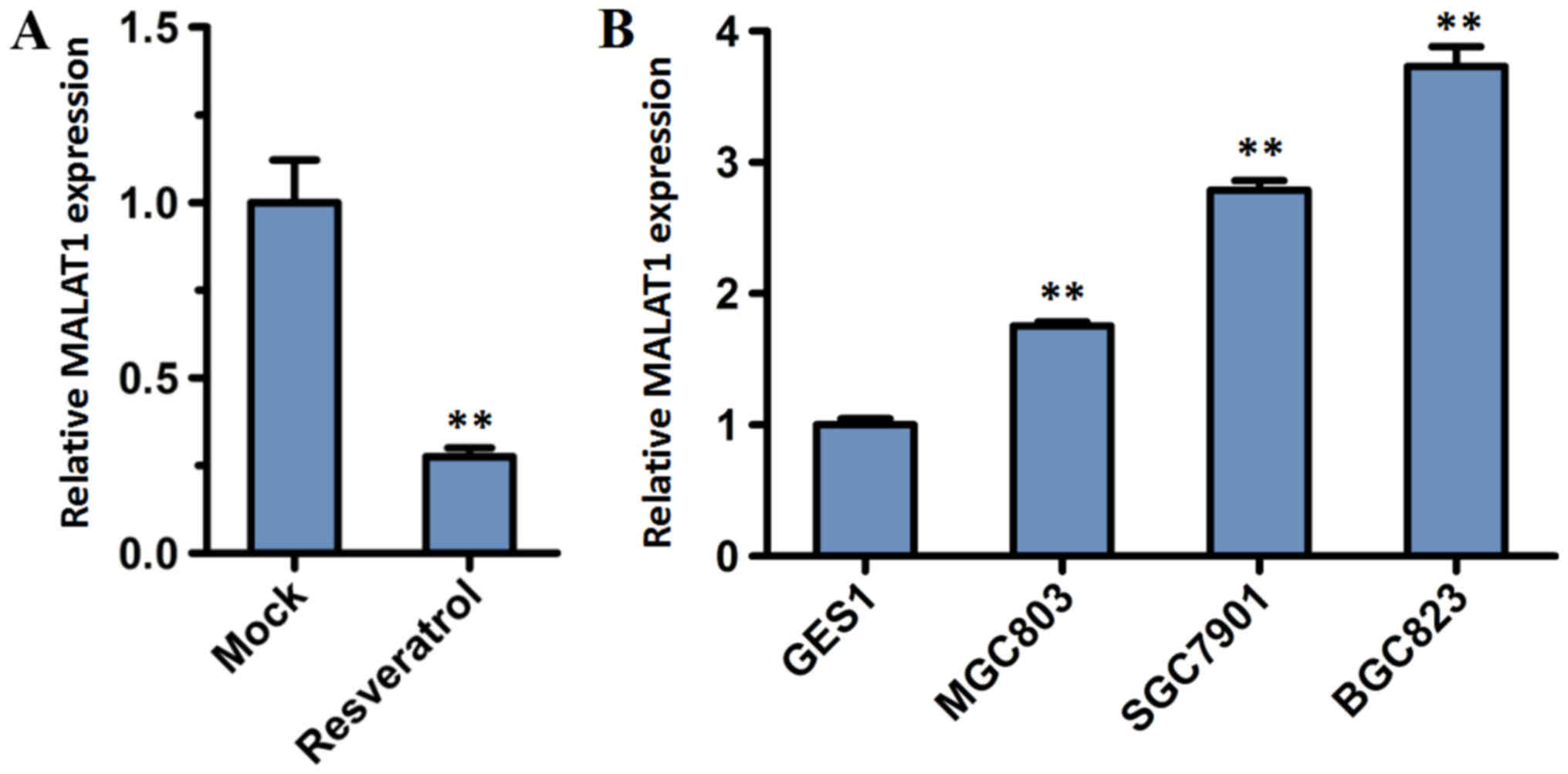
through the role of MALAT1, the expression level of lncRNA MALAT1
was analyzed. BGC823 cells were incubated for 48 h with or without
200 µM resveratrol. RT-qPCR analysis demonstrated that the relative
expression of lncRNA MALAT1 in BGC823 cells decreased significantly
following treatment with resveratrol, compared with the mock group
(P<0.01; Fig. 4A).
MALAT1 is overexpressed in gastric
cancer cells
To investigate the lncRNA MALAT1 expression in tumor
cells, a normal cell line (GES1) and three GC cell lines (BGC823,
MGC803 and SGC7901) were subjected to RT-qPCR analysis. The
expression level of MALAT1 increased in the BGC823, MGC803 and
SGC7901 cell lines compared with the normal GES1 cell line
(Fig. 4B). Furthermore, the
expression level of MALAT1 was highest in BGC823 cells.
