Introduction
Neural crest cells originate from the dorsal margin
of the neural plate, and can differentiate into various types of
cells and tissues (1,2). Numerous neural crest cells have been
isolated and characterized from different organs and tissues
(3–7). During embryonic development, neural
crest cells arise from the trunk region of the neural crest,
migrate ventrally and aggregate adjacently to the dorsal aorta to
form the primary sympathetic chain (8). The determination of neural crest cell
fate is regulated by environmental factors from the extracellular
surroundings (9). In addition,
neural crest cells differentiate into various cell lineages
according to their position in the embryo, inducing the formation
of different cell types, including neurons, melanocytes, glial
cells of the peripheral nervous system, endoneurial fibroblasts and
endocrine cells (10–12)
Environmental factors may determine the
differentiation fate of neural crest cells in vitro; neural
crest cells were reported to be induced by a combination of
secreted signals (11,12). Bone morphogenetic proteins (BMPs) are
a unique group of proteins encoded by the transforming growth
factor-β superfamily of genes, and have been reported as key
regulators of embryogenesis (13).
In addition, BMPs were observed to regulate the establishment of
the embryonic body plan, dorsal-ventral patterning and the
differentiation of neural cells (14–17).
Additionally, BMP signaling has been demonstrated to affect the
development of dorsal neural tube cells and formation of neural
crest cells during a critical period prior to neural tube closure
(18). Neuregulins (NRGs) are
members of the epidermal growth factor protein family; it has been
reported that NRGs are primarily expressed and secreted by neurons,
and act on the surrounding glial cells (19). NRGs were demonstrated to induce the
growth and differentiation of glial, epithelial and muscle cells
in vitro (20–22). It has been reported that
NRG−/− embryos died during embryogenesis and displayed
heart malformations (23). NRGs may
affect the survival, proliferation, migration, differentiation and
myelination potential of Schwann cells (24–29);
developing Schwann cells originate from neural crest cells that
migrated along developing nerve fibers (10,30–32).
Collectively, these findings suggest that environmental factors
serve a critical role in neural crest cell differentiation. The
present study aimed to determine the mechanism underlying neural
crest cell differentiation in response to treatment with BMP4 and
NRGs.
Myc activity has been reported to be a critical
factor for the development and maintenance of stem cell properties;
Myc has been demonstrated to control stem cell functions, including
proliferation, differentiation and survival (33). Neural crest cells are generated from
neural crest stem cells; as a migratory and multipotent cell
population, neural crest cells can give rise to a variety of cell
lineages during vertebrate development (34). N-Myc (MYCN) expression was observed
in ~25% of neuroblastoma cases (35). A neuroblastoma is a tumor of the
peripheral sympathetic nervous system and MYCN overexpression has
been proposed as a tumorigenic event in the development of this
disease (36,37). Furthermore, MYCN expression may be
associated with the self-renewal ability and tumorigenic potential
of neuroblastoma cells (36,38). Therefore, another aim of the present
study was to determine whether MYCN could regulate the self-renewal
ability of neural crest cells, and how the interaction between BMP4
or NGR and MYCN affects the fate of neural crest
differentiation.
Materials and methods
Experimental animals
In the present study, 3 male and 9 female C57BL/6J
mice (weight, ~22 g; age, ~9 weeks) were employed. All mice were
housed under specific pathogen-free conditions as previous
described (39). The animal
experiments were approved by the Institutional Animal Care and Use
Committee of Southwest University.
Cell culture and in vitro
differentiation assays
Pregnant female mice (8.5–9 days gestation) were
sacrificed via exposure to CO2. The embryos were removed
and washed in PBS. A total of 10–12 neural tube sections were
excised with a scalpel and planted in 6-well cell culture plates
containing Dulbecco's Modified Eagle Medium/Nutrient Mixture F-12
medium (DMEM/F12; Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) medium as previously described (32), and photographed at 2, 24 and 48 h
with a Nikon TS100 inverted microscope (Nikon Corporation, Tokyo,
Japan) at a magnification of ×40 or ×100. Image-Pro Plus 6.0
software (Media Cybernetics, Inc., Rockville, MD, USA) was used for
analysis. All experiments were conducted using neural crest cells
and their descendants that had not been cultured for >12
passages. For agent-induced differentiation assays, neural crest
cells were cultured with 50 ng/ml BMP4 or 130 ng/ml NRG (both
R&D Systems, Inc., Minneapolis, MN, USA) for 10 days in 37°C.
Neural crest cells treated with 1 µl/ml DMSO (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) served as the negative control.
Immunofluorescence
The tenth passage neural crest cells treated with
BMP4, NRG or DMSO were fixed in 4% paraformaldehyde at room
temperature for 15 min, permeated with PBS with Tween-20 (0.3%
Triton X-100) at room temperature for 5 min and blocked with 10%
goat serum (Beyotime Institute of Biotechnology, Haimen, China) at
room temperature for 1 h. The cells were then incubated with
primary antibodies at 4°C overnight. The primary antibodies were as
follows: Rabbit anti-glial fibrillary acidic protein (GFAP; cat.
no. ab7260; 1:200; Sigma-Aldrich; Merck KGaA), chicken anti-Nestin
(1:1,000; cat. no. NB100-1604; Novus Biologicals, LLC, Littleton,
CO, USA), rabbit anti-SRY-related HMG-box 10 (Sox10; 1:300; cat.
no. ab155279; Abcam, Cambridge UK) and mouse anti-neuronal-specific
class III β-tubulin (TuJ1; 1:300; cat. no. ab78078; Abcam).
Following washing with PBS, cells were incubated with secondary
antibodies at room temperature for 2 h. All secondary antibodies
were purchased from Invitrogen (Thermo Fisher Scientific, Inc.) and
used at 1:1,000 dilution. The secondary antibodies were as follows:
Alexa Fluor® 488-conjugated goat anti-mouse (cat. no.
A-11001), anti-rabbit (cat. no. A-11008) and anti-chicken (cat. no.
A-11039), and Alexa Fluor 594-conjugated goat anti-rabbit (cat. no.
A-11012) immunoglobulin G. Then, all cells were washed with PBS and
counterstained with DAPI (Beyotime Institute of Biotechnology) at
room temperature for 20 min to detect nuclei, and images were
captured with a Nikon Eclipse TE2000-E fluorescence microscope
(Nikon Corporation) at a magnification of ×100 or ×200. Image-Pro
Plus 6.0 software was used for analysis.
