Introduction
In developed countries, cardiovascular disease is
the primary cause of mortality and disability worldwide, and its
incidence is increasing. Among these, coronary artery disease (CAD)
has an important role and is a frequent cause of death. Numerous
environmental factors, including obesity and hyperlipemia, have
been confirmed to lead to the development of CAD (1). However, it remains a challenge to fully
elucidate the pathogenesis of CAD and determine risk factors
(1,2).
Identification of individuals at risk and the rapid
and accurate diagnosis of CAD are warranted to further enhance the
success of preventative or therapeutic procedures. A decrease in
the morbidity of CAD and the associated mortality may be attributed
to primary and secondary care and prevention programs (3).
Certain studies have indicated an association
between genetic variants and the occurrence of CAD (4,5). S100
calcium binding protein B (S100B), a ligand for the receptor for
advanced glycation end products (RAGE), has a crucial role in the
development and progression of atherosclerosis (6–8). RAGE
interacting with its ligand, S100B, induces lymphocytes to enter
atherosclerotic plaque lesions through the damaged endothelial
barrier, leading to the accumulation of inflammatory cells in
coronary artery atherosclerotic plaques, thus contributing to the
development of atherosclerosis. Furthermore, enhanced RAGE
expression has been observed in atherosclerotic lesions from
apolipoprotein E-deficient mice (9–12). It
has also been reported that variants of RAGE and its ligand are
associated with susceptibility to type 2 diabetes (7,9,13). Therefore, it is vital to elucidate
the functions of RAGE and thereby enhance the current understanding
of the pathogenesis of CAD.
To the best of our knowledge, only few studies have
reported an association between RAGE variants and the occurrence of
CAD in the Han Chinese population. Therefore, the association
between RAGE tag single nucleotide polymorphisms (tagSNPs) and the
risk of cardiovascular disease was explored in the present study.
The present study aimed to increase the understanding of the effect
of RAGE polymorphisms on atherogenesis in subjects with CAD.
Patients and methods
Study participants
A total of 1,719 Han Chinese subjects who were
treated at Shenyang Military General Hospital (Shenyang, China)
between July 2014 and August 2016 were included in the present
study. These included 852 patients with CAD and 867 control
subjects without any cardiovascular disease. In these participants,
the presence of conventional cardiovascular risk factors according
to the Guidelines for Assessment and Management of cardiovascular
risk from 2007 (14) were determined
by the treating doctor using a standardized interview, which
included the history of smoking, hypertension and diabetes, to
assess the patient's symptoms. The body weight (kg) divided by
height (m)2 was used to determine the body mass index.
The conventional criteria for diagnosis of hypertension and type 2
diabetes mellitus were based on the World Health Organization
guidelines. Hypertension was diagnosed according to either a
diastolic pressure of ≥90 mmHg or arterial blood pressure ≥140 mmHg
according to the Seventh Report of the Seventh Report of the Joint
National Committee on Prevention, Detection, Evaluation, and
Treatment of High Blood Pressure (15). All participants underwent coronary
angiography at the Shenyang Military General Hospital to evaluate
the suspected or established CAD. Patients with CAD recruited for
the present study were those who had undergone coronary angiography
after being diagnosed with CAD based on an angiography examination
(>70% stenosis affecting at least one coronary vessel). Among
the 852 CAD patients, they were the patients with mixed acute
coronary syndrome. The participants with a normal coronary
angiogram or major coronary artery stenosis of ≤20% according the
results of coronary arteriography were defined as the control
group. Furthermore, the control group did not have a history of CAD
or ultrasonic cardiogram abnormalities (13). The patients with CAD were a mixed
group of patients with acute coronary syndrome (ACS) and stable CAD
patients. ACS based on the pathological basis of rupture or
invasion of atherosclerotic plaque and subsequent complete or
incomplete occlusive thrombosis included the acute T-segment
elevation myocardial infarction, acute non-ST-segment elevation
myocardial infarction and unstable angina (16). Cardiac syndrome X, also known as
microvascular angina, is defined as angina pectoris caused by
abnormalities of the small coronary arteries, and is characterized
by the patient experiencing chest pain during exercise and evidence
of myocardial ischemia with a non-invasive stress test (17). With these patients the coronary
angiography can appear normal (18).
This is usually diagnosed based on chest pain typical for angina,
exercise-induced significant ST-segment depression and normal
coronary angiograms (19). Syndrome
X is diagnosed based on chest pain, exercise-induced ST-segment
depression and the absence of either significant epicardial
coronary stenosis or vasospasm (20). Certain patients with syndrome X
exhibit pathologic changes around microvessels themselves (21). As control subjects, those patients
with a normal coronary angiogram and suspected syndrome X were
subjected to the treadmill exercise test. If the test was negative,
CAD was excluded and the subjects were enrolled in the control
group.
Patients with active inflammatory disease,
autoimmune disease, severe heart failure, hemodynamic instability,
suspected myocarditis or pericarditis, diseases of the
hematopoietic system, extension of kidney or liver disease,
malignant disease, intake of immunosuppressive drugs and renal or
hepatic diseases were excluded from this study.
Prior to any invasive procedures or medical
therapies, peripheral blood samples of the control and CAD patients
were drawn at the time-point of admission to the hospital. The
blood was collected in pyrogen-free EDTA-containing tubes, and
after isolation, the plasma samples were immediately paced on ice
and then stored at −80°C.
Selection of tagSNPs
The Tiangen DNA extraction kit (Beijing, China) was
used to extract DNA from peripheral blood. The Han population
Hapmap data (http://www.hapmap.org) was used to
identify TagSNPs that covered the entire region of the RAGE gene,
from 10,000 bp upstream to 4,000 bp downstream (22). Lewontin's D′ statistics and
r2 correlation statistics were used to assess the
pairwise linkage disequilibrium difference. The variants with a
frequency at a cut-off of 0.05 and an r2 threshold of
0.8 were further investigated. A total of 25 tagSNPs of RAGE
(rs10754558, rs10925025, rs12048215, rs12143966, rs12564791,
rs1800624, rs2027432, rs204993, rs204994, rs2071290, rs2269421,
rs2269422, rs2269423, rs2282659, rs3006476, rs3014880, rs3014885,
rs3130349, rs3134943, rs3134946, rs3738448, rs4772, rs4925648,
rs7528887 and rs7529058) were selected from different haplotype
regions.
Genotyping
The 25 tagSNPs of the RAGE gene in samples from the
CAD and control patients were analyzed by matrix-assisted laser
desorption time of flight (MALDI-TOF) mass spectrometry. Polymerase
chain reaction (PCR) was generally adopted (23); however, the phosphorylation state was
also analyzed using shrimp alkaline phosphatase (Hoffman-La Roche,
Basel, Switzerland) (24). All
analyses were performed using a silicon chip made of a 384-well
porous plate. PCR products were put onto a MALDI matrix and were
analyzed using the Bruker Biflex III MALDI-TOF SpectroReader mass
spectrometer (Bruker Corp., Billerica, MA, USA) (24). The mass spectra were analyzed using
SpectroTyper software (Sequenom, Inc., San Diego, CA, USA).
Collection of carotid artery
tissues
A total of 10 carotid and coronary arteries, with
and without atherosclerotic lesions, were collected from eight
donors. Atherosclerotic carotid and coronary arteries were acquired
from four patients who died of atherosclerotic heart disease during
autopsy. Normal carotid and coronary arteries were acquired from
four subjects that were healthy prior to death during autopsy. The
donors' relatives provided written informed consent.
