Introduction
Renal cell carcinoma (RCC), originates from cells in
the renal cortex and is the most common type of kidney cancer in
adults worldwide, with ~270,000 new cases and ~116,000
RCC-associated mortalities each year (1,2). RCC is
divided into three major subtypes, including clear cell RCC
(ccRCC), papillary RCC and chromophobe RCC (3). ccRCC is the most common and aggressive
of the three subtypes, and accounts for ~80–90% of all RCC cases
(4). Despite recent advances in
cancer treatment, the long-term prognosis of patients with advanced
ccRCC is poor, with a median survival rate of 1.5 years (5). Metastasis, lymph node recurrence and
cancer recurrence are the main factors associated with poor
prognosis in patients with ccRCC (6). In ~33% of cases, patients with ccRCC
are diagnosed with local or distant metastases due to its variable
clinical presentation (7). In
addition, there is a 40% recurrence rate in patients with ccRCC
following surgery (8,9). Therefore, identifying novel therapeutic
targets may improve the survival outcome and treatment response of
patients with advanced ccRCC.
MicroRNAs (miRNAs/miRs) are a group of endogenous,
short, non-coding RNAs ~18–23 nucleotides in length (10). MiRNAs regulate multiple biological
and pathological processes, including carcinogenesis and cancer
progression (11). miRNAs promote
mRNA degradation or inhibition of mRNA translation by binding to
the complementary binding sites in the 3′-untranslated region
(3′-UTR) of target genes (12).
Several studies have demonstrated that miRNA deregulation is common
in almost all types of human cancer (13–15).
Previous studies have identified several deregulated miRNAs
associated with ccRCC, whereby oncogenic miRNAs are upregulated and
tumor suppressor miRNAs are downregulated (16–18).
Aberrantly expressed miRNAs are involved in the pathogenesis of
ccRCC and are involved in numerous processes, including cell
proliferation, cell cycle progression, apoptosis, metastasis,
epithelial-to-mesenchymal transition and chemoresistance (19–21).
miRNAs may therefore be used as potential biomarkers to improve
early diagnosis and prognosis of patients with ccRCC.
Previous studies have demonstrated that miR-508 is
abnormally expressed in several types of human cancer, including
ovarian (22) and colorectal cancer
(23) and esophageal squamous cell
carcinoma (24). An additional study
revealed that miR-508 is downregulated in RCC and may be involved
in regulating cell migration and apoptosis (25). Therefore, the aim of the current
study was to investigate the expression pattern and biological
function of miR-508 in the pathogenesis of ccRCC, as well as
investigate the underlying molecular mechanisms of miR-508 in the
development and progression of ccRCC.
Materials and methods
Human tissue samples
A total of 21 ccRCC tissue and paired adjacent
normal tissue samples were collected from patients that had
undergone surgical resection at the Lanzhou University Second
Hospital (Lanzhou, China) between May 2015 and February 2017. None
of the patients had received radiotherapy or chemotherapy treatment
prior to surgical resection. All tissue samples were taken
immediately after tumor resection, frozen in liquid nitrogen and
stored at −80°C. The current study was approved by the Ethics
Committee of Lanzhou University Second Hospital and written
informed consent was obtained from each patient. The
clinicopathological features of patients with ccRCC are summarized
in Table I.
 | Table I.Clinicopathological characteristics
of patients with ccRCC. |
Table I.
Clinicopathological characteristics
of patients with ccRCC.
| Patient no. | Gender | Age (years) | TNM stage |
|---|
| 1 | M | 47 | T1aN0M0 |
| 2 | M | 51 | T2bN0M0 |
| 3 | M | 44 | T1bN0M0 |
| 4 | M | 26 | T3aN1M0 |
| 5 | F | 58 | T2bN0M0 |
| 6 | M | 63 | T2bN0M0 |
| 7 | M | 68 | T1aN0M0 |
| 8 | F | 53 | T2aN0M0 |
| 9 | F | 57 | T1aN0M0 |
| 10 | M | 64 | T1aN0M0 |
| 11 | M | 49 | T2bN0M0 |
| 12 | M | 63 | T1aN0M0 |
| 13 | M | 56 | T1aN0M0 |
| 14 | F | 55 | T3aN1M0 |
| 15 | M | 72 | T2bN0M0 |
| 16 | F | 64 | T2aN0M0 |
| 17 | M | 68 | T2bN0M0 |
| 18 | M | 53 | T2bN0M0 |
| 19 | F | 47 | T1bN0M0 |
| 20 | F | 55 | T2aN0M0 |
| 21 | M | 60 | T2aN0M0 |
Cell culture
The normal human renal cell line HK-2, the papillary
RCC cell lines Caki-2 and ACHN, and two ccRCC cell lines 786-O and
A498 were purchased from the Cell Bank of the Chinese Academy of
Sciences (Shanghai, China). Cells were cultured in Dulbecco's
modified Eagle's medium (DMEM) supplemented with 10% fetal bovine
serum (FBS), 100 U/ml penicillin and 100 mg/ml streptomycin (all
purchased from Invitrogen; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) and maintained at 37°C in a 5% CO2-humidified
incubator.
Cell transfection
miR-508 mimics, negative control miRNA mimics
(miR-NC), small interfering RNA (siRNA) targeting the expression of
zinc finger E-box-binding homeobox 1 (ZEB1 siRNA) and negative
control siRNA (NC siRNA) were synthesized by Shanghai GenePharma
Co., Ltd. (Shanghai, China). The miR-508 mimics sequence was
5′-UGAUUGUAGCCUUUUGGAGUAGA-3′ and the miR-NC sequence was
5′-UUCUCCGAACGUGUCACGUTT-3′. The ZEB1 siRNA sequence was
5′-GUCGCUACAAACAGUUGUATT-3′ and the NC siRNA sequence was
5′-UUCUCCGAACGUGUCACGUTT-3′. The ZEB1-overexpression plasmid,
pcDNA3.1-ZEB1 and empty pcDNA3.1 plasmid were synthesized by
GeneCopoeia, Inc., (Rockville, MD, USA). Cells were seeded into
six-well plates at a density of 6×105 cells/well.
miR-508 mimics (100 pmol), miR-NC (100 pmol), ZEB1 siRNA (100
pmol), NC siRNA (100 pmol), pcDNA3.1-ZEB1 (4 µg) or empty pcDNA3.1
plasmids (4 µg) were transfected into cells using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. At 6 h
following incubation at 37°C in a 5% CO2-humidified
incubator, the medium was replaced with complete culture medium.
Reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) and cell proliferation assay was performed at 24 h
post-transfection. Transwell invasion assay was performed in
transfected cells following 48 h of incubation. Following 72 h
culture, western blot analysis was utilized for the measurement of
protein expression.