Retroviral production and
transfection
The pBabe-puro/MYCN plasmid (Youbio, Hunan, China)
was used to overexpress mouse MYCN in neural crest cells
(MYCN-overexpressing neural crest cells), as previously reported
(40), and the empty pBabe-puro
plasmid as the control. Retroviral production and transfection were
conducted as described previously (41). One day after retroviral transfection,
the cells were cultured at 37°C in the presence of 2 µg/ml
puromycin for 3 days for resistance-based selection.
Soft agar clonogenic and sphere
formation assays
For soft agar colony assay, a total of 1,500
pBabe-puro/MYCN or empty pBabe-puro neural crest cells in
suspension were mixed with 0.3% low melting point agar containing
DMEM supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.). Cells were planted onto 6-well plates (1,500
cells/well) with a solidified bottom layer (0.6% low melting point
agar in the same growth medium) for 14 days at 37°C. For sphere
formation assay, the medium in the aforementioned wells was
replaced with DMEM supplemented with 20 ng/ml epidermal growth
factor and 10 ng/ml basic fibroblast growth factor (both Gibco;
Thermo Fisher Scientific, Inc.). After the 14-day culture, the
colonies or the spheres were examined and photographed using a
Nikon TS100 inverted microscope at a magnification of ×100.
Western blot analysis
MYCN-overexpressing neural crest cells treated with
BMP4, NRG or DMSO were suspended in RIPA lysis buffer and the total
protein concentration determined using the Enhanced BCA protein
assay kit (both Beyotime Institute of Biotechnology). Following
this, 50 µg/lane of protein were separated by SDS-PAGE on 10% gel,
transferred to polyvinylidene difluoride membranes. Membranes were
then blocked with 5% no fat milk at room temperature for 1 h and
then incubated with anti-MYCN (cat. no. ab24193; 1:1,000; Abcam) or
anti-α-tubulin (cat. no. SAB4500087; 1:1,000; Sigma-Aldrich; Merck
KGaA). Horseradish peroxidase-conjugated goat anti-mouse (cat. no.
5220–0341) or anti-rabbit (cat. no. 5220-0336) secondary antibodies
(both 1:20,000; Kirkegaard & Perry Laboratories; SeraCare Life
Sciences, Inc., Milford, MA, USA) were used as secondary
antibodies. Proteins were visualized with BeyoECL Plus (Beyotime
Institute of Biotechnology).
Results
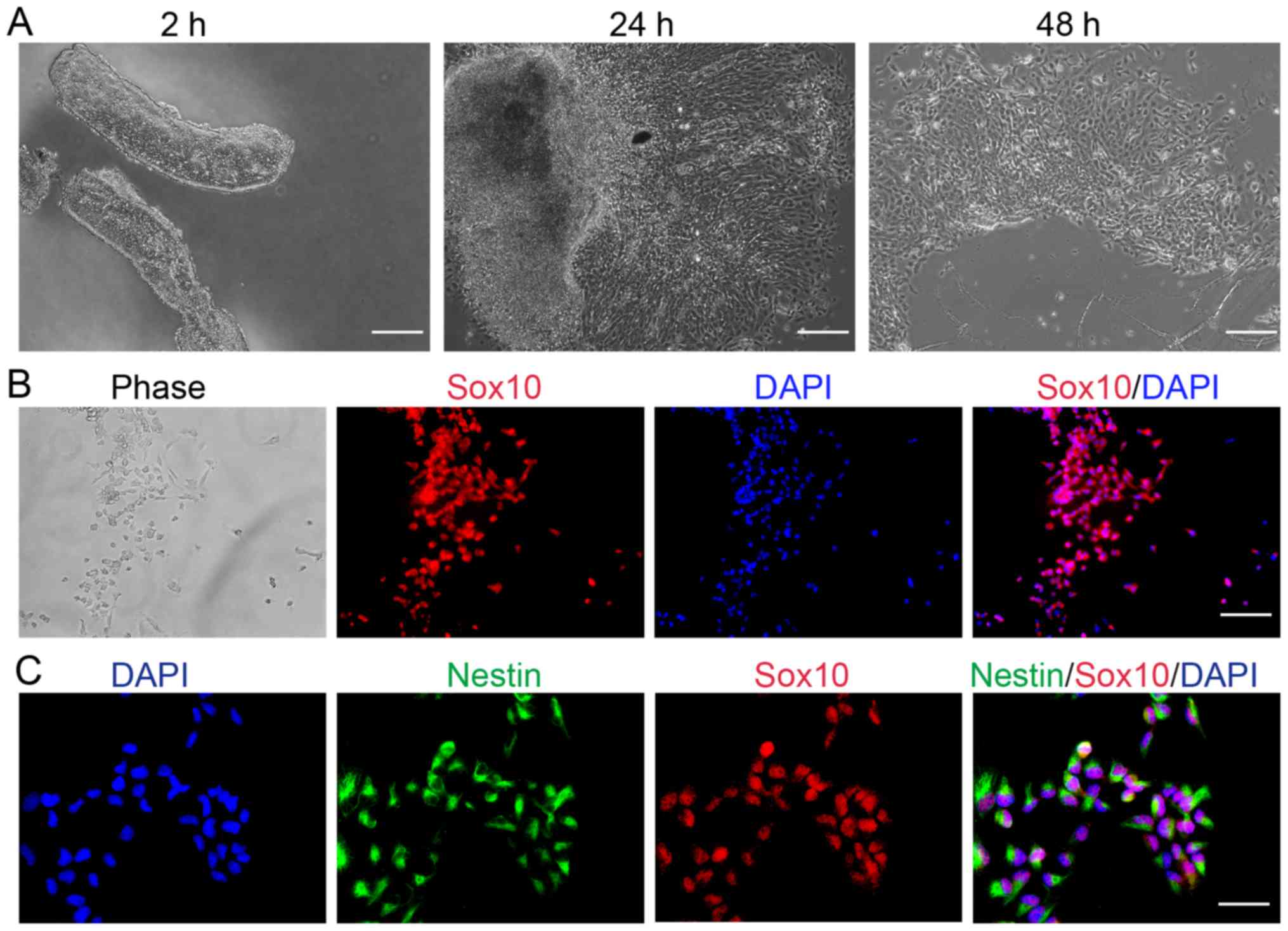
Cells migrating from the neural tube
are neural crest cells, which are characterized by Sox10 and Nestin
expression
Explant culture has been successfully used to
identify the properties of neural crest cells originating from the
stem cells of developing rodents (42,43).
