Introduction
Glioma is one of the most common types of malignant
brain tumour, with a morbidity rate of 5/10,000 each year (1,2). During
glioma progression, a series of molecular events result in the
expression of genes associated with tumour cell proliferation and
motility (3,4). Investigating the underlying molecular
mechanisms in glioma cell growth and progression by gene expression
profiling may facilitate the identification of potentially novel
biomarkers and therapeutic targets.
MicroRNAs (miRs), which are small endogenous
noncoding RNAs comprising 22–25 nucleotides, regulate gene
expression at the transcriptional and/or post-transcriptional level
by binding to the 3′untranslated region (UTR) of their target mRNA
(5,6). By repressing protein translation or
promoting mRNA degradation, miRs play important roles in multiple
cellular processes, including cell proliferation, survival,
apoptosis, differentiation and motility (6–8). Several
studies have demonstrated that miRs are deregulated in glioma, and
some miRs have been found to play promoting or suppressive roles in
different types of cancer (9,10). For
instance, miR-365 is downregulated in glioma and inhibits glioma
cell proliferation, migration and invasion via targeting
phosphoinositide-3-kinase (PI3K) regulatory subunit 3 (9), while miR-93 is upregulated in glioma
and promotes glioma cell proliferation via activation of the
PI3K/Akt signalling pathway (11).
Among the glioma-related miRs, miR-25 is upregulated in several
types of human cancer and often serves an oncogenic role (12,13). For
instance, miR-25 promotes the proliferation of triple-negative
breast cancer by targeting BTG anti-proliferation factor 2
(12). Furthermore, exosome-derived
miR-25 stimulates the malignant progression of liposarcoma
(13). Zhang et al (14) reported that miR-25 promoted glioma
cell proliferation by targeting cyclin dependent kinase inhibitor
1C, while Peng et al (15)
demonstrated that miR-25 promoted glioblastoma cell proliferation
and invasion by directly targeting neurofilament light. However,
the clinical significance of miR-25 expression in glioma as well as
the underlying molecular mechanism of miR-25 in glioma cell
proliferation and migration remains unknown.
F-box and WD repeat domain containing 7 (FBXW7), a
substrate adaptor for an E3 Skp1-Cul1-F-box ubiquitin ligase
complex, negatively regulates the abundance of several oncoproteins
(16). Studies have demonstrated
that miRs may serve promoting roles in several types of human
cancer via targeting FBXW7 (17,18). For
instance, miR-92a is upregulated in cervical cancer and promotes
cell proliferation and invasion by targeting FBXW7 (17). In addition, FBW7 inhibits malignancy
and enhances temozolomide sensitivity in glioblastoma cells
(18). However, the relationship
between miR-25 and FBXW7 in glioma has not previously been
identified.
Dickkopf Wnt signaling pathway inhibitor 3 (DKK3), a
member of the Dickkopf family, interacts with and suppresses the
Wnt signalling pathway and tumourigenesis (19,20).
DKK3 is downregulated in several types of human cancer and is a
tumour suppressor (20). A previous
study demonstrated that DKK3 induced glioma cell death (21). However, the relationship between
miR-25 and DKK3 in glioma has not previously been examined.
In the present study, upregulation of miR-25
expression in glioma was associated with poor survival in patients
with glioma. In addition, FBXW7 and DKK3 were identified as
potential targets of miR-25 in glioma cells. Furthermore, miR-25
promoted glioma cell survival, proliferation and migration via
targeting FBXW7 and DKK3.
Materials and methods
Tissue samples
A total of 60 primary glioma tissue and 10 normal
brain tissue samples were collected from patients undergoing
surgical resection at the Department of Neurosurgery of Xiangya
Hospital (Changsha, China) between March 2010 and May 2014. The
glioma tissue was obtained from 60 patients with glioma (female,
n=25; male, n=35; age range, 32–66 years; mean age, 55.1 years),
and normal tissue adjacent to the tumour was obtained from 10
patients (female, n=4; male, n=6; age range, 40–65 years; mean age,
53.6 years). None of the patients had received chemotherapy or
radiotherapy prior to surgical resection. The WHO stage was
determined by pathologists (22).
