Introduction
The incidence of thyroid cancer (TC) is the highest
among endocrine malignancies (1–4), with
~62,450 new diagnoses and ~1,950 TC mortalities in 2015 alone
(5). Furthermore, the incidence of
TC has increased rapidly in recent years and as such has attracted
more scientific attention; the age-adjusted incidence of TC is
estimated to be 9.1 per 100,000 females and 2.9 per 100,000 males
in developed countries (6–9). TC can be divided into four histologic
groups, including papillary TC (PTC), poorly differentiated
carcinoma, follicular TC and anaplastic TC (10). Among these, PTC accounts for ~80–90%
of all patients with TC (10). PTC
has a poor prognosis, so studies assessing the molecular mechanism
of PTC development are urgently required (11–14).
MicroRNAs (miRNAs) are small non-coding RNA
molecules comprised of 20–22 nucleotides, which inhibit mRNA
expression at the post-transcriptional level (15,16).
Several studies have demonstrated that various miRNAs function as
promoters or suppressors in many types of tumor, meaning that the
identification of miRNAs may serve as a useful diagnostic and
therapeutic approaches to cancer (10,17,18).
Furthermore, previous studies have reported the use of miRNAs as
biomarkers and their impact on the development of therapeutic
strategies in various malignancies, including lung cancer (18), TC (19) and prostate cancer (20). Therefore, the identification of novel
biomarkers and molecular targets may provide more effective
treatment options for patients with PTC.
The results of the current study determined the
expression of miR-30a in two TC cell lines and identified the
effect of miR-30a on the viability, migration and invasion of PTC
cells. These data indicated that miR-30a was downregulated in PTC
cells, while its ectopic overexpression significantly inhibited the
viability, migration and invasion of PTC cells. Therefore, miR-30a
may have the potential to function as a diagnostic biomarker or a
curative target in the future diagnosis and treatment of patients
with PTC.
Materials and methods
Study sample
A total of 15 pairs of PTC tissues and matched
adjacent non-tumor tissues were obtained from Fenyang Prison
Hospital (Fenyang, China). All tissue samples were obtained
following the receipt of written informed consent. The current
study was approved by the Ethics Review Board of Fenyang College
Shanxi Medical University (Shanxi, China). Seven tissue samples
were from males and eight were from females. The average age of the
study population was 58.4 years (range, 47–78 years). The data
range of sample collection was between October 2017 and May 2018.
The patients who were diagnosed with PTC were included in the
study. The diagnoses were made by pathologists. Adjacent non-tumor
tissues were isolated ≥2 cm away from the tumor border and were
shown to be free of tumor cells via microscopy. The tissues were
fixed with 10% formalin for 24 h at room temperature, processed in
paraffin and sectioned using a microtome. The thickness of sections
was 20 µm. Hematoxylin and eosin staining was used to confirm the
diagnosis. Briefly, the tissue was stained in hematoxylin for 4 min
at room temperature, washed under running tap water for 5 min,
differentiated in 1% acid alcohol for 5 min at room temperature,
and under running tap water for 5 min. The tissues were stained in
1% eosin Y for 10 min at room temperature, washed under running tap
water for 5 min, and then dehydrated 95% ethanol and absolute
ethanol. Subsequently, the tissues were cleared in xylene. The
samples were observed under a light microscope at a magnification
of ×100. Following tissue collection, samples were frozen in liquid
nitrogen immediately, transported to the laboratory and stored at
−80°C for RNA isolation. Following resection, tissues were washed
with PBS, immediately frozen in liquid nitrogen and stored at
−80°C. The expression of miR-30a in PTC tissue was then compared
with adjacent non-tumor tissues.
Cell culture
The TPC-1 cell line and the normal PTC cell line
(HT-ori3) were purchased from the American Type Culture Collection
(Manassas, VA, USA). Cells were maintained in RPMI-1640 medium
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (Invitrogen; Thermo Fisher
Scientific, Inc.), 100 IU/ml of penicillin and 100 µg/ml of
streptomycin. All cells were cultured in a humidified incubator
containing 5% CO2 at 37°C.
miRNA transfection
miR-30a mimics (20 nmol/l) and negative control (NC)
miRNA (20 nmol/l) were acquired from Shanghai GenePharma Co., Ltd.
(Shanghai, China). The miR-30a mimics sequence was
5′-UGUAAACAUCCUCGACUGGAAG-3′. The NC miRNA sequence was
5′-ACAUUUGUAGGAGCUGACCGGC-3′. Lipofectamine 2000®
(Invitrogen; Thermo Fisher Scientific, Inc.) was subsequently
utilized to perform transient transfection following incubation at
37°C for 6 h. Transfected cells were collected and purified after
48 h incubation.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA was extracted from cultured cells and tissues
using the TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Subsequently, RNA was reverse transcribed using
the RevertAid First Strand cDNA Synthesis kit (Thermo Fisher
Scientific, Inc.). qPCR was performed to determine the expression
of miR-30a and E2F transcription factor 7 (E2F7) using SYBRGreen
(Qiagen, Inc., Valencia, CA, USA). RT-qPCR amplification was
performed for 40 cycles using the following thermal profile:
Denaturation for 15 sec at 95°C, annealing and extension for 1 min
at 60°C. The following primers were utilized: miR-30a forward,
5′-CGCGATGTGTAAACATCCTCGAC-3′ and reverse,
5′-ATCCAGTGCAGGGTCCGAGG-3′; E2F7 forward,
5′-ACCCTCAGATTCCACAGACC-3′ and reverse, 5′-AGTTTGCTGTTGCCTTTCCT-3′;
U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′. U6 served as an internal control.