Therefore, explant culture was selected in the present study to
determine whether any cells migrated from the neural tube of mice
embryos. After a 2-h explant culture, the neural tube was clearly
observed; after 24 h, neural crest cells were detected at the edges
of the tissue blocks and some cells had migrated from the neural
tube explants into the culture medium (Fig. 1A). The number of cells in the culture
increased in what appeared to be a time-dependent manner,
suggesting that the cells continued to migrate from the neural
tube; after 2 days, more cells floated freely in the medium.
Sox10 is a unique HMG-box transcription factor
expressed throughout the neural crest and in oligodendrocyte
progenitor cells of the central nervous system (9,44,45). In
the present study, cells were characterized via immunofluorescence
to determine whether cells migrating from the neural tube expressed
neural stem cell-associated markers. Compared with the phase image,
immunofluorescent analysis revealed that all cells expressed Sox10
(Fig. 1B). Therefore, suggesting
that cells migrating from the mouse embryo neural tube, which
express Sox10, may be characterized as neural crest cells.
Nestin has been reported as a marker of neural stem
or progenitor cells (46,47). Embryonic stem cell-derived neural
precursor cells that had been further induced to differentiate into
neurons may be selected based on the aforementioned strategy
(48). The results of the present
study demonstrated that neural crest cells were positive for Nestin
(Fig. 1C), suggesting that neural
crest cells may possess neural stem cell characteristics.
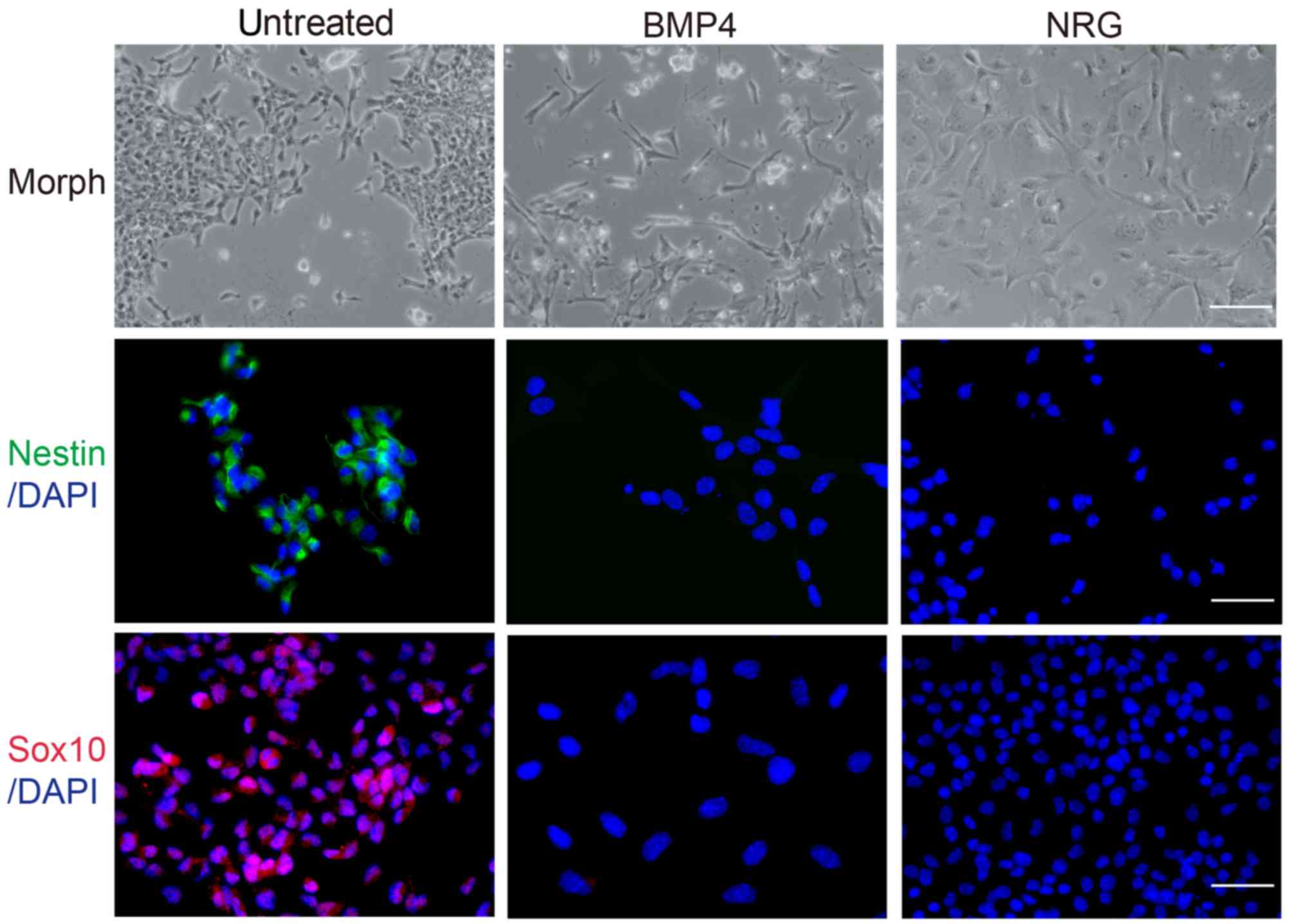
Neural crest cells maintain the
potential of multilineage differentiation
The present study investigated the differentiation
potential of neural crest cells in response to a variety of agents.