All of the tissue samples collected were immediately snap-frozen in
liquid nitrogen and stored at −80°C. According to the Helsinki
Declaration, all participants or their families were well informed
of the study details and written informed consent was obtained
prior to the study. The current study was approved by the Ethics
Committee of Xiangya Hospital of Central South University
(Changsha, China).
Cell culture
Normal human astrocytes (NHAs) were obtained from
Lonza Group Ltd. The human glioma cell lines U-373MG Uppsala,
U-87MG Uppsala, U251 and T98G were obtained from the Cell Bank of
Type Culture Collection of Chinese Academy of Sciences (Shanghai,
China). Cells were cultured in Dulbecco's modified Eagle's medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10%
FBS (Thermo Fisher Scientific, Inc.), 1% penicillin and
streptomycin (Thermo Fisher Scientific, Inc.), and 1% glutamine
(Thermo Fisher Scientific, Inc.) at 37°C in a humidified atmosphere
containing 5% CO2.
Cell transfection
U251 and T98G cells in the logarithmic phase were
seeded at a density of 5×105 cells/well in a six-well
plate for 24 h. The cells were then transfected with 100 nM
negative control (NC) inhibitor (cat. no. 4464076; Thermo Fisher
Scientific, Inc.), 100 nM miR-25 inhibitor (cat. no. 4464084;
Thermo Fisher Scientific, Inc.), 100 nM miR-NC (cat. no. 4464058;
Thermo Fisher Scientific, Inc.) or 100 nM miR-25 mimics (cat. no.
4464066; Thermo Fisher Scientific, Inc.); or co-transfected with
100 nM miR-25 inhibitor and 100 nM NC siRNA (cat. no. 4390843;
Thermo Fisher Scientific, Inc.; group termed miR-25 in + siNC), 100
nM miR-25 inhibitor and 100 nM FBXW7 siRNA (cat. no. HSS124318;
Thermo Fisher Scientific, Inc.; group termed miR-25 in + siFBXW7)
or 100 nM miR-25 inhibitor and 100 nM DKK3 siRNA (cat. no.
HSS146899; Thermo Fisher Scientific, Inc.; group termed miR-25 in +
siDKK3) using Lipofectamine® 2000 (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Following transfection for 48 h, transfected cells were collected
and used in subsequent experimentation.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from tissue samples or cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Total RNA was subsequently reverse transcribed
into cDNA using the miScript Reverse Transcription kit (Qiagen,
Inc.), according to the manufacturer's protocol using the following
conditions: 37°C for 60 min and 95°C for 5 min, followed by storage
at 4°C. qPCR was subsequently performed using the miScript
SYBR® Green PCR kit (Qiagen, Inc.) on a Roche
LightCycler 480 Real-Time PCR system (Roche Diagnostics). The
following thermocycling conditions were used for qPCR: Initial
denaturation at 95°C for 1 min; 40 cycles of 95°C for 15 sec, 55°C
for 30 sec and 72°C for 30 sec. The mRNA expression levels were
quantified using the 2−ΔΔCq method (23). The following primer pairs were used
for the qPCR: miR-25 forward, 5′-CATTGCACTTGTCTCGGTCTGA-3′ and
reverse, 5′-GCTGTCAACGATACGCTACGTAACG-3′; U6 forward,
5′-CTCGCTTCGGCAGCACA-3′, and reverse, 5′-AACGCTTCACGAATTTGCGT-3′;
DKK3 forward, 5′-AGGACACGCAGCACAAATTG-3′ and reverse,
5′-CCAGTCTGGTTGTTGGTTATCTT-3′; FBXW7 forward,
5′-GGCCAAAATGATTCCCAGCAA-3′ and reverse,
5′-ACTGGAGTTCGTGACACTGTTA-3′; and GAPDH forward,
5′-CTGGGCTACACTGAGCACC-3′ and reverse,
5′-AAGTGGTCGTTGAGGGCAATG-3′.
Cell Counting kit-8 (CCK-8) assay
CCK-8 assays were performed to assess cell
proliferation. Transfected cells were seeded into 96-well plates at
a density of 5×103 cells/well. Following incubation at
37°C for 0, 24, 48 and 72 h, 10 µl CCK-8 reagent (Beyotime
Institute of Biotechnology) was add to each well. Cells were
further incubated 37°C for 30 min and the absorbance was measured
at a wavelength of 490 nM using a microplate reader.