Immunohistochemistry and
immunofluorescence
H&E staining and Masson staining was performed
to observe the morphological features of the carotid tissues
collected. Immunohistochemistry and immunofluorescence were
performed to determine RAGE and CD68 expression in the carotid
tissue. In brief, carotid arteries (formalin-fixed,
paraffin-embedded) were serially sectioned (4 µm; ≥3 sections per
sample). Subsequently, anti-RAGE (cat. no. ab37647; Abcam,
Cambridge, MA, USA) and anti-CD68 (cat no. GMO81404; Dako,
Glostrup, Denmark) antibodies were used to analyze protein
expression in the serial sections. The samples were incubated with
the primary antibodies (dilution, 1:100) at 4°C overnight, the
unbound antibodies were then washed away. Goat anti-rabbit Alexa
Fluor 488 (cat. no. 11034) and donkey anti-mouse Alexa Fluor 555
secondary antibodies (cat. no. 31570; both; 1:1,000; Invitrogen;
Thermo Fisher Scientific, Inc.) were incubated at room temperature
for 2 h to detect the bound primary antibodies.
Avidin-biotin-peroxidase was incubated at room temperature for 15
sec to detect the bound antibodies (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). Staining with 3,3′-diaminobenzidine (cat. no.
E2886; Sigma-Aldrich; Merck KGaA) was performed for 2 h at 4°C to
detect immunoreactivity. Images were obtained using a light
microscope and a confocal laser scanning microscope (FV500;
Olympus, Tokyo, Japan).
Semi-quantitative reverse
transcription (RT)-PCR analysis of RAGE
RNA from the coronary artery tissues from the CAD
and control patients was isolated using the Eastep™
total RNA extraction kit (cat. no. LS1030; Promega Corp., Madison,
WI, USA). The RNA from the CAD and control groups was
reverse-transcribed using the First Strand complementary (c)DNA
Synthesis kit (Takara Bio, Inc., Otsu, Japan) according to the
manufacturer's protocol. A PCR analysis was then used to amplify
the cDNA of RAGE using the Superscript II kit (Takara Bio, Inc.)
with Taq polymerase and the primers listed in Table I, which also states the thermocycling
conditions and product lengths. The amplified cDNA was resolved
using 2% agarose gel electrophoresis and visualized with ethidium
bromide. ImageJ (version 1.8.0; National Institutes of Health,
Bethesda, MD, USA) software was used to analyze the results as
previous described (25).
 | Table I.Primer sequences used for the
genotyping analysis of RAGE and GAPDH variations. |
Table I.
Primer sequences used for the
genotyping analysis of RAGE and GAPDH variations.
| Name | Primer sequence
(5′-3′) | Size of PCR product
(bp) | Thermocycling
protocol |
|---|
| RAGE | F:
AGGTGAGTGGAGAAAGCCAG | 121 | 94°C for 3 min; |
|
| R:
ATGTGTCAGGTGTTTAATCA |
| 35 cycles of 94°C for
30 sec, 56°C for |
|
|
|
| 30 sec and 72°C for
30 sec; |
|
|
|
| 72°C for 7 min; |
|
|
|
| 4°C ∞ |
|
|
|
| 94°C for 3 min; |
| GAPDH | F:
AGGATGGTGTGGCTCCCTTG | 105 | 35 cycles of 94°C for
30 sec, 58°C for |
|
| R:
GCAGGGCTGAGACAGCTTCC |
| 25 sec and 72°C for
25 sec; |
|
|
|
| 72°C for 7 min; |
|
|
|
| 4°C ∞ |
Western blot analysis
Tissues were lysed in lysis buffer containing
freshly added protease inhibitor (cat. no. 78425; Invitrogen;
Thermo Fisher Scientific, Inc.). After centrifugation at 12,000 × g
for 10 min at 4°C, the protein in the supernatant was subjected to
western blot analysis. Protein concentrations were determined using
the bicinchoninic acid protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.). Proteins were resolved by 10% SDS-PAGE and
transferred to polyvinylidene difluoride membranes. After blocking
with 5% skimmed milk, the membranes were incubated with anti-RAGE
(1:1,000 dilution; cat. no. ab37647) and anti-GAPDH antibody
(1:1,000 dilution; cat no. ab9484; both Abcam) overnight at 4°C.
Specific binding was detected by subsequent incubation with
horseradish peroxidase-conjugated secondary antibodies at 4°C for 2
h and an enhanced chemiluminescence kit (cat. no. NCI4106; Pierce;
Thermo Fisher Scientific, Inc.). The blots were quantified using a
Bio-Rad Gel system (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
ELISA
The plasma levels of S100B in CAD and control
patients were assessed using S100B ELISA kits (cat. no. DY1820-05;
R&D Systems, Minneapolis, MN, USA). The absorbance of each well
was detected at 450 nm and the plasma concentration of S100B was
determined using a standard curve according to the manufacturer's
protocol. When determining whether the S100B plasma levels were
associated with the genotype of RAGE rs2269422, the rare AA
homozygote carriers were grouped together with the AT heterozygotes
due to the small number of AA homozygotes present in the control
and CAD groups (26).
Statistical analysis
Values are expressed as the mean ± standard error of
the mean and were analyzed using SPSS software version 21.0 (IBM
Corp., Armonk, NY, USA). Student's t-test or the Mann-Whitney
U-test was used to compare among the different groups, and multiple
comparisons between groups were assessed by one-way analysis of
variance (ANOVA) followed by Tukey's Honest Significant Difference
post-hoc test analysis. The difference in the plasma levels of
S100B between CAD and control groups were also analyzed by ANOVA.
The distribution of genotype and allele frequencies between the CAD
and control groups were analyzed by determining the Hardy-Weinberg
equilibrium. The CAD risk was evaluated by the P-values, 95%
confidence intervals (95% CIs) and odds ratios (ORs). The
Bonferroni adjustment method was used for multiple comparisons
among the 25 tagSNPs and a significance level of <0.002
(0.05/25=0.002) was required.
Results
Clinical characteristics of the study
participants
The demographic data of the study population,
including 852 CAD and 867 control subjects, are presented in
Table II. In the control group, the
distribution of genotypes was in a Hardy-Weinberg equilibrium. The
CAD group was characterized by a higher prevalence of traditional
atherosclerotic risk factors compared with that in the control
group. The conventional vascular risk factors, including cigarette
smoking, hypertension and diabetes mellitus had a higher prevalence
in patients with CAD compared with the control subjects
(P<0.05). In addition, increased levels of triglycerides, as
well as low-density and very low-density lipoprotein-cholesterol,
and lower levels of high-density lipoprotein-cholesterol were
detected in the CAD patients compared with those in the control
subjects (Table II).
| Table II.Clinical Characteristics of CAD
patients and control subjects. |
Table II.
Clinical Characteristics of CAD
patients and control subjects.
| Characteristic | Control group
(n=867) | CAD group
(n=852) | P-value |
|---|
| Age (years) | 58.3±7.9 | 59.8±8.2 | 0.001 |
| Sex (M/F) | 477/390 | 440/412 | 0.161 |
| BMI
(kg/m2) | 24.8±4.7 | 25.2±4.2 | 0.001 |
| DM | 104 (12.0) | 266 (31.2) | P<0.001 |
| Hypertension | 432(49.8) | 514 (60.3) | P<0.001 |
| Smoking | 268 (30.9) | 369 (43.3) | P<0.001 |
| TG (mmol/l) | 1.74±1.38 | 2.41±1.48 | P<0.001 |
| TC (mmol/l) | 4.07±1.03 | 4.79±1.14 | P<0.001 |
| LDL-C (mmol/l) | 1.92±0.62 | 2.61±0.81 | P<0.001 |
| HDL-C (mmol/l) | 1.81±0.39 | 1.29±0.35 | P<0.001 |
Association of tagSNPs with the risk
of CAD
All study participants were genotyped for the RAGE
tagSNP rs2269422. The primary data for the tagSNPs are listed in
Table III. The frequency
distribution of the 25 tagSNPs in the controls was in accordance
with the Hardy-Weinberg equilibrium (P≥0.05). Table III presents the genotype and allele
frequencies for the 25 tagSNPs of RAGE in the cases and controls.