A cell suspension was prepared from individual secondary colonies
and plated onto glass coverslips. The results of the
immunofluorescence analysis demonstrated that cells expressed the
stem cell markers, Nestin and Sox10. In addition, neural crest
cells were treated with BMP4 or NRG for 10 days. The majority of
DMSO-treated cells exhibited a round and prominent nucleus, and
abundant cytoplasm (Fig. 2). Cells
treated with BMP4 exhibited neuronal cell morphology (49), with numerous long neuritic processes
and small cell bodies that frequently formed aggregates (Fig. 2); however, cells treated with NRG
exhibited Schwann-like cell morphology (50,51),
with ovoid cell bodies, a prominent nucleus and natural bipolar
extensions (Fig. 2). Therefore,
these data suggested that BMP4 and NRG treatment may induce neural
crest cell differentiation into neurons and Schwann cells.
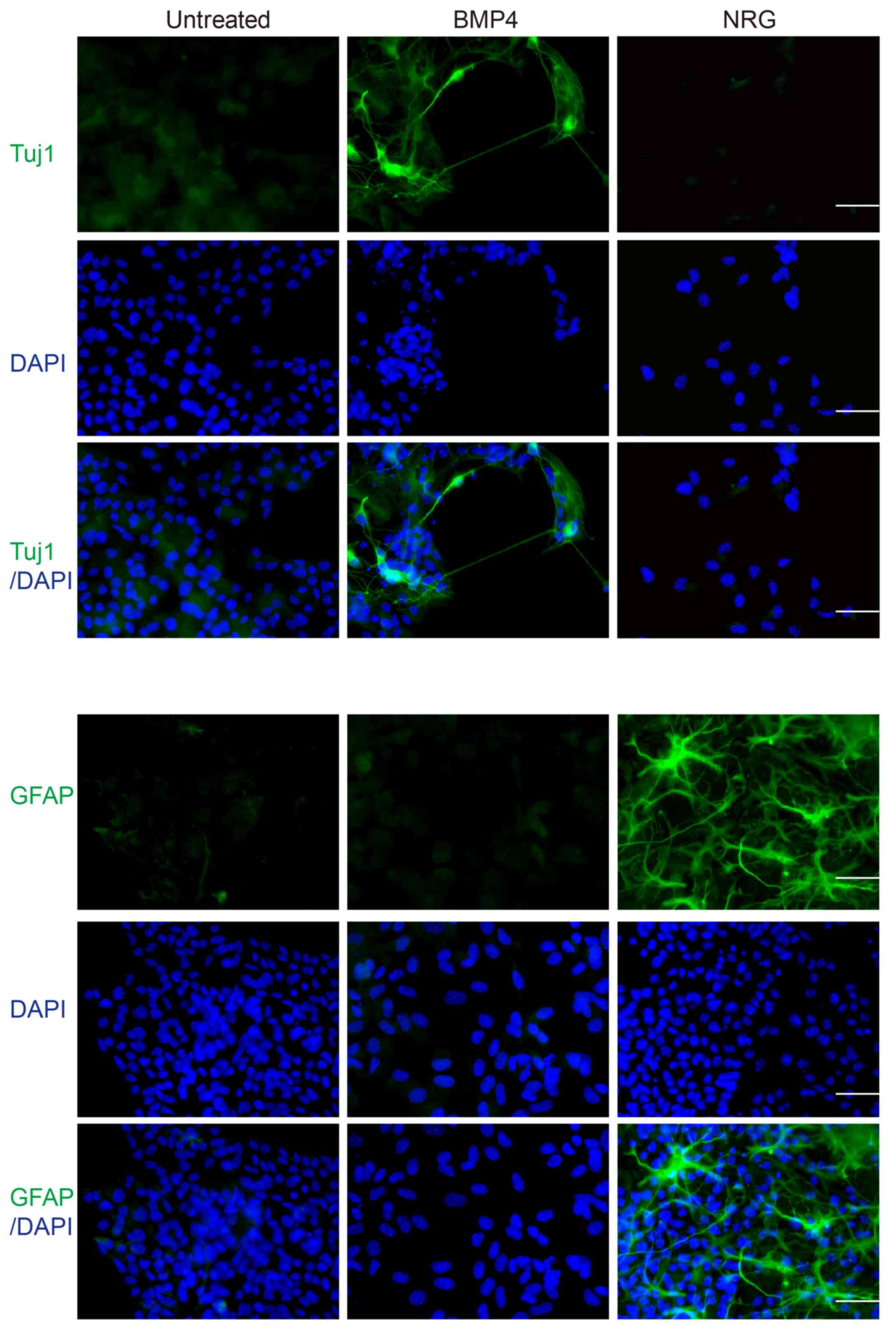
To confirm the cell phenotype following treatment
with BMP4 or NRG, immunofluorescence was conducted using
anti-Nestin and anti-Sox10 antibodies. Compared with the untreated
cell group, treatment with BMP4 or NRG resulted in a marked
reduction in the number of cells expressing stem cell markers
(Fig. 2), accompanied by a marked
increase in the number of cells expressing differentiation markers
(Fig. 3). BMP4-treated cells
expressed the neuronal marker, Tuj1 (52), whereas NRG-treated cells expressed
GFAP, which is a common marker of Schwann and glial cells (53), upon induction of differentiation. On
the contrary, untreated cells expressed differentiation markers at
markedly lower levels (Fig. 3).
These differentiation analyses suggested that neural crest cells
may possess the potential for multilineage differentiation and that
environmental factors may control their fate.