RT-qPCR
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), according to on the manufacturer's protocol. To examine the
expression of miR-508, total miRNA was first reverse transcribed
into cDNA using the TaqMan® MicroRNA Reverse
Transcription kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The temperature protocol for reverse transcription was as
follows: 16°C for 30 min, 42°C for 30 min and 85°C for 5 min. qPCR
was subsequently performed using the TaqMan MicroRNA Assay kit
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The
temperature protocol for qPCR were as follows: 50°C for 2 min, 95°C
for 10 min; 40 cycles of denaturation at 95°C for 15 sec; and
annealing/extension at 60°C for 60 sec. To analyze ZEB1 mRNA
expression levels, total RNA was first reverse transcribed into
cDNA using the PrimeScript RT Reagent kit (Takara Biotechnology,
Co., Ltd., Dalian, China). The temperature protocol for reverse
transcription was as follows: 37°C for 15 min and 85°C for 5 sec.
qPCR was subsequently performed using the SYBR Premix Ex Taq™ kit
(Takara Biotechnology, Co., Ltd.). The temperature protocol for
qPCR was as follows: 5 min at 95°C, followed by 40 cycles of 95°C
for 30 sec and 65°C for 45 sec. ZEB1 and miR-508 expression was
quantified using the 2−ΔΔCq method and normalized to the
internal reference gene GAPDH or U6 small nuclear RNA, respectively
(26). The primer sequences used
were as follows: miR-508 forward, 5′-TTCAAGAGACATGAGTGAC-3′ and
reverse, 5′-TCTCTTGAACATGAGTGACG-3′; U6 forward,
5′-TGCGGGTGCTCGCTTCGGCAGC-3′ and reverse,
5′-CCAGTGCAGGGTCCGAGGT-3′; ZEB1 forward, 5′-AAGTGGCGGTAGATGGTA-3′
and reverse, 5′-TTGTAGCGACTGGATTTT-3′; and GAPDH forward,
5′-AACGGATTTGGTCGTATTG-3′ and reverse,
5′-GGAAGATGGTGATGGGATT-3′.
Cell proliferation assay
The proliferative ability of ccRCC cells was
analyzed using the cell counting kit-8 assay (CCK-8; Dojindo
Molecular Technologies, Inc., Kumamoto, Japan). Briefly,
transfected cells were collected and seeded into 96-well plates at
a density of 3×103 cells/well and cultured for 0, 24, 48
and 72 h at 37°C. Following incubation, 10 µl CCK-8 reagent was
then added to each well and the cells were incubated for an
additional 2 h at 37°C. Cell proliferation was determined by
measuring the absorbance at a wavelength of 450 nm using the
SpectraMax Microplate® Spectrophotometer (Molecular
Devices LLC, Sunnyvale, CA, USA).
Transwell invasion assay
ccRCC cell invasion was examined using an in
vitro Transwell invasion chamber assay using Transwell™
polycarbonate membrane inserts for 24-well plates containing 8-µm
pores (Corning Inc., Corning, NY, USA) precoated with
Matrigel® (BD Biosciences, San Jose, CA, USA). Following
transfection, 5×104 cells in serum-free DMEM were added
to the upper chamber of the Transwell insert. In the lower chamber,
600 µl DMEM supplemented with 10% FBS was used as a
chemo-attractant, and the plates were incubated at 37°C in a 5%
CO2-humidified incubator for 24 h. Non-invasive cells
remaining on the polycarbonate membrane were carefully removed and
invaded cells were fixed with 100% methanol at room temperature for
30 min and stained with 0.1% crystal violet at room temperature for
30 min. The invasive capacity of ccRCC cells was quantified by
counting the number of invaded cells in five randomly selected
visual fields/chamber under an inverted light microscope (×200
magnification).
Bioinformatics analysis and
dual-luciferase reporter assay
TargetScan (http://www.targetscan.org) and miRDB (http://mirdb.org) databases were used to identify
putative targets of miR-508. ZEB1 was predicted as a potential
target gene of miR-508. The wild-type (wt) and mutant (mut)
fragments of the ZEB1 3′-UTR (synthesized by Shanghai GenePharma
Co., Ltd.) were amplified by PCR and cloned into the firefly
luciferase-expressing pmirGLO luciferase reporter plasmid (Promega
Corporation, Madison, WI, USA). Cells were seeded into 24-well
plates and incubated at 37°C in a CO2-humidified
incubator for 24 h prior to transfection. Cells were subsequently
co-transfected with miR-508 mimics or miR-NC and wt or mut 3′-UTR
ZEB1 luciferase reporter plasmids using Lipofectamine®
2000. Following incubation for 48 h, luciferase activities were
detected using the Dual-Luciferase Reporter assay system (Promega
Corporation), according to the manufacturer's protocol. Firefly
luciferase activity was normalized to that of Renilla
luciferase activity.
Western blot analysis
Total protein was extracted from tissues or cells
using ice-cold radioimmunoprecipitation assay buffer
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) supplemented with
proteinase inhibitor cocktail (Roche Applied Science, Penzberg,
Germany). Total protein was quantified using a bicinchoninic acid
assay kit (Pierce; Thermo Fisher Scientific, Inc.) and equal
quantities of protein (30 µg/lane) were separated via SDS-PAGE on a
10% gel. The separated proteins were transferred onto
polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA,
USA) and blocked for 1 h at room temperature with 5% skimmed milk
in Tris-buffered saline containing 0.05% Tween-20. The membranes
were incubated with the following primary antibodies: Mouse
anti-human monoclonal ZEB1 (dilution, 1:1,000; cat. no. sc-81428)
or mouse anti-human monoclonal GAPDH (dilution, 1:1,000; cat. no.
sc-47724; both Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
overnight at 4°C. Following primary incubation, the membranes were
subsequently incubated with goat-anti mouse horseradish
peroxidase-labeled secondary antibody (dilution, 1:5,000; cat. no.
sc-516102; Santa Cruz Biotechnology, Inc.) at room temperature for
2 h. Protein bands were visualized using enhanced chemiluminescence
detection reagent (EMD Millipore), according to the manufacturer's
protocol. Protein expression was quantified by normalizing target
protein expression to the GAPDH loading control. Quantity One
software version 4.62 (Bio-Rad Laboratories, Inc., Hercules, CA,
USA) was used for densitometry analysis.