Wound-healing assay
Cell migration was examined using wound-healing
assays. Transfected U251 and T98G cells were seeded into 12-well
plates at a density of 5×105 cells/well. Following
incubation at 37°C for 24 h, cells were scratched with a sterile
200-µl pipette tip and PBS was used to wash away any detached
cells. Cell migration was observed following culture at 37°C for 0
and 24 h, and images were captured under a light microscope using a
digital camera system (magnification, ×200 or ×40).
Dual-luciferase reporter assay
TargetScan software (version 7.1; www.targetscan.org) was used to predict target genes
of miR-25, identifying FBXW7 and DKK3. The 3′UTR of human FBW7
containing the wild type (WT) or mutant (MT) putative miR-25
targeted sequence was cloned into the pMIR-REPORT™ miRNA Expression
Reporter Vector system (Ambion; Thermo Fisher Scientific, Inc.).
Subsequently, the luciferase reporter plasmid containing either
WT-FBXW7 3′UTR or mutant MT-FBXW7 3′UTR was co-transfected with
miR-25 mimics or miR-NC mimics into 293T cells (Cell Bank of Type
Culture Collection of Chinese Academy of Sciences) using
Lipofectamine® 2000, according to the manufacturer's
protocol. Similarly, the WT and MT 3′UTR of human DKK3 containing
the putative miR-25 targeted sequence was cloned into the
pMIR-REPORT™ miRNA Expression Reporter Vector system. Subsequently,
the luciferase reporter plasmid containing either WT-DKK3 3′UTR or
MT-DKK3 3′UTR was co-transfected with miR-25 mimics or miR-NC
mimics into 293T cells using Lipofectamine® 2000,
according to the manufacturer's protocol. After transfection for 48
h, cultured cells were collected and reporter activities were
detected using the Dual-Luciferase Reporter assay system (Promega
Corporation), according to the manufacturer's protocol. The
activity of firefly luciferase was normalized to the activity of
Renilla luciferase.
Western blot analysis
Total protein was extracted from tissues or cells
using radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology). Cell lysates were centrifuged at 12,000 × g for 5
min at 4°C, and the supernatant was collected. Total protein was
quantified using the Pierce BCA Protein Assay kit (Thermo Fisher
Scientific, Inc.) and 50 µg protein/lane was separated via SDS-PAGE
on a 12% gel. The separated proteins were transferred onto PVDF
membranes (EMD Millipore) and blocked for 3 h at room temperature
with 5% skimmed milk in TBS containing Tween-20 (0.5% TBST). The
membranes were incubated with rabbit anti-human FBXW7 primary
antibody (1:200; ab105752; Abcam, Cambridge, MA, USA), rabbit
anti-human DKK3 primary antibody (1:500; ab187532; Abcam), rabbit
anti-human GAPDH primary antibody (1:200; ab9485; Abcam), at room
temperature for 3 h. Membranes were washed three times with TBST.
Following primary antibody incubation, the membranes were incubated
with horseradish peroxidase-conjugated goat anti-rabbit secondary
antibody (1:5,000; ab6721; Abcam) at room temperature for 40 min.
The protein bands were visualised using enhanced chemiluminescence
reagents (ECL; Thermo Fisher Scientific, Inc.). Protein expression
was quantified using ImageJ software version 1.46 (National
Institutes of Health).
Statistical analysis
Data presented as the mean ± standard deviation from
three independent experiments. All statistical analyses were
performed using GraphPad Prism software (ver. 5.0; GraphPad
Software, Inc.). Student's t-test (two-tailed) was used to examine
differences between two groups. One-way analysis of variance
followed by Tukey's post hoc test was used to examine differences
among multiple groups. Pearson's correlation analysis was used to
assess the relationship of miR-25 expression with FBXW7 or DKK3.
Chi-square test was used to assess the association between miR-25
expression and clinicopathological characteristics in patients with
glioma. P<0.05 was considered to indicate a statistically
significant difference.