It was revealed that the A allele of the RAGE SNP rs2269422 was
associated with an enhanced risk of CAD (P<0.001, OR=0.505, 95%
CI, 0.409–0.625) in a Han Chinese population. Thus, it was
indicated that this tagSNP of RAGE is closely associated with the
risk of CAD in this population. Of note, the allele or genotype
frequencies for the remaining tagSNPs of RAGE were not
significantly different between the CAD and the control subjects
(Table III).
| Table III.Genotypic and allelic frequencies of
receptor for advanced glycation end products polymorphisms in the
control (n=867) and CAD group (n=852). |
Table III.
Genotypic and allelic frequencies of
receptor for advanced glycation end products polymorphisms in the
control (n=867) and CAD group (n=852).
| Polymorphism |
Genotypes/alleles | Control | Patients with
CAD | P-value | OR (95% CI) |
|---|
| Rs10754558 | Genotypes |
|
|
|
|
|
| CC | 229 (26.4) | 265 (31.1) |
| 1.00 |
|
| CG | 459 (52.9) | 439 (51.5) | 0.089 | 0.826
(0.663–1.030) |
|
| GG | 179 (20.6) | 148 (17.4) | 0.019 | 0.714
(0.540–0.946) |
|
| Alleles |
|
|
|
|
|
| C | 917 (52.9) | 969 (56.9) |
| 1.00 |
|
| G | 817 (47.1) | 735 (43.1) | 0.019 | 0.851
(0.744–0.974) |
| Rs10925025 | Genotypes |
|
|
|
|
|
| AA | 156 (18.0) | 156 (18.3) |
| 1.00 |
|
| AG | 455 (52.5) | 405 (47.5) | 0.379 | 0.890
(0687–1.153) |
|
| GG | 256 (29.5) | 291 (34.2) | 0.367 | 1.137
(0.861–1.501) |
|
| Alleles |
|
|
|
|
|
| A | 767 (44.2) | 717 (42.1) |
| 1.00 |
|
| G | 967 (55.8) | 987 (57.9) | 0.202 | 1.091
(0.953–1.249) |
| Rs12048215 | Genotypes |
|
|
|
|
|
| AA | 477 (54.9) | 456 (53.5) |
| 1.00 |
|
| AG | 344 (39.6) | 354 (41.5) | 0.462 | 1.076
(0.885–1.310) |
|
| GG | 46 (5.5) | 42 (4.9) | 0.955 | 0.955
(0.617–1.479) |
|
| Alleles |
|
|
|
|
|
| A | 1,298 (74.9) | 1,266 (74.3) | 0.019 | 0.844
(0.733–0.973) |
|
| G | 436 (25.1) | 438 (25.7) |
|
|
| Rs12143966 | Genotypes |
|
|
|
|
|
| AA | 177 (20.4) | 201 (23.6) |
| 1.00 |
|
| AG | 204 (23.5) | 211 (24.7) | 0.837 | 0.955
(0.617–1.479) |
|
| GG | 486 (56.1) | 440 (51.6) | 0.064 | 0.797
(0.627–1.013) |
|
| Alleles |
|
|
|
|
|
| A | 558 (32.2) | 613 (35.9) |
| 1.00 |
|
| G | 1,176 (67.8) | 1,091 (64.0) | 0.091 | 0.827
(0.718–0.953) |
| Rs12564791 | Genotypes |
|
|
|
|
|
| CC | 633 (73.0) | 639 (75.0) |
| 1.00 |
|
| CT | 222 (25.6) | 189 (22.2) | 0.134 | 0.843
(0.675–11.054 |
|
| TT | 12 (1.4) | 24 (2.8) | 0.052 | 1.981
(0.982–3.996) |
|
| Alleles |
|
|
|
|
|
| C | 1,488 (85.8) | 1,467 (86.1) | 0.814 | 0.977
(0.806–1.185) |
|
| T | 246 (14.2) | 237 (13.9) |
|
|
| Rs1800624 | Genotypes |
|
|
|
|
|
| AA | 588 (67.8) | 594 (69.7) |
| 1.00 |
|
| AT | 261 (30.1) | 234 (27.5) | 0.265 | 0.887
(0.719–1.095) |
|
| TT | 18 (2.1) | 24 (2.8) | 0.380 | 1.320
(0.709–2.456) |
|
| Alleles |
|
|
|
|
|
| A | 1,437 (82.9) | 1,422 (83.5) | 0.682 | 0.960
(0.802–1.147) |
|
| T | 297 (17.1) | 237 (16.5) |
|
|
| Rs2027432 | Genotypes |
|
|
|
|
|
| GG | 747 (86.2) | 732 (85.9) |
