Introduction
Sepsis is a pathophysiological syndrome caused by
infection and is a major public health problem (1). Its incidence rate has been reported to
be 535 cases per 100,000 person-years and rising; in-hospital
mortality remains high at 25–30% (2). Despite advances in care, sepsis is a
major cause of mortality and critical illness in intensive care
units worldwide (3). The heart is
the most vulnerable organ during sepsis, and ~50% of patients with
sepsis develop heart dysfunction (4,5).
Hydrogen sulfide (H2S) has been confirmed to serve an
important role in the physiological and pathological processes of
various systems, including the circulatory, nervous and respiratory
systems (6). H2S has a
protective role in cells as a regulator of vascular tone,
inflammatory responses and the clearance of reactive oxygen
species, and it may even reduce the risk of myocardial ischemia
(7,8). Multiple studies have confirmed that an
appropriate dose of H2S has cardiac protective effects
(7,9,10), which
may be associated with anti-apoptotic, anti-inflammatory and
anti-oxidative mechanisms (11). A
previous study by the present research team demonstrated that low
doses of H2S are able to improve the cardiac dysfunction
caused by sepsis (12); however, the
specific mechanisms involved are not clear.
Phosphoinositol-3-kinases (PI3Ks) are a conserved
family of signaling transducers involved in the regulation of cell
proliferation and survival. In studies of the mechanisms of cardiac
dysfunction caused by sepsis, the PI3K/protein kinase B (Akt)
signaling pathway has been proposed to have a key role in the
development of dysfunction (13,14).
Previous studies have revealed that PI3K and its downstream
mediator, Akt, serve a role in sepsis via the regulation of cell
activation, inflammation and apoptosis (15–17).
Additionally, studies have demonstrated that H2S is
involved in the PI3K/Akt signaling pathway in pancreatitis and
myocardial ischemia (18–20). However, it is unclear whether the
protective effect of H2S against myocardial damage
during sepsis is associated with the PI3K/Akt pathway. Therefore,
the current study used cecal ligation and puncture (CLP) to induce
a rat model of sepsis, and the effect and mechanism of
H2S on myocardial injury in sepsis were evaluated by the
administration of H2S and inhibitors of PI3K in this
model.
Materials and methods
Reagents
Sodium hydrosulfide (NaHS) was purchased from
Sigma-Aldrich (Merck KGaA; Darmstadt, Germany; cat. no. 161527).
LY294002 (a PI3K inhibitor) was purchased from ApexBio (Houston,
TX, USA; cat. no. 8250–200 mg). Akt, phospho (p)-Akt and caspase-3
antibodies were obtained from Abcam (Cambridge, MA, USA; cat. nos.
ab179463, ab192623 and ab13847, respectively). PI3K, p-PI3K and
nuclear factor-κB (NF-κB) antibodies were obtained from Cell
Signaling Technology, Inc. (Danvers, MA, USA; cat. nos. 4249, 4228
and 8242, respectively). B-cell lymphoma-2 (Bcl-2) and
Bcl-2-associated X protein (Bax) antibodies were obtained from
Boster Biological Technology, Ltd. (Wuhan, China; cat. nos. BA0412
and BA0315-2, respectively). Peroxidase-conjugated AffiniPure goat
anti-rabbit immunoglobulin G (IgG), peroxidase-conjugated
AffiniPure goat anti-mouse IgG and mouse anti-β-actin monoclonal
antibodies were obtained from OriGene Technologies, Inc. (Beijing,
China; cat. nos. ZB-2301, ZB-2305 and TA-09, respectively). In
situ Cell Death Detection kit, POD was obtained from Roche
Diagnostics GmbH (Mannheim, Germany; cat. no. 11684817910). Rat
troponin I (TnI), interleukin (IL)-10, IL-6 and tumor necrosis
factor-α (TNF-α) ELISA kits were provided by Elabscience
Biotechnology Co., Ltd. (Wuhan, China; cat. nos. E-EL-R0055c,
E-EL-R0016c, E-EL-R0015c and E-EL-R0019c, respectively).
Experimental animals
Adult male Sprague Dawley rats (7–8 weeks old; body
weight 211.58±11.42 g) were provided by the animal center of
Xinjiang Medical University [Urumqi, China; animal use license no.
SYXK (Sinkiang) 2011-010101]. The animals were housed in cages with
pathogen-free conditions, and free access to food and water, under
a 12-h light/dark cycle at a room temperature of 22°C with 45%
humidity, and normal air conditions (21% O2, 78%
N2 and 0.03% CO2). Chloral hydrate (350
mg/kg; intraperitoneal injection) was used to anesthetize rats for
surgery and blood collection. Subsequently, rapid cervical
dislocation was used to sacrifice the animals painlessly. All
experimental protocols in this study complied with the Guidelines
for the Care and Use of Laboratory Animals published by the US
National Institutes of Health (1996) and were approved by the
Animal Protection and Use Committee of Shihezi University (Shihezi,
China).
Animal model
Prior to surgery, all mice were fasted for 8 h, with
water freely available. The CLP procedure was performed as
previously described (21). Briefly,
the rats were anesthetized with chloral hydrate and then secured to
a sterile operating table. Under aseptic conditions, a 2-cm
incision was made in the abdomen, and the cecum was exposed layer
by layer. Prior to ligation of the cecum, feces were gently
squeezed to the distal end of the cecum, then the cecum was ligated
with a thin wire, and the cecum was pierced with an 18-gauge
needle. Any remaining contents were extruded through the puncture
site. Subsequently, the cecum was pushed back into the abdominal
cavity, the abdominal cavity was closed and the layers were
sutured. The sham group underwent the same procedure without CLP.
All rats were resuscitated using 0.9% sodium chloride brine
(subcutaneous injection, ~40 ml/kg), and following surgery, rats
were returned their cages with free access to food and water.
Experimental grouping
Rats (n=56) were randomly divided into 7 groups as
follows: Sham group, underwent exposure of the cecum, but not
ligation and perforation; sham + NaHS group, received the sham
procedure and was administered a 2-ml/kg intraperitoneal
administration of 8.9 µmol/kg NaHS 1 h following the surgery; sham
+ LY294002 group, underwent the sham procedure and was administered
a 2-ml/kg intraperitoneal injection of 40 mg/kg LY294002 following
the surgery; CLP group, treated with the CLP surgery; CLP + NaHS
group, underwent CLP and was administered NaHS (dosage and method
as in the sham + NaHS group); CLP + LY294002 group, underwent CLP
and was administered LY294002 following the surgery (dosage and
method as in the sham + LY294002 group); CLP + NaHS + LY294002
group, underwent CLP and was administered an intraperitoneal
injection of NaHS and LY294002 at the aforementioned doses. The
doses of LY294002 and NaHS were selected according to previous
studies (5,12). Following the procedures, all the rats
were free to drink and eat, and 12 h later they were anesthetized
again. The rat abdominal cavity was opened and blood was obtained
from the abdominal aorta. Following coagulation, blood samples were
centrifuged (3,500 × g, 15 min) and clear supernatants were
collected. A portion of each serum sample was sent to the First
Affiliated Hospital of Shihezi University for the measurement of
creatine kinase-MB (CK-MB) and lactate dehydrogenase (LDH). The
remaining samples were frozen at −80°C for subsequent analysis. The
rats were sacrificed by cervical dislocation immediately after
blood collection. Heart tissue was obtained immediately after
sacrifice. Half of each heart tissue sample was fixed in
paraformaldehyde (4%) for histomorphological analysis, and the
other half was stored at −80°C for western blot analysis and other
experiments.
Measurements of CK-MB, LDH and cardiac
TnI (cTnI) levels in serum
Serum CK-MB and LDH levels in the rats were measured
using an automatic biochemical analyzer (Modular DPP H7600; Roche
Diagnostics, Basel, Switzerland). An ELISA kit was used to measure
serum cTnI levels according to the manufacturer's instructions.