Results
Upregulation of miR-25 is associated
with glioma progression
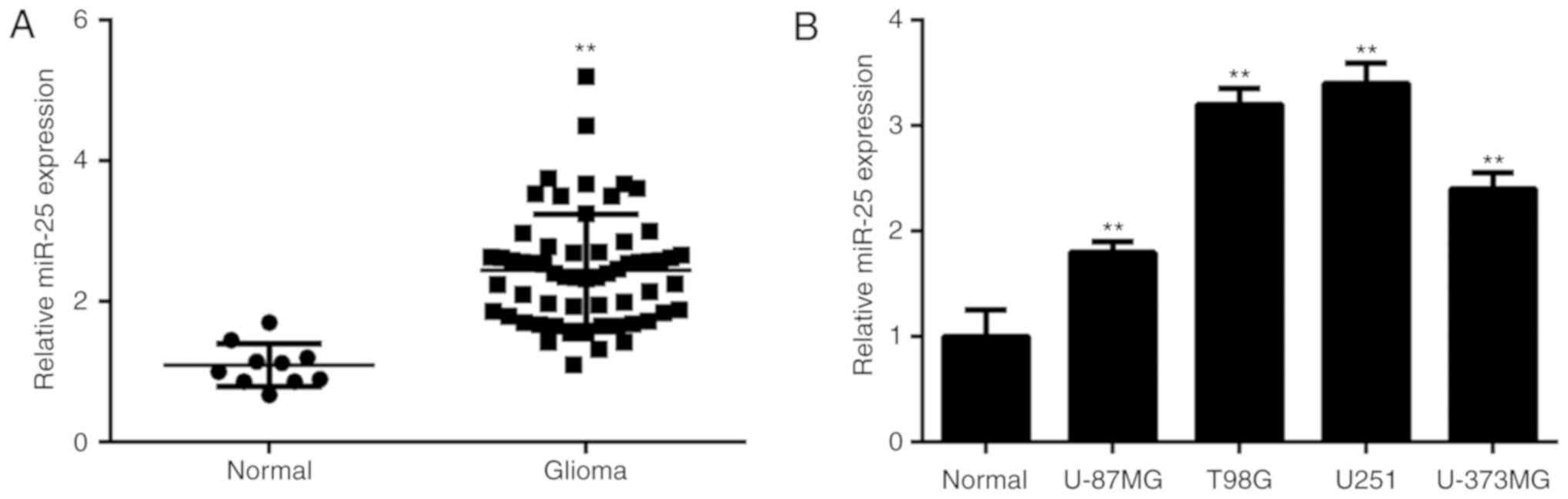
In the current study, miR-25 expression levels were
significantly higher in glioma tissue compared with normal brain
tissue (Fig. 1A). Glioma patients
were divided into high and low expression groups, based on the mean
value of miR-25 expression in the glioma tissue samples. A
significant association of high miR-25 expression with advanced
clinical stages was identified (Table
I). Furthermore, miR-25 expression was significantly
upregulated in several common glioma cell lines compared with NHAs
(Fig. 1B).
 | Table I.Association between miR-25 expression
and clinicopathological characteristics in patients with
glioma. |
Table I.
Association between miR-25 expression
and clinicopathological characteristics in patients with
glioma.
| Variables | Number (n=60) | Low miR-25
(n=31) | High miR-25
(n=29) | P-value |
|---|
| Age |
|
|
| 0.208 |
| <50 years | 32 | 14 | 18 |
|
| ≥50 years | 28 | 17 | 11 |
|
| Gender |
|
|
| 0.794 |
| Male | 35 | 19 | 16 |