| 1.00 |
|
| AG | 117 (13.5) | 111 (13.0) | 0.820 | 0.968
(0.732–1.280) |
|
| AA | 3 (0.3) | 9 (1.1) | 0.078 | 3.061
(0.826–11.353) |
|
| Alleles |
|
|
|
|
|
| A | 123 (7.1) | 129 (7.6) |
|
|
|
| G | 1,611 (92.9) | 1,575 (92.4) | 0.601 | 0.932
(0.721–1.205) |
| Rs204993 | Genotypes |
|
|
|
|
|
| AA | 312 (36.0) | 294 (34.5) |
| 1.00 |
|
| AG | 435 (50.2) | 408 (47.9) | 0.965 | 0.995
(0.808–1.227) |
|
| GG | 120 (13.8) | 150 (17.6) | 0.054 | 1.327
(0.994–1.769) |
|
| Alleles |
|
|
|
|
|
| A | 123 (7.1) | 129 (7.6) |
| 1.00 |
|
| G | 1,611 (92.9) | 1,575 (92.4) | 0.118 | 1.115
(0.973–1.278) |
| Rs204994 | Genotypes |
|
|
|
|
|
| CC | 554 (63.9) | 528 (62.0) |
| 1.00 |
|
| CT | 289 (33.3) | 288 (33.8) | 0.665 | 1.046
(0.854–1.280) |
|
| TT | 24 (2.8) | 36 (4.2) | 0.091 | 1.574
(0.926–2.674) |
|
| Alleles |
|
|
|
|
|
| C | 1,397 (80.5) | 1,344 (78.9) |
| 1.00 |
|
| T | 337 (19.4) | 360 (21.1) | 0.710 | 1.110
(0.637–1.322) |
| Rs2269421 | Genotypes |
|
|
|
|
|
| CC | 809 (93.2) | 816 (95.8) |
| 1.00 |
|
| CG | 55 (6.5) | 33 (3.8) | 0.020 | 0.595
(0.382–0.926) |
|
| GG | 3 (0.3) | 3 (0.4) | 0.990 | 0.991
(0.200–4.927) |
|
| Alleles |
|
|
|
|
|
| C | 1,674 (96.4) | 1,685 (97.7) |
| 1.00 |
|
| T | 62 (3.6) | 39 (2.3) | 0.026 | 0.625
(0.416–0.938) |
| Rs2071290 | Genotypes |
|
|
|
|
|
| CC | 804 (92.7) | 798 (93.7) |
|
1.00 |
|
| CT | 63 (7.3) | 51 (6.0) | 0.295 | 0.816
(0.557–1.195) |
|
| TT | 0 (0.0) | 3 (0.4) | 0.082 | 1.004
(1.000–1.008) |
|
| Alleles |
|
|
|
|
|
| C | 1,674 (96.4) | 1,685 (97.7) |
| 1.00 |
|
| G | 62 (3.6) | 39 (2.3) | 0.710 | 0.918
(0.637–1.322) |
| Rs2269422 | Genotypes |
|
|
|
|
|
| TT | 723 (83.4) | 598 (70.2) |
|
1.00 |
|
| AT | 138 (15.9) | 239 (28.1) | <0.001 | 2.094
(1.654–2.651 |
|
| AA | 6 (0.7) | 15 (1.8) | 0.017 | 3.023
(1.166–7.838) |
|
| Alleles |
|
|
|
|
|
| A | 150 (8.7) | 269 (15.8) | <0.001 | 0.505
(0.409–0.625) |
|
| T | 1,584 (91.3) | 1,435 (84.2) |
|
|
| Rs2269423 | Genotypes |
|
|
|
|
|
| AA | 78 (9.0) | 81 (9.5) |
| 1.00 |
|
| AC | 351 (40.5) | 363 (42.6) | 0.981 | 0.996
(0.706–1.404) |
|
| CC | 438 (50.5) | 408 (47.9) | 0.530 | 0.897
(0.639–1.259) |
|
| Alleles |
|
|
|
|
|
| A | 507 (29.2) | 525 (30.8) |
| 1.00 |
|
| T | 1,277 (70.8) | 1,179 (69.2) | 0.315 | 0.928
(0.802–1.074) |
| Rs300647 | Genotypes |
|
|
|
|
|
| CC | 693 (79.9) | 687 (80.6) |
| 1.00 |
|
| AC | 168 (19.4) | 165 (19.4) | 0.016 | 0.991
(0.780–1.259 |
|
| AA | 6 (0.7) | 0 (0.0) | 0.015 | 0.991
(0.985–0.998) |
|
| Alleles |
|
|
|
|
|
| A | 180 (79.8) | 165 (9.7) |
| 1.00 |
|
| C | 1,554 (20.2) | 1,539 (90.3) | 1.080 | 0.532
(0.799–1.251) |
| Rs2282659 | Genotypes |
|
|
|
|
|
| AA | 552 (63.7) | 514 (60.3) |
| 1.00 |
|
| AG | 279 (32.2) | 301 (35.3) | 0.154 | 1.159
(0.949–1.419) |
|
| GG | 36 (4.2) | 37 (4.3) | 0.683 | 1.104
(0.687–1.774) |
|
| Alleles |
|
|
|
|
|
| A | 1,383 (79.8) | 1,329 (78.0) |
| 1.00 |
|
| G | 351 (20.2) | 375 (22.0) | 0.210 | 1.113
(0.944–1.311) |