Measurement of inflammatory cytokines
in serum
ELISA kits were used to determine the TNF-α, IL-6
and IL-10 levels in serum according to the manufacturer's
instructions.
Measurement of H2S
levels
H2S levels in serum were detected using a
H2S testing kit purchased from Nanjing Institute of
Bioengineering (Nanjing, China). H2S reacts with zinc
acetate, N,N-dimethylphenylenediamine and ammonium ferric sulfate
in the kit to form methylene blue, which has a maximum absorption
peak at 665 nm. The H2S content was calculated by
determining the absorbance value of methylene blue, and the plasma
H2S level is expressed in µmol/l.
Histological analysis
The tissue fixed in paraformaldehyde was
conventionally embedded, sliced (4-µm thick) and dewaxed with
xylene, then washed with ethanol at various levels. Subsequently,
sections were stained with hematoxylin and eosin (H&E) at room
temperature for ~2 min, dehydrated, dewaxed and sealed. Finally,
images of sections were captured using an optical microscope
(Olympus Corporation, Tokyo, Japan).
Western blot analysis
The frozen myocardial tissue was lysed with lysis
buffer (cat. no. R0030; Beijing Solarbio Science & Technology
Co., Ltd., Beijing, China) on ice, and following protein
extraction, a NanoDrop spectrophotometer (Thermo Fisher Scientific,
Inc., Waltham, MA, USA) was used to determine the concentration of
tissue proteins. A total of 10 µl of protein samples (40 mg/ml)
were separated by SDS-PAGE using 10% gels, and then transferred to
a polyvinylidene fluoride membrane. The membrane was blocked with
5% bovine serum albumin (cat. no. 4240GR100; BioFroxx; NeoFROXX
GmbH, Hesse, Germany) for 2 h at room temperature. Membranes were
incubated with antibodies against Akt (1:800 dilution), p-Akt
(1:800 dilution), NF-κB (1:800 dilution), PI3K (1:800 dilution),
p-PI3K (1:800 dilution), Bcl-2 (1:200 dilution), Bax (1:200
dilution), caspase-3 (1:500 dilution) and β-actin (1:1,000
dilution) at 4°C for 12–18 h. The membranes were then washed six
times in TBS-Tween (5 min each time), and incubated with
horseradish peroxidase (HRP)-conjugated goat anti-rabbit secondary
antibody (1:5,000 dilution) for 2–3 h at room temperature (β-actin
was detected using HRP-conjugated goat anti-mouse secondary
antibody under the same conditions). Following incubation, the
membranes were washed six times over a total of 30 min. The
membranes were reacted with an enhanced chemiluminescence reagent
(cat. no. WBKLS0100; EMD Millipore, Billerica, MA, USA) and
developed using X-ray film. The band intensities were quantified
using ImageJ software (Java 1.6.0.24; version 1.51k; National
Institutes of Health, Bethesda, MD, USA).
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) assay
Apoptosis of myocardial cells was determined by a
TUNEL assay using the in situ Cell Death Detection kit, POD.
Slices (4-µm thick) of heart tissue were incubated in a 68°C oven
for 1 h, and then dewaxed in xylene and gradient alcohol. The
sliced tissue was then washed three times with PBS (5 min each
wash). After washing, the sections were placed in hydrogen peroxide
and soaked for 10 min. Subsequently, further washes with PBS were
performed, and then sections were steamed in a pressure cooker for
5 min with citrate buffer at pH 6.0, and washed again three times
with PBS (5 min each wash). Samples were incubated with 50 µl TUNEL
reaction mixture (1:12) at 37°C for 1 h in the dark. The slides
were then rinsed three times with PBS, 50 µl POD substrate was
added to each section, and the samples were incubated at 37°C for
30 min in the dark. Following incubation, the sections were washed
three times with PBS, and then 50 µl diaminobenzidine coloring
solution was added dropwise, and the color development was stopped
with distilled water when an appropriate degree of coloration was
achieved. The sections were counterstained with hematoxylin for 2
min, then dehydrated in alcohol and sealed with neutral gum.
Finally, a light microscope was used to calculate the apoptotic
index. Dark brown staining indicated TUNEL-positive cells, and five
high power fields of the slices were randomly selected to calculate
the proportion of positive cells.
Statistical analysis
Each experiment was repeated three times. All data
were analyzed using SPSS 22.0 software (IBM Corp., Armonk, NY,
USA). All data are expressed as the mean ± standard error. The data
were analyzed using one-way analysis of variance followed by least
significant difference tests. P<0.05 was considered to indicate
a statistically significant difference.
Results
Confirmation of the rat model of
sepsis
The rats in the CLP group exhibited the behavioral
characteristics of sepsis, including malaise, fever, chills,
piloerection, generalized weakness and reduced gross motor
activity, as well as weight loss and increased proinflammatory
cytokine levels in the serum. One mortality in the CLP group and
two in the CLP+LY294002 group were observed.
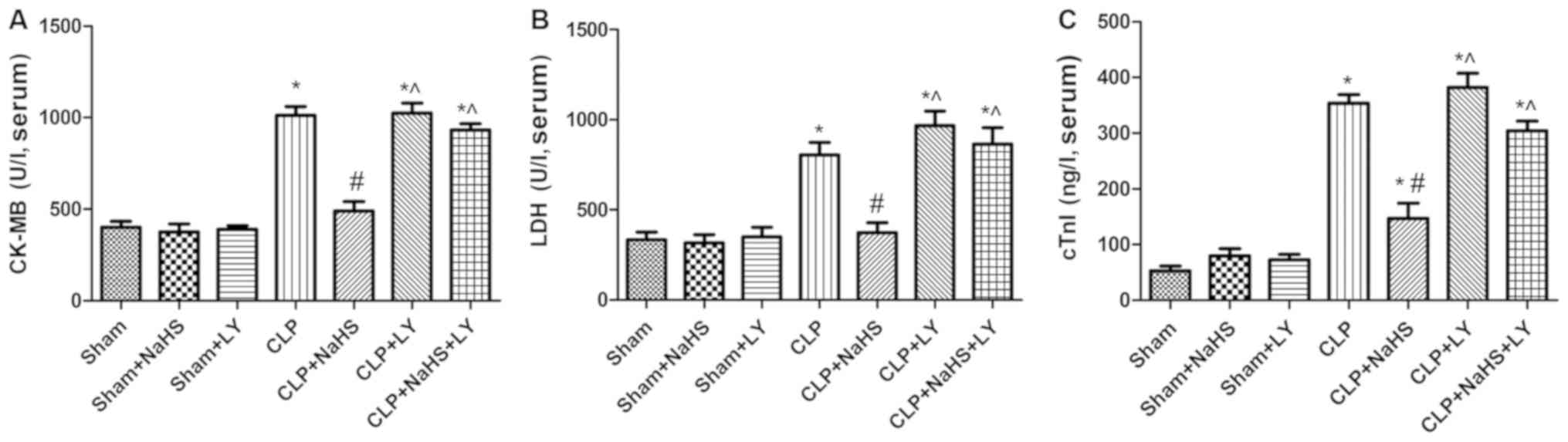
Effects of NaHS on myocardial enzyme
serum levels
As presented in Fig.
1, CK-MB, LDH and cTnI concentrations exhibited no significant
differences among the sham, sham + NaHS and sham + LY groups. There
was a significant increase in serum CK-MB, LDH and cTnI in the CLP
group compared with the sham group, and rats in the NaHS treatment
group exhibited reductions in CK-MB, LDH and cTnI levels compared
with those in the CLP group. However, these reductions were
attenuated by LY294002 (a PI3K/Akt pathway-specific inhibitor).
