Introduction
Colorectal cancer (CRC) is the third most common
type of cancer and the fourth-leading cause of cancer-associated
mortality worldwide (1). Genetic and
epigenetic genomic alterations, leading to gene amplification, the
activation of oncogenes and the loss of tumor suppressor genes, are
involved in the tumorigenesis of CRC (2). Risk factors associated with CRC include
environmental and dietary mutagens, intestinal microorganisms and
pathogens, and chronic intestinal inflammation (3). The connection between inflammation and
tumorigenesis is well established. The processes of
colitis-associated cancer (CAC) and non-inflammatory CRC feature
common events, including the development of localized hyperplasia,
polyps, adenoma and ultimately, carcinoma (4). CAC is the CRC subtype preceded by
inflammatory bowel disease (IBD), Crohn's disease or ulcerative
colitis (UC) (5). UC is associated
with a cumulative risk of CAC of ≤20% within 30 years of disease
onset (5,6). Azoxymethane (AOM) is a mutagenic agent
that induces multiple colonic tumors following a single injection
in chronic colitis mouse models within short periods (7). This mouse model of CAC is valuable for
the understanding of the mechanisms of inflammation in
tumorigenesis (8).
Endoplasmic reticulum (ER) stress is associated with
both the pathogenesis of intestinal inflammation and tumorigenesis
(9–11). The ER is crucial for the folding of
secretory and membrane proteins, lipid biosynthesis, the regulation
of intracellular Ca2+ and redox signaling. During ER
stress, the disturbance of ER homeostasis due to oxidative stress,
energy deprivation, altered metabolic status or inflammatory
stimuli causes ER calcium depletion and the accumulation of
unfolded and misfolded proteins in the ER lumen (12). In mammalian cells, there are three
types of ER stress sensors: Protein-kinase-RNA-like-ER kinase,
activating transcription factor 6 and inositol-requiring enzyme
(IRE) 1. These transmembrane ER proteins regulate the unfolded
protein response to address ER stress (13,14).
IRE1 is the most evolutionally conserved among the three ER stress
response sensors (15). There are
two IRE1 paralogs, α and β, present in mammals (16). IRE1α is a ubiquitously expressed
kinase and site-specific RNA endonuclease (17,18),
that processes the mRNA encoding X-box binding protein (XBP) 1 to
maintain ER homeostasis (19). IRE1α
excises a 26-nucleotide-intron sequence from the unspliced XBP1u
mRNA to produce spliced XBP-1s mRNA, resulting in the activation of
XBP1 protein expression. The XBP1 protein is a transcription factor
involved in tumor growth and survival (10,20).
Although IRE1β is a paralog of IRE1α (21), its expression is restricted to the
epithelium of the gastrointestinal and respiratory tracts (16,22),
while IRE1α is expressed in a variety of tissues (23). IRE1α and IRE1β protect mice from
dextran sulfate sodium (DSS)-induced colitis (16,24);
however, these two IRE1 proteins have different functions in ER
stress and varying substrate specificities (25). For example, IRE1β has a decreased
XBP1u mRNA RNase activity compared with IRE1α (22) and IRE1β degrades 28S rRNA more
efficiently than IRE1α (26). IRE1β
overexpression in HeLa cells results in the cleavage of 28S rRNA,
leading to cell apoptosis (27).
IRE1β, but not IRE1α, regulates the secretory protein mucin (MUC) 2
in intestinal goblet cells (26,28).
MUC2 is a putatively vital molecule in the innate immune defense of
the colon; a lack of the MUC2 gene results in spontaneous colitis
and colon cancer in mice (29–32).
The current study induced colonic tumor formation in
mice using AOM and dextran sulfate sodium (DSS). It was observed
that IRE1β and MUC2 expression was downregulated in the tumors.
This indicated that the inhibition of the IRE1β-MUC2 signaling
pathway may promote the occurrence and development of colonic
tumors.
Materials and methods
Mice
Female C57BL/6 (B6) mice (Vital River Laboratory
Animal Technology Co., Ltd., Beijing, China) were maintained in the
animal experimental center of The First Affiliated Hospital of
Henan University of Science and Technology. Mice were housed with a
controlled temperature of 20–22°C, 50% humidity and a 12-h
light/dark cycle; they were fed with rodent chow from Beijing HFK
Bioscience Co., Ltd. (Beijing, China). The Animal Care and Use
Committee of The First Affiliated Hospital of Henan University of
Science and Technology (Luoyang, China) approved all the animal
procedures in this study.
Colitis-mediated colon tumors were induced using a
modified protocol previously described by Neufert et al
(7). A total of 20 mice (age, 7–8
weeks; weight, 19–23 g) were randomly allocated into the control
and tumor groups (n=10/group). Mice in the tumor group received an
intraperitoneal injection of 1 mg/ml AOM (12 mg/kg; molecular
weight (MW), 74.08 Da; cat. no. MFCD00126912; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) and control mice received saline (12
ml/kg saline). In primary experiments, the high mortality of mice
was associated with direct administration following intraperitoneal
injection, so on the 7th day after intraperitoneal injection of
AOM, mice in the tumor group received 1.0% DSS (MW 36–50 kDa; cat.
no. 160110; MP Biomedicals, LLC, Santa Ana, CA, USA) for 7 days to
induce colitis. The 1% DSS solution was prepared by dissolving
fine-grain DSS powder (1 g) in 100 ml drinking water. DSS solution
was freshly prepared prior to administration. Water bottles
containing DSS were replaced at 5 days with fresh DSS solution for
the remaining 3 days. On day 8, the DSS solution was replaced with
normal drinking water for 14 days; mice in the control group did
not receive DSS. The cycle of 7 days DSS/14 days normal drinking
water was repeated three times. Control mice were provided with
normal drinking water throughout the experiment. Mice in both
groups had free access to food and water. The mice were monitored
once every two days for total of 69 days to record body weight,
stool consistency, rectal bleeding and ulceration.
Mice were euthanized at the end of the third cycle.
The entire colon was excised and measured. Colons were cut open and
laid flat, lumen-side up. The number and size of colonic tumors was
assessed. Colon tissues and tumor tissues were frozen at −80°C for
subsequent reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) and western blot analyses. RNAlater
Stabilization reagent (Qiagen GmbH, Hilden, Germany) was used to
prevent mRNA degradation according to the manufacturers'
instructions. Further samples from the colon and tumor tissue were
fixed with 10% neutral-formalin for 24 h at room temperature and
embedded in paraffin 62°C for 3 h. Sections were stained by
hematoxylin and eosin (H&E) and immunohistochemistry (IHC) as
described below. The severity of colitis was evaluated based on the
disease activity index (DAI) using a previously described method
(Table I) (33).
 | Table I.Disease activity index score. |
Table I.