| Rs3014880 | Genotypes |
|
|
|
|
|
| CC | 702 (81.0) | 684 (80.3) |
| 1.00 |
|
| CG | 159 (18.3) | 168 (19.7) | 0.510 | 1.084
(0.852–1.380) |
|
| GG | 6 (0.7) | 0 (0.0) | 0.016 | 0.992
(0.985–0.998) |
|
| Alleles |
|
|
|
|
|
| C | 1,563 (90.1) | 1,329 (78.0) |
| 1.00 |
|
| G | 171 (9.9) | 375 (22.0) | 0.042 | 1.000
(0.799–1251) |
| Rs3014885 | Genotypes |
|
|
|
|
|
| CC | 690 (79.6) | 681 (79.9) |
|
1.00 |
|
| CT | 171 (19.7) | 171 (20.1) | 0.914 | 1.013
(0.799–1,284) |
|
| TT | 6 (0.7) | 0 (0.0) | 0.015 | 0.991
(0.985–0.998) |
|
| Alleles |
|
|
|
|
|
| C | 1,551 (89.4) | 1,533 (90.0) |
| 1.00 |
|
| T | 183 (10.6) | 171 (10.0) | 0.998 | 0.933
(0.749–1.163) |
| Rs3130349 | Genotypes |
|
|
|
|
|
| TT | 645 (74.4) | 624 (73.2) |
| 1.00 |
|
| CT | 216 (24.9) | 213 (25.0) | 0.864 | 1.019
(0.890–1.269) |
|
| CC | 6 (0.7) | 15 (1.8) | 0.043 | 2.584
(0.996–6.073) |
|
| Alleles |
|
|
|
|
|
| C | 228 (13.1) | 243 (14.3) |
| 1.00 |
|
| T | 1,506 (86.9) | 1,461 (85.7) | 0.129 | 0.910
(0.749–1.106) |
| Rs3134943 | Genotypes |
|
|
|
|
|
| CC | 699 (80.6) | 663 (77.8) |
| 1.00 |
|
| CT | 153(17.6) | 171 (20.1) | 0.185 | 1.178
(0.924–1.502) |
|
| TT | 15 (1.7) | 18 (20.1) | 0.505 | 1.265
(0.632–0.531) |
|
| Alleles |
|
|
|
|
|
| C | 1,551 (89.4) | 1,497 (87.9) |
| 1.00 |
|
| T | 183 (10.6) | 207 (12.1) | 0.346 | 1.172
(0.949–1.448) |
| Rs3134946 | Genotypes |
|
|
|
|
|
| CC | 684 (78.9) | 663 (77.8) |
| 1.00 |
|
| CG | 177 (20.4) | 171 (20.1) | 0.978 | 0.997
(0.787–1.262) |
|
| GG | 6 (0.7) | 18 (20.1) | 0.012 | 3.095
(1.221–7.885) |
|
| Alleles |
|
|
|
|
|
| C | 1,545 (89.4) | 1,505 (88.3) |
| 1.00 |
|
| G | 189 (10.6) | 199 (11.7) | 0.484 | 1.081
(0.875–1.335) |
| Rs3738448 | Genotypes |
|
|
|
|
|
| GG | 558 (64.5) | 574 (67.4) |
| 1.00 |
|
| CT | 264 (30.4) | 241 (28.3) | 0.265 | 0.887
(0.719–1.095) |
|
| TT | 45 (5.2) | 37 (4.3) | 0.329 | 0.799
(0.570–1.254) |
|
| Alleles |
|
|
|
|
|
| G | 1,380 (79.6) | 1,389 (81.5) | 0.155 | 0.884
(0.747–1.047) |
|
| T | 354 (20.4) | 315 (18.5) |
|
|
| Rs4772 | Genotypes |
|
|
|
|
|
| AA | 693 (79.9) | 690 (81.0) |
| 1.00 |
|
| G | 168 (19.4) | 159 (18.7) | 0.680 | 0.951
(0.747–1.210) |
|
| GG | 6 (0.7) | 3 (0.3) | 0.322 | 0.502
(0.125–2.016) |
|
| Alleles |
|
|
|
|
|
| A | 1,544 (89.6) | 1,539 (90.3) |
| 1.00 |
|
| G | 180 (10.4) | 165 (9.7) | 0.532 | 1.767
(1.452–2.150) |
| Rs4925648 | Genotypes |
|
|
|
|
|
| CC | 576 (64.4) | 570 (66.9) |
| 1.00 |
|
| CT | 267 (30.8) | 246 (28.9) | 0.501 | 0.931
(0.756–1.147) |
|
| TT | 24 (2.8) | 36 (4.2) | 0.121 | 1.516
(0.893–2.573) |
|
| Alleles |
|
|
|
|
|
| A | 1,419 (81.8) | 1,386 (81.3) |
| 1.00 |
|
| T | 315 (18.2) | 318 (18.7) | 0.708 | 1.034
(0.870–1.228) |
| Rs7528887 | Genotypes |
|
|
|
|
|
| AA | 465 (53.6) | 450 (52.8) |
| 1.00 |
|
| AG | 330 (38.0) | 351 (41.2) | 0.351 | 1.099
(0.901–1.340) |
|
| GG | 72 (8.3) | 51 (6.0) | 0.108 | 0.732
(0.500–1.072) |
|
| Alleles |
|
|
|
|
|
| A | 1,260 (72.7) | 1,251 (73.4) |