Furthermore, the 2-∆∆Cq method was utilized for RNA quantification
and associated analysis (21).
MTT assay
Cell viability was assessed via an MTT assay.
Cultured TPC-1 cells were first transfected with the miRNA mimics,
then seeded into 96-well plates (~4,000 cells/well). Subsequently,
MTT solution (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was
added to each well and incubated at 37°C for 4 h. A solubilization
solution, dimethyl sulfoxide, was added to dissolve the insoluble
purple formazan. A microplate reader was used for detecting the
viability of cells at an absorbance at 490 nm.
Invasion and migration assays
Transwell inserts that were precoated with Matrigel
(BD Biosciences, San Jose, CA, USA) were utilized to identify the
effect of miR-30a on the invasive capacity of PTC cells. PTC cells
(1×105) were suspended in Dulbecco's Modified Eagle's medium (DMEM;
Thermo Fisher Scientific, Inc.) without serum. Subsequently, these
cells and medium were seeded into the upper chamber of the system
and DMEM containing 20% FBS was added to the lower chamber. Cells
were incubated at 37°C for 24 h and fixed using 4% polyoxymethylene
at 4°C for 15 min. Following 15 min of incubation at 4°C, samples
were stained using 0.1% crystal violet dye for 10 min at 37°C. The
experimental procedure for the migration assay was the same as
aforementioned, except the transwell inserts were not coated with
Matrigel prior to experimentation. A light microscope was utilized
to visualize the results at magnification, ×200. Each assay was
performed in triplicate and repeated three times.
Western blotting
Protein from TPC-1 cells was extracted using
modified radioimmunoprecipitation assay buffer containing 0.5%
sodium dodecyl sulfate (SDS) in the presence of a proteinase
inhibitor cocktail (all Roche Applied Science, Madison, WI, USA) at
96 h following transfection. A bicinchoninic acid assay was used
for the detection of protein concentration. Protein (2 µg per lane)
was then separated using 10% SDS-PAGE and transferred onto a
polyvinylidene difluoride membrane. Membranes were blocked with 5%
non-fat at room temperature for 1 h and incubated with the
following primary antibodies at 4°C overnight: Anti-E2F7 (cat. no.
sc-32574; 1:5,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
and anti-GAPDH (cat. no. 2118; 1:5,000; Cell Signaling Technology,
Inc.). Samples were then incubated with horseradish
peroxidase-conjugated goat anti-rabbit immunoglobulin G secondary
antibodies (cat. no. 65-6120; 1:5,000; Invitrogen; Thermo Fisher