Disease activity index score.
|
| Assigned score |
|---|
|
|
|
|---|
| Characteristic | 0 | 1 | 2 | 3 | 4 |
|---|
| Body mass decrease
(%) | 0 | 1–5 | 5–10 | 10–15 | >15 |
| Stool
appearance | Normal | – | Loose | – | Diarrhea |
| Rectal
bleeding | Normal | – | Hemoccult
positive | – | Gross bleeding |
Histopathology examination
Tissue slices (4-µm) were prepared from distal colon
tissue samples and stained with H&E for histological
examination. Staining was performed at room temperature and 1%
Hematoxylin (stained for 1–10 min) and 0.5% Eosin (stained for 2–5
min) were used. Pathology and level of inflammation were scored in
a blinded fashion using a previously described scoring system
(34) to quantify: The extent of
neutrophil and lymphocyte infiltration (0–3 points); Paneth and
goblet cell degranulation (0–2 points); epithelium reactivity,
including crypt distortion (0–3 points); and inflammatory foci (0–3
points).
IHC was conducted with sections mounted on
poly-L-lysine-coated slides using a modified biotin-peroxidase
complex method as previously described (35). Briefly, tissue sections were
incubated overnight at 4°C with rabbit polyclonal antibodies
against IRE1α (dilution, 1:150; cat. no. sc-20790; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), IRE1β (dilution, 1:100; cat.
no. GTX87426; GeneTex, Inc., Irvine, CA, USA) and MUC2 (dilution,
1:100; cat. no. ab76774; Abcam, Cambridge, UK). Antigen-antibody
complexes were detected with a biotinylated goat anti-rabbit
antibody (dilution, 1:300; cat. no. SA1020; Boster Biological
Technology Co., Ltd., Wuhan, China) conjugated with
streptavidin-horseradish peroxidase (HRP) for 30 min at 37°C. At
room temperature and visualized by reacting with the
3,3-diaminobenzidine reagent kit (Solarbio Science & Technology
Co., Ltd., Beijing, China; cat. no. DA1010) at room temperature for
5 min. Sections were counterstained with 1% hematoxylin for 1–10
min at room temperature. Negative control sections were obtained by
omitting the primary antibody or by using an unspecific antibody.
Images at a magnification of ×400 were captured using a Nikon
Ds-Fi2 500w light microscope, (Nikon Corporation, Tokyo, Japan).
IHC staining was used to semi-quantitatively determine protein
levels as previously described (35): A total of 10 fields were randomly
selected on each slide and 100 cells per field were counted and
scored for IRE1α, IRE1β and MUC2 staining. The semi-quantitative
scores were obtained from the staining intensity (none, 0; weak, 1;
moderate, 2; strong, 3) multiplied by the percentage of positively
stained cells (≤5%, 0; 6–25%, 1; 26–50%, 2; 51–75%, 3; and >75%,
4).
RT-qPCR analysis
Total RNA from the colon tissue samples was
extracted using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
instructions. cDNA was synthesized using PrimeScript-RT Master mix
(Takara Bio, Inc., Otsu, Japan) with the following temperature
protocol: 37°C for 15 min, 85°C for 5 sec and 4°C for 10 min. qPCR
was performed as described previously, using undiluted cDNA
templates (35). Primer sequences
(Table II) were designed with
Primer 3.0 (36) and synthesized by
Sangon Biotech Co., Ltd. (Zhengzhou, China). qPCR was performed
using a CFX96 Real-Time PCR system (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) and SYBR green (Takara Bio, Inc.) was used to
detect PCR products. The reaction followed an initial step at 95°C
for 30 sec, followed by 40 cycles of 95°C for 5 sec, 58°C for 30
sec and 72°C for 30 sec. For annealing temperatures >60°C, a
two-step method was used (37). Each
sample was assayed in triplicate. Amplification efficiency was
>97%. A melting curve analysis was performed for the PCR
products of each target gene and β-actin to evaluate primer
specificity. The relative abundance of target gene mRNA level was
evaluated using 2−ΔΔCq method, normalized to β-actin
(38). mRNA expression levels were
presented as relative fold by comparing the quantity of mRNA
between different groups. When Cq >35, the gene was considered
to not be expressed.
| Table II.Reverse transcription-quantitative
polymerase chain reaction primers. |
Table II.
Reverse transcription-quantitative
polymerase chain reaction primers.
|
| Primer (5′-3′) |
|---|
|
|
|
|---|
| Gene | Forward | Reverse |
|---|
| IL-6 |
AACGATGATGCACTTGCAGA |
TGGTACTCCAGAAGACCAGAGG |
| TNF-α |
CCACCACGCTCTTCTGTCTACT |
TGCTACGACGTGGGCTACA |
| IL-8 |
CTAGGCATCTTCGTCCGTCC |
TTGGGCCAACAGTAGCCTTC |
| XBP1u |
GGTCTGCTGAGTCCGCAGCACTC |
AGGCTTGGTGTATACATGG |
| XBP1s |
GGTCTGCTGAGTCCGCAGCAGG |
AGGCTTGGTGTATACATGG |
| IRE1α |
GCATCACCAAGTGGAAGTATC |
ACCATTGAGGGAGAGGCATAG |
| IRE1β |
CACAACCTATCGCCGCTACT |
CATCCTGGTGCCATGTGTAA |
| MUC2 |
CTGACCAAGAGCGAACACAA |
CATGACTGGAAGCAACTGGA |
| β-actin |
GGCTGTATTCCCCTCCATCG |
CCAGTTGGTAACAATGCCATGT |
Western blotting
Protein lysates were prepared from the extracted
tissues in radioimmunoprecipitation assay lysis buffer (cat. no.
R0020; Solarbio Science & Technology Co., Ltd.) on ice by
homogenization with a grinder. The supernatant was obtained by
centrifugation (15 min; 10,800 × g; 4°C. The bicinchoninic acid
(cat. no. PC0020; Solarbio Science & Technology Co., Ltd.)
method was used to determine protein concentration. Protein (30 µg)
from each sample was denatured, resolved on 10% SDS-PAGE gels and
transferred onto polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA). Rabbit polyclonal antibodies
against IRE1α (dilution, 1:1,000; cat. no. 14C10; Cell Signaling
Technology, Inc., Danvers, MA, USA) and IRE1β (dilution, 1:300;
cat. no. ab135795; Abcam) and mouse monoclonal antibodies against
GAPDH (dilution, 1:1,000; cat. no. CW0100A; CWBIO Technology Co.
Ltd, Beijing, China) were incubated with the membranes at 4°C
overnight, followed by HRP-conjugated anti-rabbit (dilution,
1:1,000; cat. no. BA1054) or anti-mouse IgG (dilution, 1:3,000;
cat. no. BA1050; both Boster Biological Technology Co. Ltd.) for 1
h at room temperature, and enhanced chemiluminescence reagent
(Pierce; Thermo Fisher Scientific, Inc.) according to the
manufacturers' protocols. GAPDH was used to normalize protein
expression. A ChemiDoc XRS (Bio-Rad Laboratories, Inc.) was used to
capture images, which were quantified using ImageJ software v1.48
(National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
All data are presented as the mean ± standard
deviation. Experiments were repeated at least three times. All
statistical analyses were performed using SPSS 20.0 (IBM Corp.,
Armonk, NY, USA). Student's t-test or Mann-Whitney U test were used
to determined significant differences between groups. Wilcoxon
signed-rank test was used for non-parametric data. P<0.05 was
considered to indicate a statistically significant difference.