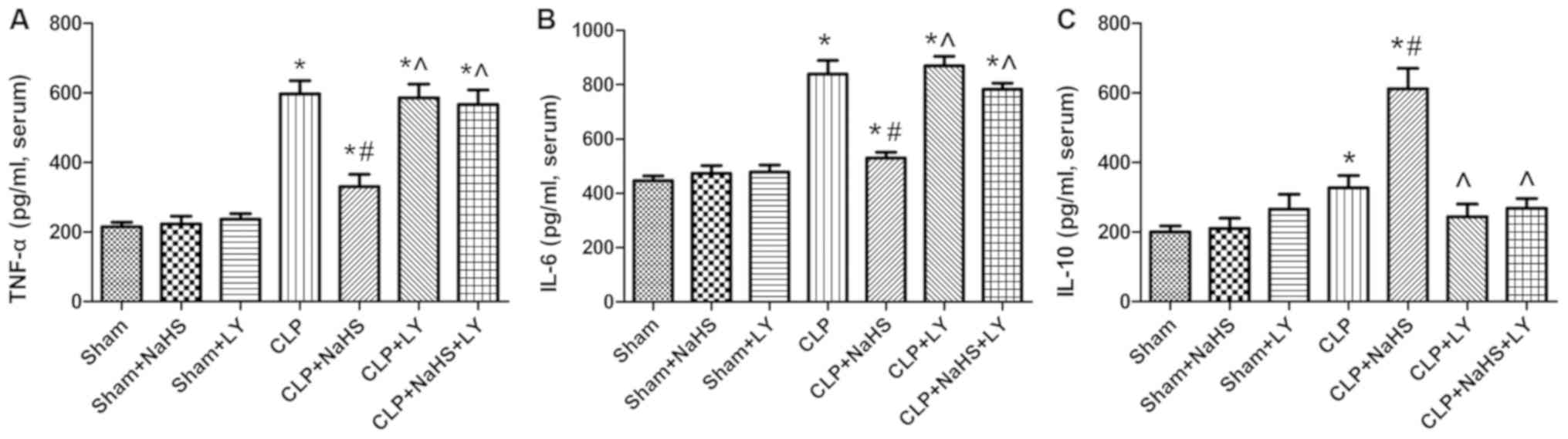
NaHS reduces inflammatory reactions in
septic rats
Inflammatory factors were measured in the serum of
model rats. Following CLP, the levels of the inflammatory factors
TNF-α, IL-6 and IL-10 in the serum of rats were increased
significantly compared with those in the sham group (P<0.05;
Fig. 2). The administration of NaHS
decreased the levels of TNF-α and IL-6 in the serum compared with
those in the CLP group (P<0.05), whereas the level of IL-10 was
significantly increased. However, the administration of LY294002
eliminated the effects of NaHS (P<0.05 vs. CLP + NaHS group;
Fig. 2).
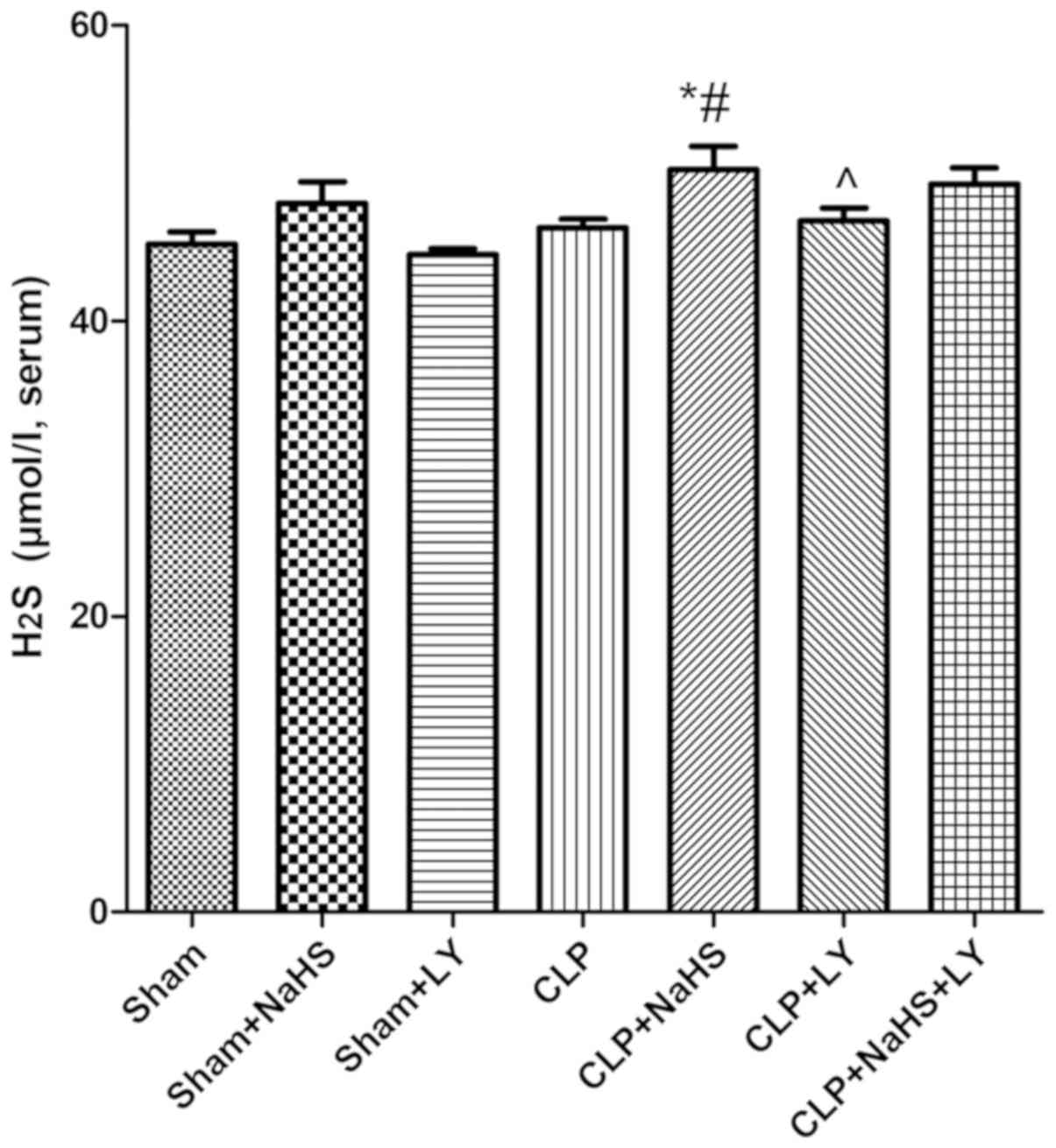
H2S levels
Although level of H2S was increased in
the CLP group, there was no significant difference in
H2S levels between the CLP and sham groups. Following
the administration of NaHS, the serum H2S content was
increased significantly compared with that in the CLP group
(P<0.05; Fig. 3). This finding
indicated that the administration of NaHS increases serum
H2S content (Fig. 3).
NaHS alleviates myocardial damage
caused by sepsis
H&E staining demonstrated that the sham group
had normal myocardial fiber morphology and a regular arrangement of
myocardial cells, with no abnormalities of the interstices and
microvessels (Fig. 4). However,
myocardial cells were disordered in the CLP group, with broken
myocardial fibers, and parts of the myocardium exhibited
vacuolization changes and inflammatory cell infiltration. The
administration of NaHS reduced the degree of myocardial damage, and
LY294002 eliminated the protective effect of NaHS (Fig. 4).