MALAT1 knockdown suppresses the
viability of BGC823 cells
To determine the involvement of MALAT1 in the
biological function of gastric cancer, MALAT1 was knocked down in
the present study. Firstly, two individual siRNA oligonucleotides
of MALAT1 (siRNA-1 and siRNA-2) were synthesized and transfected
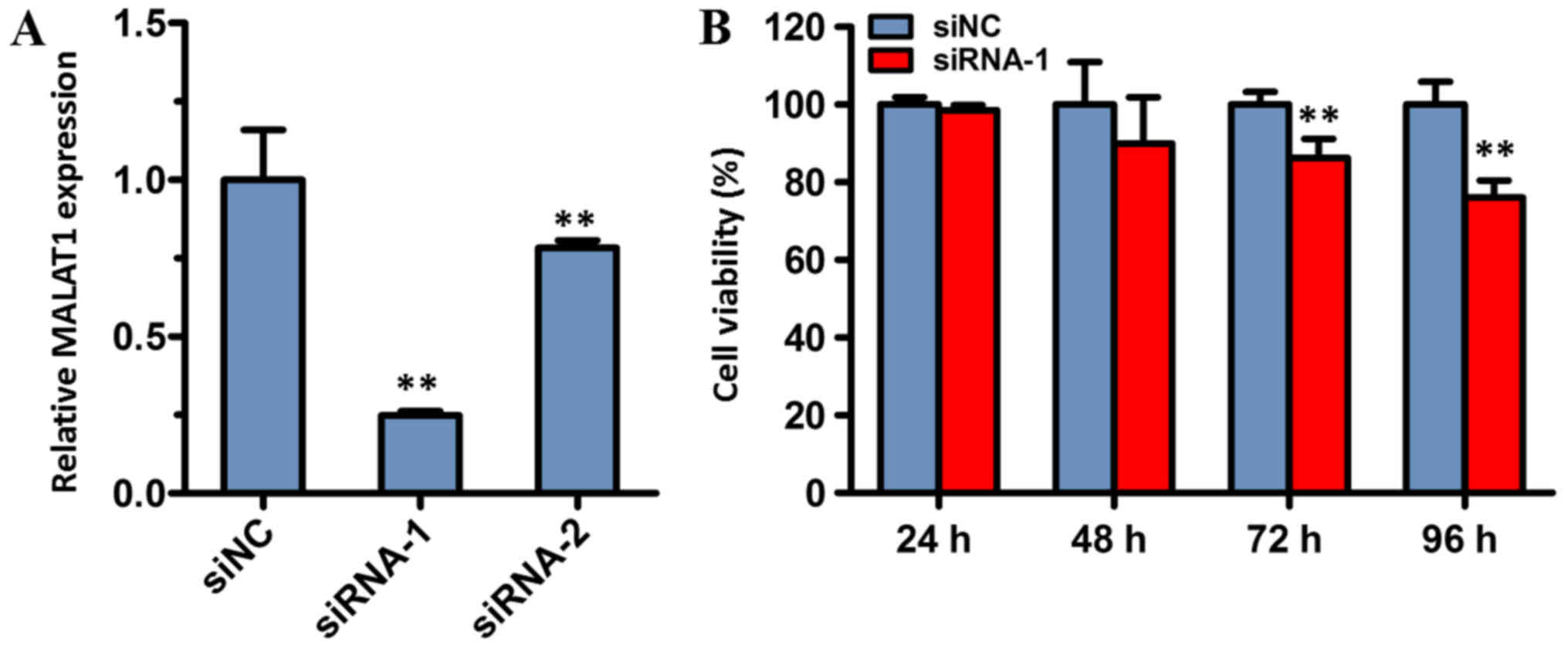
into BGC823 cells. The results demonstrated that the expression
level of MALAT1 in cells transfected with siRNA-1 or siRNA-2 was
significantly reduced compared with cells transfected with siNC
(both P<0.01; Fig. 5A). The
expression of MALAT1 was the lowest in the siRNA-1 group, and,
therefore, this group was used in the subsequent experiments.
To assess the effect of siRNA-mediated MALAT1
knockdown on the viability of cancer cells, the viability of BGC823
cells was determined with a CCK-8 assay. The results of the CCK-8
assay revealed that cell viability of the siRNA-1 treatment group
was significantly decreased compared with the siNC group, following
72 and 96 h of incubation. However, there was no significant
difference between the siRNA-1 group and the control group
following transfection for 24 or 48 h. These data indicated that
MALAT1 may serve a role in cell viability.