Results
Treatment with AOM/DSS induces tumors
in mouse colons
According to Thaker et al (39) and preliminary experiments, the most
consistent results were observed using female mice; hence, only
female mice were utilized in the current study. No signs of
inflammation or colonic tumors were observed in the control group.
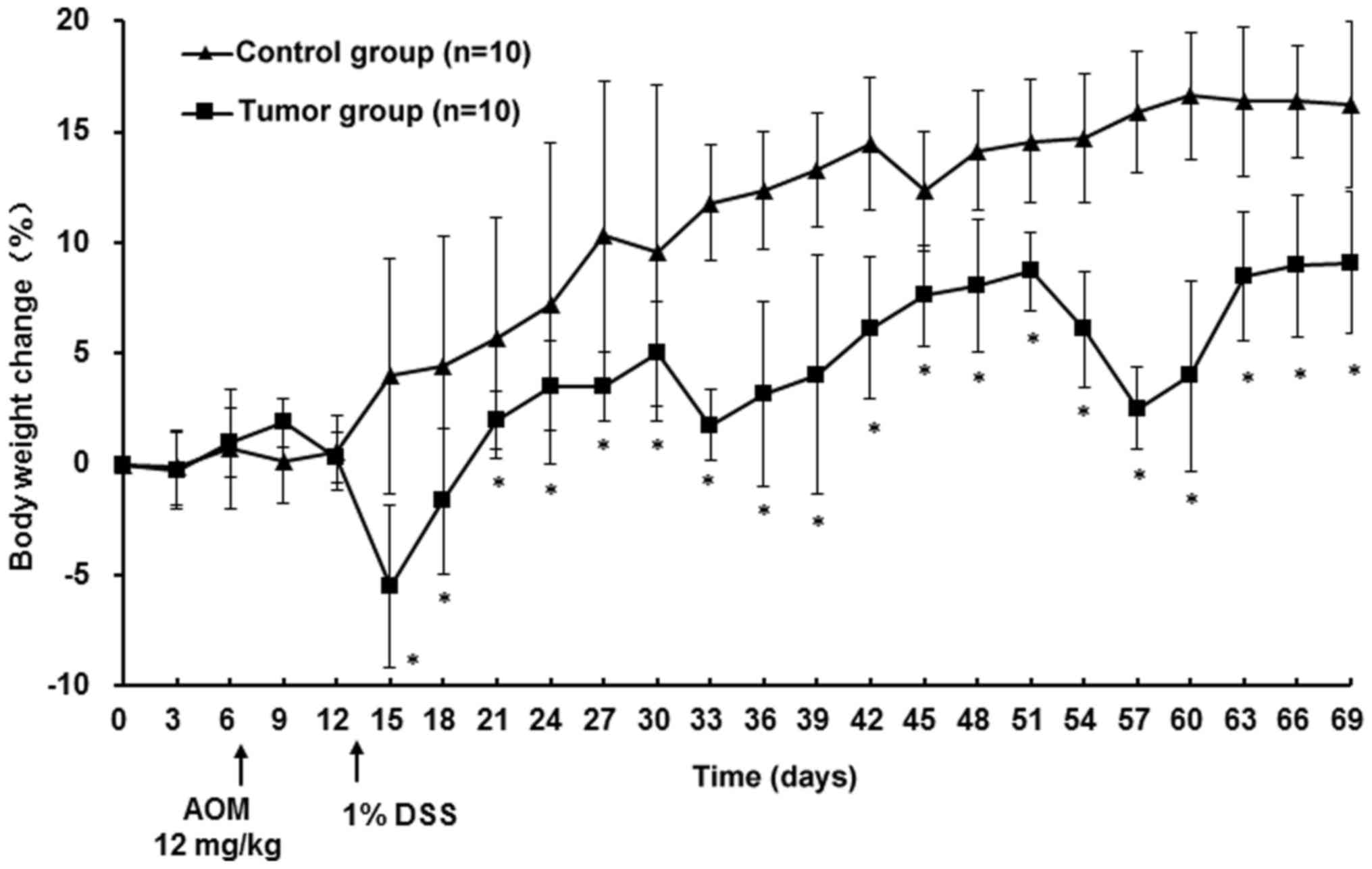
Mice in the tumor group exhibited no sign of illness following AOM
treatment and reduced appetite and drank less starting on day 6 of
the 1% DSS treatment. By day 7, all mice of the tumor group
presented with bloody stools and declining body weight; symptoms
lasted until day 4 after 1% DSS treatment. The weight loss of the
mice in the tumor group was the lowest with −5.5% during the first
cycle. In the following two cycles, mice exhibited similar
symptoms, but less severe. Body weights of mice in the tumor group
increased, although slower compared with the control mice;
particularly during DSS treatment. At the endpoint, there was a
significant difference in body weight between the tumor and the
control group (P<0.05; Fig. 1)
and DAI scores in the tumor group were significantly higher
compared with the control group (P<0.05; Table III). No mortality was observed in
either group. The colon length in the tumor group was significantly
decreased compared with the control group (P<0.05; Table III). A total of 36 (average number
of tumors per mouse, 3.6; range, 2–5) colonic tumors were observed
in mice of the tumor group. The mean tumor diameter was 3.1 mm
(range, 1–4 mm). Histological examination revealed that the mucosa
of the colons from the tumor group were disordered and the crypt
structure was destroyed compared with the control group: Colonic
epithelial cells were atypical, with decreased differentiation,
increased cell size and large nuclei; the nucleus-to-cytoplasm
ratio of the cells was increased, the number of goblet cells was
decreased and the number of inflammatory cells was increased
(Fig. 2A and B).