Statistical analysis
The results are presented as the mean ± standard
deviation from at least three independent experiments. All
statistical analyses were performed using SPSS software (version
19.0; IBM Corp., Armonk, NY, USA). The statistical significance of
differences between two groups was analyzed using a two-tailed,
paired Student's t-test. One-way analysis of variance followed by
the Student-Newman-Keuls post hoc test was used to analyze
differences among multiple groups. The association between miR-508
and ZEB1 mRNA expression in ccRCC tissue samples was determined
using Spearman's correlation analysis. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-508 expression is downregulated in
ccRCC tissue samples and cell lines
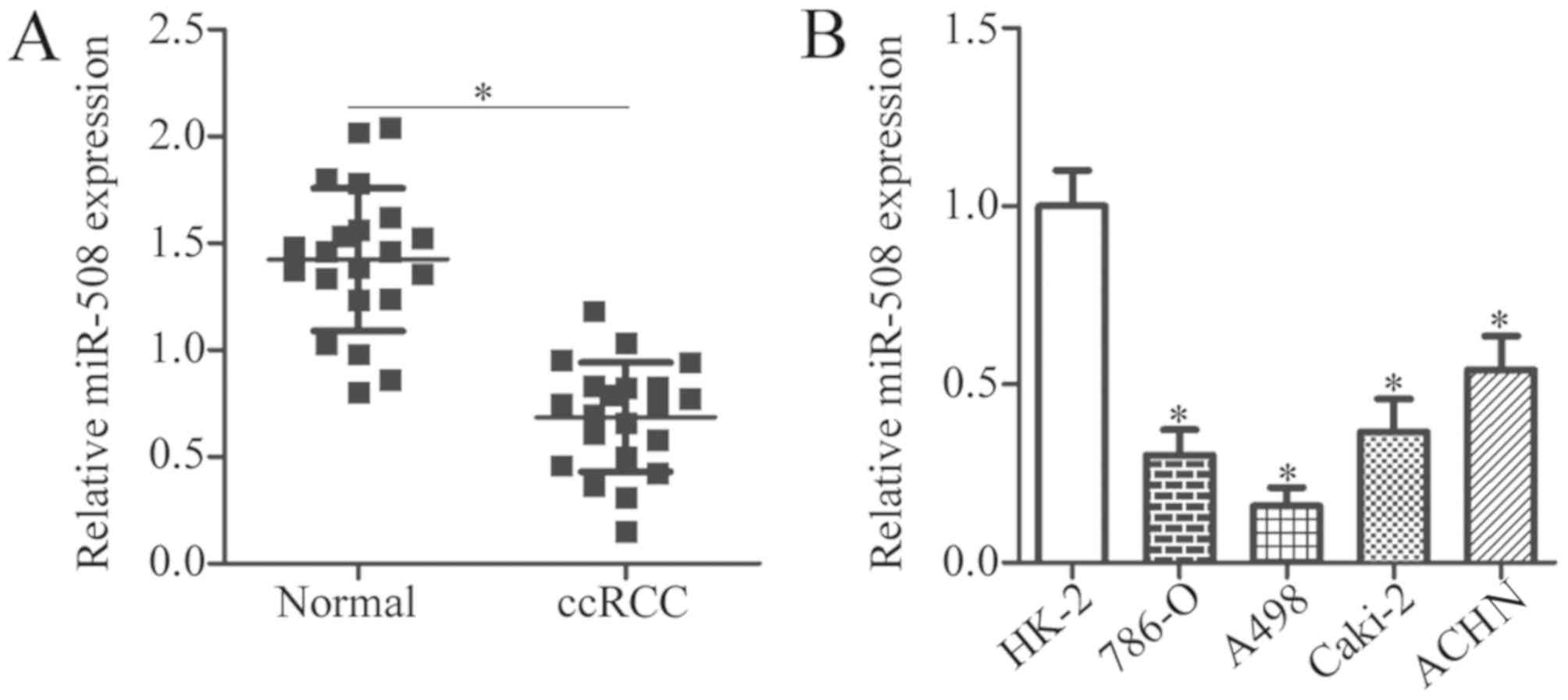
To investigate the expression pattern of miR-508 in
ccRCC, the expression levels of miR-508 in ccRCC tissue samples and
adjacent normal tissue samples from patients was determined by
RT-qPCR analysis. The miR-508 expression level was significantly
decreased in ccRCC tissue samples when compared with adjacent
normal tissue samples (P<0.05; Fig.
1A). RT-qPCR was then used to examine miR-508 expression in two
ccRCC cell lines (786-O and A498), two papillary RCC cell lines
(Caki-2 and ACHN), and the normal human renal cell line HK-2. The
expression of miR-508 was significantly decreased in ccRCC and
papillary RCC cell lines when compared with the normal human renal
cell line (P<0.05; Fig. 1B).
These results suggest that downregulation of miR-508 may be
associated with ccRCC progression.
miR-508 overexpression suppresses the
proliferation and invasion of ccRCC cells
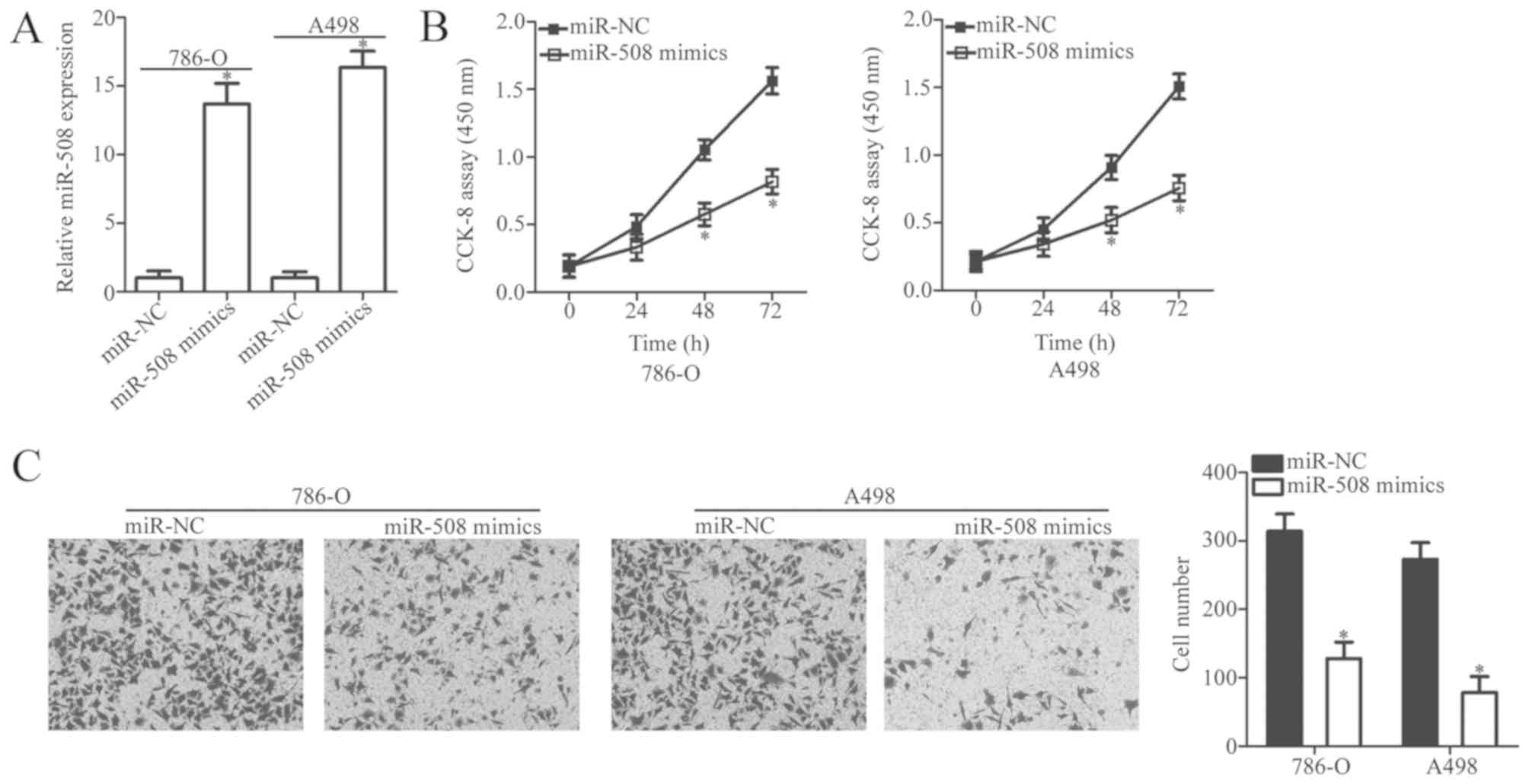
To investigate the biological function of miR-508 in
ccRCC progression, ccRCC cell lines 786-O and A498 (as they
exhibited the lowest miR-508 expression) were transfected with