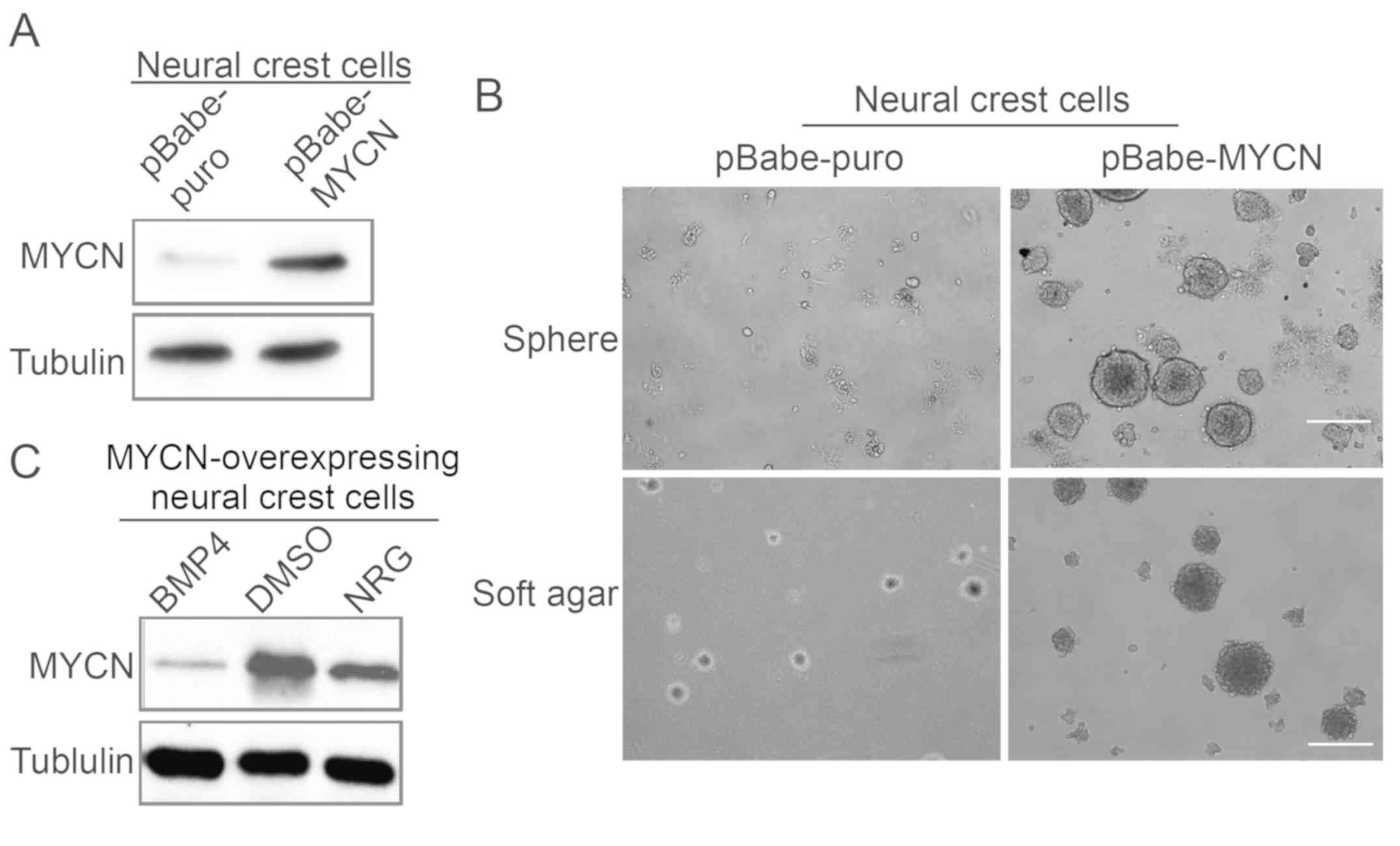
Clonal sphere formation assay of
MYCN-overexpressing neural crest cells suggests clonogenic
self-renewal potential in vitro
MYCN upregulation was validated by western blotting
(Fig. 4A). To investigate the
function of MYCN in neural crest cells, the self-renewal and
clonogenic abilities of MYCN-overexpressing neural crest cells were
analyzed via soft agar and sphere formation assays. The result
demonstrated that MYCN-overexpressing neural crest cells exhibited
a higher number of colonies and larger colony sizes compared with
the control group of neural crest cells (Fig. 4B). Similarly, the neurosphere
formation ability of MYCN-overexpressing neural crest cells
appeared to be enhanced compared with the neural crest cell control
group under the same culture conditions. These results demonstrated
that MYCN may promote the self-renewal ability of neural crest
cells. The authors further investigated the effects of BMP4 and NRG
on MYCN expression in MYCN-overexpressing neural crest cells; the
protein expression levels of MYCN were markedly decreased in cells
treated with BMP4 or NRG compared with the control cells (Fig. 4C).
Discussion
Neural crest cells have stem cell characteristics
and have the ability to generate various types of cells and tissues
during vertebrate development (54).
In the present study, neural crest cells were isolated from mouse
embryos and characterized by specific stem cell markers, including
Sox10 and Nestin. Factors of the extracellular environment may
affect the direction of neural crest cell differentiation. It has
been reported that partitioning defective 3 homolog regulates the
contact between neural crest cells and the timing of Schwann cell
differentiation (55). In human
neural crest stem cells, aligned electrospun fibers were revealed
to promote differentiation towards the Schwann cell lineage
(56). A recent study suggested
that, during the neural differentiation of embryonic stem cells,
miR-29b promoted the differentiation of embryonic stem cells into
neural tube epithelial cells and inhibited their differentiation
into neural crest cells (57).
It was previously reported that embryonic stem cells
cultured with BMP4 differentiated into germ cells (58,59).
BMP4 also regulated the proliferation and differentiation in
epithelial and mesenchymal tissue compartments of the developing
mouse ureter (60). In addition,
BMP4 was reported to serve a key role in the differentiation of
auditory neuron-like cells from bone-derived mesenchymal stromal
cells (61). NRG is a type of
polypeptide growth factor that serves a key role in the development
and differentiation of the heart and nervous system (23,62).
Generally, neural crest cells differentiate into glia, neurons and
melanocytes in the mouse embryo (9,63). In
the present study, the induction of differentiation via specific
agents, BMP4 or NRG, revealed that neural crest cells may
differentiate into neurons or Schwann cells, respectively,
indicating that neural crest cells may alter their direction of
differentiation according to their environments. Therefore, BMP4
and NRG could be considered as key factors in neural crest cell
differentiation.
Self-renewal ability is an essential characteristic
of stem cells, and enables the generation of daughter cells with
the same developmental potential as their parental cells (64,65).
Colony and sphere formation assays have been widely used to
evaluate the self-renewal ability of individual stem cells
(66–68). MYCN was reported as a key factor in
the maintenance of embryonic stem cell-derived neural crest stem
cells (69). The soft agar
clonogenic and sphere formation assays revealed that
MYCN-overexpressing neural crest cells were able to self-renew and
generate progeny cells with the same self-renewal ability,
MYCN-overexpressing neural crest cells developed more colonies
compared with the neural crest cells transfected with empty
vectors. Neuroblastoma is a cancer of neural crest stem cell
lineage, many reports demonstrated that MYCN acted as an oncogene
in neuroblastoma (35,36,70).