|
| Female | 25 | 12 | 13 |
|
| WHO stage |
|
|
| 0.019a |
| I–II | 33 | 22 | 11 |
|
| III–IV | 27 | 9 | 18 |
|
Knockdown of miR-25 supresses glioma
cell proliferation and migration
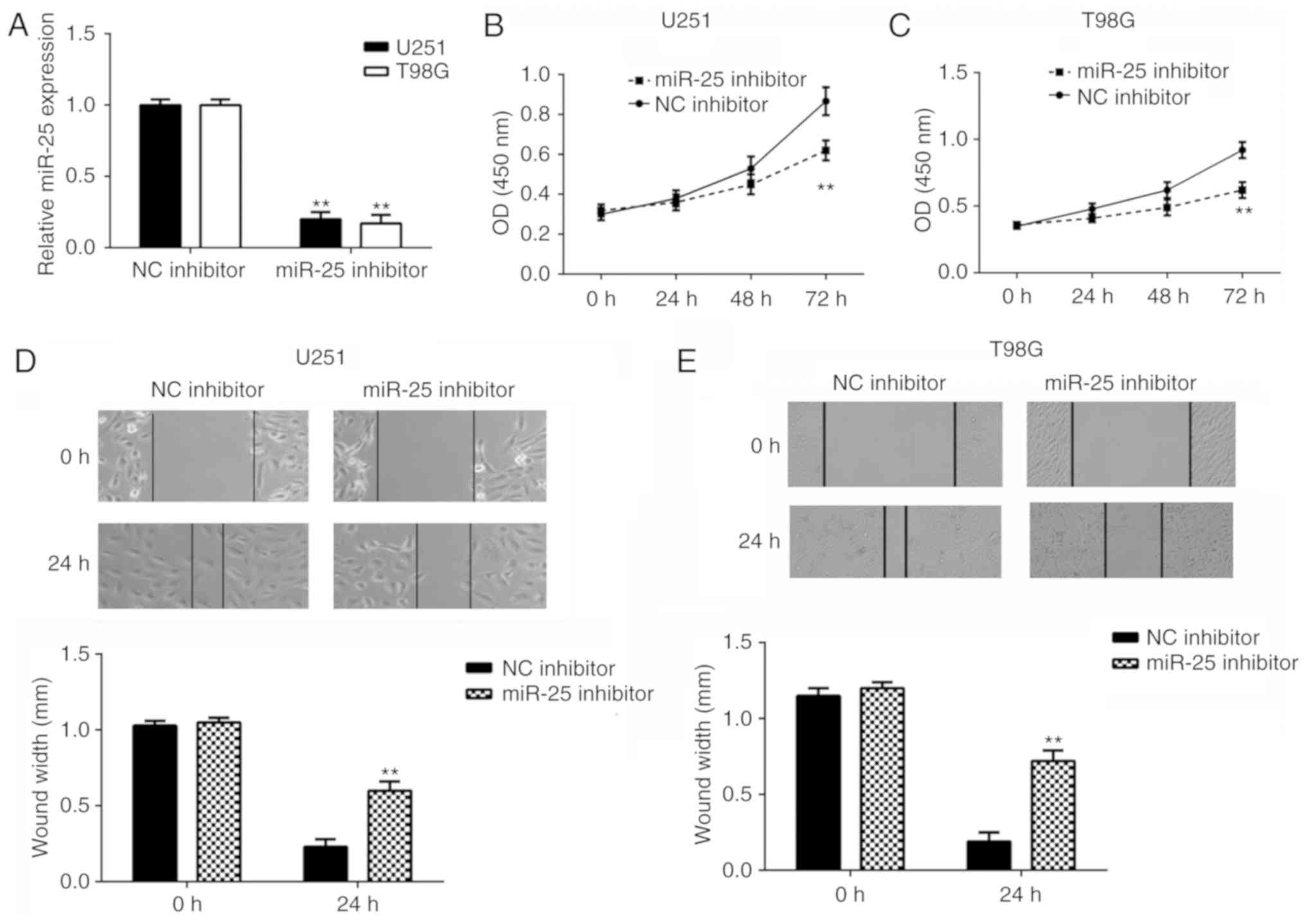
To further examine the function of miR-25 in glioma,
U251 and T98G cells were transfected with miR-25 inhibitor and NC
inhibitor. The miR-25 expression levels were significantly reduced
in cells transfected with miR-25 inhibitor compared with those
transfected with the NC inhibitor (Fig.
2A). In addition, cell proliferation was examined using CCK8
assays, which revealed that knockdown of miR-25 significantly
inhibited U251 and T98G cell proliferation compared with the
negative control (Fig. 2B and C).
Furthermore, cell migration was examined using wound-healing
assays, which revealed that knockdown of miR-25 significantly
inhibited U251 and T98G cell migration compared with the respective
negative control (Fig. 2D and E).
These results suggest that miR-25 knockdown inhibits glioma cell
proliferation and migration.