miR-508 mimics or miR-NC. The relative expression of miR-508 was
significantly increased in ccRCC cells following transfection with
miR-508 mimics compared with miR-NC (P<0.05; Fig. 2A). A CCK-8 assay was then used to
examine the effect of miR-508 overexpression on ccRCC cell
proliferation. The results demonstrated that miR-508 overexpression
significantly inhibited the proliferative ability of 786-O and A498
cells when compared with the negative controls (P<0.05; Fig. 2B). Subsequently, the in vitro
Transwell invasion assay demonstrated that miR-508 overexpression
significantly inhibited the invasive capacity of 786-O and A498
cells compared with the negative controls (P<0.05; Fig. 2C). Taken together, these results
suggest that miR-508 may serve a tumor-suppressive role in the
development of ccRCC.
ZEB1 is a direct target gene of
miR-508 in ccRCC cells
It is well established that miRNAs directly target
mRNAs of target genes, thereby exerting their role as
post-transcriptional regulators (12). To further investigate the mechanisms
underlying the tumor suppressive role of miR-508 in ccRCC in the
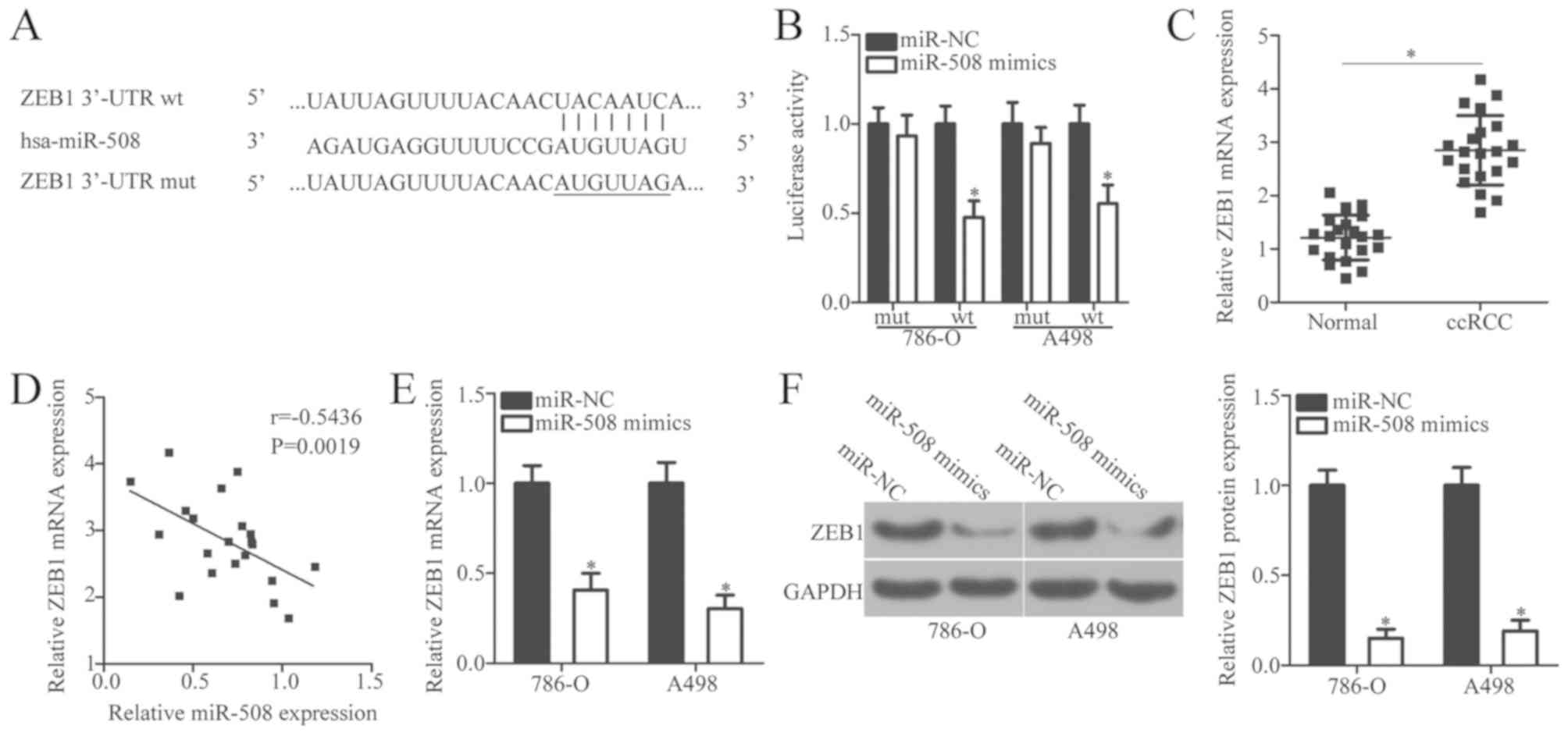
present study, putative targets of miR-508 were examined by
bioinformatics analysis. TargetScan and miRDB databases identified
ZEB1, which contains a 3′-UTR sequence complementary to the seed
sequence of miR-508 (Fig. 3A).
Therefore, ZEB1 was selected for verification using a
dual-luciferase reporter assay. This assay was used to confirm
whether miR-508 binds directly to the partially complimentary
sequence within the 3′-UTR of ZEB1 in ccRCC cells. The results
demonstrated that miR-508 overexpression significantly decreased
the luciferase activity of the reporter plasmid containing the wt
miR-508 binding site, whereas co-transfection with the mut miR-508
binding site plasmid demonstrated no effect on luciferase activity
(P<0.05; Fig. 3B).