MYCN served important roles in balancing proliferation,
differentiation and cell death in neuroblastoma and normal neural
crest cells (71,72). Therefore, it was hypothesized that
MYCN-overexpressing neural crest cells acquire tumorigenic
potential and that MYCN may regulate the development of
neuroblastoma that originate from neural crest cells.
Of note, BMP4 could reduce MYCN expression and
promote differentiation in neuroblastoma cells (73), and NRG was involved in neuroblastoma
cell differentiation (74). These
reports suggested BMP4 and NRG regulated neuroblastoma development.
In the present study, BMP4 and NRG were shown to suppress MYCN
expression in MYCN-overexpressing neural crest cells, which
indicated that BMP4 and NRG may inhibit the maintenance of
MYCN-induced stemness, suggesting there was cross-talk between MYCN
and the BMP4 or NRG signaling pathway. Collectively, the findings
of the current study indicated a molecular mechanism through which
MYCN may promote the stemness of neural crest cells. Specific
agents, including BMP4 and NRG, may decrease MYCN expression and
induce neural crest cell differentiation. Therefore, the present
study revealed that the direction of cell differentiation would be
altered through modifying environmental factors. MYCN could serve a
key role in regulating neural crest cell differentiation, and BMP4
and NRG may be regarded as novel inhibitors of MYCN amplification
in neuroblastoma.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 31501100), the
Chongqing Science and Technology Commission (grant no.
cstc2016shmszx80101), the Ph.D. Start-up Foundation of Southwest
University (grant no. SWU 2015021) and the Chongqing Research
Program of Basic Research and Frontier Technology (grant no.
cstc2015jcyjA10120), the China Postdoctoral Science Foundation
(grant no. 2016M592624) and the Chongqing Postdoctoral Science
Special Foundation (grant no. Xm2016087). The funders had no role
in the study design, data collection and analysis, decision to
publish or preparation of the manuscript.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SZ performed the all the experiment with the
exception of retroviral production and transfection, and wrote the
manuscript. WL performed the statistical analyses. HFD and HC
conducted the experiments. LY designed the current study and
revised this manuscript. All the authors have read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Animal Care and Use Committee of Southwest University (Chongqing,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gonzalez Malagon SG, Lopez Munoz AM, Doro
D, Bolger TG, Poon E, Tucker ER, Adel Al-Lami H, Krause M, Phiel
CJ, Chesler L and Liu KJ: Glycogen synthase kinase 3 controls
migration of the neural crest lineage in mouse and Xenopus. Nat
Commun. 9:11262018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang T, Moore M and He F: Pten regulates
neural crest proliferation and differentiation during mouse
craniofacial development. Dev Dyn. 247:304–314. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang J, Duan X, Zhang H, Deng Z, Zhou Z,
Wen N, Smith AJ, Zhao W and Jin Y: Isolation of neural
crest-derived stem cells from rat embryonic mandibular processes.
Biol Cell. 98:567–575. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yoshida S, Shimmura S, Nagoshi N, Fukuda
K, Matsuzaki Y, Okano H and Tsubota K: Isolation of multipotent
neural crest-derived stem cells from the adult mouse cornea. Stem
Cells. 24:2714–2722. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Morrison SJ, White PM, Zock C and Anderson
DJ: Prospective identification, isolation by flow cytometry, and in
vivo self-renewal of multipotent mammalian neural crest stem cells.
Cell. 96:737–749. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jiang X, Gwye Y, McKeown SJ,
Bronner-Fraser M, Lutzko C and Lawlor ER: Isolation and
characterization of neural crest stem cells derived from in
vitro-differentiated human embryonic stem cells. Stem Cells Dev.
18:1059–1070. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chung KF, Sicard F, Vukicevic V, Hermann
A, Storch A, Huttner WB, Bornstein SR and Ehrhart-Bornstein M:
Isolation of neural crest derived chromaffin progenitors from adult
adrenal medulla. Stem Cells. 27:2602–2613. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lumb R and Schwarz Q: Sympathoadrenal
neural crest cells: The known, unknown and forgotten? Dev Growth
Differ. 57:146–157. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sauka-Spengler T and Bronner-Fraser M: A
gene regulatory network orchestrates neural crest formation. Nat
Rev Mol Cell Biol. 9:557–568. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Joseph NM, Mukouyama YS, Mosher JT, Jaegle
M, Crone SA, Dormand EL, Lee KF, Meijer D, Anderson DJ and Morrison
SJ: Neural crest stem cells undergo multilineage differentiation in
developing peripheral nerves to generate endoneurial fibroblasts in
addition to Schwann cells. Development. 131:5599–5612. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wakamatsu Y, Maynard TM and Weston JA:
Fate determination of neural crest cells by NOTCH-mediated lateral
inhibition and asymmetrical cell division during gangliogenesis.
Development. 127:2811–2821. 2000.PubMed/NCBI
|
|
12
|
Steventon B, Araya C, Linker C, Kuriyama S
and Mayor R: Differential requirements of BMP and Wnt signalling
during gastrulation and neurulation define two steps in neural
crest induction. Development. 136:771–779. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sykaras N and Opperman LA: Bone
morphogenetic proteins (BMPs): How do they function and what can
they offer the clinician. J Oral Sci. 45:57–73. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sykaras N and Opperman LA: Bone
morphogenetic proteins (BMPs): How do they function and what can
they offer the clinician? J Oral Sci. 45:57–73. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ripamonti U and Reddi AH: Tissue
engineering, morphogenesis, and regeneration of the periodontal
tissues by bone morphogenetic proteins. Crit Rev Oral Biol Med.