Effect of MALAT1 knockdown on the
migration and invasion of gastric cancer cells
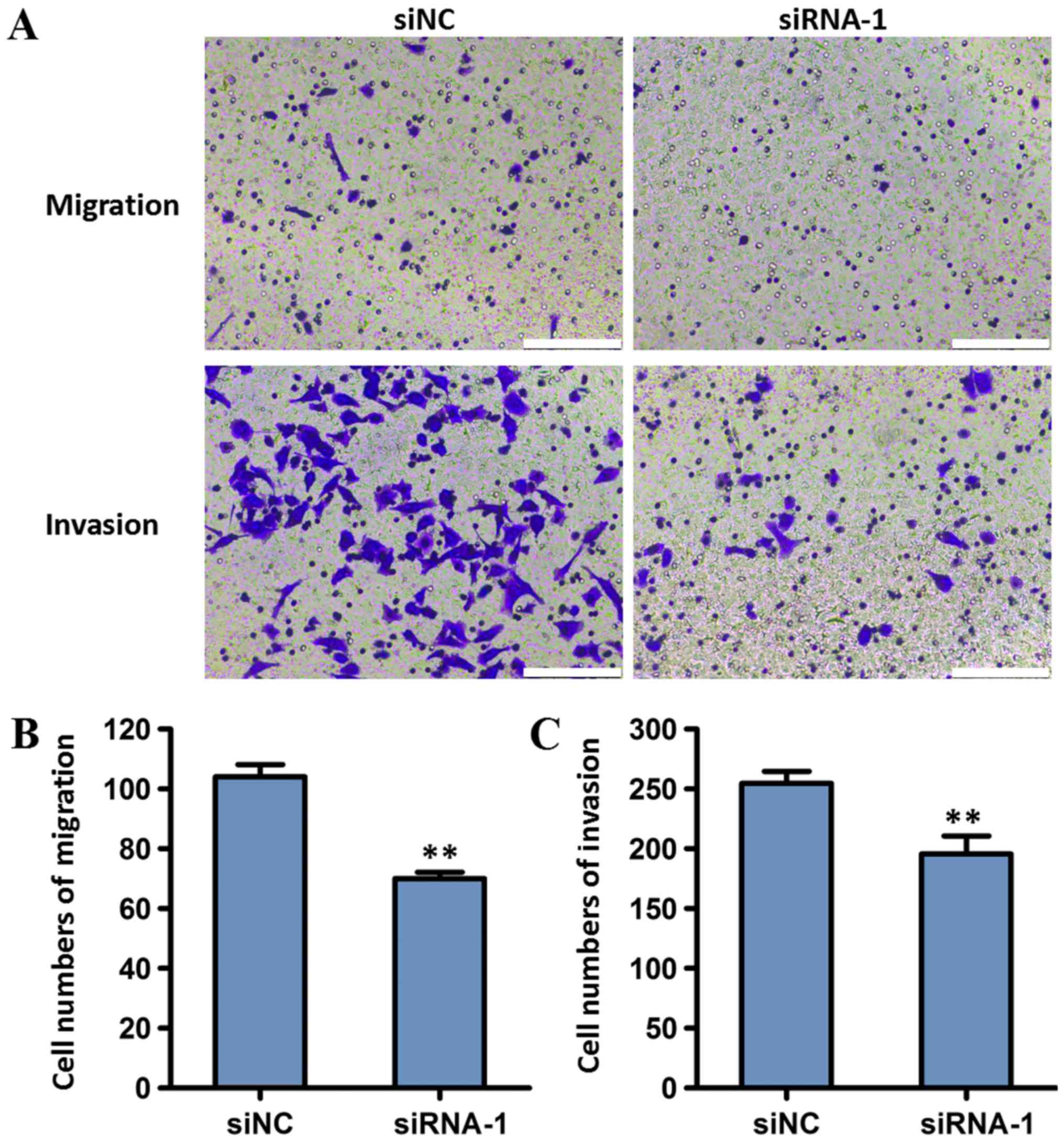
To investigate the effect of MALAT1 on the invasion
and migration of tumor cells, BGC823 cells transfected with MALAT1
siRNA-1 were used in transwell migration and Matrigel invasion
assays. Compared with the siNC group, cells transfected with
siRNA-1 exhibited decreased invasion and migration (P<0.01;
Fig. 6). These results indicated
that MALAT1 knockdown may reduce the migration and invasion of
gastric cancer cells.
MALAT1 enhances
epithelial-to-mesenchymal transition in BGC823 cells
Previous studies indicated that
epithelial-to-mesenchymal transition serves an important role in
tumor invasion and metastasis (17,18).