To further understand the association between
miR-508 and ZEB1 in ccRCC, the expression of ZEB1 was analyzed
using RT-qPCR in ccRCC and normal adjacent tissue samples from
patients with ccRCC. The mRNA levels of ZEB1 were significantly
increased in ccRCC tissue samples when compared with adjacent
normal tissue samples (P<0.05; Fig.
3C). In addition, Spearman's correlation analysis indicated a
statistically significant negative correlation between miR-508 and
ZEB1 mRNA expression levels in ccRCC tissue samples (r=−0.5436,
P=0.0019; Fig. 3D). To investigate
whether ZEB1 expression was regulated by miR-508 in ccRCC cells,
ZEB1 expression levels were determined by RT-qPCR in 786-O and A498
cells transfected with miR-508 mimics or miR-NC. The mRNA and
protein expression levels of ZEB1 significantly decreased following
miR-508 overexpression compared with the negative controls
(P<0.05; Fig. 3E and F). These
results suggest that ZEB1 may be a direct target of miR-508 in
ccRCC cells.
Inhibition of ZEB1 suppresses ccRCC
cell proliferation and invasion
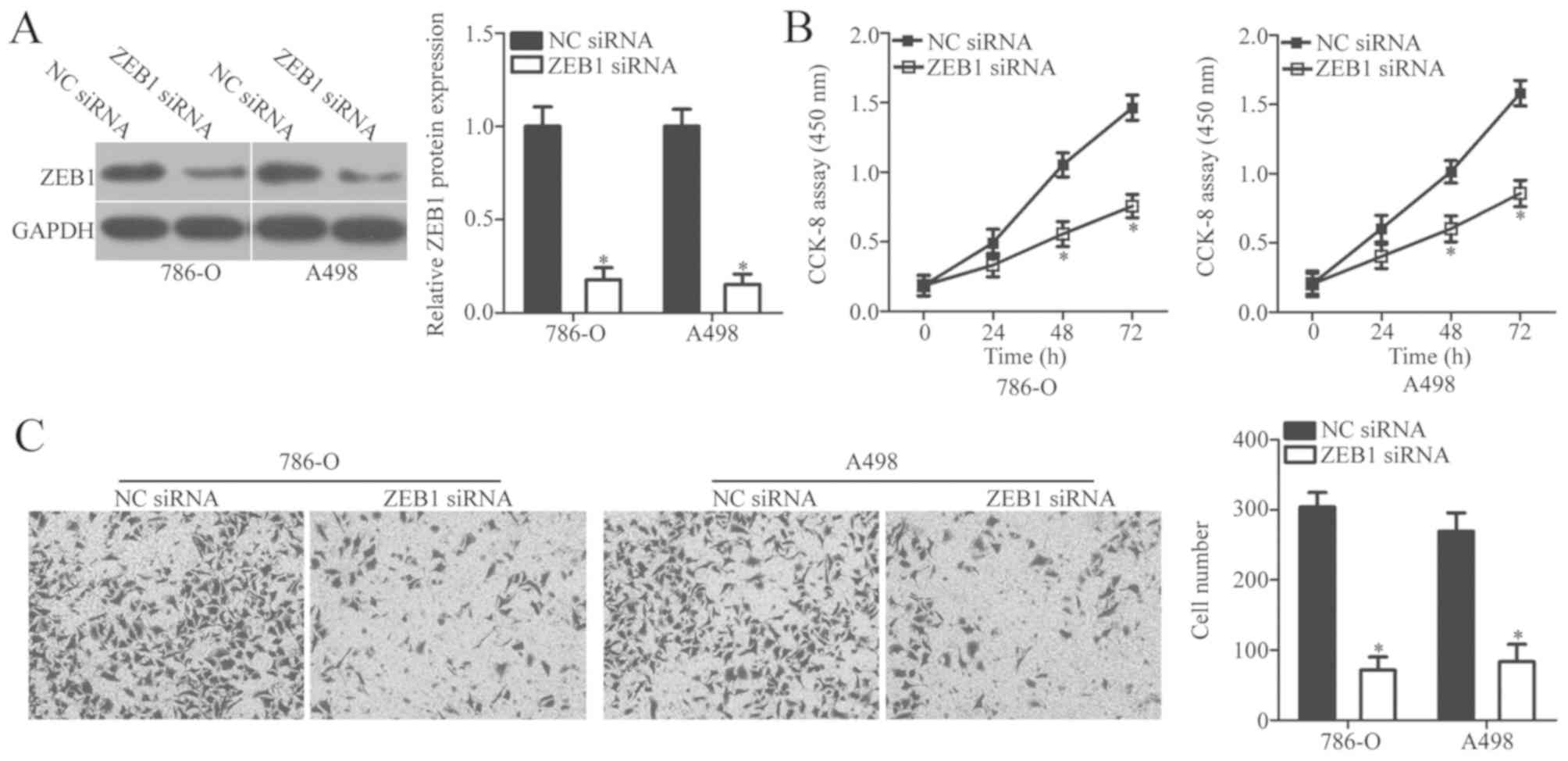
To examine the biological role of ZEB1 in ccRCC,
786-O and A498 cells were transfected with ZEB1 siRNA or NC siRNA.
Endogenous ZEB1 protein expression was significantly decreased in
ccRCC cells following transfection with ZEB1 siRNA when compared
with NC siRNA (P<0.05; Fig. 4A).
CCK-8 and in vitro Transwell invasion assays were used to
examine the effect of ZEB1 knockdown on ccRCC cell proliferation
and invasion, respectively. The results demonstrated that ZEB1
knockdown significantly inhibited the proliferative and invasive
ability of 786-O and A498 cells when compared with the negative
controls (P<0.05; Fig. 4B and C).
Taken together, these results demonstrated that ZEB1 knockdown
exerts a functional role similar to that of miR-508 overexpression
in ccRCC cells, which suggests that ZEB1 may be a functional target
of miR-508 in ccRCC.