8:154–163. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Paralkar VM, Weeks BS, Yu YM, Kleinman HK
and Reddi AH: Recombinant human bone morphogenetic protein 2B
stimulates PC12 cell differentiation: Potentiation and binding to
type IV collagen. J Cell Biol. 119:1721–1728. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shah NM, Groves AK and Anderson DJ:
Alternative neural crest cell fates are instructively promoted by
TGFbeta superfamily members. Cell. 85:331–343. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stottmann RW and Klingensmith J: Bone
morphogenetic protein signaling is required in the dorsal neural
folds before neurulation for the induction of spinal neural crest
cells and dorsal neurons. Dev Dyn. 240:755–765. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Raabe TD, Francis A and DeVries GH:
Neuregulins in glial cells. Neurochem Res. 23:311–318. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wen D, Peles E, Cupples R, Suggs SV, Bacus
SS, Luo Y, Trail G, Hu S, Silbiger SM, Levy RB, et al: Neu
differentiation factor: A transmembrane glycoprotein containing an
EGF domain and an immunoglobulin homology unit. Cell. 69:559–572.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Marchionni MA, Goodearl AD, Chen MS,
Bermingham-McDonogh O, Kirk C, Hendricks M, Danehy F, Misumi D,
Sudhalter J, Kobayashi K, et al: Glial growth factors are
alternatively spliced erbB2 ligands expressed in the nervous
system. Nature. 362:312–318. 1993. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Carraway KL III and Cantley LC: A neu
acquaintance for erbB3 and erbB4: A role for receptor
heterodimerization in growth signaling. Cell. 78:5–8. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Meyer D and Birchmeier C: Multiple
essential functions of neuregulin in development. Nature.
378:386–390. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Garratt AN, Britsch S and Birchmeier C:
Neuregulin, a factor with many functions in the life of a schwann
cell. Bioessays. 22:987–996. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jessen KR and Mirsky R: Origin and early
development of Schwann cells. Microsc Res Tech. 41:393–402. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jessen KR and Mirsky R: Schwann cells and
their precursors emerge as major regulators of nerve development.
Trends Neurosci. 22:402–410. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Freeman MR: Sculpting the nervous system:
Glial control of neuronal development. Curr Opin Neurobiol.
16:119–125. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mirsky R, Jessen KR, Brennan A, Parkinson
D, Dong Z, Meier C, Parmantier E and Lawson D: Schwann cells as
regulators of nerve development. J Physiol Paris. 96:17–24. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Falls DL: Neuregulins: Functions, forms,
and signaling strategies. Exp Cell Res. 284:14–30. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li L, Habbes HW, Eiberger J, Willecke K,
Dermietzel R and Meier C: Analysis of connexin expression during
mouse Schwann cell development: Connexin29 as a novel marker for
the transition of neural crest to precursor cells. Glia. 55:93–103.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Aquino JB, Hjerling-Leffler J, Koltzenburg
M, Edlund T, Villar MJ and Ernfors P: In vitro and in vivo
differentiation of boundary cap neural crest stem cells into mature
Schwann cells. Exp Neurol. 198:438–449. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Buchstaller J, Sommer L, Bodmer M,
Hoffmann R, Suter U and Mantei N: Efficient isolation and gene
expression profiling of small numbers of neural crest stem cells
and developing Schwann cells. J Neurosci. 24:2357–2365. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Laurenti E, Varnum-Finney B, Wilson A,
Ferrero I, Blanco-Bose WE, Ehninger A, Knoepfler PS, Cheng PF,
MacDonald HR, Eisenman RN, et al: Hematopoietic stem cell function
and survival depend on c-Myc and N-Myc activity. Cell Stem Cell.
3:611–624. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Crane JF and Trainor PA: Neural crest stem
and progenitor cells. Annu Rev Cell Dev Biol. 22:267–286. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang M and Weiss WA: Neuroblastoma and
MYCN. Cold Spring Harb Perspect Med. 3:a0144152013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Alam G, Cui H, Shi H, Yang L, Ding J, Mao
L, Maltese WA and Ding HF: MYCN promotes the expansion of
Phox2B-positive neuronal progenitors to drive neuroblastoma
development. Am J Pathol. 175:856–866. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mao L, Ding J, Perdue A, Yang L, Zha Y,
Ren M, Huang S, Cui H and Ding HF: Cyclin E1 is a common target of
BMI1 and MYCN and a prognostic marker for neuroblastoma
progression. Oncogene. 31:3785–3795. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Brodeur GM: Neuroblastoma: Biological
insights into a clinical enigma. Nat Rev Cancer. 3:203–216. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Martinewski A, Correia CS, de Souza NL and
Merusse JL: Mouse housing system using pressurized cages
intraventilated by direct-current microfans. J Am Assoc Lab Anim
Sci. 51:177–180. 2012.PubMed/NCBI
|
|
40
|
Itahana K, Zou Y, Itahana Y, Martinez JL,
Beausejour C, Jacobs JJ, Van Lohuizen M, Band V, Campisi J and
Dimri GP: Control of the replicative life span of human fibroblasts
by p16 and the polycomb protein Bmi-1. Mol Cell Biol. 23:389–401.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cui H, Schroering A and Ding HF: p53
mediates DNA damaging drug-induced apoptosis through a
caspase-9-dependent pathway in SH-SY5Y neuroblastoma cells. Mol
Cancer Ther. 1:679–686. 2002.PubMed/NCBI
|
|
42
|
Lo L, Dormand E, Greenwood A and Anderson
DJ: Comparison of the generic neuronal differentiation and neuron
subtype specification functions of mammalian achaete-scute and
atonal homologs in cultured neural progenitor cells. Development.
129:1553–1567. 2002.PubMed/NCBI
|
|
43
|
Greenwood AL, Turner EE and Anderson DJ:
Identification of dividing, determined sensory neuron precursors in
the mammalian neural crest. Development. 126:3545–3559.
1999.PubMed/NCBI
|
|
44
|
Kelsh RN: Sorting out Sox10 functions in
neural crest development. Bioessays. 28:788–798. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kawaguchi J, Nichols J, Gierl MS, Faial T
and Smith A: Isolation and propagation of enteric neural crest
progenitor cells from mouse embryonic stem cells and embryos.
Development. 137:693–704. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Selander L and Edlund H: Nestin is
expressed in mesenchymal and not epithelial cells of the developing
mouse pancreas. Mech Dev. 113:189–212. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lendahl U, Zimmerman LB and McKay RD: CNS
stem cells express a new class of intermediate filament protein.
Cell. 60:585–595. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lee H, Mok KH, Muhandiram R, Park KH, Suk
JE, Kim DH, Chang J, Sung YC, Choi KY and Han KH: Local structural
elements in the mostly unstructured transcriptional activation
domain of human p53. J Biol Chem. 275:29426–29432. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Trainor PA: Specification of neural crest
cell formation and migration in mouse embryos. Semin Cell Dev Biol.