Scientific, Inc.) at 4°C overnight. Immunoreactive bands were
visualized on film by enhanced chemiluminescent substrate (Pierce;
Thermo Fisher Scientific, Inc.). GAPDH was utilized as an internal
control and data were analyzed using ImageJ software version 1.8
(National Institutes of Health, Bethesda, MD, USA).
Luciferase reporter assay
TargetScan (www.targetscan.org/) was used to predict the target
gene of miR-30a. To determine the association between miRNA and
target genes, PCR was used to amplify the 3′-untranslated region
(UTR) of human E2F7, which was then cloned into a pmirGLO vector
(Promega Corporation, Madison, WI, USA). The PCR method used is
detailed in the previous subsection. Subsequently, Lipofectamine
2000® was utilized to transfect cells in the 24-well
plates. The obtained wild type or mutant pmirGLO vector, pRL-SV40
Renilla luciferase construct (5 ng; Promega Corporation) and
miR-30a mimic or the respective negative control were
co-transfected to each well. After 48 h transfection, cells were
extracted and the luciferase activity was determined using the Dual
luciferase reporter assay system (DLR® Assay) following
48 h. Firefly luciferase activity in the vectors was normalized to
Renilla luciferase activity.
Statistical analysis
Data were analyzed using GraphPad Prism 6.0 software
(GraphPad Software, Inc., La Jolla, CA, USA) and presented as the
mean ± standard deviation. One-way analysis of variance followed by
a Tukey's post-hoc test was used for comparisons between multiple
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
miR-30a is downregulated in human PTC
tissue
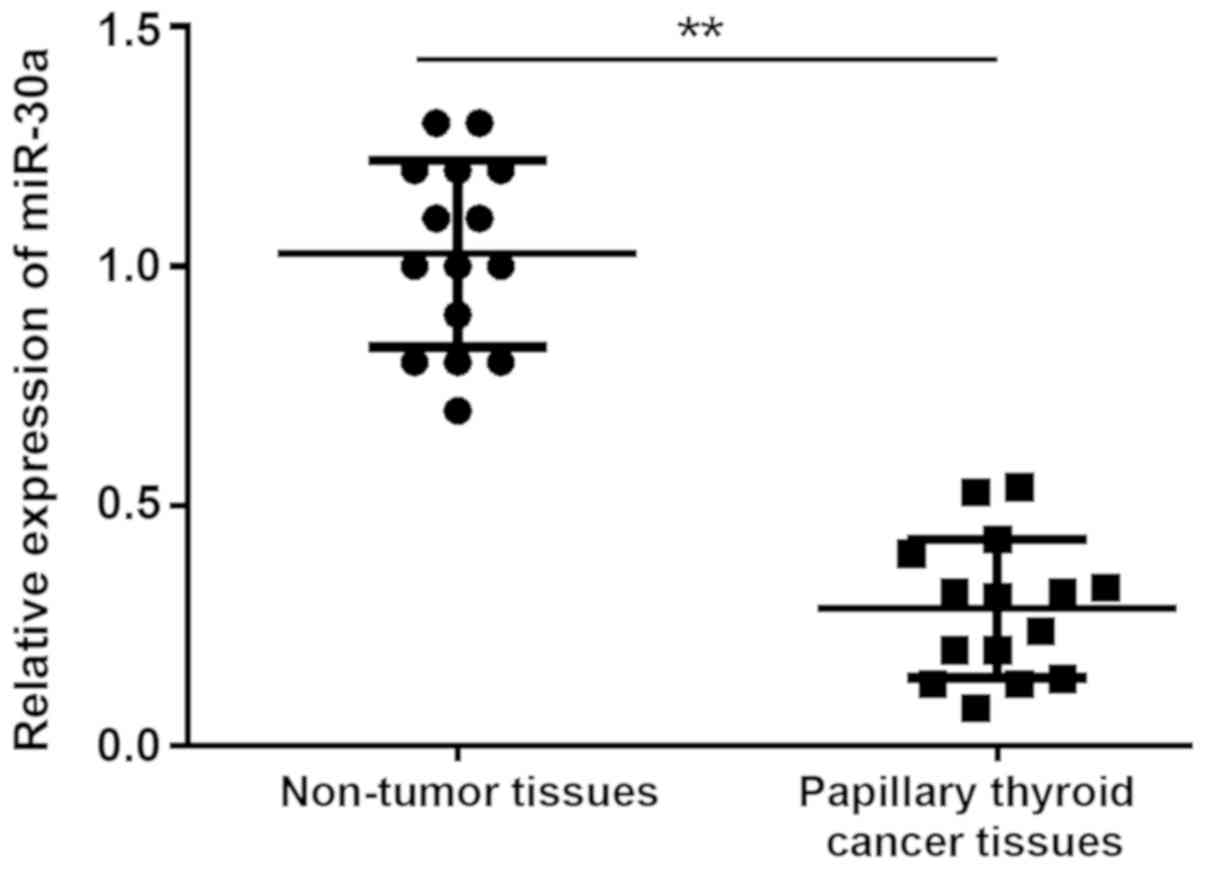
To obtain a greater understanding of the clinical
interrelation of the expression of miR-30a in PTC tissue, 15 pairs
of PTC tissue samples and non-tumor tissues were assessed via
RT-qPCR. The results revealed that the expression of miR-30a was
significantly downregulated in PTC tissue compared with non-tumor
tissue (Fig. 1).
miR-30a is downregulated and E2F7 is
upregulated in human PTC cells
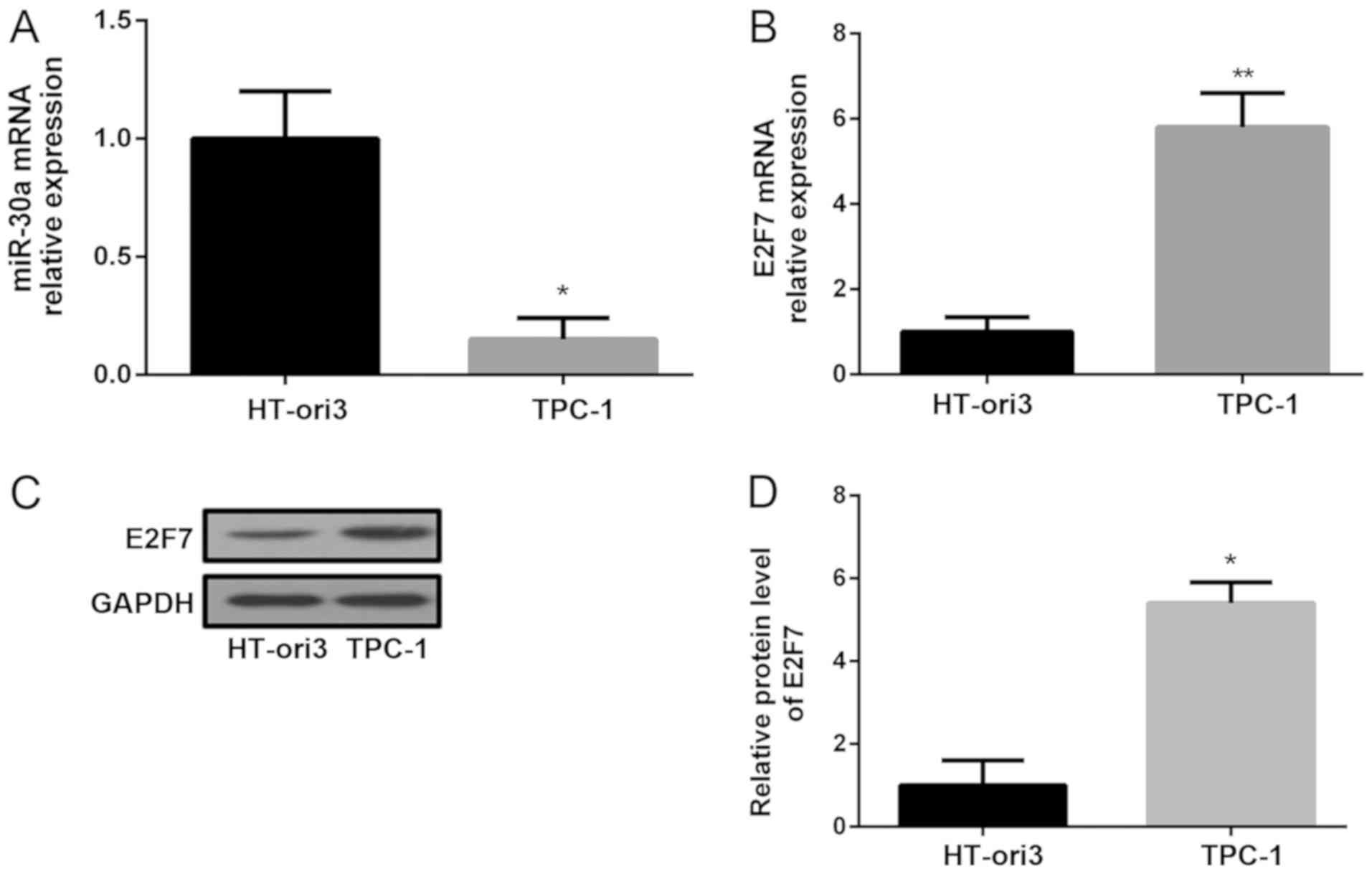
The current study aimed to determine the expression
of miR-30a in a PTC cell line (TPC-1) and a normal thyroid cell
line (HT-ori3). Compared with HT-ori3 cells, the expression of
miR-30a and the levels of E2F7 in TCP-1 cells were significantly
decreased and increased, respectively (Fig. 2A-B). Western blotting was
subsequently performed to detect the protein expression of E2F7 in
these cell lines. The results revealed that E2F7 levels in TPC-1
cells were markedly increased compared with HT-ori3 cells (Fig. 2C-D).