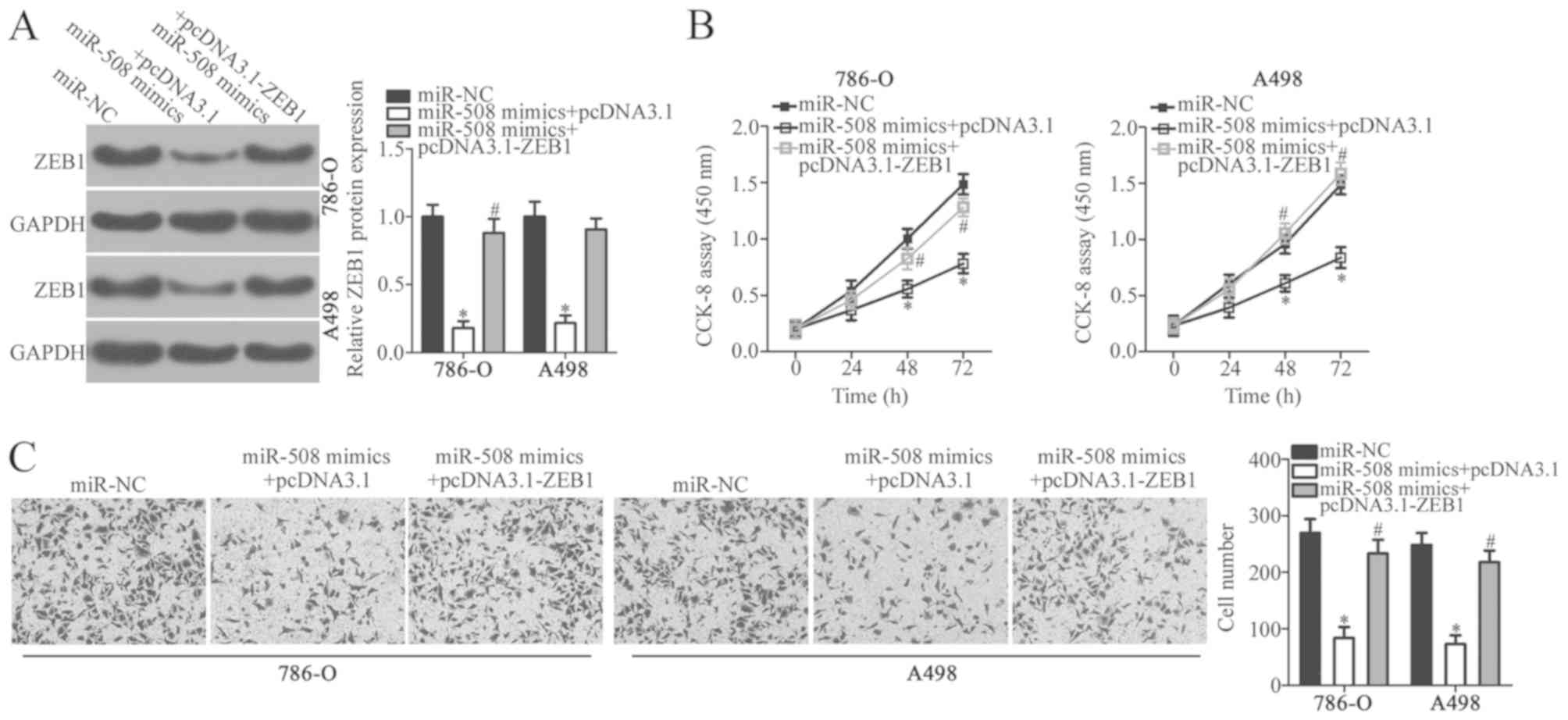
Restoration of ZEB1 expression
reverses the suppressive effects of miR-508 on the malignant
phenotype of ccRCC cells
As ZEB1 was identified as a direct target of miR-508
in ccRCC, rescue assays were performed to determine whether ZEB1
knockdown was responsible for the miR-508-induced inhibitory
effects on ccRCC cell proliferation and invasion. ccRCC cell lines
786-O and A498 were co-transfected with miR-508 mimics and pcDNA3.1
or pcDNA3.1-ZEB1. Western blot analysis confirmed that the relative
protein expression levels of ZEB1 were significantly increased in
ccRCC cells following co-transfection with miR-508 mimics and
pcDNA3.1-ZEB1 compared with miR-508 mimics and the pcDNA3.1
negative control (P<0.05; Fig.
5A). Furthermore, functional analysis demonstrated that
restoration of ZEB1 expression significantly reversed the
inhibitory effects of miR-508 on ccRCC cell proliferation and
invasion (P<0.05; Fig. 5B and C).
These results suggest that miR-508 may serve a tumor suppressive
role in ccRCC and that the inhibitory effects of this miRNA may be,
in part, mediated by regulating ZEB1 expression.
Discussion
A number of recent studies have identified a variety
of deregulated miRNAs in ccRCC, including miR-224 (27), miR-502 (28), miR-543 (18) and miR-645 (29). miRNAs are involved in several
biological and pathological processes, including ccRCC occurrence
and development (30–32). Previous studies have also identified
several deregulated miRNAs in ccRCC, whereby oncogenic miRNAs are
upregulated and tumor suppressor miRNAs are downregulated (17,33).
Therefore, understanding the expression pattern and biological
function of miRNAs in ccRCC, as well as the underlying mechanisms
of miRNAs in the development and progression of ccRCC, may be
useful for the identification of novel therapeutic targets in the
treatment of patients with ccRCC. In the current study, miR-508
expression in ccRCC tissue samples and cell lines was examined and
the regulatory effects of miR-508 in the development of ccRCC in
vitro were examined. Furthermore, the underlying molecular
mechanisms of miR-508 in ccRCC cells were investigated. The current
study identified miR-508 as a putative novel biomarker in the
diagnosis of ccRCC, as well as a potentially novel and efficient
therapeutic target for the treatment of patients with ccRCC.
Previous studies have demonstrated that miR-508
expression is downregulated in ovarian cancer (22) and colorectal cancer (23) compared with normal tissues of the
same type. By contrast, miR-508 was observed to be upregulated in
esophageal squamous cell carcinoma compared with normal esophageal
tissues (24). Patients with
esophageal squamous cell carcinoma expressing high levels of
miR-508 exhibited worse survival rates when compared with patients
with low miR-508 expression (24).
However, the expression status of miR-508 in ccRCC requires further
investigation. In the present study, miR-508 expression was
significantly decreased in ccRCC tissue samples and cell lines when
compared with adjacent normal tissue samples and the normal human
renal cell line HK-2, respectively. These conflicting results
suggest that miR-508 may exert tissue-specific expression patterns,
and miR-508 may therefore present a potential diagnostic biomarker
for patients with these types of cancer.
Previous studies have demonstrated that aberrant
miR-508 expression may contribute to the development and
progression of several types of human cancer (22–24).
Upregulation of miR-508 suppressed cell proliferation, metastasis
and epithelial-to-mesenchymal transition in ovarian cancer via the
mitogen-activated protein kinase 1 signaling pathway (22). In addition, Yan et al
(23) reported that ectopic
expression of miR-508 in colorectal cancer cells attenuated
epithelial-to-mesenchymal transition, stemness and metastasis in
vitro and in vivo. By contrast, miR-508 exerts an
oncogenic role in esophageal squamous cell carcinoma by promoting
tumorigenicity both in vivo and in vitro (24). These studies suggest that the role of
miR-508 in cancer development and progression may be tissue
specific. miR-508 may function as tumour suppressor or promoter in
different types of human cancer, depending on the characteristics
of corresponding target genes (34).