FBXW7 and DKK3 are target genes of
miR-25 in glioma cells
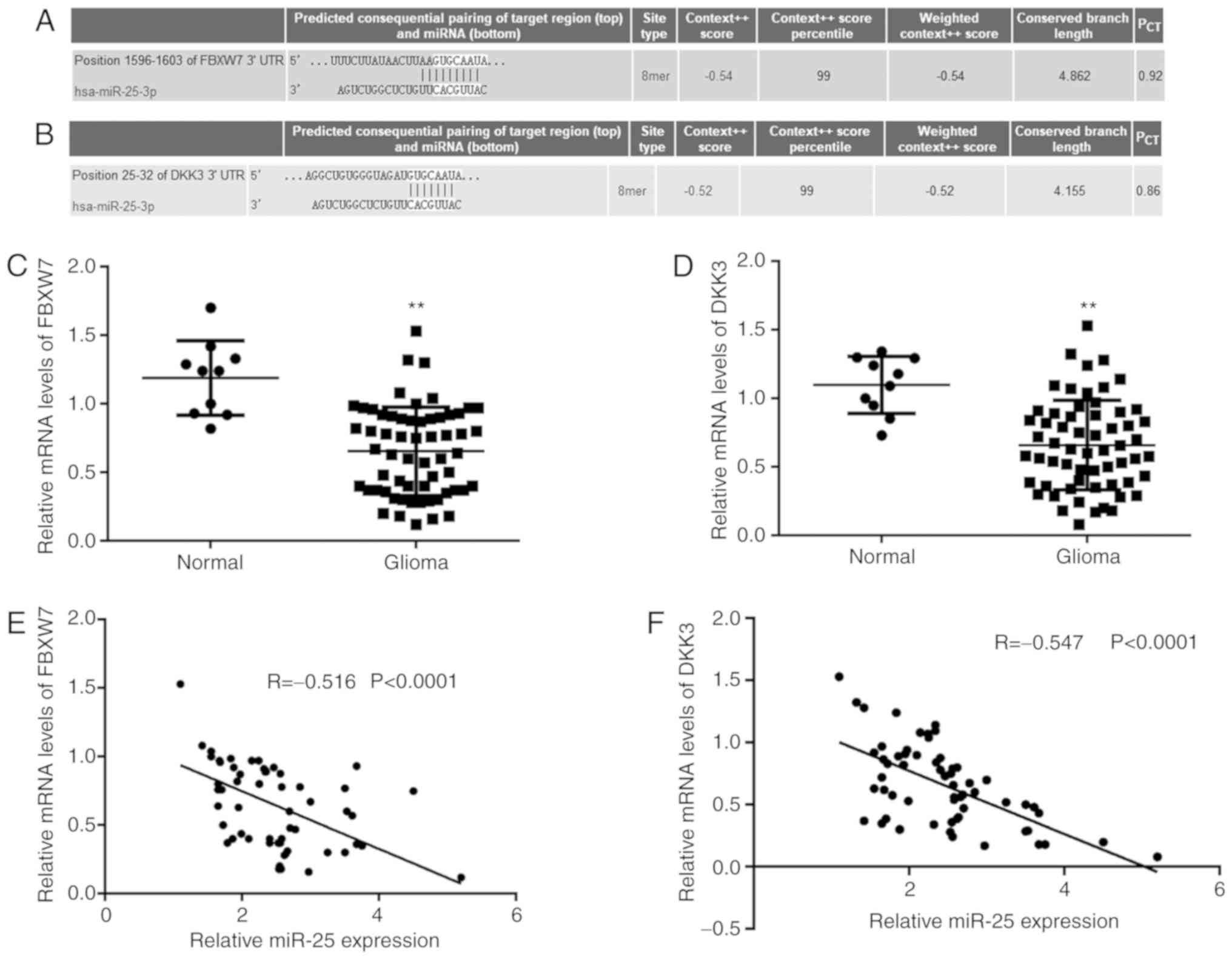
To further examine the potential underlying
regulatory mechanism of miR-25 in glioma progression, TargetScan
was used to predict potential target genes of miR-25. FBXW7 and
DKK3 were identified as potential functional target genes of miR-25
(Fig. 3A and B). The mRNA expression
levels of FBXW7 and DKK3 were significantly downregulated in glioma
tissue compared with normal brain tissue (Fig. 3C and D). Furthermore, an inverse
correlation between FBXW7 and miR-25 expression and between DKK3
and miR-25 expression was observed in glioma tissues (Fig. 3E and F).