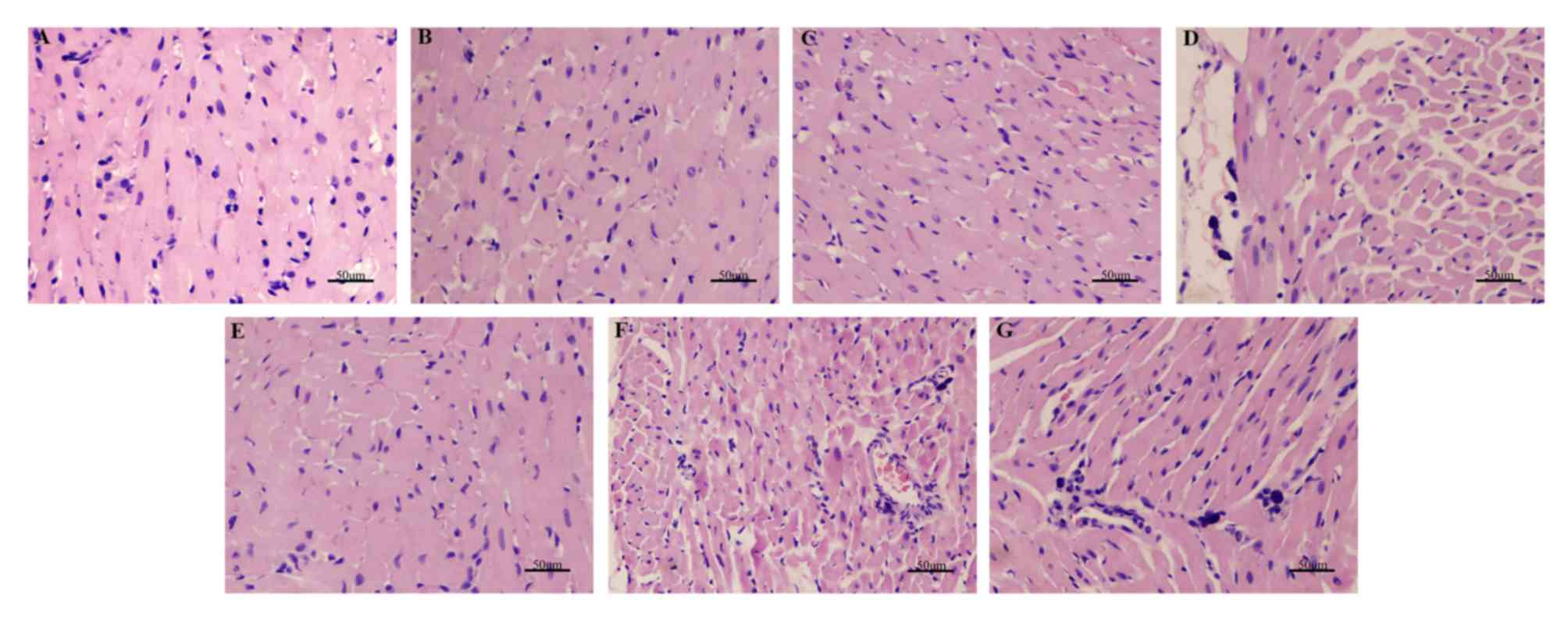 | Figure 4.H&E staining demonstrating the
morphology of myocardial cells in each group. Magnification, ×200.
(A) Sham, (B) sham + NaHS, (C) sham + LY, (D) CLP, (E) CLP + NaHS,
(F) CLP + LY and (G) CLP + NaHS + LY group. H&E, hematoxylin
and eosin; NaHS, sodium hydrosulfide; CLP, cecal ligation and
puncture; LY, LY294002. |
NaHS reduces myocardial apoptosis in
septic rats
TUNEL staining was performed to investigate the
effect of NaHS on myocardial apoptosis in rats with sepsis
(Fig. 5). Myocardial apoptosis was
significantly increased in the CLP group compared with the sham
group (P<0.05). However, following the administration of NaHS,
the apoptosis rate of cardiomyocytes was significantly reduced
compared with that in the CLP group, and LY294002 treatment
reversed these alterations (Fig.
5).
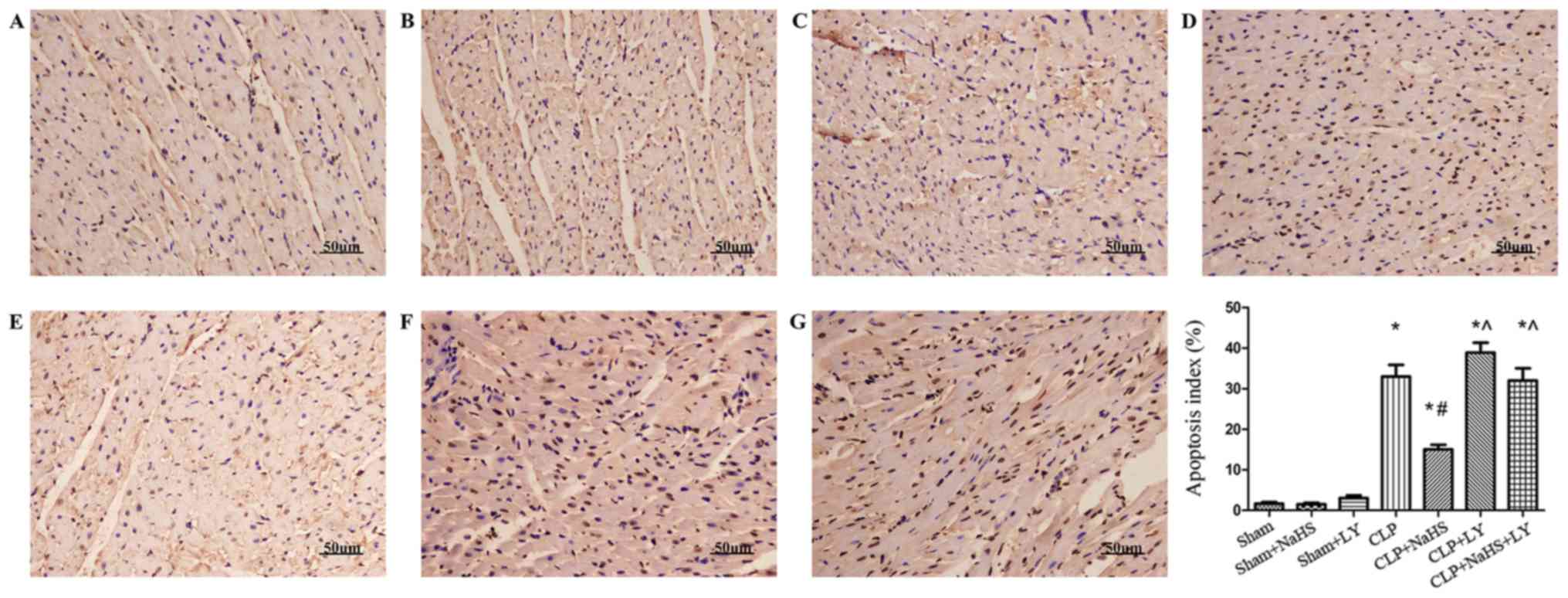 | Figure 5.Apoptosis detected by TUNEL assay.
Magnification, ×200. (A) Sham, (B) sham + NaHS, (C) sham + LY, (D)
CLP, (E) CLP + NaHS, (F) CLP + LY and (G) CLP + NaHS + LY group.
The subgroup apoptosis index was calculated. *P<0.05 vs. sham
group; #P<0.05 vs. CLP group; ^P<0.05 vs. CLP +
NaHS group. TUNEL, terminal deoxynucleotidyl transferase dUTP nick
end labeling; NaHS, sodium hydrosulfide; CLP, cecal ligation and
puncture; LY, LY294002. |
Effect of NaHS on PI3K, Akt, NF-κB,
Bcl-2, Bax and caspase-3
The p-PI3K/PI3K and p-Akt/Akt ratios, and the levels
of NF-κB, Bcl-2, Bax and caspase-3 were determined using western
blot analysis to further understand the protective mechanism of
NaHS against the myocardial injury caused by sepsis. The
p-PI3K/PI3K and p-Akt/Akt ratios, and NF-κB, Bax and caspase-3
levels were increased by CLP compared with those in the sham group,
whereas the level of Bcl-2 was decreased (Figs. 6 and 7). Following the administration of NaHS,
the phosphorylation of PI3K and Akt was further increased and the
expression of Bcl-2 was increased compared with that in the CLP
group, whereas the expression levels of NF-κB, Bax and caspase-3
were decreased. The addition of LY294002 eliminated these
NaHS-induced effects.