| 1.00 |
|
| G | 474 (27.3) | 453 (26.6) | 0.963 | 0.963
(0.828–1.119) |
| Rs7529058 | Genotypes |
|
|
|
|
|
| CC | 234 (27.0) | 228 (26.8) |
| 1.00 |
|
| CG | 407 (46.9) | 390 (45.8) | 0.887 | 0.983
(0.782–1.237) |
|
| GG | 226 (26.1) | 234 (27.5) | 0.645 | 1.063
(0.821–1.376) |
|
| Alleles |
|
|
|
|
|
| C | 875 (50.5) | 846 (49.6) |
| 1.00 |
|
| G | 859 (49.5) | 858 (50.4) | 0.657 | 1.033
(0.904–1.181) |
RAGE expression in atherosclerotic
carotid arteries
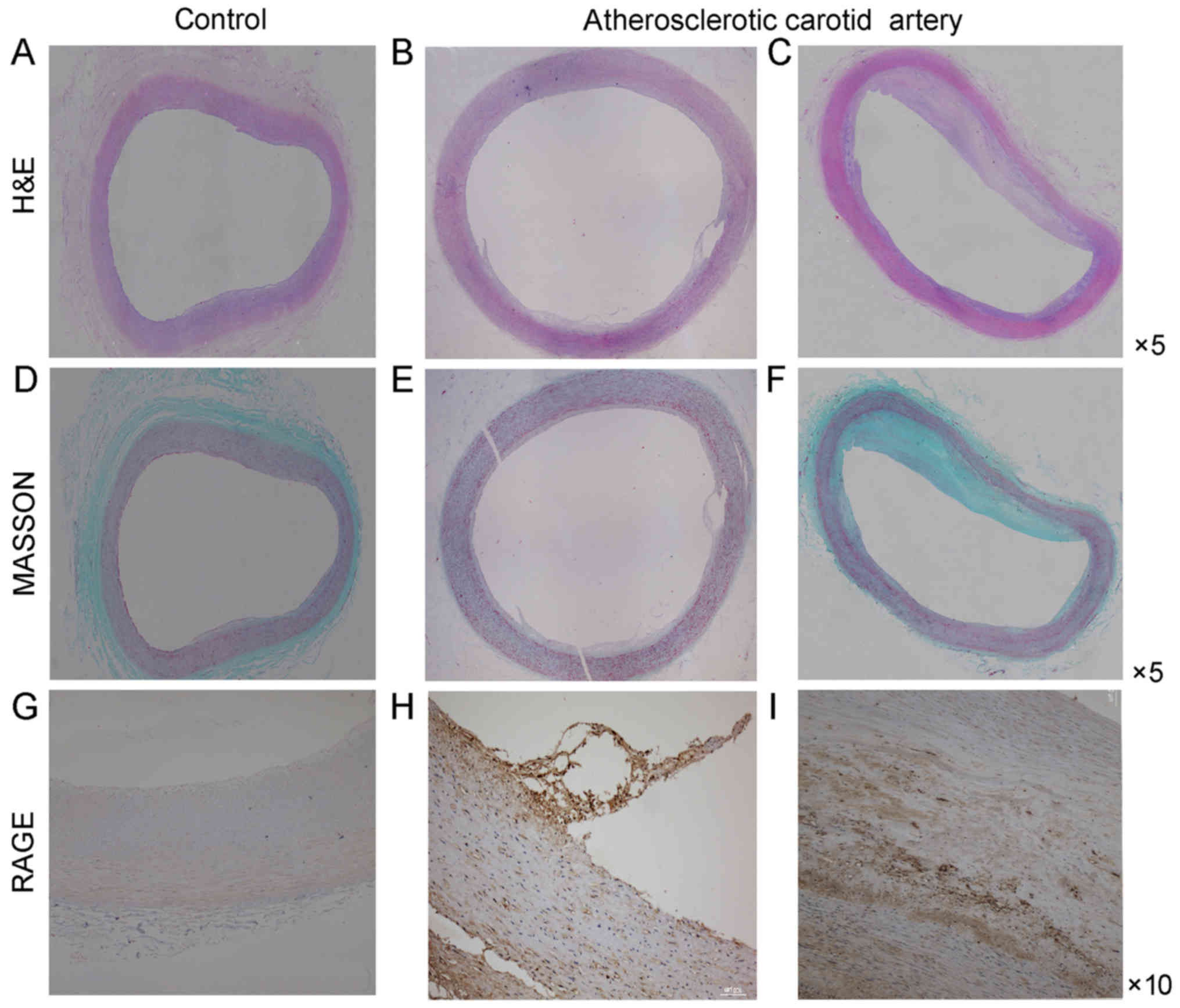
Sections stained with H&E and Masson
demonstrated the basic morphological features of human carotid
arteries (Fig. 1). The basic
morphology of normal carotid artery and the morphologic change of
atherosclerotic carotid artery were revealed. Endothelial cells in
normal carotid arteries displayed a continuous single layer
distribution and smooth muscle cells lined the vascular wall. On
the contrary, endothelial cells in atherosclerotic carotid arteries
did not have a continuous single layer distribution and smooth
muscle cells were disorganized. At the same time macrophages
infiltrated the atherosclerotic plaque.
Immunohistochemical analysis of cross-sections of
the carotid artery tissue specimens was performed to detect RAGE
expression. The expression of RAGE was more prominent in the
atherosclerotic carotid artery plaque specimens compared with that
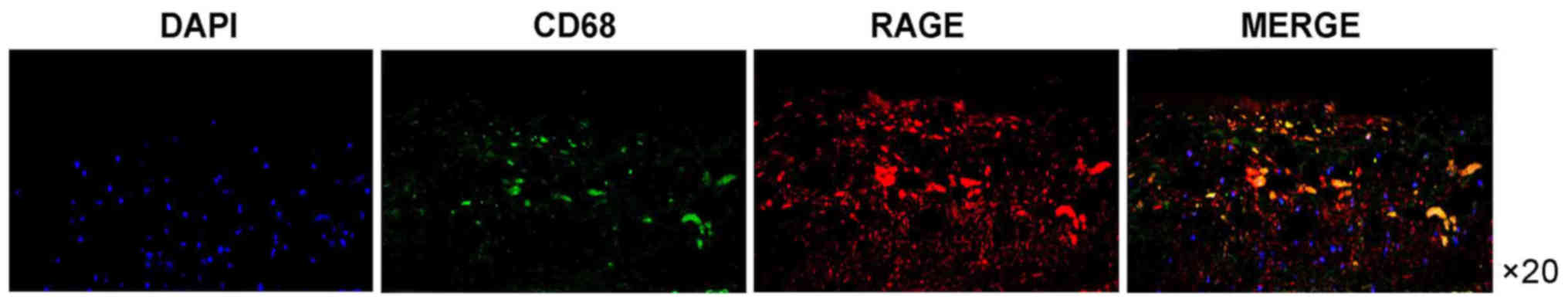
in the control carotid artery specimens (Fig. 1). Furthermore, the RAGE expression
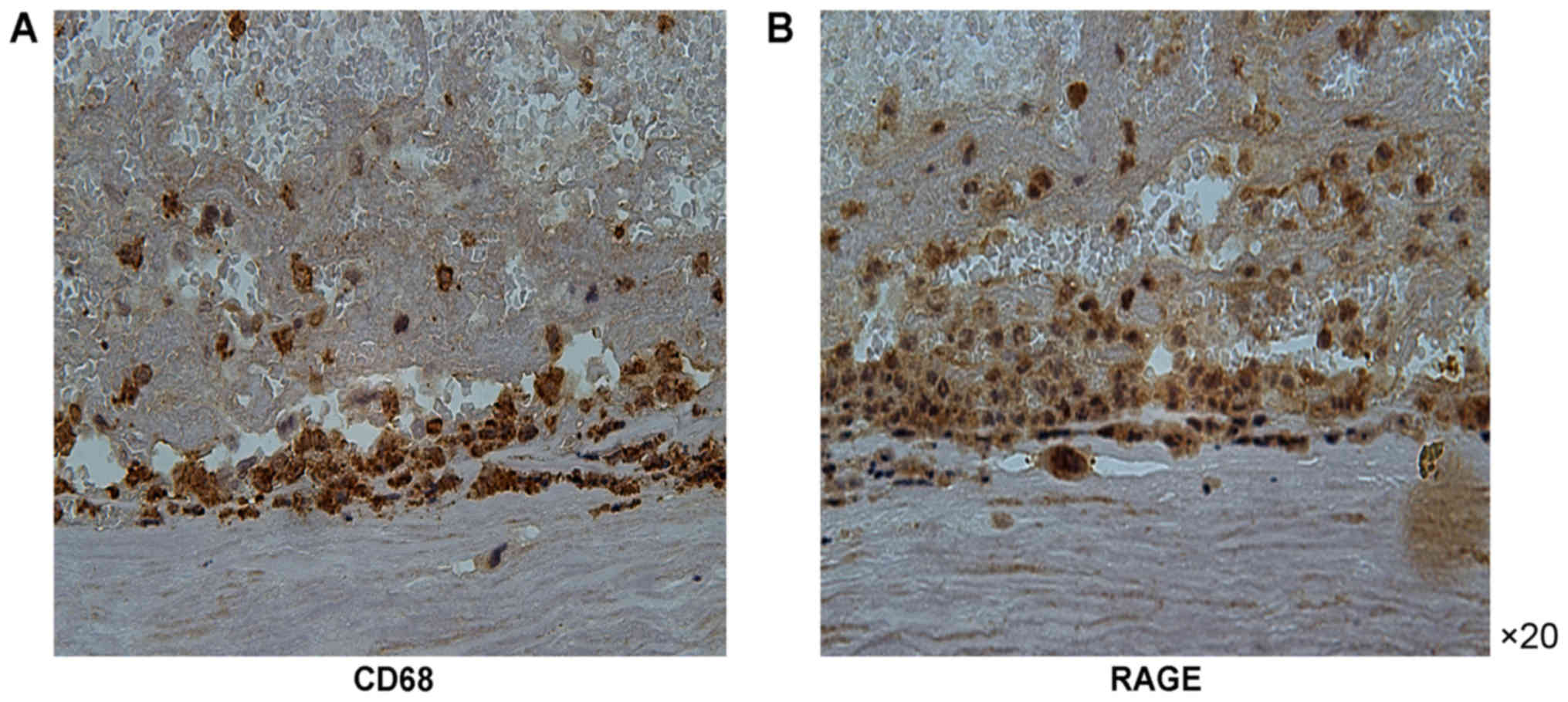
was more prominent in the atherosclerotic carotid tissue specimens
containing CD68-positive macrophages compared with that in the
carotid artery specimens of the control group. RAGE expression was
also increased in areas of macrophage infiltration (Fig. 2). Of note, immunofluorescence
analysis of RAGE indicated that it was closely colocalized with the
CD68-positive regions. RAGE-positive regions were also positive for
CD68 (Figs. 1–3).
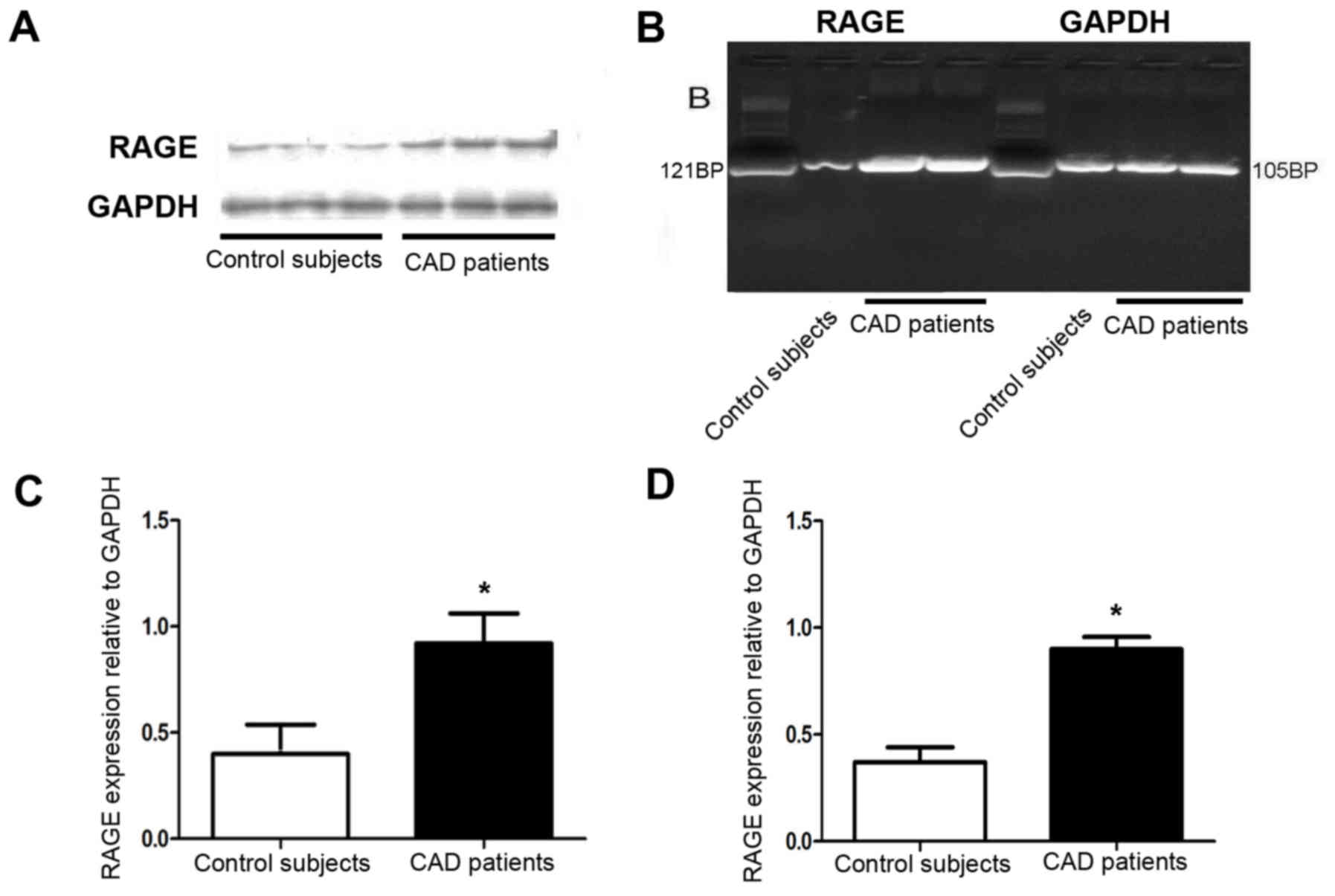
Furthermore, RT-PCR and western blot analysis
respectively indicated that the mRNA and protein expression of RAGE
was increased in the atherosclerotic coronary artery compared with
that in the normal coronary artery tissues (Fig. 4A-D).
Plasma S100B levels in RAGE genotype
subsets
The calcium-binding protein S100B binds to RAGE,
which leads to activation of the RAGE-dependent signal transduction
pathway, resulting in atherogenesis. Therefore, potential
differences in plasma S100B levels between CAD patients and control
subjects were assessed. The plasma S100B levels in the CAD patients
were upregulated in comparison with those in the control group
(243.97±48.29 vs. 102.4 ±25.16 pg/ml; P<0.05). Of note, patients
with the RAGE rs2269422 AA/AT variant had higher plasma levels of
S100B compared with those of the TT genotype in the CAD and control
groups (286.69±63.91 vs. 201.24±32.68 pg/ml and 129.43±21.34 vs.