Therefore, the potential effect of MALAT1 on
epithelial-to-mesenchymal transition was investigated in the
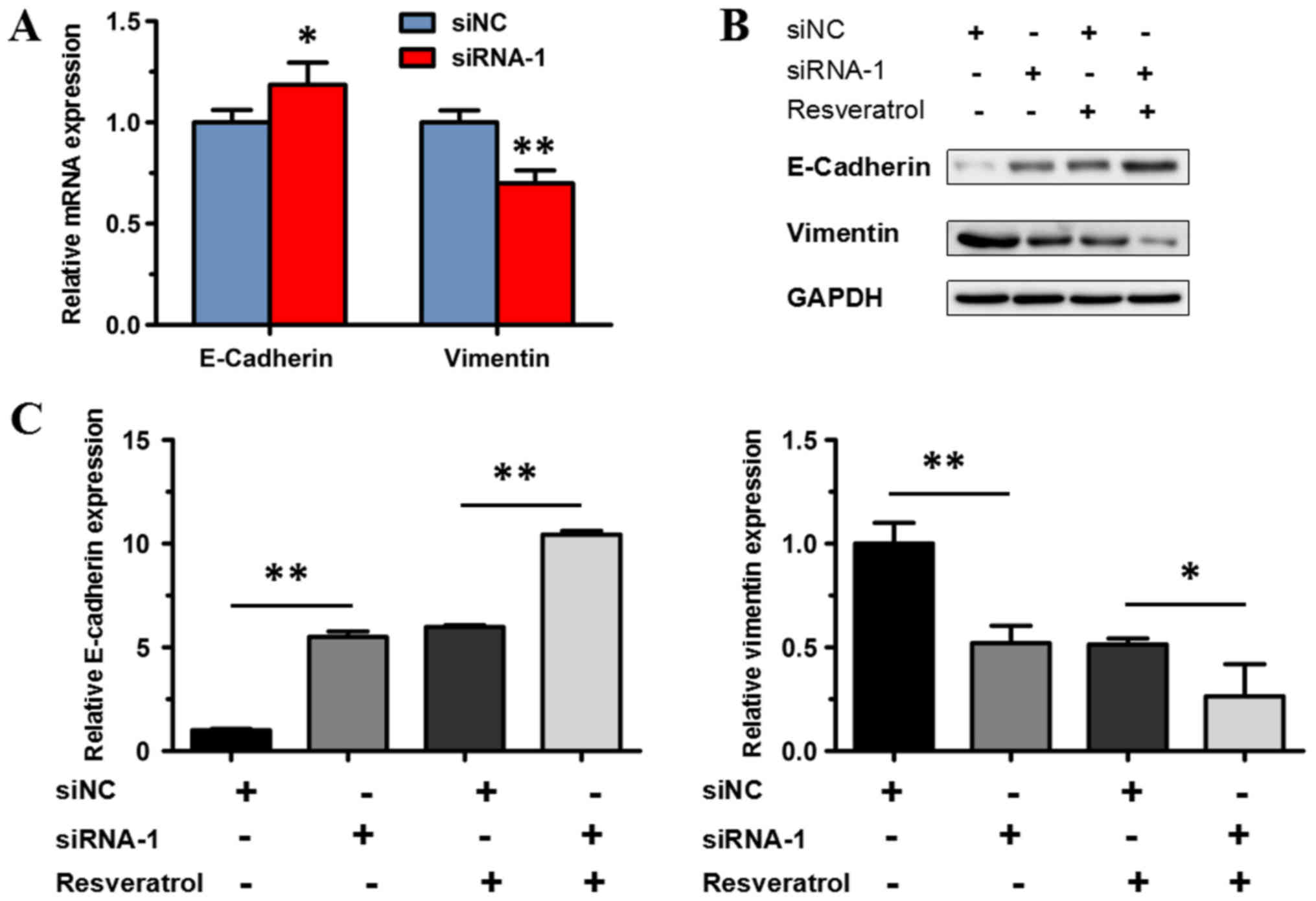
present study. siRNA-1 was used to knock down the expression of
MALAT1 in the transfected BGC823 cells. RT-qPCR and western blot
analysis were used to detect E-cadherin and vimentin expression in
BGC823 cells treated with siRNA-1. mRNA expression of E-cadherin
increased significantly in BGC823 cells transfected with siRNA-1,
compared with the siNC group (P<0.05); while the expression of
vimentin significantly decreased (P<0.01; Fig. 7A). Western blot analysis revealed
that protein expression of E-cadherin increased significantly in
BGC823 cells transfected with siRNA-1 compared with the siNC group
(P<0.01), while the expression of vimentin decreased
significantly (P<0.01; Fig. 7B and
C).
In order to investigate whether MALAT1 knockdown
abolishes the effect of resveratrol on epithelial-to-mesenchymal
transition, resveratrol was administered to BGC823 cells
transfected with siRNA-1. The protein expression of vimentin
decreased significantly (P<0.05), and the protein expression of
E-cadherin increased significantly in BGC823 cells transfected with
siRNA-1 following treatment with 200 µM resveratrol, compared with
the control (the siNC group treated with resveratrol; P<0.01;
Fig. 7B and C). The aforementioned
result indicated that transfection with siRNA-1 could not abolish
the effect of resveratrol, and, therefore, resveratrol may suppress
MALAT1-mediated epithelial-to-mesenchymal transition. These results
demonstrated that downregulated expression of MALAT1 inhibited
epithelial-to-mesenchymal transition in BGC823 cells, indicating
that MALAT1 may enhance epithelial-to-mesenchymal transition in
gastric cancer cells.