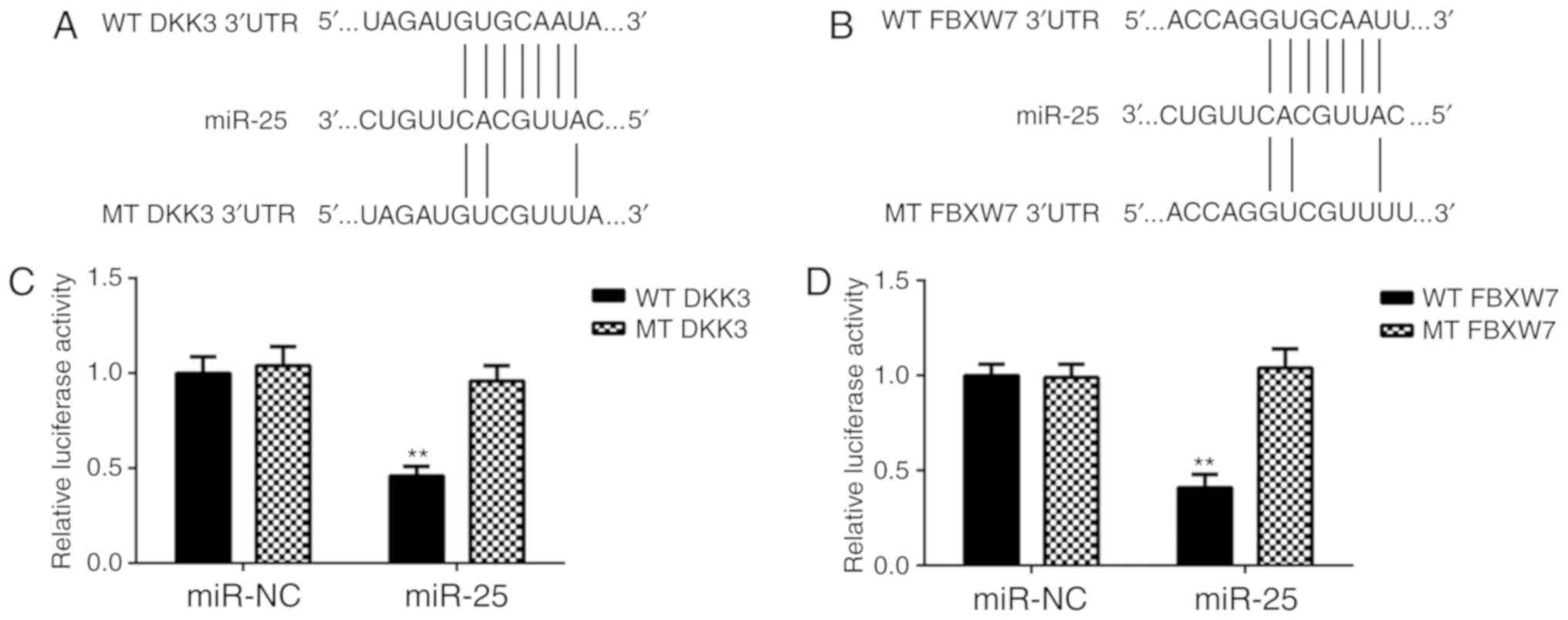
Dual-luciferase reporter assays were performed to
confirm the association between miR-25 and FBXW7, and between
miR-25 and DKK3, in glioma cells. Luciferase reporter plasmids
containing the WT-DKK3 3′UTR or MT-DKK3 3′UTR, and WT-FBXW7 3′UTR
or MT-FBXW7 3′UTR were generated (Fig.
4A and B). The results of the dual-luciferase reporter assays
demonstrated that miR-25 overexpression significantly reduced the
luciferase activity of the plasmid containing the WT-DKK3 or
WT-FBXW7 3′UTR compared with the MT-DKK3 or WT-FBXW7 3′UTR,
respectively, which did not affect the luciferase activity. These
results confirm that FBXW7 and DKK3 are target genes of miR-25 in
glioma cells.