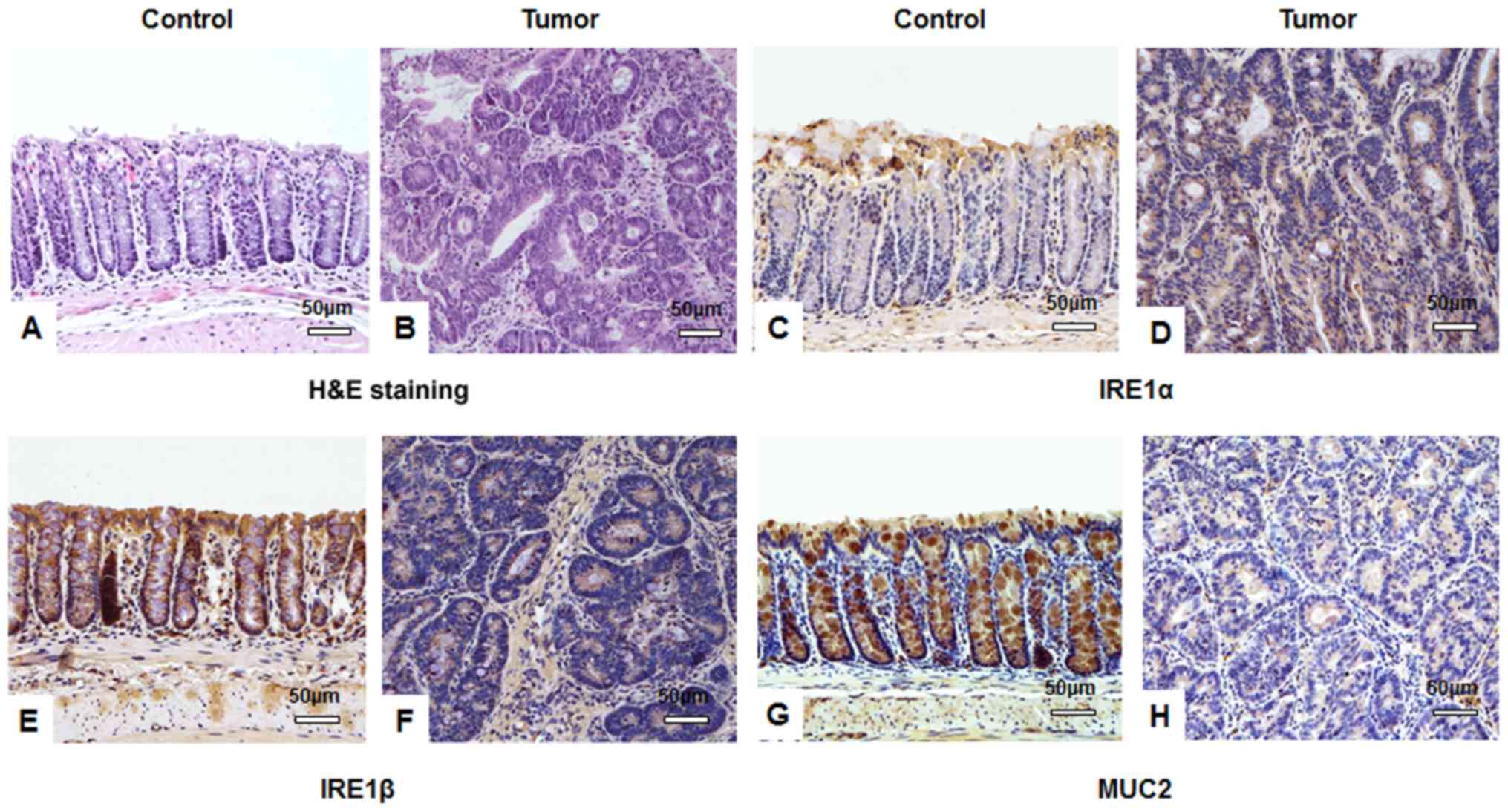 | Figure 2.IHC staining suggests that AOM/DSS
treatment affects IRE1α, IRE1β and MUC2 expression in tissues.
Following a one-off treatment with AOM and three cycles of DSS,
tumor tissues from mice in the tumor group (n=10) and colon tissues
from mice in the untreated control group (n=10) assessed by IHC.
Hematoxylin and eosin staining of (A) control and (B) tumor tissues
identifying typical tumor characteristics, including disordered
mucosa, destroyed crypt structure, less differentiated cells, a
larger cell and nuclei size, and an increased nucleus-to-cytoplasm
ratio. IRE1α protein was stained using IHC in (C) control and (D)
tumor tissues and expression in the cytoplasm of colon submucosa
cells was observed. IHC visualized IRE1β protein in (E) control and
(F) tumor tissues and identified cytoplasmic expression in colonic
mucosa epithelial cells. MUC2 protein expression was visualized
using IHC in goblet cells of (G) control and (H) tumor tissues. All
images were obtained using a Nikon Ds-Fi2 500w [Ds-Fi2; light
microscope (magnification, ×200)]; scale bar, 50 µm. IHC,
immunohistochemistry; AOM, azoxymethane; DSS, dextran sulfate
sodium; IRE, inositol-requiring enzyme; MUC, mucin. |
| Table III.Disease activity index scores and
colon length. |
Table III.
Disease activity index scores and
colon length.
| Group | N | DAI | Colon length
(cm) |
|---|
| Control | 10 | 0 | 8.7±0.6 |
| Tumor | 10 | 3.3a |
6.4±0.6a |
mRNA expression differs between the
control and tumor groups
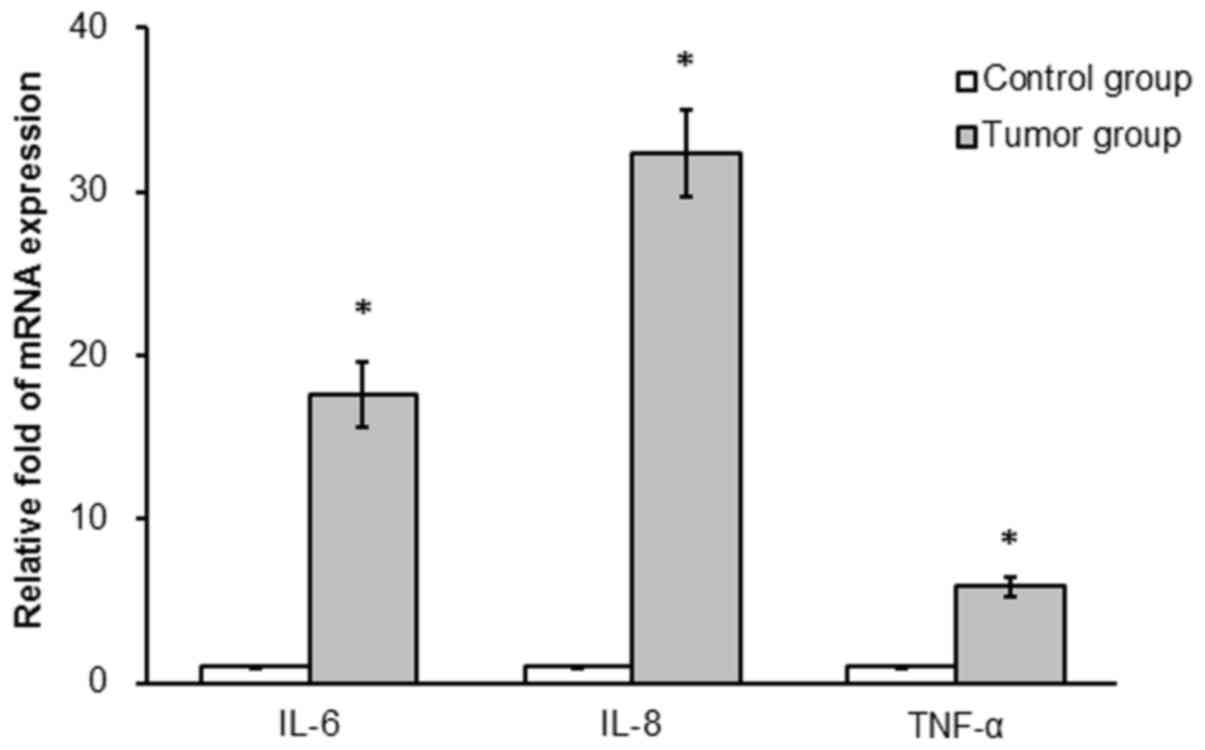
mRNA expression of IL-6, IL-8 and TNF-α was
determined in colonic tissues from the two groups. IL-6, IL-8 and
TNF-α mRNA expression was significantly increased in the tumor
compared with the control group (17.6-, 32.3- and 5.9-fold,
respectively; P<0.05; Fig.