Discussion
lncRNAs are >200 nucleotides in length and
function in various aspects of cell biological processes, including
proliferation, apoptosis, invasion, metastasis and
epithelial-to-mesenchymal transition, by transcriptional and
post-transcriptional regulation of gene expression or epigenetic
regulation (21,22). A number of lncRNAs, including H19,
HOX antisense intergenic RNA and gastric carcinoma high expressed
transcript 1, were upregulated in different gastric cancer cell
lines (23–26). Furthermore, MALAT1, also known as
nuclear-enriched abundant transcript 2, is a frequently expressed
lncRNA that has been associated with metastatic potential and poor
prognosis in lung adenocarcinoma (19). Numerous studies have demonstrated
that MALAT1 is associated with various tumor types, including lung
cancer, hepatocellular carcinoma, prostate cancer, gallbladder
cancer, bladder cancer, colorectal cancer, renal cell carcinoma and
pancreatic cancer (27–36). Additionally, a number of reports also
provided evidence for the biological and clinical significance of
MALAT1 expression as a prognostic biomarker of gastric cancer
(20,37–39). Xia
et al (20) and Okugawa et
al (37) revealed that the
tissue MALAT1 expression level was significantly increased in
gastric cancer tissues, compared with normal mucosa, and high
expression of MALAT1 was independently associated with a poor
prognosis for patients with gastric cancer. The study by Wang et
al (38) demonstrated that
MALAT1 expression was notably increased in gastric cancer cell
lines, compared with the levels in GES1 cells (38). In the present study, it was
determined that the expression level of MALAT1 increased in gastric
cancer cells lines BGC823, MGC803 and SGC7901, compared with normal
gastric epithelial cell line GES1, and the expression of MALAT1 was
highest in BGC823 cells. The present results were supported by the
studies by Wang et al (38)
and Li et al (39), but
partly conflicted with data reported by Xia et al (20), whose study indicated that the
expression level of MALAT1 was increased in gastric cancer cell
lines MKN45, CTC105 and CTC141, but lower in SGC7901 and BGC823,
compared with GES1 cells (17).
Therefore, further studies are necessary to investigate the
expression of MALAT1 in various tumor cells.
MALAT1 is a potential biomarker of gastric cancer
(19). Functional studies have
demonstrated that the knockdown of MALAT1 could suppress cell
viability, migration and invasion in gastric cancer cell lines
MKN45, CTC141, SGC996 and NOZ (20,40).
Another study indicated that mice administered with MALAT1-depleted
BGC823 or NOZ cells exhibited a reduced degree of lung metastasis
compared with mice treated with wild type BGC823 and NOZ cell lines
(39). The current study revealed
that MALAT1 knockdown notably suppressed the viability, migration
and invasion of gastric cancer BGC823 cells, indicating the
potential function of MALAT1 in promoting metastasis.
Epithelial-to-mesenchymal transition is an important
process for human in wound healing, organ fibrosis and initiation
of metastasis for cancer progression (18). When epithelial-to-mesenchymal
transition occurs, epithelial cells lose polarity and relinquish
cell-cell adhesion to gain migratory and invasive properties and
become mesenchymal stem cells (40,41).
MALAT1 induces epithelial-to-mesenchymal transition in multiple
cancer types, including tongue cancer, cervical cancer, lung
adenocarcinoma and oral squamous cell carcinoma (42–45). The
present results also revealed that MALAT1 enhanced
epithelial-to-mesenchymal transition in gastric cancer BGC823
cells. These results indicated that MALAT1 can promote tumor growth
and metastasis by inducing epithelial-to-mesenchymal transition in
various cancer types.
Previous studies have demonstrated beneficial
effects of resveratrol in preventing and treating cancer (46–48). In
the present study, it was determined that resveratrol inhibited
cell viability in a dose-dependent manner in human gastric cancer
BGC823 cells and suppressed cell migration and invasion. Notably,
the IC50 value of resveratrol was high (~200 µM) in gastric cancer
BGC823 cells after 48 h and resveratrol caused cytotoxicity to
normal gastric GES1 cells. Additionally, this similar concentration
of resveratrol in gastric cancer cells has been reported in other
published articles. Using an MTT assay, Gao et al (49) determined that the viability of
SGC7901 cells was inhibited by ~80% when they were exposed to 200
µM resveratrol for 48 h. Jing et al (50) obtained similar results using an MTT
assay and trypan blue staining, with >80% of MGC803 cell growth
being inhibited after 24 and 48 h of exposure to 200 µM resveratrol
and the IC50 of resveratrol for MGC803 cells was 127 µM. Yang et
al (51) determined that in
three gastric cancer cell lines (AGS, BGC823 and SGC7901), 100 mM
resveratrol was sufficient to induce >50% growth inhibition
following treatment for 24 h (56.78% for AGS; 54.87% for BGC-823;
and 52.16% for SGC7901) using an MTS assay. The present results
demonstrated a similar growth inhibition rate of SGC7901 using a
CCK-8 assay, as well as ~53.8% reduction in the cell viability of
the gastric cancer cell line BGC823. The differences in doses of
resveratrol in previous studies may be due to different methods and
gastric cell lines used.