95.52±28.98 pg/ml, respectively; P<0.05; Table IV). Furthermore, exclusion of
diabetic patients from the analysis did not significantly affect
the results, and it was also indicated that the RAGE s2269422
variant was associated with the occurrence of CAD and that genetic
variations of RAGE have a significant impact on the risk of CAD in
the present Han Chinese population.
| Table IV.Plasma S100B levels according to the
TT and AA/AT variants of the rs2269422 polymorphism of receptor for
advanced glycation end products. |
Table IV.
Plasma S100B levels according to the
TT and AA/AT variants of the rs2269422 polymorphism of receptor for
advanced glycation end products.
|
| S100B (pg/ml) |
|---|
|
|
|
|---|
| Group | TT | AA/AT |
|---|
| Control (n=90) | 95.52±28.98 | 129.43±21.34 |
| CAD (n=70) |
201.24±32.68a |
286.69±63.91a |
Discussion
CAD is an atherosclerotic inflammatory disease, and
oxidative stress and inflammation have an important role in its
pathogenesis (1). RAGE, as a
transmembrane receptor, binds to cellular ligands, which induces
signal transduction and cellular dysfunction, including the
activation of nuclear factor κB and cyclic adenosine monophosphate
pathways, leading to the amplification of atherosclerotic
inflammatory responses (27–29). The present study demonstrated an
increase in RAGE expression in atherosclerotic carotid plaques
compared with that in normal arteries. Furthermore, RT-PCR and
immunohistochemistry revealed increased RAGE mRNA levels and
protein expression in coronary arteries of CAD patients vs. that in
normal controls. It was further indicated that compared with the
corresponding wild-type genotype, the rs2269422 SNP of RAGE was
associated with an increased susceptibility to atherosclerosis.
Furthermore, S100B expression in subjects with the AA/AT genotype
of the RAGE SNP rs2269422 was higher compared with that in the TT
genotype carriers; as this was also identified in control subjects,
it may not be CAD-specific.
Previous studies have established the association
between RAGE, the pathogenesis of atherosclerosis and the
occurrence of CAD, highlighting that RAGE has a central role in the
formation of unstable atherosclerotic plaques and the pathogenesis
and progression of CAD (6,30). However, at present, the pathogenesis
of CAD, including the association between the tagSNPs of RAGE and
the occurrence of CAD, remains to be fully elucidated. In the
present study, the association between 25 tagSNPs of RAGE and CAD
was determined, and it was revealed that the presence of the A
allele of the rs2269422 SNP of RAGE significantly increases the
risk of CAD.
The results of the present study suggested that
macrophages in atherosclerotic plaques of the carotid artery
express more RAGE than the normal carotid tissues. Furthermore, the
RAGE rs2269422 AA allele carriers had a 1.9-fold increased risk of
CAD compared with that of the TT allele carriers. Of note, CAD
patients also had increased plasma S100B levels compared with the
control patients. Furthermore, the plasma S100B levels in RAGE
rs2269422 AA/AT genotype carriers were also significantly increased
in comparison with those in the RAGE rs2269422 TT genotype
carriers, which were demonstrated in the control group as well as
the CAD group.
Several studies have investigated the effect of RAGE
polymorphisms on atherosclerotic disease. However, the effect of
tagSNPs of RAGE on CAD has remained to be fully elucidated. Watson
et al (31) indicated that
RAGE p.Gly82Ser (G82S) mutation have a major effect on the
susceptibility of type 2 diabetes mellitus patients to CAD.
Subjects with the SS genotype of the G82S variant of RAGE had an
increased risk of cardiovascular disease compared with that in the
control group (6,30). To date, only a small number of
candidate genes have been identified to be associated with CAD in
Han Chinese populations. To the best of our knowledge, the
association between the RAGE rs2269422 TT genotype with the
progression and development of CAD has not been previously
reported.
In the present study, an increased expression of
RAGE was identified in macrophages, which may be activated and lead
to the production of proteins, including S100B and pro-inflammatory
cytokines, promoting inflammation and cytotoxicity. Upon S100B
binding to RAGE, a series of cellular signaling pathways is
activated to induce a massive release of inflammatory factors into
the blood. The expression of RAGE by inflammatory cells attracts
macrophages into the lesion region, followed by accumulation of
excess cholesterol in the macrophages, contributing to the
formation of foam cells in atherosclerotic lesions and then
enhancing the severity of the coronary artery lesion (6,32). Thus,
RAGE-positive macrophages and their ligands, including S100B, have
an intermediary part in regulating cell proliferation,
differentiation and apoptosis in the development of atherosclerotic
plaques.
The rs2269422 SNP located in the intron region of
the RAGE gene has not been previously reported to be associated
with the occurrence of CAD in Han Chinese populations. The present
study identified that the intronic rs2269422 SNP either directly or
indirectly impaired the function of RAGE. An increasing number of
studies have indicated that variations in the intron region
influence gene splicing. Furthermore, the rs2269422 RAGE variant
did not obviously affect gene translation, but it may indirectly
influence the RAGE mRNA splicing function. Of note, relative to the
control group, the plasma S100B levels of patients with CAD were
also significantly higher and this requires further
investigation.
The present study has several limitations that
require to be addressed. First, the subjects enrolled in the study
were from a homogenous Northern Han Chinese population. Thus, the
results of the present case-control study should be verified in
other populations. Furthermore, the 1,719 patients included in the
present study may be too few to extend this conclusion to the
entire Han Chinese population. In addition, an investigation of
changes in potential novel signal transduction pathways, such as
the RAGE-MAPK-NF-κB signaling pathway, caused by variations in RAGE
and its ligands is required in future studies.
In conclusion, the present study indicates a
correlation between the RAGE rs2269422 SNP and the occurrence of
CAD in a Han Chinese population, and suggests that RAGE may
contribute to the formation of coronary atherosclerotic plaques in
patients with CAD. The rs2269422 variant of RAGE may thus be a
genetic risk factor for the occurrence of CAD.
Acknowledgements
The authors would like to thank Miss Li Na and Miss
Jun-Yan Du (Department of Cardiology, Shenyang Military General
Hospital, Shenyang, China) for their efforts in collecting the data
for the preliminary study.
Funding
The present study was supported by grants from the
National Key Research and Development Program of China for the 13th
five-year plan (grant no. 2016YFC1301300), the National Science
Foundation of China (grant no. 81670340) and the Liaoning Science
and Technology Project (grant no. 2015010400-301).
Availability of data and materials
Data from the current study are available from the
corresponding author on reasonable request.
Authors' contributions
XZ, MC, FT and XS collected and statistically
processed the clinical data. XY and MC performed the
immunohistochemistry and immunofluorescence assays. XZ and FT
performed the RT-PCR assay. XZ and XS performed the ELISA.
Ethics approval and consent to
participate
The present study complied with the Declaration of
Helsinki and informed consent had been obtained from the subjects
or their relatives. The Shenyang Military General Hospital Ethics
Committee approved the study protocol.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests regarding this study.
References
|
1
|
Kriszbacher I, Koppan M and Bodis J:
Inflammation, atherosclerosis, and coronary artery disease. N Engl
J Med. 353:429–430. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Khera AV, Emdin CA, Drake I, Natarajan P,
Bick AG, Cook NR, Chasman DI, Baber U, Mehran R, Rader DJ, et al:
Genetic risk, adherence to a healthy lifestyle, and coronary
disease. N Engl J Med. 375:2349–2358. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Abaci A: Management of cardiovascular risk
factors for primary prevention: Evaluation of Turkey results of the
EURIKA study. Turk Kardiyol DernArs. 40:135–142. 2012. View Article : Google Scholar
|
|
4
|
Arbab-Zadeh A and Fuster V: The myth of
the ‘vulnerable plaque’: Transitioning from a focus on individual
lesions to atherosclerotic disease burden for coronary artery
disease risk assessment. J Am Coll Cardiol. 65:846–855. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bjorkegren JLM, Kovacic JC, Dudley JT and
Schadt EE: Genome-wide significant loci: How important are they?