3).
IRE1α and XBP1u mRNA expression levels were not
significantly different between the tumor and the control group
(P>0.05; Fig. 4). XBP1s mRNA
levels were significantly increased in the tumor compared with the
control group (~2-fold; P<0.05; Fig.
4) and IRE1β and MUC2 mRNA levels in the tumor group were
significantly decreased compared with the control group (42 and
30%, respectively; P<0.05; Fig.
4).
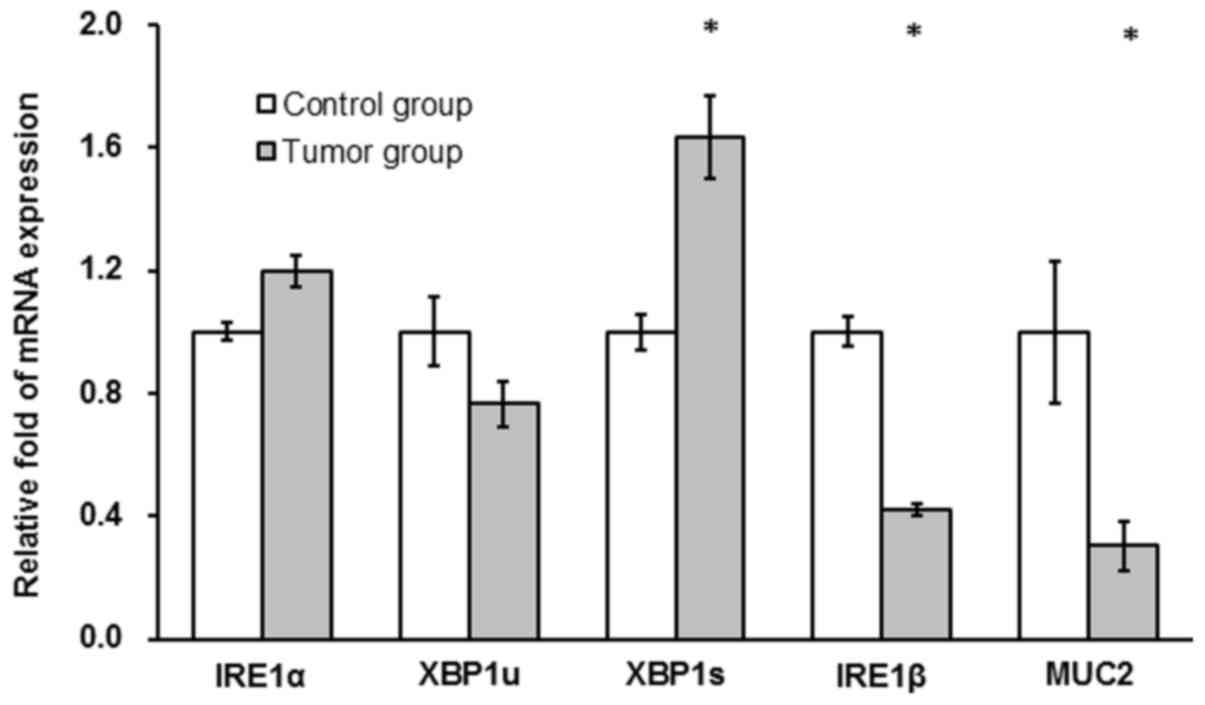 | Figure 4.AOM/DSS treatment affects mRNA levels
of XBP1s, IRE1β and MUC2 in colon tissues of mice. Following a
one-off treatment with AOM and three cycles of DSS, mRNA expression
of IRE1α, XBP1u, XBP1s, IRE1β and MUC2 were determined in tissues
from mice in the untreated control and tumor groups (n=10/group) at
the end point of the experiment using reverse
transcription-quantitative polymerase chain reaction assays.
*P<0.05 vs. control group. AOM, azoxymethane; DSS, dextran
sulfate sodium; IRE, inositol-requiring enzyme; XBP, X-box binding
protein; MUC, mucin. |
IRE1β and MUC2 protein expression is
downregulated in tumor tissues
As determined by IHC staining, there was no
significant difference in IRE1α protein expression between the
tumor and control group (P>0.05; Table IV). IRE1α was primarily expressed in
the cytoplasm of colonic submucosa cells. A low level of IRE1α
expression was observed in the cytoplasm of colonic epithelial
cells, particularly in brush border cells (Fig. 2C and D). IRE1β protein expression in
the tumor group was significantly decreased compared with the
control group (P<0.05; Table
IV). IRE1β protein was predominately expressed in the cytoplasm
of colonic mucosa epithelial cells (Fig.
2E and F). Similar to IRE1β, the expression of MUC2 protein in
the tumor group was significantly lower than in the control group
(P<0.05; Table IV). MUC2 protein
was primarily expressed in the cytoplasm of colonic mucosa
epithelial cells, particularly in goblet cells in the normal mucosa
(Fig. 2G and H).
| Table IV.IRE1α, IRE1β and MUC2 protein
expression in colonic tissues determined by immunohistochemistry
analysis. |
Table IV.
IRE1α, IRE1β and MUC2 protein
expression in colonic tissues determined by immunohistochemistry
analysis.