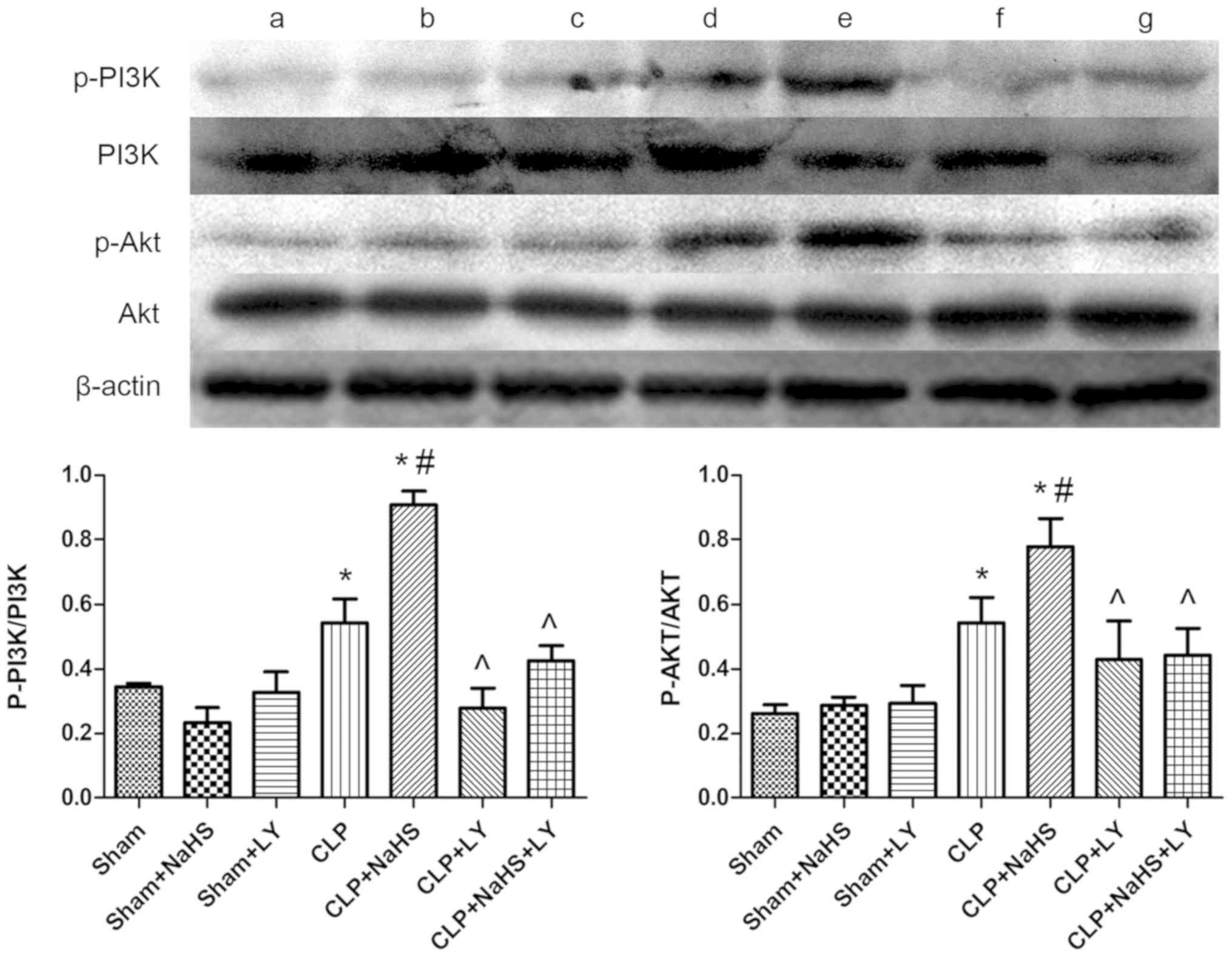 | Figure 6.Changes in PI3K, p-PI3K, Akt and
p-Akt in each group. Representative western blotting images are
presented. Lanes: a, Sham; b, sham + NaHS; c, sham + LY; d, CLP; e,
CLP + NaHS; f, CLP + LY; g, CLP + NaHS + LY. Quantified data are
expressed as the mean ± standard error. *P<0.05 vs. sham group;
#P<0.05 vs. CLP group; ^P<0.05 vs. CLP + NaHS
group. PI3K, phosphatidylinositol-3-kinase; p-, phospho-; Akt,
protein kinase B; NaHS, sodium hydrosulfide; CLP, cecal ligation
and puncture; LY, LY294002. |
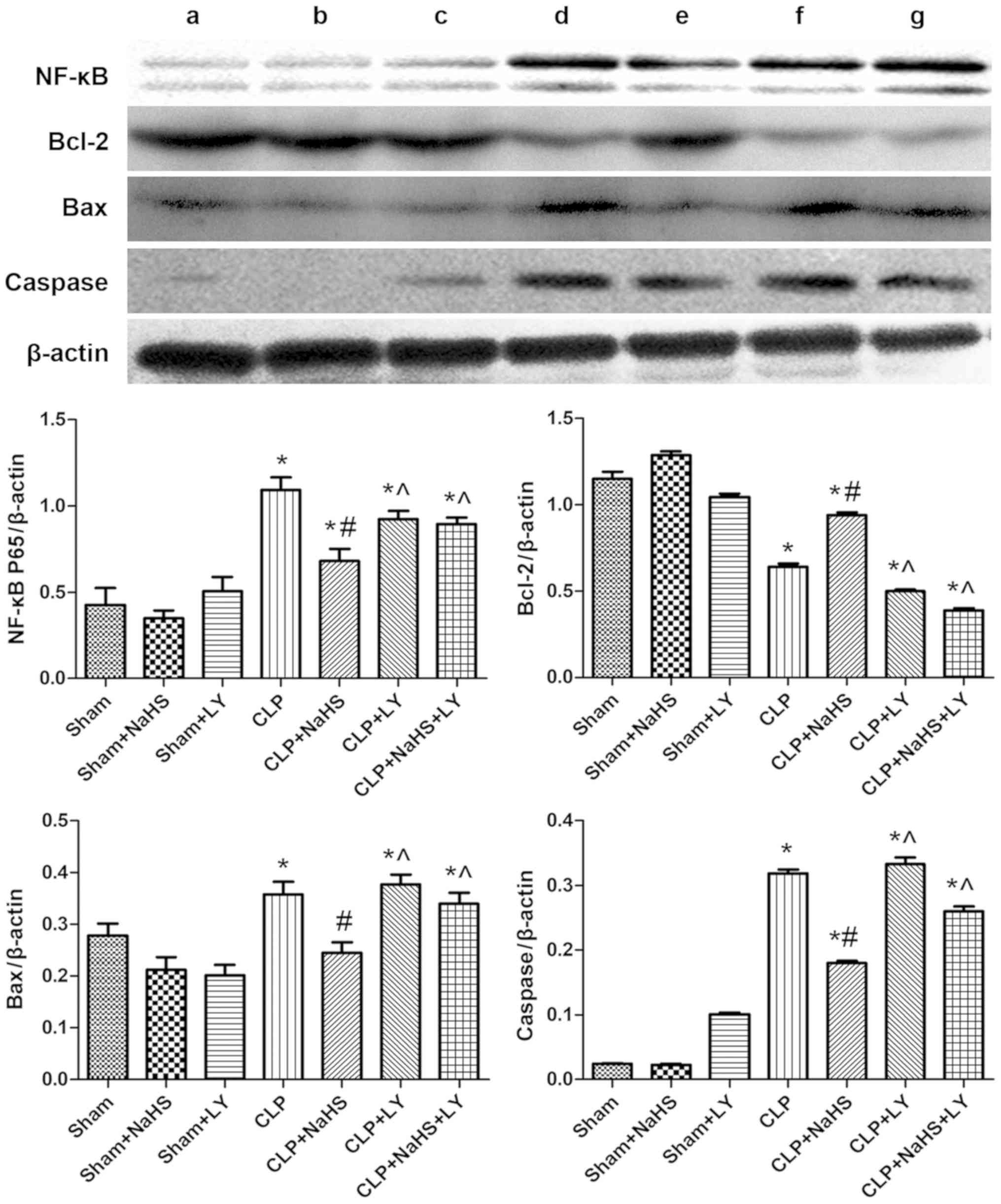 | Figure 7.Expression of NF-κB, Bcl-2, Bax and
caspase in each group. Representative western blotting images are
presented. Lanes: a, Sham; b, sham + NaHS; c, sham + LY; d, CLP; e,
CLP + NaHS; f, CLP + LY; g, CLP + NaHS + LY. Quantified data are
expressed as the mean ± standard error. *P<0.05 vs. sham group;
#P<0.05 vs. CLP group; ^P<0.05 vs. CLP + NaHS
group. NF-κB, nuclear factor-κB; Bcl-2, B-cell lymphoma-2; Bax,
Bcl-2-associated X protein; NaHS, sodium hydrosulfide; CLP, cecal
ligation and puncture; LY, LY294002. |
Discussion
As reported in our previous study, exogenous
H2S can improve sepsis-induced heart dysfunction
(12). In the current study,
exogenous H2S significantly reduced the expression of
myocardial injury markers (CK-MB, LDH and cTnI), reduced the
expression of pro-inflammatory factors (TNF-α and IL-6) and induced
the expression of the anti-inflammatory factor IL-10 following CLP.