miR-30a decreases the proliferation of
TPC-1 cells
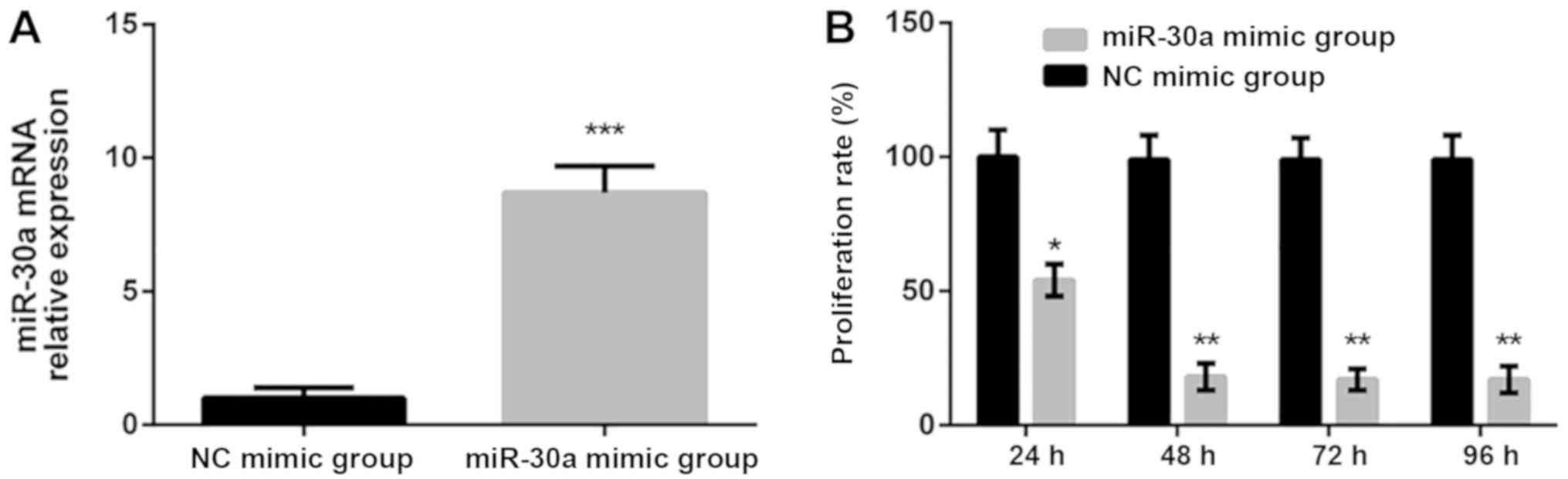
To determine whether miR-30a regulates the
proliferation of PTC cells, miR-30a mimics were utilized to
increase the expression of miR-30a. Following transfection for 6 h,
a significant increase in miR-30a expression was observed when
compared with the NC mimic group (Fig.
3A). An MTT assay was subsequently performed to determine the
proliferation of TPC-1 cells following miR-30a mimic or NC mimic
transfection. The results revealed that transfection with miR-30a
mimics significantly decreased cell proliferation compared with the
NC mimics group. Furthermore, the proliferation of cells in the
miR-30a mimic group achieved the minimal viability rate and become
stable after 48 h (Fig. 3B). These
results confirmed that the upregulation of miR-30a inhibits the
proliferation of PTC cells.