Systems genetics to understand heritability of coronary artery
disease and other common complex disorders. J Am Coll Cardiol.
65:830–845. 2015.PubMed/NCBI
|
|
6
|
Gao J, Deng L, Wang Y, Shi Y, Xiao X,
Zheng X, Ren H and Xu D: Relationship between RAGE gene
polymorphisms and cardiovascular disease prognosis in the Chinese
Han population. Mol Genet Genomics. 292:1139–1149. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kajikawa M, Nakashima A, Fujimura N,
Maruhashi T, Iwamoto Y, Iwamoto A, Matsumoto T, Oda N, Hidaka T,
Kihara Y, et al: Ratio of serum levels of AGEs to soluble form of
RAGE is a predictor of endothelial function. Diabetes Care.
38:119–125. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Katakami N: Can soluble receptor for
advanced glycation end-product (sRAGE) levels in blood be used as a
predictor of cardiovascular diseases? Atherosclerosis. 266:223–225.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nam MH, Son WR, Lee YS and Lee KW:
Glycolaldehyde-derived advanced glycation end products
(glycol-AGEs)-induced vascular smooth muscle cell dysfunction is
regulated by the AGES-receptor (RAGE) axis in endothelium. Cell
Commun Adhes. 22:67–78. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Puri R, Nicholls SJ, Shao M, Kataoka Y,
Uno K, Kapadia SR, Tuzcu EM and Nissen SE: Impact of statins on
serial coronary calcification during atheroma progression and
regression. J Am Coll Cardiol. 65:1273–1282. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Reichert S, Triebert U, Santos AN, Hofmann
B, Schaller HG, Schlitt A and Schulz S: Soluble form of receptor
for advanced glycation end products and incidence of new
cardiovascular events among patients with cardiovascular disease.
Atherosclerosis. 266:234–239. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rowisha M, El-Batch M, El Shikh T, El
Melegy S and Aly H: Soluble receptor and gene polymorphism for AGE:
Relationship with obesity and cardiovascular risks. Pediatr Res.
80:67–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ligthart S, Sedaghat S, Ikram MA, Hofman
A, Franco OH and Dehghan A: EN-RAGE: A novel inflammatory marker
for incident coronary heart disease. Arterioscler Thromb Vasc Biol.
34:2695–2699. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
American College of Cardiology
Foundation/American Heart Association Task Force on Practice
Guidelines; American Society of Echocardiography; American Society
of Nuclear Cardiology; Heart Rhythm Society; Society of
Cardiovascular Anesthesiologists; Society for Cardiovascular
Angiography and Interventions; Society for Vascular Medicine;
Society for Vascular Surgery, ; Fleisher LA, Beckman JA, et al:
2009 ACCF/AHA focused update on perioperative beta blockade
incorporated into the ACC/AHA 2007 guidelines on perioperative
cardiovascular evaluation and care for noncardiac surgery. J Am
Coll Cardiol. 54:e13–e118. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Elisaf M, Tzouvelekis E and Nikas N; Greek
EURIKA Investigators, : Primary prevention of cardiovascular
disease in Greece: Greek results of the EURIKA study. Hellenic J
Cardiol. 55:217–226. 2014.PubMed/NCBI
|
|
16
|
Ford TJ, Rocchiccioli P, Good R,
McEntegart M, Eteiba H, Watkins S, Shaukat A, Lindsay M, Robertson
K, Hood S, et al: Systemic microvascular dysfunction in
microvascular and vasospastic angina. Eur Heart J. 1–12.
2018.PubMed/NCBI
|
|
17
|
Ong P, Athanasiadis A and Sechtem U:
Treatment of angina pectoris associated with coronary microvascular
dysfunction. Cardiovasc Drugs Ther. 30:351–356. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Loffler AI and Bourque JM: Coronary
microvascular dysfunction, microvascular angina, and management.
Curr Cardiol Rep. 18:12016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ezhumalai B, Ananthakrishnapillai A,
Selvaraj RJ, Satheesh S and Jayaraman B: Cardiac syndrome X:
Clinical characteristics revisited. Indian Heart J. 67:328–331.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chhabra L and Kowlgi NG: Low incidence of
diabetes mellitus in coronary microvascular dysfunction: An
intriguing association. JACC Cardiovasc Interv. 9:395–396. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Suzuki H: Different definition of
microvascular angina. Eur J Clin Invest. 45:1360–1366. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Morrison AC, Fu YP and O'Donnell CJ;
Cohorts for Heart and Aging Research in Genomic Epidemiology
(CHARGE) Consortium Subclinical Atherosclerosis and CHD Working
Working, : Working Group: Variants in ANGPTL4 and the risk of
coronary artery disease. N Engl J Med. 375:23032016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu Y, Tian X, Li Y, Liu D, Liu M, Zhang
X, Zhang Q, Yan C and Han Y: Up-Regulation of CREG expression by
the transcription factor GATA1 inhibits high glucose- and high
palmitate-induced apoptosis in human umbilical vein endothelial
cells. PLoS One. 11:e01548612016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cai W, Tao J, Zhang X, Tian X, Liu T, Feng
X, Bai J, Yan C and Han Y: Contribution of homeostatic chemokines
CCL19 and CCL21 and their receptor CCR7 to coronary artery disease.
Arterioscler Thromb Vasc Biol. 34:1933–1941. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Okon I, Ding Y and Zou MH: Ablation of
interferon regulatory factor 3 promotes the stability of
atherosclerotic plaques. Hypertension. 69:407–408. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang M, Han Y, Zhang X, Pei F, Deng J,
Kang J and Yan C: An intron polymorphism in the CXCL16 gene is
associated with increased risk of coronary artery disease in
Chinese Han population: A large angiography-based study.
Atherosclerosis. 210:160–165. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao D, Tong L, Zhang L, Li H, Wan Y and
Zhang T: Tanshinone II A stabilizes vulnerable plaques by
suppressing RAGE signaling and NF-κB activation in
apolipoprotein-E-deficient mice. Mol Med Rep. 14:4983–4990. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu Y, Wang L, Delguste F, Durand A,
Guilbaud A, Rousselin C, Schmidt AM, Tessier F, Boulanger E and
Neviere R: Advanced glycation end products receptor RAGE controls
myocardial dysfunction and oxidative stress in high-fat fed mice by
sustaining mitochondrial dynamics and autophagy-lysosome pathway.
Free Radic Biol Med. 112:397–410. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Muniyappa R and Srinivas PR: Dicarbonyl
stress and atherosclerosis: Is it all RAGE? Diabetes. 63:3587–3589.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ma WQ, Qu QR, Zhao Y and Liu NF:
Association of RAGE gene Gly82Ser polymorphism with coronary artery
disease and ischemic stroke: A systematic review and meta-analysis.
Medicine (Baltimore). 95:e55932016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Watson AM, Li J, Samijono D, Bierhaus A,
Thomas MC, Jandeleit-Dahm KA and Cooper ME: Quinapril treatment
abolishes diabetes-associated atherosclerosis in
RAGE/apolipoprotein E double knockout mice. Atherosclerosis.
235:444–448. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li Y, Zhu J, Chen L, Hu W, Wang M, Li S,
Gu X, Tao H, Zhao B, Ma G and Li K: Genetic predisposition to
ischaemic stroke by RAGE and HMGB1 gene variants in Chinese Han
population. Oncotarget. 8:100150–100164. 2017.PubMed/NCBI
|