| Group | N | IRE1α | IRE1β | MUC2 |
|---|
| Control | 10 | 1.50±0.23 | 5.26±0.38 | 4.77±0.47 |
| Tumor | 10 | 1.63±0.18 |
2.53±0.23a |
1.83±0.41a |
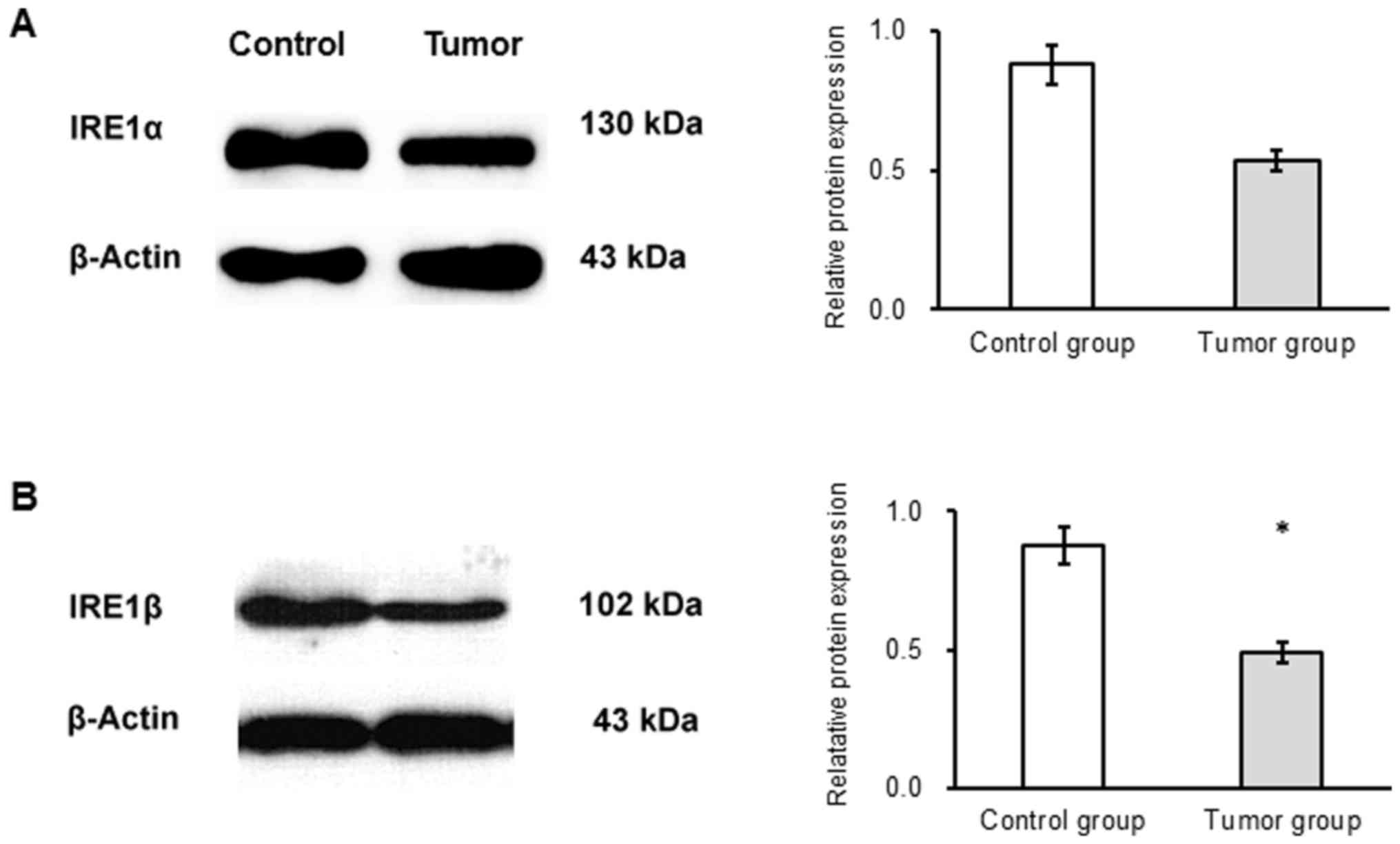
The results of western blot analysis revealed no
significant difference in the expression of IRE1α protein between
the groups (Fig. 5A). IRE1β protein
expression was significantly decreased in the tumor compared with
the control group (P<0.05; Fig.
5B).
Discussion
IRE1α serves a protective role against ER stress and
colitis (24,40). The IRE1-XBP1 signaling pathway serves
a pivotal role in cell survival under conditions of ER stress and
is associated with tumorigenesis of various cancer types (41–44). In
the current study, mRNA expression of IRE1α, XBP1u and XBP1s and
IRE1α protein expression were compared in tumor and normal tissues.
XBP1 mRNA, a signaling molecule in the IRE1α-XBP1 signaling
pathway, is processed by IRE1α (19). XBP1s mRNA expression was
significantly increased in tumor compared with control tissues.
This suggested that the activity of the IRE1α-XBP1 pathway was
increased, indicating that the ER stress response may be associated
with tumorigenesis. The present study further observed that IRE1α
mRNA levels were not increased in tumor tissues compared with the
control, while XBP1s mRNA levels were. Under ER stress, IRE1α is
activated through trans-autophosphorylation and dimerization, or
oligomerization (45). It was
speculated that activated IRE1α levels may be increased, while gene
expression levels remained unchanged. Thus, the determination of
phosphorylated-IRE1α levels requires future assessment.
IRE1α is a transmembrane RNAse that initiates the
splicing of XBP1u to XBP1s. The latter encodes a transcription
activator; its activity induces the expression of ER chaperones
(46). IRE1α promotes cell
survival-associated signal transduction and the transmission of
apoptotic signals (47). It has been
demonstrated that the IRE1-XBP1 signaling pathway is associated to
the pathogenesis of IBD (48,49) and
human cancer (50), leading to the
consideration of the IRE1-XBP1 signaling pathway as a potential
target for cancer therapy (51). A
recent study suggested that IRE1α promotes cell survival by
splicing XBP1 mRNA and promoting regulated IRE1-dependent decay
(RIDD). During the RIDD process, IRE1 promotes the degradation of
mRNAs that predominately encode ER-targeting proteins (52), which may be the mechanism for the
involvement of the IRE1α-XBP1 signaling pathway in the pathogenesis
of inflammation-mediated CRC. It was previously reported that IRE1α
and XBP1 protein expression levels were unchanged in human CRC
tissue compared with adjacent normal tissue (44). Therefore, the role of the IRE1-XBP1
signaling pathway in the pathogenesis of colonic tumors may be
discrepant between humans and mice.
IRE1β, a homolog of IRE1α, protects from colitis and
IRE1β−/− mice are more sensitive to experimentally
induced colitis (16). In contrast
to IRE1α, IRE1β affects different substrates, regulating the mRNA
that encodes ER proteins to maintain ER homeostasis in highly
differentiated secretory cells (28). Divergent effects of IRE1α and IRE1β
on the cell fate may be implicated by their varying roles in
tumorigenesis (25,46). Therefore, the current study analyzed
IRE1β expression in tumor tissues. The results suggested that IRE1β
mRNA and protein levels were decreased in the tumor compared with
the control tissues. Although Tsuru et al (28) reported that IRE1β is specifically
expressed in the ER of goblet cells, present IHC results revealed
that IRE1β positive staining was observed in goblet and absorptive
cells, indicating that IRE1β may affect multiple cells types. IRE1β
acts to protect the intestine from inflammation (52). IRE1β expression in the epithelium of
the gastrointestinal tract is consistent with promoting resistance
to DSS-induced colitis at the epithelial cell level (16). When DSS is ingested, the
gastrointestinal tract epithelial cells are the first to be exposed
to the treatment (16). Toxicity
likely has a primary role in the development of inflammation among
colonic epithelia. DSS acts on the epithelial cell barrier, leading
to reduced IRE1β expression in epithelial cells and weakening the
protection against colonic inflammation (16). As IRE1β is a protective factor in
colitis that can induce apoptosis (23) it is expected that a decrease in IRE1β
expression is associated with tumorigenesis, as apoptosis is
putatively associated with tumor occurrence (53). IRE1β has been reported to regulate
the expression of microsomal triacylglycerol transfer protein, a
regulator of lipid absorption in the colonic epithelium (54); however, it is unclear whether this
function of IRE1β affects tumorigenesis. In an in vitro
study, Dai et al (55)
reported that IRE1β mRNA expression increases in undifferentiated
and decreases in differentiated Caco-2 cells. IRE1β has been
revealed to be involved in cell apoptosis (27). In a previous study, it was reported
that IRE1β and MUC2 expression are downregulated in human colonic
tissues (44). These results
indicated that IRE1β serves an important role in the pathogenesis
of colitis and tumors.
MUC2 is the major component of the intestinal mucus
gel barrier, which serves an important role in the protection of
intestinal function (31).
Furthermore, MUC2 is putative to protecting the intestine against
colitis and colorectal cancer (30).