miR-30a inhibits the migration and
invaison of TPC-1 cells
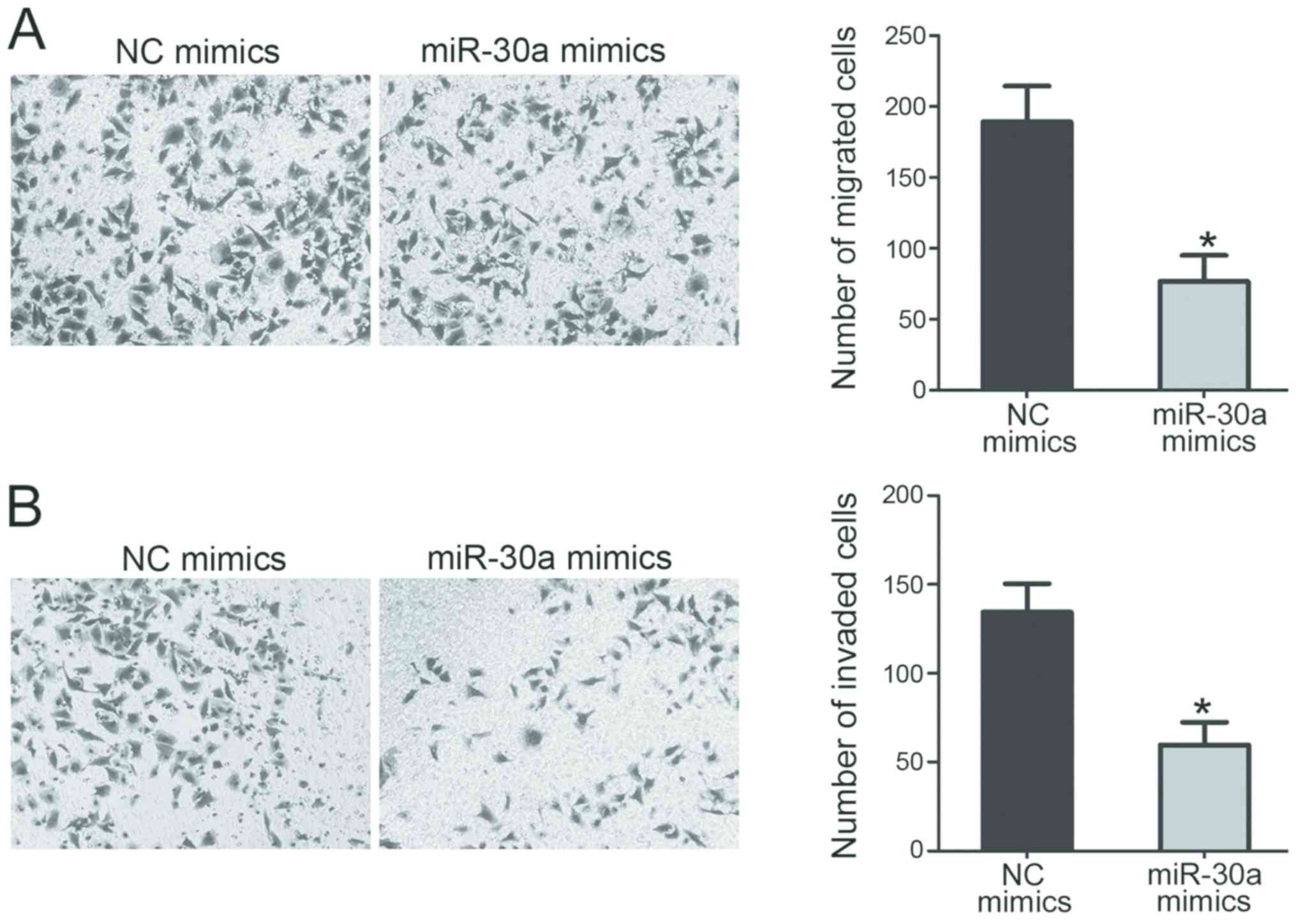
To assess the roles of miR-30a in PTC cell migration
and invasion, transwell migration and invasion assays were
performed in TPC-1 cells transfected with miR-30a or NC mimics. The
results indicated that the overexpression of miR-30a, induced via
the transfection of a miR-30a mimic, significantly suppressed the
migration (Fig. 4A) and invasion
(Fig. 4B) of TPC-1 cells compared
with the NC mimics group.
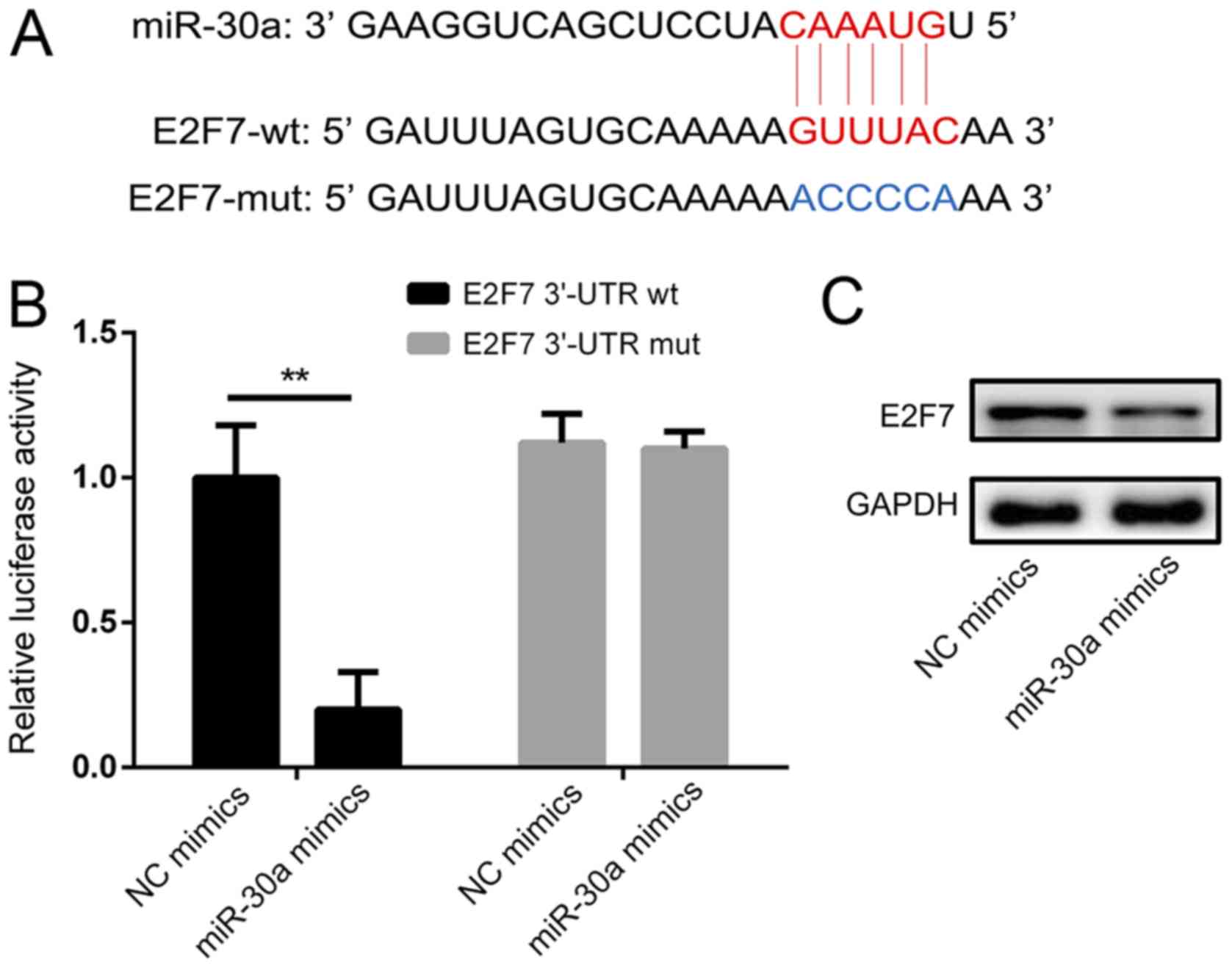
miR-30a targets the E2F7 3′-UTR
directly
To identify the molecular mechanism of miR-30a in
the regulation of cell survival, putative miR-30a targets were
predicted via bioinformatics analysis (Fig. 5A). The results revealed that E2F7 is
a direct target of miR-30a. A luciferase reporter assay was then
performed to confirm whether miR-30a targets E2F7 directly
(Fig. 5B). The results demonstrated
that, compared with the NC mimics group, co-transfection with
miR-30a mimics significantly decreased the luciferase activity of
the wild-type E2F7-3′-UTR luciferase vector in TPC-1 cells.
However, no significant effect on the luciferase activity of mutant
E2F7-3′-UTR following transfection of miR-30a was observed.
Furthermore, the protein levels of E2F7 were markedly decreased
following miR-30a transfection (Fig.
5C). These data indicate that E2F7 may be a direct target of
miR-30a in PTC cells.
Discussion
TC is a common endocrine malignancy that is
primarily derived from follicular thyroid or parafollicular C cells
(22). The overall 10-year survival
rate of differentiated TC was >80%, with ~5–20% of TC patients
developing local or regional recurrence and 10–15% developing
distant metastases in a 2014 study (23). Furthermore, PTC accounts for ~80–90%
of all TC patients. In recent years, previous studies have aimed to
assess certain miRNAs as possible regulators of tumorigenesis and
PTC development (24–26). The current study aimed to assess the
role of miR-30a in PTC via the regulation of E2F7. The results
demonstrated that the overexpression of miR-30a suppressed the
proliferation, migration and invasion of PTC cells by directly
targeting E2F7.
Previous studies have demonstrated that miRNAs serve
as tumor suppressors in various types of cancer (27–30). A
previous study revealed that miR-141 suppressed TC cell growth and
metastasis by suppressing insulin receptor substrate 2, indicating
that miR-141 may serve as a potential therapeutic target for the
treatment of patients with TC (31).
Furthermore, miR-497 is also considered to be a TC tumor suppressor
that acts by repressing brain derived neurotrophic factor (32). A previous study also indicated that