16:683–693. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sugimoto T, Kato T, Sawada T, Horii Y,
Kemshead JT, Hino T, Morioka H and Hosoi H: Schwannian cell
differentiation of human neuroblastoma cell lines in vitro induced
by bromodeoxyuridine. Cancer Res. 48:2531–2537. 1988.PubMed/NCBI
|
|
51
|
Ross RA, Spengler BA, Domenech C, Porubcin
M, Rettig WJ and Biedler JL: Human neuroblastoma I-type cells are
malignant neural crest stem cells. Cell Growth Differ. 6:449–456.
1995.PubMed/NCBI
|
|
52
|
Lee S, Lee B, Lee JW and Lee SK: Retinoid
signaling and neurogenin2 function are coupled for the
specification of spinal motor neurons through a chromatin modifier
CBP. Neuron. 62:641–654. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jessen KR and Mirsky R: The origin and
development of glial cells in peripheral nerves. Nat Rev Neurosci.
6:671–682. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Bhatt S, Diaz R and Trainor PA: Signals
and switches in Mammalian neural crest cell differentiation. Cold
Spring Harb Perspect Biol. 5(pii): a0083262013.PubMed/NCBI
|
|
55
|
Blasky AJ, Pan L, Moens CB and Appel B:
Pard3 regulates contact between neural crest cells and the timing
of Schwann cell differentiation but is not essential for neural
crest migration or myelination. Dev Dyn. 243:1511–1523. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ren YJ, Zhang S, Mi R, Liu Q, Zeng X, Rao
M, Hoke A and Mao HQ: Enhanced differentiation of human neural
crest stem cells towards the Schwann cell lineage by aligned
electrospun fiber matrix. Acta Biomater. 9:7727–7736. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Xi J, Wu Y, Li G, Ma L, Feng K, Guo X, Jia
W, Wang G, Yang G, Li P and Kang J: Mir-29b mediates the neural
tube versus neural crest fate decision during embryonic stem cell
neural differentiation. Stem Cell Reports. 9:571–586. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Makoolati Z, Movahedin M and
Forouzandeh-Moghadam M: In vitro germ cell differentiation from
embryonic stem cells of mice: Induction control by BMP4 signaling.
Biosci Rep. 36:e004072016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yang S, Yuan Q, Niu M, Hou J, Zhu Z, Sun
M, Li Z and He Z: BMP4 promotes mouse iPS cell differentiation to
male germ cells via Smad1/5, Gata4, Id1 and Id2. Reproduction.
153:211–220. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Mamo TM, Wittern AB, Kleppa MJ, Bohnenpoll
T, Weiss AC and Kispert A: BMP4 uses several different effector
pathways to regulate proliferation and differentiation in the
epithelial and mesenchymal tissue compartments of the developing
mouse ureter. Hum Mol Genet. 26:3553–3563. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Peng T, Zhu G, Dong Y, Zeng J, Li W, Guo
W, Chen Y, Duan M, Hocher B and Xie D: BMP4: A possible key factor
in differentiation of auditory neuron-like cells from bone-derived
mesenchymal stromal cells. Clin Lab. 61:1171–1178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Britsch S, Li L, Kirchhoff S, Theuring F,
Brinkmann V, Birchmeier C and Riethmacher D: The ErbB2 and ErbB3
receptors and their ligand, neuregulin-1, are essential for
development of the sympathetic nervous system. Genes Dev.
12:1825–1836. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Milet C and Monsoro-Burq AH: Neural crest
induction at the neural plate border in vertebrates. Dev Biol.
366:22–33. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Borowicz S, Van Scoyk M, Avasarala S,
Karuppusamy Rathinam MK, Tauler J, Bikkavilli RK and Winn RA: The
soft agar colony formation assay. J Vis Exp. 92:e519982014.
|
|
65
|
Franken NA, Rodermond HM, Stap J, Haveman
J and van Bree C: Clonogenic assay of cells in vitro. Nat Protoc.
1:2315–2319. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hamburger AW and Salmon SE: Primary
bioassay of human tumor stem cells. Science. 197:461–463. 1977.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Cui H, Ma J, Ding J, Li T, Alam G and Ding
HF: Bmi-1 regulates the differentiation and clonogenic self-renewal
of I-type neuroblastoma cells in a concentration-dependent manner.
J Biol Chem. 281:34696–34704. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Reynolds BA and Weiss S: Generation of
neurons and astrocytes from isolated cells of the adult mammalian
central nervous system. Science. 255:1707–1710. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhang JT, Weng ZH, Tsang KS, Tsang LL,
Chan HC and Jiang XH: MycN is critical for the maintenance of human
embryonic stem cell-derived neural crest stem cells. PLoS One.
11:e01480622016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Ke XX, Zhang D, Zhao H, Hu R, Dong Z, Yang
R, Zhu S, Xia Q, Ding HF and Cui H: Phox2B correlates with MYCN and
is a prognostic marker for neuroblastoma development. Oncol Lett.
9:2507–2514. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Newman EA, Chukkapalli S, Bashllari D,
Thomas TT, Van Noord RA, Lawlor ER, Hoenerhoff MJ, Opipari AW and
Opipari VP: Alternative NHEJ pathway proteins as components of MYCN
oncogenic activity in human neural crest stem cell differentiation:
Implications for neuroblastoma initiation. Cell Death Dis.
8:32082017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Althoff K, Beckers A, Bell E, Nortmeyer M,
Thor T, Sprüssel A, Lindner S, De Preter K, Florin A, Heukamp LC,
et al: A Cre-conditional MYCN-driven neuroblastoma mouse model as
an improved tool for preclinical studies. Oncogene. 34:3357–3368.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Ferlemann FC, Menon V, Condurat AL, Rößler
J and Pruszak J: Surface marker profiling of SH-SY5Y cells enables
small molecule screens identifying BMP4 as a modulator of
neuroblastoma differentiation. Sci Rep. 7:136122017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Schlitter AM, Dorneburg C, Barth TF, Wahl
J, Schulte JH, Brüderlein S, Debatin KM and Beltinger C: CD57(high)
neuroblastoma cells have aggressive attributes ex situ and an
undifferentiated phenotype in Patients. PLoS One. 7:e420252012.
View Article : Google Scholar : PubMed/NCBI
|