However, the potential role of miR-508 in ccRCC remains unknown. In
the current study, functional experiments indicated that miR-508
overexpression inhibited the proliferation and invasion of ccRCC
cells. These findings suggest that miR-508 may serve as a potential
therapeutic target in the treatment of patients with ccRCC.
miRNAs regulate several biological and pathological
processes, including tumorigenesis, by binding to specific sites
within the 3′-UTR of target mRNA sequences to inhibit their
translation and expression (12). In
the current study, the underlying mechanism of miR-508 in ccRCC
cells was investigated. Bioinformatics analysis identified ZEB1,
which contains a 3′-UTR sequence complementary to the seed sequence
of miR-508. Therefore, ZEB1 was considered a potential target gene
of miR-508 in the present study. A dual-luciferase reporter assay
was used to confirm a direct interaction between the 3′-UTR of ZEB1
and miR-508 in ccRCC cells. RT-qPCR and western blot analyses
revealed that the mRNA and protein expression levels of ZEB1 were
negatively regulated by miR-508 in ccRCC cells. In addition, the
current study demonstrated that ZEB1 expression was significantly
increased in ccRCC tissue samples, and ZEB1 expression was
inversely correlated with miR-508 expression in these tissue
samples. Finally, restoration of ZEB1 expression reversed the
inhibitory effect of miR-508 on the malignant phenotype of ccRCC
cells. Taken together, these results suggest that ZEB1 is a direct
and functional downstream target of miR-508 in ccRCC cells.
ZEB1, located on the short arm of human chromosome
10, is a member of the ZEB family of transcription factors
(35). Previous studies have
demonstrated that ZEB1 is upregulated in several types of cancer,
including lung (36), gastric
(37), colorectal (38) and endometrial cancers (39). Overexpression of ZEB1 has also been
identified in ccRCC, and ZEB1 overexpression was significantly
correlated with tumor grade, tumor, node and metastasis stage,
lymph node metastasis and distant metastases (40). In addition, patients with ccRCC
expressing high ZEB1 expression levels exhibited worse overall and
progression-free survival when compared with low ZEB1 expression
(40). Furthermore, ZEB1 was
demonstrated to be involved in several biological processes in
ccRCC development and progression (41). The current study demonstrated that
miR-508 directly targets ZEB1 and suppresses ccRCC cell
proliferation and invasion. The miR-508-ZEB1 axis may therefore be
a novel and efficient therapeutic target, which could be used to
inhibit the rapid growth and metastasis of ccRCC.
In conclusion, to the best of the authors'
knowledge, the current study is the first to demonstrate that
miR-508 is downregulated in ccRCC and that miR-508 may inhibit the
development of ccRCC by directly targeting ZEB1. The miR-508-ZEB1
axis may present a potential therapeutic target in the treatment of
patients with ccRCC patients. However, the current study has
several limitations that will need to be addressed in future
studies. The effect of miR-508 in cell apoptosis, in vivo
tumor growth, angiogenesis, energy metabolism and drug resistance
were not examined. Due to the relatively small sample size, the
correlation between clinicopathological features and miR-508
expression in ccRCC tissue samples was also not examined.
Furthermore, future studies will be required to validate miR-508 as
a potential biomarker in the early diagnosis and prognosis of
patients with ccRCC.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundations of China (grant no. 31700720),
Project of Chengguan District Science and Technology Bureau of
Lanzhou city (grant no. 2017SHFZ0029) and the Doctoral Research
Fund of the Second Hospital of Lanzhou University (grant no.
ynbskyjj2015-1-13).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZY and WW designed the study. WW and WH performed
RT-qPCR, CCK-8 and in vitro Transwell invasion assays. YW
performed the dual-luciferase reporter assay. JY performed western
blotting and statistical analysis. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Lanzhou University Second Hospital (Lanzhou, China).
The present study was performed in accordance with the Declaration
of Helsinki and the guidelines of the Ethics Committee of Lanzhou
University Second Hospital. Written informed consent was obtained
from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Liu Y, Han X, Yu Y, Ding Y, Ni C, Liu W,
Hou X, Li Z, Hou J, Shen D, et al: A genetic polymorphism affects
the risk and prognosis of renal cell carcinoma: Association with
follistatin-like protein 1 expression. Sci Rep. 6:266892016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Garcia JA, Cowey CL and Godley PA: Renal
cell carcinoma. Curr Opin Oncol. 21:266–271. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Patard JJ, Leray E, Rioux-Leclercq N,
Cindolo L, Ficarra V, Zisman A, De La Taille A, Tostain J, Artibani
W, Abbou CC, et al: Prognostic value of histologic subtypes in
renal cell carcinoma: A multicenter experience. J Clin Oncol.
23:2763–2771. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nerich V, Hugues M, Paillard MJ, Borowski
L, Nai T, Stein U, Nguyen Tan Hon T, Montcuquet P, Maurina T,
Mouillet G, et al: Clinical impact of targeted therapies in
patients with metastatic clear-cell renal cell carcinoma. Onco
Targets Ther. 7:365–374. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu D, Li M, Wang L, Zhou Y, Zhou J, Pan H
and Qu P: microRNA145 inhibits cell proliferation, migration and
invasion by targeting matrix metallopeptidase-11 in renal cell
carcinoma. Mol Med Rep. 10:393–398. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Crispen PL, Breau RH, Allmer C, Lohse CM,
Cheville JC, Leibovich BC and Blute ML: Lymph node dissection at
the time of radical nephrectomy for high-risk clear cell renal cell
carcinoma: Indications and recommendations for surgical templates.
Eur Urol. 59:18–23. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Flanigan RC, Campbell SC, Clark JI and
Picken MM: Metastatic renal cell carcinoma. Curr Treat Options
Oncol. 4:385–390. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Breda A, Konijeti R and Lam JS: Patterns
of recurrence and surveillance strategies for renal cell carcinoma
following surgical resection. Expert Rev Anticancer Ther.
7:847–862. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang YQ and Chen J: Predictive role of
vascular endothelial growth factor polymorphisms in the survival of
renal cell carcinoma patients. Genet Mol Res. 13:5011–5017. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hayes J, Peruzzi PP and Lawler S:
MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol
Med. 20:460–469. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bryzgunova OE, Konoshenko MY and Laktionov
PP: MicroRNA-guided gene expression in prostate cancer: Literature
and database overview. J Gene Med. 20:e30162018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pratap P, Raza ST, Abbas S and Mahdi F:
MicroRNA-associated carcinogenesis in lung carcinoma. J Cancer Res
Ther. 14:249–254. 2018.PubMed/NCBI
|
|
15
|
Vannini I, Fanini F and Fabbri M: Emerging
roles of microRNAs in cancer. Curr Opin Genet Dev. 48:128–133.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun X, Lou L, Zhong K and Wan L:
MicroRNA-451 regulates chemoresistance in renal cell carcinoma by
targeting ATF-2 gene. Exp Biol Med (Maywood). 242:1299–1305. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xiao W, Lou N, Ruan H, Bao L, Xiong Z,
Yuan C, Tong J, Xu G, Zhou Y, Qu Y, et al: Mir-144-3p promotes cell
proliferation, metastasis, sunitinib resistance in clear cell renal
cell carcinoma by downregulating ARID1A. Cell Physiol Biochem.