It has been demonstrated that MUC2-deficient mice spontaneously
develop colon cancer (32).
Consistent with this, the results of the current study revealed
that MUC2 mRNA and protein levels were significantly decreased in
tumor compared with normal tissue. It has been reported that IRE1β
is involved in the ER homeostasis of colon goblet cells and the
synthesis and secretion of MUC2 (28). IRE1β is believed to control the
levels of translatable cytosolic MUC2 mRNA to maintain MUC
production (28). An in vitro
study has revealed that IRE1β regulates secretory proteins in cells
via degrading mRNA in the ER (26).
In addition, IRE1β is essential for airway epithelial MUC
production (22). These data suggest
that IRE1β may regulate MUC2 expression and the downregulation of
IRE1β and MUC2 may reduce the protection of the colon to promote
occurrence and development of tumors.
The current findings suggested that the ER
stress-associated IRE1α-XBP1 signaling pathway was activated during
the occurrence and development of CRC, and potentially contributed
to these processes. The decreased expression of two protective
molecules, IRE1β and MUC2, promoted the occurrence and development
of inflammation and ultimately tumors in the colonic epithelium.
The mechanism of these processes requires further analysis to
identify potential therapeutic targets for the treatment or
prevention of colitis-associated CRC.
Acknowledgements
The authors would like to thank Dr Lei Gao, Dr
Tengfei Guo and Dr Dandan Feng from the Department of
Gastroenterology and Hepatology, The First Affiliated Hospital,
Henan University of Science and Technology (Luoyang, China) for
providing reagents and materials, and for technical support.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81370487) and The
Scientific Research Fund of Beijing Rehabilitation Hospital,
Capital Medical University (grant no. 2017-004).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FD performed the majority of experiments, analyzed
the data and drafted the manuscript. SD, QX, PC and MC provided
technical support, purchased reagents and analyzed the data. ZR and
YF analyzed the data and provided their guidance for the study and
manuscript writing. QG provided funding, designed the current
study, analyzed the data and wrote the manuscript.
Ethics approval and consent to
participate
The Animal Care and Use Committee of The First
Affiliated Hospital of Henan University of Science and Technology
approved all the animal procedures in this study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rabeneck L, Horton S, Zauber AG and Earle
C: Colorectal cancer. JAMA J Am Med Assoc. 270:23022015.
|
|
2
|
Arvelo F, Sojo F and Cotte C: Biology of
colorectal cancer. Ecancermedicalscience. 9:5202015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Raskov H, Pommergaard HC, Burcharth J and
Rosenberg J: Colorectal carcinogenesis-update and perspectives.
World J Gastroenterol. 20:18151–18164. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Grivennikov SI: Inflammation and
colorectal cancer: Colitis-associated neoplasia. Semin
Immunopathol. 35:229–244. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Terzic J, Grivennikov S, Karin E and Karin
M: Inflammation and colon cancer. Gastroenterology.
138:2101–2114.e5. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bye WA, Nguyen TM, Parker CE, Jairath V
and East JE: Strategies for detecting colon cancer in patients with
inflammatory bowel disease. Cochrane Database Syst Rev.
9:CD0002792017.PubMed/NCBI
|
|
7
|
Neufert C, Becker C and Neurath MF: An
inducible mouse model of colon carcinogenesis for the analysis of
sporadic and inflammation-driven tumor progression. Nat Protoc.
2:1998–2004. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kanneganti M, Mino-Kenudson M and
Mizoguchi E: Animal models of colitis-associated carcinogenesis. J
Biomed Biotechnol. 2011:3426372011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luo K and Cao SS: Endoplasmic reticulum
stress in intestinal epithelial cell function and inflammatory
bowel disease. Gastroenterol Res Pract. 2015:3287912015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dicks N, Gutierrez K, Michalak M,
Bordignon V and Agellon LB: Endoplasmic reticulum stress, genome
damage, and cancer. Front Oncol. 5:112015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cao SS, Luo KL and Shi L: Endoplasmic
reticulum stress interacts with inflammation in human diseases. J
Cell Physiol. 231:288–294. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Parmar VM and Schröder M: Sensing
endoplasmic reticulum stress. Adv Exp Med Biol. 738:153–168. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gardner BM, Pincus D, Gotthardt K,
Gallagher CM and Walter P: Endoplasmic reticulum stress sensing in
the unfolded protein response. Cold Spring Harb Perspect Biol.
5:a0131692013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
McMillan DR, Gething MJ and Sambrook J:
The cellular response to unfolded proteins: Intercompartmental
signaling. Curr Opin Biotechnol. 5:540–545. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cox JS, Shamu CE and Walter P:
Transcriptional induction of genes encoding endoplasmic reticulum
resident proteins requires a transmembrane protein kinase. Cell.
73:1197–1206. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bertolotti A, Wang X, Novoa I, Jungreis R,
Schlessinger K, Cho JH, West AB and Ron D: Increased sensitivity to
dextran sodium sulfate colitis in IRE1beta-deficient mice. J Clin
Invest. 107:585–593. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Manie SN, Lebeau J and Chevet E: Cellular
mechanisms of endoplasmic reticulum stress signaling in health and
disease. 3. Orchestrating the unfolded protein response in
oncogenesis: An update. Am J Physiol Cell Physiol. 307:C901–C907.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Papandreou I, Denko NC, Olson M, Van
Melckebeke H, Lust S, Tam A, Solow-Cordero DE, Bouley DM, Offner F,
Niwa M and Koong AC: Identification of an Ire1alpha endonuclease
specific inhibitor with cytotoxic activity against human multiple
myeloma. Blood. 117:1311–1314. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yoshida H, Matsui T, Yamamoto A, Okada T
and Mori K: XBP1 mRNA is induced by ATF6 and spliced by IRE1 in
response to ER stress to produce a highly active transcription
factor. Cell. 107:881–891. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Koong AC, Chauhan V and Romero-Ramirez L:
Targeting XBP-1 as a novel anti-cancer strategy. Cancer Biol Ther.