miR-7 may function as a tumor suppressor by directly targeting
serine/threonine protein kinase 1, potentially serving as a novel
therapeutic target for TC (30).
Therefore, miRNAs have been demonstrated to be associated with
tumor development and progression, highlighting their potential use
as biomarkers for the diagnosis and prognosis of PTC. However, the
prognostic relevance and functions of miR-30a in PTC remain largely
unclear. The current study assessed the expression of miR-30a and
their role in PTC cells. The results indicated that miR-30a was
downregulated in PTC cell lines compared with normal cell lines.
Furthermore, the ectopic expression of miR-30a inhibited PTC cell
proliferation, migration and invasion.
It is well known that E2F7 serves predominantly as a
transcriptional repressor, binding to miRNA promoters and
protein-coding genes bearing E2F consensus motifs, thereby
inhibiting their expression (33).
The E2F family of transcription factors are important regulators of
cellular proliferation (34). A
previous study has revealed that the E2F7 protein serves an
essential role in the regulation of cell cycle progression and may
be a key component of a negative feedback loop required to turn off
the transcription of E2F-driven G1/S target genes, thus allowing
progression through the cell cycle (35). The p53-dependent transcriptional
upregulation of E2F7 results in the repression of relevant gene
expression and the E2F7-dependent mechanism contributes to
p53-dependent cell cycle arrest in response to DNA damage (36). Furthermore, E2F7 may serve as an
independent prognostic factor of glioma and may therefore
constitute a potential therapeutic target for this disease
(37). A recent study demonstrated
that the inhibition of E2F7 inhibits gallbladder cancer cell
proliferation, migration and metastasis (38). In the current study, miR-30a was
confirmed to target E2F7 directly and the increased expression of
miR-30a significantly suppressed E2F7 expression in human PTC
cells. Furthermore, the results of transwell migration and invasion
assays indicated that the overexpression of miR-30a, induced via
transfection with miR-30a mimics, significantly suppressed the
migration and invasion of PTC cells. In future experiments, studies
will aim to perform other functional assays to determine the effect
of miR-30a on the proliferation and metastasis of PTC cells in
vitro and in vivo, including cell cycle analysis by flow cytometry
and wound healing assay. Collectively, the current study revealed
that miR-30a inhibited cell proliferation, migration and invasion,
partially by targeting E2F7. Therefore, the current study provided
evidence for the critical regulatory axis of miR-30a/E2F7 in the
developmental process of PTC.