Furthermore, the apoptosis of cardiomyocytes was reduced by
H2S in the CLP-induced sepsis model. However, the
effects of exogenous H2S on myocardial injury in sepsis
were counteracted by the PI3K signaling pathway inhibitor LY294002,
indicating that the PI3K/Akt signaling pathway may partially
mediate the effect of H2S.
Sepsis is caused by a dysfunctional response to
infection in the host, resulting in life-threatening organ
dysfunction (22). Sepsis has high
morbidity and mortality rates, and significant treatment costs in
developed and developing countries (23). During sepsis, various factors,
including cytokines, eicosanoids, reactive oxygen species and
nitrogen species, accelerate the development of the symptoms
observed in patients with sepsis and cause severe systemic
inflammatory reactions, which can ultimately lead to multiple organ
failure (24). The heart is one of
the most vulnerable target organs in sepsis (4,5).
Currently, there is no specific treatment for sepsis-induced
cardiac insufficiency.
H2S is one of a family of gaseous
signaling molecules, which includes nitric oxide (NO) and carbon
monoxide. H2S is typically considered to be a toxic gas;
however, in the past 30 years, the understanding of the effects
H2S has changed markedly (25). Numerous in vitro and in
vitro experiments have demonstrated that H2S has
protective effects in various tissues and cells (26–28).
Abdelrahman et al (29)
reported that H2S has a protective effect on CLP-induced
cardiac dysfunction, by reducing tachycardia, mortality, serum
CK-MB, cTnI, C-reactive protein and LDH, and cardiac and aortic
malondialdehyde levels. Chen et al (30) demonstrated that the exogenous
administration of NaHS ameliorated septic-induced renal dysfunction
by inhibiting inflammation and oxidative stress through the
Toll-like receptor 4/NACHT, LRR and PYD domains-containing protein
3 signaling pathway. Furthermore, Ahmad et al (31) reported that the intraperitoneal
injection of an appropriate dose of H2S improved the
survival rate of septic rats and reduced the inflammatory reaction.
This finding is supported by the results of the present study,
demonstrating that the exogenous administration of NaHS is able to
attenuate inflammation during sepsis and reduce myocardial cell
apoptosis. However, the findings of Zhang et al (32,33) were
contrary to the results of the present study. These previous
studies indicated that H2S may exert pro-inflammatory
effects during sepsis by regulating the inflammatory response via
extracellular signal-regulated kinase-NF-κB pathway activation. We
concur with the in-depth analysis by Ahmad et al (31) regarding the differences in findings
from those of Zhang et al The differences between studies
may be due to the following: i) The dose of NaHS used (0.89 µmol/kg
was used in the current study, whereas Zhang et al
administered a high dose (10 mg/kg) by intraperitoneal injection);
and ii) the time of administration. The serum H2S level
in each group was measured in the present study. The level of
endogenous H2S in the serum of rats appeared to increase
following CLP, which was consistent with the findings of Bee et
al (34). This phenomenon may be
associated with the upregulation of inducible nitric oxide synthase
(iNOS) expression during septic shock. Due to the increase in iNOS,
the level of NO is also increased, which can induce cystathionine
γ-lyase expression, resulting in higher H2S levels
(35). No significant difference in
H2S level between the sham and CLP groups was observed
and, although the production of endogenous H2S was
increased following the occurrence of sepsis, the increase was not
significant. Following the administration of exogenous NaHS, a
statistically significant increase in the serum level of
H2S was detected, indicating that the administration of
NaHS produced the desired effect.
CK, CK-MB and LDH are indicators used for the
diagnosis and detection of myocardial injury (36,37).
cTnI and cTnT have high specificity, and Tn levels are directly
proportional to the degree of myocardial damage (38). In the current study, the analysis of
serum myocardial injury markers in each group revealed that
myocardial enzymes were significantly elevated in the sepsis group,
indicating that sepsis had an adverse effect on the myocardium. The
intraperitoneal injection of NaHS reduced the serum myocardial
enzyme level. Similar changes in Tn levels were observed. H&E
staining of the heart tissue revealed that the sham-operated group
exhibited tightly arranged myocardial fibers with uniform coloring,
and no edema, congestion or exudation. Following CLP, the presence
of edema, degeneration and myocardial fiber breakage was observed
in parts of the myocardial tissue. Interstitial blood vessels were
congested and inflammatory cells had infiltrated the tissue. The
administration of NaHS reduced the degree of myocardial damage
compared with that in the sepsis group. The results of Abdelrahman
et al (29) support the
findings of the present study.
Following the onset of sepsis, a large amount of
endotoxin is released, causing a series of pro-inflammatory factor
cascades to be activated, including TNF-α and IL-1 (39,40).
Among the pro-inflammatory cytokines, TNF-α induces cardiomyocyte
apoptosis (41). A previous study
reported that the infusion of TNF-α monoclonal antibodies to septic
mice transiently improves ventricular function, suggesting that
TNF-α may reduce damage to the myocardium during sepsis (42). Another study demonstrated that the
inflammation and apoptosis induced during sepsis is associated with
the production and release of reactive oxygen species and
inflammatory mediators. These substances may be involved in the
activation of inducible pathways, such as NF-κB (43). Additionally, sepsis induces the
expression of iNOS, which in turn produces high levels of NO.
Excessive NO may cause cytotoxicity due to an increase in
peroxynitrite, which can lead to cardiac dysfunction (44). In the current study, the CLP-induced
inflammatory responses and cytotoxicity were characterized by
elevated levels of TNF-α, IL-6, CK-MB and LDH in serum. Bcl-2 was
decreased, and Bax and caspase levels were increased by CLP, and
the TUNEL assay confirmed that the apoptosis rate was increased
following CLP. These results indicated that CLP induced
inflammation and apoptosis. Furthermore, the administration of NaHS
to rats that received CLP reduced the levels of the inflammatory
response markers in the serum and reduced cardiomyocyte apoptosis.