5:756–759. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Welihinda AA, Tirasophon W and Kaufman RJ:
The cellular response to protein misfolding in the endoplasmic
reticulum. Gene Expr. 7:293–300. 1999.PubMed/NCBI
|
|
22
|
Martino MB, Jones L, Brighton B, Ehre C,
Abdulah L, Davis CW, Ron D, O'Neal WK and Ribeiro CM: The ER stress
transducer IRE1beta is required for airway epithelial mucin
production. Mucosal Immunol. 6:639–654. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen Ya and Brandizzi F: IRE1: ER stress
sensor and cell fate executor. Trends Cell Biol. 23:547–555. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang HS, Chen Y, Fan L, Xi QL, Wu GH, Li
XX, Yuan TL, He SQ, Yu Y, Shao ML, et al: The endoplasmic reticulum
stress sensor IRE1α in intestinal epithelial cells is essential for
protecting against colitis. J Biol Chem. 290:15327–15336. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Oikawa D, Kitamura A, Kinjo M and Iwawaki
T: Direct association of unfolded proteins with mammalian ER stress
sensor, IRE1β. PLoS One. 7:e512902012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nakamura D, Tsuru A, Ikegami K, Imagawa Y,
Fujimoto N and Kohno K: Mammalian ER stress sensor IRE1β
specifically down-regulates the synthesis of secretory pathway
proteins. FEBS Lett. 585:133–138. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iwawaki T, Hosoda A, Okuda T, Kamigori Y,
Nomura-Furuwatari C, Kimata Y, Tsuru A and Kohno K: Translational
control by the ER transmembrane kinase/ribonuclease IRE1 under ER
stress. Nat Cell Biol. 3:158–164. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tsuru A, Fujimoto N, Takahashi S, Saito M,
Nakamura D, Iwano M, Iwawaki T, Kadokura H, Ron D and Kohno K:
Negative feedback by IRE1β optimizes mucin production in goblet
cells. Proc Natl Acad Sci USA. 110:2864–2869. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Johansson ME, Phillipson M, Petersson J,
Velcich A, Holm L and Hansson GC: The inner of the two Muc2
mucin-dependent mucus layers in colon is devoid of bacteria. Proc
Natl Acad Sci USA. 105:15064–15069. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kawashima H: Roles of the gel-forming MUC2
mucin and its O-glycosylation in the protection against colitis and
colorectal cancer. Biol Pharm Bull. 35:1637–1641. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Van der Sluis M, De Koning BA, De Bruijn
AC, Velcich A, Meijerink JP, Van Goudoever JB, Büller HA, Dekker J,
Van Seuningen I, Renes IB and Einerhand AW: Muc2-deficient mice
spontaneously develop colitis, indicating that MUC2 is critical for
colonic protection. Gastroenterology. 131:117–129. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Velcich A, Yang W, Heyer J, Fragale A,
Nicholas C, Viani S, Kucherlapati R, Lipkin M, Yang K and
Augenlicht L: Colorectal cancer in mice genetically deficient in
the mucin Muc2. Science. 295:1726–1729. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jackson LN, Zhou Y, Qiu S, Wang Q and
Evers BM: Alternative medicine products as a novel treatment
strategy for inflammatory bowel disease. Am J Chin Med. 36:953–965.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Esworthy RS, Kim BW, Larson GP, Yip ML,
Smith DD, Li M and Chu FF: Colitis locus on chromosome 2 impacting
the severity of early-onset disease in mice deficient in GPX1 and
GPX2. Inflamm Bowel Dis. 17:1373–1386. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li H, Zhou Y, Zheng Y, Guo H, Gao L, Chen
P, Feng D, Wu L, Yang M, Qi Y, et al: The gastric mucosa from
patients infected with CagA+ or VacA+ Helicobacter pylori has a
lower level of dual oxidase-2 expression than uninfected or
infected with CagA-/VacA-H. pylori. Dig Dis Sci. 61:2328–2337.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Untergasser A, Cutcutache I, Koressaar T,
Ye J, Faircloth BC, Remm M and Rozen SG: Primer3-new capabilities
and interfaces. Nucleic Acids Res. 40:e1152012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wong ML and Medrano JF: Real-time PCR for
mRNA quantitation. Biotechniques. 39:75–85. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Thaker AI, Shaker A, Rao MS and Ciorba MA:
Modeling colitis-associated cancer with azoxymethane (AOM) and
dextran sulfate sodium (DSS). J Vis Exp:. (pii):
41002012.PubMed/NCBI
|
|
40
|
Tabas I and Ron D: Integrating the
mechanisms of apoptosis induced by endoplasmic reticulum stress.
Nat Cell Biol. 13:184–190. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen C and Zhang X: IRE1alpha-XBP1 pathway
promotes melanoma progression by regulating IL-6/STAT3 signaling. J
Transl Med. 15:422017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fujimoto T, Onda M, Nagai H, Nagahata T,
Ogawa K and Emi M: Upregulation and overexpression of human X-box
binding protein 1 (hXBP-1) gene in primary breast cancers. Breast
Cancer. 10:301–306. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Fujimoto T, Yoshimatsu K, Watanabe K,
Yokomizo H, Otani T, Matsumoto A, Osawa G, Onda M and Ogawa K:
Overexpression of human X-box binding protein 1 (XBP-1) in
colorectal adenomas and adenocarcinomas. Anticancer Res.
27:127–131. 2007.PubMed/NCBI
|
|
44
|
Jiang Y, Zhou Y, Zheng Y, Guo H, Gao L,
Chen P, Feng D, Qi R, Li X, Chang Y, et al: Expression of
inositol-requiring enzyme 1β is downregulated in colorectal cancer.
Oncol Lett. 13:1109–1118. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hetz C, Martinon F, Rodriguez D and
Glimcher LH: The unfolded protein response: Integrating stress
signals through the stress sensor IRE1α. Physiol Rev. 91:1219–1243.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lin JH, Li H, Yasumura D, Cohen HR, Zhang
C, Panning B, Shokat KM, Lavail MM and Walter P: IRE1 signaling
affects cell fate during the unfolded protein response. Science.
318:944–949. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lencer WI, DeLuca H, Grey MJ and Cho JA:
Innate immunity at mucosal surfaces: The IRE1-RIDD-RIG-I pathway.
Trends Immunol. 36:401–409. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kaser A, Adolph TE and Blumberg RS: The
unfolded protein response and gastrointestinal disease. Semin
Immunopathol. 35:307–319. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kaser A, Lee AH, Franke A, Glickman JN,
Zeissig S, Tilg H, Nieuwenhuis EE, Higgins DE, Schreiber S,
Glimcher LH and Blumberg RS: XBP1 links ER stress to intestinal
inflammation and confers genetic risk for human inflammatory bowel
disease. Cell. 134:743–756. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Romero-Ramirez L, Cao H, Regalado MP,
Kambham N, Siemann D, Kim JJ, Le QT and Koong AC: X box-binding
protein 1 regulates angiogenesis in human pancreatic
adenocarcinomas. Transl Oncol. 2:31–38. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yadav RK, Chae SW, Kim HR and Chae HJ:
Endoplasmic reticulum stress and cancer. J Cancer Prev. 19:75–88.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Coelho DS and Domingos PM: Physiological
roles of regulated Ire1 dependent decay. Front Genet. 5:762014.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Moradi Marjaneh R, Hassanian SM, Ghobadi
N, Ferns GA, Karimi A, Jazayeri MH, Nasiri M, Avan A and Khazaei M:
Targeting the death receptor signaling pathway as a potential
therapeutic target in the treatment of colorectal cancer. J Cell
Physiol. 233:6538–6549. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Iqbal J, Dai K, Seimon T, Jungreis R,
Oyadomari M, Kuriakose G, Ron D, Tabas I and Hussain MM: IRE1beta
inhibits chylomicron production by selectively degrading MTP mRNA.
Cell Metab. 7:445–455. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Dai K, Khatun I and Hussain MM: NR2F1 and
IRE1beta suppress microsomal triglyceride transfer protein
expression and lipoprotein assembly in undifferentiated intestinal
epithelial cells. Arterioscler Thromb Vasc Biol. 30:568–574. 2010.
View Article : Google Scholar : PubMed/NCBI
|