The underlying mechanism of resveratrol varies in
different tumor types. In prostate cancer, the beneficial effect of
resveratrol was due to inhibition of the protein kinase B
(Akt)/microRNA-21 signaling pathway (52). The study of colorectal cancer
indicated that resveratrol inhibited cell invasion and metastasis
through regulating the MALAT1 and Wnt/β-catenin signaling pathways
(34). Another report revealed that
resveratrol inhibited the progression of the cell cycle by
inhibiting the activity of phosphatase and tensin homolog, which
suppresses the phosphoinositide 3-kinase/Akt pathway in MGC803
cells (50). Furthermore,
resveratrol could suppress invasion and metastasis and inhibit the
hedgehog signaling pathway in gastric cancer SGC7901 cells
(49). Therefore, the anticancer
mechanism of resveratrol remains unclear. In the present study, it
was determined that resveratrol exerted an anticancer function by
suppressing MALAT1-mediated epithelial-to-mesenchymal transition in
human gastric cancer cells. Additionally, the present study
provided novel evidence to understand the anticancer mechanism of
resveratrol.
In conclusion, the present study determined that
resveratrol decreased MALAT1 expression in two gastric cancer cell
lines (BGC823 and SGC7901) and inhibited cell viability in a
dose-dependent manner. Further experiments demonstrated that cell
migration and invasion were inhibited by resveratrol via
suppressing MALAT1-mediated epithelial epithelial-to-mesenchymal
transition in gastric cancer cell line BGC823. These data provided
novel evidence for understanding the anticancer mechanism in
vitro and demonstrated the potential use of resveratrol as an
anticancer drug.
Acknowledgements
The authors would like thank Dr Baoying Fei and Mr.
Junxian Chen in the Department of Gastroenterology, Tongde Hospital
of Zhejiang Province (Hangzhou, China), Mr. Guanping Chen in the
Laboratory of Tumor Traditional Chinese Medicine Therapy in
Institute of Cancer Research, Tongde Hospital of Zhejiang Province
and Dr. Huwei Hou in the Laboratory of Cell Molecular Biology of
Cell-Land Biological Technology Institute (Hangzhou, China) for the
help in statistical analysis and technical assistance in this
research.
Funding
This study was supported by grants from Zhejiang
Provincial Natural Science Foundation (grant no. LY13H290012) and
Zhejiang Provincial Science and Technology Program of Traditional
Chinese Medicine (grant no. 2017ZA029).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZY and PC proposed the hypothesis and designed the
majority of the experiments. ZY wrote the manuscript. ZY, QX and
ZhaC performed the experiments and analyzed the results. HN, LX, QZ
and ZhiC assisted in the execution of some experiments. QX, HN and
LX were involved in revising the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Takaoka M: Resveratrol, a new phenolic
compound, from Veratrum grandiflorum. J Chem Society Japan.
60:1090–1100. 1939.
|
|
2
|
Frémont L: Biological effects of
resveratrol. Life Sci. 66:663–673. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang Y, Catana F, Yang Y, Roderick R and
van Breemen RB: An LC-MS method for analyzing total resveratrol in
grape juice, cranberry juice, and in wine. J Agric Food Chem.
50:431–435. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Burns J, Yokota T, Ashihara H, Lean ME and
Crozier A: Plant foods and herbal sources of resveratrol. J Agric
Food Chem. 50:3337–3340. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jang M, Cai L, Udeani GO, Slowing KV,
Thomas CF, Beecher CW, Fong HH, Farnsworth NR, Kinghorn AD, Mehta
RG, et al: Cancer chemopreventive activity of resveratrol, a
natural product derived from grapes. Science. 275:218–220. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Clément MV, Hirpara JL, Chawdhury SH and
Pervaiz S: Chemopreventive agent resveratrol, a natural product
derived from grapes, triggers CD95 signaling-dependent apoptosis in
human tumor cells. Blood. 92:996–1002. 1998.PubMed/NCBI
|
|
7
|
Nakagawa H, Kiyozuka Y, Uemura Y, Senzaki
H, Shikata N, Hioki K and Tsubura A: Resveratrol inhibits human
breast cancer cell growth and may mitigate the effect of linoleic
acid, a potent breast cancer cell stimulator. J Cancer Res Clin
Oncol. 127:258–264. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Garvin S, Ollinger K and Dabrosin C:
Resveratrol induces apoptosis and inhibits angiogenesis in human
breast cancer xenografts in vivo. Cancer Lett. 231:113–122. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kalra N, Roy P, Prasad S and Shukla Y:
Resveratrol induces apoptosis involving mitochondrial pathways in
mouse skin tumorigenesis. Life Sci. 82:348–358. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shukla Y and Singh R: Resveratrol and
cellular mechanisms of cancer prevention. Ann N Y Acad Sci.