These results indicated that H2S ameliorated the degree
of sepsis-induced myocardial damage, potentially via
anti-inflammatory and anti-apoptotic effects.
In numerous studies of sepsis, PI3K and downstream
Akt have been demonstrated to be involved in the regulation of cell
activation, inflammation and apoptosis (5,45,46).
Studies have reported that heat shock protein A12B and heat shock
protein 27 can attenuate cardiac dysfunction caused by endotoxins.
The mechanism of this effect may be associated with the activation
of PI3K/Akt (47,48). Additionally, other studies have
demonstrated that H2S activates the PI3K/Akt pathway
(18,49). The results of the current study
suggest that the exogenous administration of H2S
significantly increases PI3K and Akt phosphorylation in rats with
CLP-induced sepsis. In order to confirm this hypothesis, a PI3K/Akt
pathway specific inhibitor, LY294002, was used. The results
revealed that the anti-inflammatory and anti-apoptotic effects of
H2S were eliminated by the intraperitoneal injection of
LY294002 in the model rats.
H2S has been widely used in animal
studies; however, it is not clear whether H2S has
beneficial or damaging effects in vivo. Currently, clinical
experiments have confirmed that H2S is
anti-inflammatory, reduces myocardial fibrosis and protects the
myocardiu (50). However, different
H2S donors may have different mechanisms of action
(51). Future studies are required
to investigate the specific mechanisms of H2S donors in
complex cardiovascular signaling pathways.
In conclusion, the present study demonstrated that
exogenous H2S therapy provides an important protective
effect against sepsis-induced myocardial damage via activation of
the PI3K/Akt pathway. H2S inhibits inflammation and
apoptosis, and reduces myocardial dysfunction during sepsis.
Acknowledgements
Not applicable.
Funding
This study was supported by a grant from XPCC
Science and Technology Research and Achievement Transformation
Project (grant no. KC0038).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JPL, JHL and QC jointly conceived and designed this
study. JPL, GW and LL conducted the animal experiment. JPL analyzed
the data and completed the first draft. JHL, PT, BG and QC reviewed
and improved the paper. PT and BG revised the manuscript critically
for important intellectual content. All authors read and approved
the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Animal
Protection and Use Committee of Shihezi University (Shihezi,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Glossary
Abbreviations
Abbreviations:
|
H2S
|
hydrogen sulfide
|
|
PI3K/Akt
|
phosphatidylinositol-3-kinase/protein
kinase B
|
|
CLP
|
cecal ligation and puncture
|
|
IL
|
interleukin
|
|
TNF-α
|
tumor necrosis factor-α
|
|
NF-κB
|
nuclear factor-κB
|
References
|
1
|
Singer M, Deutschman CS, Seymour CW,
Shankar-Hari M, Annane D, Bauer M, Bellomo R, Bernard GR, Chiche
JD, Coopersmith CM, et al: The Third International Consensus
Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA.
315:801–810. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fleischmann C, Scherag A, Adhikari NK,
Hartog CS, Tsaganos T, Schlattmann P, Angus DC and Reinhart K;
International Forum of Acute Care Trialists, : Assessment of Global
incidence and mortality of hospital-treated sepsis. Current
estimates and limitations. Am J Respir Crit Care Med. 193:259–272.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vincent JL, Marshall JC, Namendys-Silva
SA, VinFrancois B, Martin-Loeches I, Lipman J, Reinhart K,
Antonelli M, Pickkers P, Njimi H, et al: Assessment of the
worldwide burden of critical illness: The intensive care over
nations (ICON) audit. Lancet Respir Med. 2:380–386. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zaky A, Deem S, Bendjelid K and Treggiari
MM: Characterization of cardiac dysfunction in sepsis: An ongoing
challenge. Shock. 41:12–24. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
An R, Zhao L, Xi C, Li H, Shen G, Liu H,
Zhang S and Sun L: Melatonin attenuates sepsis-induced cardiac
dysfunction via a PI3K/Akt-dependent mechanism. Basic Res Cardiol.
111:82016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stein A and Bailey SM: Redox Biology of
Hydrogen Sulfide: Implications for Physiology, Pathophysiology and
Pharmacology. Redox Biol. 1:32–39. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shen Y, Shen Z, Luo S, Guo W and Zhu YZ:
The cardioprotective effects of hydrogen sulfide in heart diseases:
From molecular mechanisms to therapeutic potential. Oxid Med Cell
Longev. 2015:9251672015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu Y, Chen X, Pan TT, Neo KL, Lee SW, Khin
ES, Moore PK and Bian JS: Cardioprotection induced by hydrogen
sulfide preconditioning involves activation of ERK and PI3K/Akt
pathways. Pflugers Arch. 455:607–616. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Donnarumma E, Trivedi RK and Lefer DJ:
Protective Actions of H2S in acute myocardial infarction and heart
failure. Compre Physiol. 7:583–602. 2017. View Article : Google Scholar
|
|
10
|
Patel VB, McLean BA, Chen X and Oudit GY:
Hydrogen sulfide: An old gas with new cardioprotective effects.
Clin Sci (Lond. 128:321–323. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Calvert JW, Coetzee WA and Lefer DJ: Novel
insights into hydrogen sulfide-mediated cytoprotection. Antioxid
Redox Signal. 12:1203–1217. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li X, Cheng Q, Li J, He Y, Tian P and Xu
C: Significance of hydrogen sulfide in sepsis-induced myocardial
injury in rats. Exp Ther Med. 14:2153–2161. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhai J and Guo Y: Paeoniflorin attenuates
cardiac dysfunction in endotoxemic mice via the inhibition of
nuclear factor-κB. Biomed Pharmacother. 80:200–206. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao P, Wang Y, Zeng S, Lu J, Jiang TM and
Li YM: Protective effect of astragaloside IV on
lipopolysaccharide-induced cardiac dysfunction via downregulation
of inflammatory signaling in mice. Immunopharmacol Immunotoxicol.
37:428–433. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Luo K, Long H, Xu B and Luo Y: Apelin
attenuates postburn sepsis via a phosphatidylinositol
3-kinase/protein kinase B dependent mechanism: A randomized animal
study. Int J Surg. 21:22–27. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Williams DL, Li C, Ha T, Ozment-Skelton T,
Kalbfleisc JH, Preiszner J, Brooks L, Breuel K and Schweitzer JB:
Modulation of the phosphoinositide 3-kinase pathway alters innate
resistance to polymicrobial sepsis. J Immunol. 172:449–456. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun N, Wang H, Ma L, Lei P and Zhang Q:
Ghrelin attenuates brain injury in septic mice via PI3K/Akt
signaling activation. Brain Res Bull. 124:278–285. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shao M, Zhuo C, Jiang R, Chen G, Shan J,
Ping J, Tian H, Wang L, Lin C and Hu L: Protective effect of
hydrogen sulphide against myocardial hypertrophy in mice.
Oncotarget. 8:22344–22352. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tamizhselvi R, Moore PK and Bhatia M:
Hydrogen sulfide acts as a mediator of inflammation in acute
pancreatitis: In vitro studies using isolated mouse pancreatic
acinar cells. J Cell Mol Med. 11:315–326. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Y, Liao R, Qiang Z and Zhang C:
Pro-inflammatory cytokine-driven PI3K/Akt/Sp1 signalling and H2S
production facilitates the pathogenesis of severe acute
pancreatitis. Biosci Rep. 37(pii): BSR201604832017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rittirsch D, Huber-Lang MS, Flierl MA and
Ward PA: Immunodesign of experimental sepsis by cecal ligation and
puncture. Nat Protoc. 4:31–36. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rhodes A, Evans LE, Alhazzani W, Levy MM,
Antonelli M, Ferrer R, Kumar A, Sevransky JE, Sprung CL, Nunnally
ME, et al: Surviving Sepsis Campaign: International Guidelines for
Management of Sepsis and Septic Shock: 2016. Intensive Care Med.