Overall, the current study demonstrated that the
proliferation, migration and invasion of PTC cells are inhibited by
miR-30a overexpression by targeting E2F7. All results obtained in
the current study indicate that miR-30a may serve as a tumor
suppressor gene in the tumorigenesis and progression of PTC.
Therefore, miR-30a and E2F7 may serve as potential therapeutic
targets for the treatment of patients with PTC.
Acknowledgements
Not applicable.
Funding
No funding received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
GY conceived the current study, wrote the paper and
created the figures. GY and ZY performed the experiments, searched
the literature, analyzed the data, created the tables and reviewed
the manuscript.
Ethics approval and consent to
participate
All tissue samples were obtained following the
receipt of written informed consent. The current study was approved
by the Ethics Review Board of Fenyang College Shanxi Medical
University (Shanxi, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang J, Yang H, Si Y, Hu D, Yu Y, Zhang Y,
Gao M and Zhang H: Iodine promotes tumorigenesis of thyroid cancer
by suppressing Mir-422a and up-regulating MAPK1. Cell Physiol
Biochem. 43:1325–1336. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kaptan E, Sancar-Bas S, Sancakli A, Bektas
S and Bolkent S: The effect of plant lectins on the survival and
malignant behaviors of thyroid cancer cells. J Cell Biochem.
119:6274–6287. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sokić SI, Adanja BJ, Vlajinac HD, Janković
RR, Marinković JP and Zivaljević VR: Risk factors for thyroid
cancer. Neoplasma. 41:371–374. 1994.PubMed/NCBI
|
|
4
|
Takacsova E, Kralik R, Waczulikova I,
Zavodna K and Kausitz J: A different prognostic value of BRAFV600E
mutation positivity in various age groups of patients with
papillary thyroid cancer. Neoplasma. 64:156–164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Di W, Li Q, Shen W, Guo H and Zhao S: The
long non-coding RNA HOTAIR promotes thyroid cancer cell growth,
invasion and migration through the miR-1-CCND2 axis. Am J Cancer
Res. 7:1298–1309. 2017.PubMed/NCBI
|
|
6
|
Xing M, Haugen BR and Schlumberger M:
Progress in molecular-based management of differentiated thyroid
cancer. Lancet. 381:1058–1069. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ding ZY, Huang YJ, Tang JD, Li G, Jiang PQ
and Wu HT: Silencing of hypoxia-inducible factor-1α promotes
thyroid cancer cell apoptosis and inhibits invasion by
downregulating WWP2, WWP9, VEGF and VEGFR2. Exp Ther Med.
12:3735–3741. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lim ST, Jeon YW and Suh YJ: The prognostic
values of preoperative tumor volume and tumor diameter in T1N0
papillary thyroid cancer. Cancer Res Treat. 49:890–897. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rachinsky I, Rajaraman M, Leslie WD,
Zahedi A, Jefford C, McGibbon A, Young JE, Pathak KA, Badreddine M,
De Brabandere S, et al: Regional variation across Canadian centers
in radioiodine administration for thyroid remnant ablation in
well-differentiated thyroid cancer diagnosed in 2000–2010. J
Thyroid Res. 2016:28679162016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guan H, Liang W, Xie Z, Li H, Liu J, Liu
L, Xiu L and Li Y: Down-regulation of miR-144 promotes thyroid
cancer cell invasion by targeting ZEB1 and ZEB2. Endocrine.
48:566–574. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vasko VV and Saji M: Molecular mechanisms
involved in differentiated thyroid cancer invasion and metastasis.
Curr Opin Oncol. 19:11–17. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liao T, Wang YJ, Hu JQ, Wang Y, Han LT, Ma
B, Shi RL, Qu N, Wei WJ, Guan Q, et al: Histone methyltransferase
KMT5A gene modulates oncogenesis and lipid metabolism of papillary
thyroid cancer in vitro. Oncol Rep. 39:2185–2192.
2018.PubMed/NCBI
|
|
13
|
Lu ZL, Chen YJ, Jing XY, Wang NN, Zhang T
and Hu CJ: Detection and identification of serum peptides biomarker
in papillary thyroid cancer. Med Sci Monit. 24:1581–1587. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao W, Chen S, Hou X, Chen G and Zhao Y:
CHK2 promotes anoikis and is associated with the progression of
papillary thyroid cancer. Cell Physiol Biochem. 45:1590–1602. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shi Z, Zhou H, Lu L, Li X, Fu Z, Liu J,
Kang Y, Wei Z, Pan B, Liu L, et al: The roles of microRNAs in
spinal cord injury. Int J Neurosci. 127:1104–1115. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ouyang Q, Xu L, Cui H, Xu M and Yi L:
MicroRNAs and cell cycle of malignant glioma. Int J Neurosci.
126:1–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen X, Ba Y, Ma L, Cai X, Yin Y, Wang K,
Guo J, Zhang Y, Chen J, Guo X, et al: Characterization of microRNAs
in serum: A novel class of biomarkers for diagnosis of cancer and
other diseases. Cell Res. 18:997–1006. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang RJ, Zheng YH, Wang P and Zhang JZ:
Serum miR-125a-5p, miR-145 and miR-146a as diagnostic biomarkers in
non-small cell lung cancer. Int J Clin Exp Pathol. 8:765–771.