1215:1–8. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Alfaras I, Juan ME and Planas JM:
Tans-resveratrol reduces precancerous colonic lesions in
dimethylhydrazine-treated rats. J Agric Food Chem. 58:8104–8110.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin HC, Chen YF, Hsu WH, Yang CW, Kao CH
and Tsai TF: Resveratrol helps recovery from fatty liver and
protects against hepatocellular carcinoma induced by hepatitis B
virus X protein in a mouse model. Cancer Prev Res (Phila).
5:952–962. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen K, Xu XW, Zhang RC, Pan Y, Wu D and
Mou YP: Systematic review and meta-analysis of laparoscopy-assisted
and open total gastrectomy for gastric cancer. World J
Gastroenterol. 19:5365–5376. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Orditura M, Galizia G, Sforza V,
Gambardella V, Fabozzi A, Laterza MM, Andreozzi F, Ventriglia J,
Savastano B, Mabilia A, et al: Treatment of gastric cancer. World J
Gastroenterol. 20:1635–1649. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pretz JL, Wo JY, Mamon HJ, Kachnic LA and
Hong TS: Chemoradiation therapy: Localized esophageal, gastric, and
pancreatic cancer. Surg Oncol Clin N Am. 22:511–524. 2011.
View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Radisky DC: Epithelial-mesenchymal
transition. J Cell Sci. 118:4325–4326. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xia H, Chen Q, Chen Y, Ge X, Leng W, Tang
Q, Ren M, Chen L, Yuan D, Zhang Y, et al: The lncRNA MALAT1 is a
novel biomarker for gastric cancer metastasis. Oncotarget.
7:56209–56218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang J, Sun J, Wang J, Song Y, Gao P, Shi
J, Chen P and Wang Z: Long noncoding RNAs in gastric cancer:
Functions and clinical applications. Onco Targets Ther. 9:681–697.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun W, Yang Y, Xu C, Xie Y and Guo J:
Roles of long noncoding RNAs in gastric cancer and their clinical
applications. J Cancer Res Clin Oncol. 142:2231–2237. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang EB, Han L, Yin DD, Kong R, De W and
Chen J: c-Myc-induced, long, noncoding H19 affects cell
proliferation and predicts a poor prognosis in patients with
gastric cancer. Med Oncol. 31:9142014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou X, Ye F, Yin C, Zhuang Y, Yue G and
Zhang G: The interaction between MiR-141 and lncRNA-H19 in
regulating cell proliferation and migration in gastric cancer. Cell
Physiol Biochem. 36:1440–1452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang ZZ, Shen ZY, Shen YY, Zhao EH, Wang
M, Wang CJ, Cao H and Xu J: HOTAIR long noncoding RNA promotes
gastric cancer metastasis through suppression of poly r(C)-binding
protein (PCBP) 1. Mol Cancer Ther. 14:1162–1170. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang F, Xue X, Zheng L, Bi J, Zhou Y, Zhi
K, Gu Y and Fang G: Long non-coding RNA GHET1 promotes gastric
carcinoma cell proliferation by increasing c-Myc mRNA stability.
FEBS J. 281:802–813. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gutschner T, Hämmerle M, Eissmann M, Hsu
J, Kim Y, Hung G, Revenko A, Arun G, Stentrup M, Gross M, et al:
The noncoding RNA MALAT1 is a critical regulator of the metastasis
phenotype of lung cancer cells. Cancer Res. 73:1180–1189. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ji Q, Zhang L, Liu X, Zhou L, Wang W, Han
Z, Sui H, Tang Y, Wang Y, Liu N, et al: Long non-coding RNA MALAT1
promotes tumor growth and metastasis in colorectal cancer through
binding to SFPQ and releasing oncogene PTBP2 from SFPQ/PTBP2
complex. Br J Cancer. 111:736–748. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lai MC, Yang Z, Zhou L, Zhu QQ, Xie HY,
Zhang F, Wu LM, Chen LM and Zheng SS: Long non-coding RNA MALAT-1
overexpression predicts tumor recurrence of hepatocellular
carcinoma after liver transplantation. Med Oncol. 29:1810–1816.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ren S, Liu Y, Xu W, Sun Y, Lu J, Wang F,
Wei M, Shen J, Hou J, Gao X, et al: Long noncoding RNA MALAT-1 is a
new potential therapeutic target for castration resistant prostate
cancer. J Urol. 190:2278–2287. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schmidt LH, Spieker T, Koschmieder S,
Schäffers S, Humberg J, Jungen D, Bulk E, Hascher A, Wittmer D,
Marra A, et al: The long noncoding MALAT-1 RNA indicates a poor
prognosis in non-small cell lung cancer and induces migration and
tumor growth. J Thorac Oncol. 6:1984–1992. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu XS, Wang XA, Wu WG, Hu YP, Li ML, Ding
Q, Weng H, Shu YJ, Liu TY, Jiang L, et al: MALAT1 promotes the
proliferation and metastasis of gallbladder cancer cells by
activating the ERK/MAPK pathway. Cancer Biol Ther. 15:806–814.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ying L, Chen Q, Wang Y, Zhou Z, Huang Y
and Qiu F: Upregulated MALAT-1 contributes to bladder cancer cell
migration by inducing epithelial-to-mesenchymal transition. Mol
Biosyst. 8:2289–2294. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ji Q, Liu X, Fu X, Zhang L, Sui H, Zhou L,
Sun J, Cai J, Qin J, Ren J and Li Q: Resveratrol inhibits invasion
and metastasis of colorectal cancer cells via MALAT1 mediated
Wnt/β-catenin signal pathway. PLoS One. 8:e787002013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang HM, Yang FQ, Chen SJ, Che J and
Zheng JH: Upregulation of long non-coding RNA MALAT1 correlates
with tumor progression and poor prognosis in clear cell renal cell
carcinoma. Tumour Biol. 36:2947–2955. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pang EJ, Yang R, Fu XB and Liu YF:
Overexpression of long non-coding RNA MALAT1 is correlated with
clinical progression and unfavorable prognosis in pancreatic
cancer. Tumour Biol. 36:2403–2407. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Okugawa Y, Toiyama Y, Hur K, Toden S,
Saigusa S, Tanaka K, Inoue Y, Mohri Y, Kusunoki M, Boland CR and
Goel A: Metastasis-associated long non-coding RNA drives gastric
cancer development and promotes peritoneal metastasis.