43:304–377. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cohen J: The immunopathogenesis of sepsis.
Nature. 420:885–891. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kosir M and Podbregar M: Advances in the
diagnosis of sepsis: Hydrogen sulfide as a prognostic marker of
septic shock severity. Ejifcc. 28:134–141. 2017.PubMed/NCBI
|
|
25
|
Lavu M, Bhushan S and Lefer DJ: Hydrogen
sulfide-mediated cardioprotection: Mechanisms and therapeutic
potential. Clin Sci (Lond). 120:219–229. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang M, Shan H, Wang T, Liu W, Wang Y,
Wang L, Zhang L, Chang P, Dong W, Chen X and Tao L: Dynamic change
of hydrogen sulfide after traumatic brain injury and its effect in
mice. Neurochem Res. 38:714–725. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li L, Xiao T, Li F, Li Y, Zeng O, Liu M,
Liang B, Li Z, Chu C and Yang J: Hydrogen sulfide reduced renal
tissue fibrosis by regulating autophagy in diabetic rats. Mol Med
Rep. 16:1715–1722. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bhatia M, Wong FL, Fu D, Lau HY, Moochhala
SM and Moore PK: Role of hydrogen sulfide in acute pancreatitis and
associated lung injury. FASEB J. 19:623–625. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Abdelrahman RS, El-Awady MS, Nader MA and
Ammar EM: Hydrogen sulfide ameliorates cardiovascular dysfunction
induced by cecal ligation and puncture in rats. Hum Exp Toxicol.
34:953–964. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen Y, Jin S, Teng X, Hu Z, Zhang Z, Qiu
X, Tian D and Wu Y: Hydrogen Sulfide Attenuates LPS-Induced Acute
Kidney Injury by Inhibiting Inflammation and Oxidative Stress. Oxid
Med Cell Longev. 2018:67172122018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ahmad A, Druzhyna N and Szabo C: Delayed
treatment with sodium hydrosulfide improves regional blood flow and
alleviates cecal ligation and puncture (CLP)-Induced Septic Shock.
Shock. 46:183–193. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang H, Moochhala SM and Bhatia M:
Endogenous hydrogen sulfide regulates inflammatory response by
activating the ERK pathway in polymicrobial sepsis. J Immunol.
181:4320–4331. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang H, Zhi L, Moochhala S, Moore PK and
Bhatia M: Hydrogen sulfide acts as an inflammatory mediator in
cecal ligation and puncture-induced sepsis in mice by upregulating
the production of cytokines and chemokines via NF-kappaB. Am J
Physiol Lung Cell Mol Physiol. 292:L960–971. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bee N, White R and Petros AJ: Hydrogen
sulfide in exhaled gases from ventilated septic neonates and
children: A Preliminary Report. Pediatr Crit Care Med.
18:e327–e332. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Coletta C and Szabo C: Potential role of
hydrogen sulfide in the pathogenesis of vascular dysfunction in
septic shock. Curr Vasc Pharmacol. 11:208–221. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dahlin LG, Kagedal B, Nylander E, Olin C,
Rutberg H and Svedjeholm R: Early identification of permanent
myocardial damage after coronary surgery is aided by repeated
measurements of CK-MB. Scand Cardiovasc J. 36:35–40. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Y and Chen M: Fentanyl ameliorates
severe acute pancreatitis-induced myocardial injury in rats by
regulating NF-κB Signaling Pathway. Med Sci Monit. 23:3276–3283.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mair J: Cardiac troponin I and troponin T:
Are enzymes still relevant as cardiac markers? Clin Chim Acta.
257:99–115. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang B, Liu Y, Zhang JS, Zhang XH, Chen
WJ, Yin XH and Qi YF: Cortistatin protects myocardium from
endoplasmic reticulum stress induced apoptosis during sepsis. Mol
Cell Endocrinol. 406:40–48. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Comstock KL, Krown KA, Page MT, Martin D,
Ho P, Pedraza M, Castro EN, Nakajima N, Glembotski CC, Quintana PJ
and Sabbadini RA: LPS-induced TNF-alpha release from and apoptosis
in rat cardiomyocytes: Obligatory role for CD14 in mediating the
LPS response. J Mol Cell Cardiol. 30:2761–2775. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nakagawa T, Zhu H, Morishima N, Li E, Xu
J, Yankner BA and Yuan J: Caspase-12 mediates
endoplasmic-reticulum-specific apoptosis and cytotoxicity by
amyloid-beta. Nature. 403:98–103. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Abraham E, Wunderink R, Silverman H, Perl
TM, Nasraway S, Levy H, Bone R, Wenzel RP, Balk R, Allred R, et al:
Efficacy and safety of monoclonal antibody to human tumor necrosis
factor alpha in patients with sepsis syndrome. A randomized,
controlled, double-blind, multicenter clinical trial. TNF-alpha MAb
Sepsis Study Group. JAMA. 273:934–941. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Williams DL, Ha T, Li C, Kalbfleisch JH
and Ferguson DA Jr: Early activation of hepatic NFkappaB and NF-IL6
in polymicrobial sepsis correlates with bacteremia, cytokine
expression, and mortality. Ann Sur. 230:95–104. 1999. View Article : Google Scholar
|
|
44
|
Preiser JC, Zhang H, Vray B, Hrabak A and
Vincent JL: Time course of inducible nitric oxide synthase activity
following endotoxin administration in dogs. Nitric Oxide.
5:208–211. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kim TH, Kim SJ and Lee SM: Stimulation of
the alpha7 nicotinic acetylcholine receptor protects against sepsis
by inhibiting Toll-like receptor via phosphoinositide 3-kinase
activation. J Infect Dis. 209:1668–1677. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gao M, Ha T, Zhang X, Wang X, Liu L,
Kalbfleisch J, Singh K, Williams D and Li C: The Toll-like receptor
9 ligand, CpG oligodeoxynucleotide, attenuates cardiac dysfunction
in polymicrobial sepsis, involving activation of both
phosphoinositide 3 kinase/Akt and extracellular-signal-related
kinase signaling. J Infect Dis. 207:1471–1479. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhou H, Qian J, Li C, Li J, Zhang X, Ding
Z, Gao X, Han Z, Cheng Y and Liu L: Attenuation of cardiac
dysfunction by HSPA12B in endotoxin-induced sepsis in mice through
a PI3K-dependent mechanism. Cardiovasc Res. 89:109–118. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
You W, Min X, Zhang X, Qian B, Pang S,
Ding Z, Li C, Gao X, Di R, Cheng Y and Liu L: Cardiac-specific
expression of heat shock protein 27 attenuated endotoxin-induced
cardiac dysfunction and mortality in mice through a
PI3K/Akt-dependent mechanism. Shock. 32:108–117. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tamizhselvi R, Sun J, Koh YH and Bhatia M:
Effect of hydrogen sulfide on the phosphatidylinositol
3-kinase-protein kinase B pathway and on caerulein-induced cytokine
production in isolated mouse pancreatic acinar cells. J Pharmacol
Exp Ther. 329:1166–1177. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hackfort BT and Mishra PK: Emerging role
of hydrogen sulfide-microRNA crosstalk in cardiovascular diseases.
Am J Physiol Heart Circ Physiol. 310:H802–H812. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chatzianastasiou A, Bibli SI, Andreadou I,
Efentakis P, Kaludercic N, Wood ME, Whiteman M, Di Lisa F, Daiber
A, Manolopoulos VG, et al: Cardioprotection by H2S Donors: Nitric
oxide-dependent and independent mechanisms. J Pharmacol Exp Ther.
358:431–440. 2016. View Article : Google Scholar : PubMed/NCBI
|