2015.PubMed/NCBI
|
|
19
|
Fuziwara CS and Kimura ET: MicroRNA
deregulation in anaplastic thyroid cancer biology. Int J
Endocrinol. 2014:7434502014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Costa-Pinheiro P, Ramalho-Carvalho J,
Vieira FQ, Torres-Ferreira J, Oliveira J, Gonçalves CS, Costa BM,
Henrique R and Jerónimo C: MicroRNA-375 plays a dual role in
prostate carcinogenesis. Clin Epigenetics. 7:422015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xing M: Molecular pathogenesis and
mechanisms of thyroid cancer. Nat Rev Cancer. 13:184–199. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Perros P, Boelaert K, Colley S, Evans C,
Evans RM, Gerrard Ba G, Gilbert J, Harrison B, Johnson SJ, Giles
TE, et al: Guidelines for the management of thyroid cancer. Clin
Endocrinol (Oxf). 81 (Suppl 1):S1–S122. 2014. View Article : Google Scholar
|
|
24
|
Bi CL, Zhang YQ, Li B, Guo M and Fu YL:
MicroRNA-520a-3p suppresses epithelial-mesenchymal transition,
invasion, and migration of papillary thyroid carcinoma cells via
the JAK1-mediated JAK/STAT signaling pathway. J Cell Physiol.
234:4054–4067. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu H, Guo J, Chai H and Meng X:
MicroRNA-744 suppresses cell proliferation and invasion of
papillary thyroid cancer by directly targeting NOB1. Mol Med Rep.
19:1903–1910. 2019.PubMed/NCBI
|
|
26
|
Heidari Z, Mohammadpour-Gharehbagh A,
Eskandari M, Harati-Sadegh M and Salimi S: Genetic polymorphisms of
miRNA let7a-2 and pri-mir-34b/c are associated with an increased
risk of papillary thyroid carcinoma and clinical/pathological
features. J Cell Biochem. 14–Dec;2018.(Epub ahead of print).
|
|
27
|
Damodaran M, Paul SFD and Venkatesan V:
Genetic polymorphisms in miR-146a, miR-196a2 and miR-125a genes and
its association in prostate cancer. Pathol Oncol Res.
29–Mar;2018.(Epub ahead of print). View Article : Google Scholar
|
|
28
|
Mo JS, Park YR and Chae SC: MicroRNA 196B
regulates HOXA5, HOXB6 and GLTP expression levels in colorectal
cancer cells. Pathol Oncol Res. 12–Mar;2018.(Epub ahead of print).
View Article : Google Scholar
|
|
29
|
Seclaman E, Narita D, Anghel A, Cireap N,
Ilina R, Sirbu IO and Marian C: MicroRNA expression in laser
micro-dissected breast cancer tissue samples-a pilot study. Pathol
Oncol Res. 25:233–239. 2019. View Article : Google Scholar
|
|
30
|
Yue K, Wang X, Wu Y, Zhou X, He Q and Duan
Y: microRNA-7 regulates cell growth, migration and invasion via
direct targeting of PAK1 in thyroid cancer. Mol Med Rep.
14:2127–2134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dong S, Meng X, Xue S, Yan Z, Ren P and
Liu J: microRNA-141 inhibits thyroid cancer cell growth and
metastasis by targeting insulin receptor substrate 2. Am J Transl
Res. 8:1471–1481. 2016.PubMed/NCBI
|
|
32
|
Wang P, Meng X, Huang Y, Lv Z, Liu J, Wang
G, Meng W, Xue S, Zhang Q, Zhang P and Chen G: MicroRNA-497
inhibits thyroid cancer tumor growth and invasion by suppressing
BDNF. Oncotarget. 8:2825–2834. 2017.PubMed/NCBI
|
|
33
|
Westendorp B, Mokry M, Groot Koerkamp MJ,
Holstege FC, Cuppen E and de Bruin A: E2F7 represses a network of
oscillating cell cycle genes to control S-phase progression.
Nucleic Acids Res. 40:3511–3523. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen D, Pacal M, Wenzel P, Knoepfler PS,
Leone G and Bremner R: Division and apoptosis of E2f-deficient
retinal progenitors. Nature. 462:925–929. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mitxelena J, Apraiz A, Vallejo-Rodríguez
J, García-Santisteban I, Fullaondo A, Alvarez-Fernández M,
Malumbres M and Zubiaga AM: An E2F7-dependent transcriptional
program modulates DNA damage repair and genomic stability. Nucleic
Acids Res. 46:4546–4559. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Carvajal LA, Hamard PJ, Tonnessen C and
Manfredi JJ: E2F7, a novel target, is up-regulated by p53 and
mediates DNA damage-dependent transcriptional repression. Genes
Dev. 26:1533–1545. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yin W, Wang B, Ding M, Huo Y, Hu H, Cai R,
Zhou T, Gao Z, Wang Z and Chen D: Elevated E2F7 expression predicts
poor prognosis in human patients with gliomas. J Clin Neurosci.
33:187–193. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ye YY, Mei JW, Xiang SS, Li HF, Ma Q, Song
XL, Wang Z, Zhang YC, Liu YC, Jin YP, et al: MicroRNA-30a-5p
inhibits gallbladder cancer cell proliferation, migration and
metastasis by targeting E2F7. Cell Death Dis. 9:4102018. View Article : Google Scholar : PubMed/NCBI
|