Carcinogenesis. 35:2731–2739. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang J, Su L, Chen X, Li P, Cai Q, Yu B,
Liu B, Wu W and Zhu Z: MALAT1 promotes cell proliferation in
gastric cancer by recruiting SF2/ASF. Biomed Pharmacother.
68:557–564. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li Y, Wu Z, Yuan J, Sun L, Lin L, Huang N,
Bin J, Liao Y and Liao W: Long non-coding RNA MALAT1 promotes
gastric cancer tumorigenicity and metastasis by regulating
vasculogenic mimicry and angiogenesis. Cancer Lett. 395:31–44.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Acloque H, Adams MS, Fishwick K,
Bronner-Fraser M and Nieto MA: Epithelial-mesenchymal transitions:
The importance of changing cell state in development and disease. J
Clin Invest. 119:1438–1449. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liang J, Liang L, Ouyang K, Li Z and Yi X:
MALAT1 induces tongue cancer cells' EMT and inhibits apoptosis
through Wnt/β-catenin signaling pathway. J Oral Pathol Med.
46:98–105. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu S, Song L, Yao H, Zhang L, Xu D, Gao F
and Li Q: MiR-375 is epigenetically downregulated by HPV-16 E6
mediated DNMT1 upregulation and modulates EMT of cervical cancer
cells by suppressing lncRNA MALAT1. PLoS One. 11:e01634602016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li J, Wang J, Chen Y, Li S, Jin M, Wang H,
Chen Z and Yu W: LncRNA MALAT1 exerts oncogenic functions in lung
adenocarcinoma by targeting miR-204. Am J Cancer Res. 6:1099–1107.
2016.PubMed/NCBI
|
|
45
|
Zhou X, Liu S, Cai G, Kong L, Zhang T, Ren
Y, Wu Y, Mei M, Zhang L and Wang X: Long non coding RNA MALAT1
promotes tumor growth and metastasis by inducing
epithelial-mesenchymal transition in oral squamous cell carcinoma.
Sci Rep. 5:159722015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kundu JK and Surh YJ: Cancer
chemopreventive and therapeutic potential of resveratrol:
Mechanistic perspectives. Cancer Lett. 269:243–261. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Qadri SM, Föller M and Lang F: Inhibition
of suicidal erythrocyte death by resveratrol. Life Sci. 85:33–38.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bisht K, Wagner KH and Bulmer AC:
Curcumin, resveratrol and flavonoids as anti-inflammatory, cyto-
and DNA-protective dietary compounds. Toxicology. 278:88–100. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gao Q, Yuan Y, Gan H and Peng Q:
Resveratrol inhibits the hedgehog signaling pathway and
epithelial-mesenchymal transition and suppresses gastric cancer
invasion and metastasis. Oncol Lett. 9:2381–2387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Jing X, Cheng W, Wang S, Li P and He L:
Resveratrol induces cell cycle arrest in human gastric cancer
MGC803 cells via the PTEN-regulated PI3K/Akt signaling pathway.
Oncol Rep. 35:472–478. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang Q, Wang B, Zang W, Wang X, Liu Z, Li
W and Jia J: Resveratrol inhibits the growth of gastric cancer by
inducing G1 phase arrest and senescence in a Sirt1-dependent
manner. PLoS One. 8:e706272013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sheth S, Jajoo S, Kaur T, Mukherjea D,
Sheehan K, Rybak LP and Ramkumar V: Resveratrol reduces prostate
cancer growth and metastasis by inhibiting the Akt/MicroRNA-21
pathway. PLoS One. 7:e516552012. View Article : Google Scholar : PubMed/NCBI
|