43:2420–2433. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang F, Ma J, Tang Q, Zhang W, Fu Q, Sun
J, Wang H and Song B: MicroRNA-543 promotes the proliferation and
invasion of clear cell renal cell carcinoma cells by targeting
Krüppel-like factor 6. Biomed Pharmacother. 97:616–623. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He YH, Chen C and Shi Z: The biological
roles and clinical implications of microRNAs in clear cell renal
cell carcinoma. J Cell Physiol. 233:4458–4465. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jaswani P, Prakash S, Dhar A, Sharma RK,
Prasad N and Agrawal S: MicroRNAs involvement in renal
pathophysiology: A bird's eye view. Indian J Nephrol. 27:337–341.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ellinger J, Gevensleben H, Muller SC and
Dietrich D: The emerging role of non-coding circulating RNA as a
biomarker in renal cell carcinoma. Expert Rev Mol Diagn.
16:1059–1065. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hong L, Wang Y, Chen W and Yang S:
MicroRNA-508 suppresses epithelial-mesenchymal transition,
migration, and invasion of ovarian cancer cells through the
MAPK1/ERK signaling pathway. J Cell Biochem. 119:7431–7440. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yan TT, Ren LL, Shen CQ, Wang ZH, Yu YN,
Liang Q, Tang JY, Chen YX, Sun DF, Zgodzinski W, et al: miR-508
defines the stem-like/mesenchymal subtype in colorectal cancer.
Cancer Res. 78:1751–1765. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin C, Liu A, Zhu J, Zhang X, Wu G, Ren P,
Wu J, Li M, Li J and Song L: miR-508 sustains phosphoinositide
signalling and promotes aggressive phenotype of oesophageal
squamous cell carcinoma. Nat Commun. 5:46202014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhai Q, Zhou L, Zhao C, Wan J, Yu Z, Guo
X, Qin J, Chen J and Lu R: Identification of miR-508-3p and
miR-509-3p that are associated with cell invasion and migration and
involved in the apoptosis of renal cell carcinoma. Biochem Biophys
Res Commun. 419:621–626. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fujii N, Hirata H, Ueno K, Mori J, Oka S,
Shimizu K, Kawai Y, Inoue R, Yamamoto Y, Matsumoto H, et al:
Extracellular miR-224 as a prognostic marker for clear cell renal
cell carcinoma. Oncotarget. 8:109877–109888. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang S, Guo Z, Xu J, Wang J, Zhang J, Cui
L, Zhang H, Liu Y and Bai Y: miR-502-mediated histone
methyltransferase SET8 expression is associated with clear cell
renal cell carcinoma risk. Oncol Lett. 14:7131–7138.
2017.PubMed/NCBI
|
|
29
|
Chen J, Shu Y, Yu Q and Shen W:
MicroRNA-645 promotes cell metastasis and proliferation of renal
clear cell carcinoma by targeting GK5. Eur Rev Med Pharmacol Sci.
21:4557–4565. 2017.PubMed/NCBI
|
|
30
|
Ding D, Zhang Y, Wen L, Fu J, Bai X, Fan
Y, Lin Y, Dai H, Li Q, Zhang Y and An R: MiR-367 regulates cell
proliferation and metastasis by targeting metastasis-associated
protein 3 (MTA3) in clear-cell renal cell carcinoma. Oncotarget.
8:63084–63095. 2017.PubMed/NCBI
|
|
31
|
Fan Y, Ma X, Li H, Gao Y, Huang Q, Zhang
Y, Bao X, Du Q, Luo G, Liu K, et al: miR-122 promotes metastasis of
clear-cell renal cell carcinoma by downregulating Dicer. Int J
Cancer. 142:547–560. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu F, Wu L, Wang A, Xu Y, Luo X, Liu X,
Hua Y, Zhang D, Wu S, Lin T, et al: MicroRNA-138 attenuates
epithelial-to-mesenchymal transition by targeting SOX4 in clear
cell renal cell carcinoma. Am J Transl Res. 9:3611–3622.
2017.PubMed/NCBI
|
|
33
|
Wang C, Cai L, Liu J, Wang G, Li H, Wang
X, Xu W, Ren M, Feng L, Liu P and Zhang C: MicroRNA-30a-5p inhibits
the growth of renal cell carcinoma by modulating GRP78 expression.
Cell Physiol Biochem. 43:2405–2419. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jones KB, Salah Z, Del Mare S, Galasso M,
Gaudio E, Nuovo GJ, Lovat F, LeBlanc K, Palatini J, Randall RL, et
al: miRNA signatures associate with pathogenesis and progression of
osteosarcoma. Cancer Res. 72:1865–1877. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shen A, Zhang Y, Yang H, Xu R and Huang G:
Overexpression of ZEB1 relates to metastasis and invasion in
osteosarcoma. J Surg Oncol. 105:830–834. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Larsen JE, Nathan V, Osborne JK, Farrow
RK, Deb D, Sullivan JP, Dospoy PD, Augustyn A, Hight SK, Sato M, et
al: ZEB1 drives epithelial-to-mesenchymal transition in lung
cancer. J Clin Invest. 126:3219–3235. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jia B, Liu H, Kong Q and Li B:
Overexpression of ZEB1 associated with metastasis and invasion in
patients with gastric carcinoma. Mol Cell Biochem. 366:223–229.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang GJ, Zhou T, Tian HP, Liu ZL and Xia
SS: High expression of ZEB1 correlates with liver metastasis and
poor prognosis in colorectal cancer. Oncol Lett. 5:564–568. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Singh M, Spoelstra NS, Jean A, Howe E,
Torkko KC, Clark HR, Darling DS, Shroyer KR, Horwitz KB, Broaddus
RR and Richer JK: ZEB1 expression in type I vs. type II endometrial
cancers: A marker of aggressive disease. Mod Pathol. 21:912–923.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Harb OA, Elfeky MA, El Shafaay BS, Taha
HF, Osman G, Harera IS, Gertallah LM, Abdelmonem DM and Embaby A:
SPOP, ZEB-1 and E-cadherin expression in clear cell renal cell
carcinoma (cc-RCC): Clinicopathological and prognostic
significance. Pathophysiology. 25:335–345. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang S, Hong Z, Chai Y, Liu Z, Du Y, Li Q
and Liu Q: CSN5 promotes renal cell carcinoma metastasis and EMT by
inhibiting ZEB1 degradation. Biochem Biophys Res Commun.
488:101–108. 2017. View Article : Google Scholar : PubMed/NCBI
|