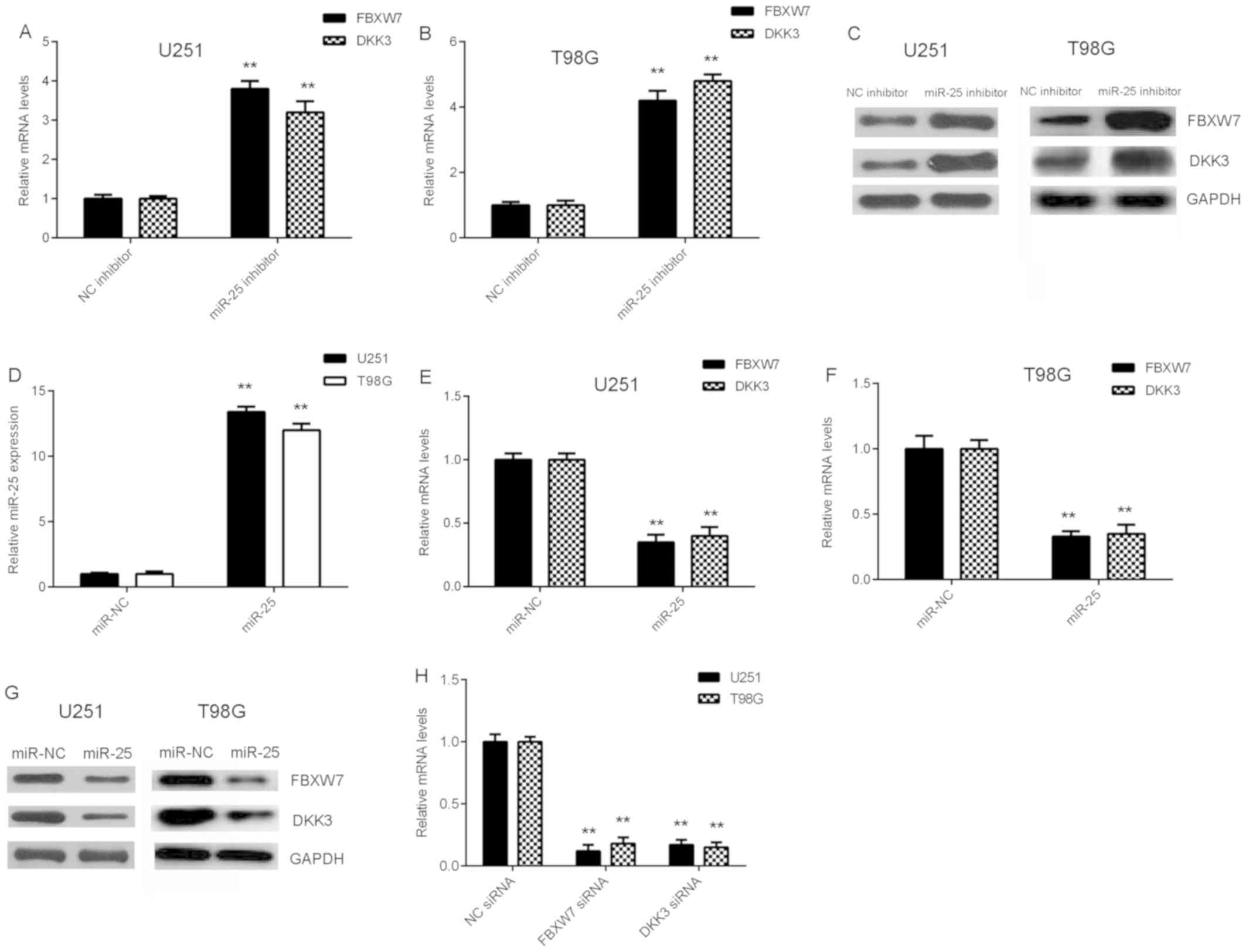
FBXW7 and DKK3 expression is
negatively regulated by miR-25 in glioma cells
The effect of miR-25 on FBXW7 and DKK3 expression
was examined in glioma cells. As shown in Fig. 5A-C, knockdown of miR-25 increased
FBXW7 and DKK3 mRNA and protein expression levels in glioma cells.
Cells with significantly increased miR-25 expression levels were
obtained by transfection with miR-25 mimics compared with negative
control (Fig. 5D). The
overexpression of miR-25 was demonstrated to decrease FBXW7 and
DKK3 expression at the mRNA and protein levels in glioma cells
(Fig. 5E-G). These results suggest
that FBXW7 and DKK3 expression is negatively regulated by miR-25 in
glioma cells.
In subsequent experiments, glioma cells were
transfected with NC siRNA, FBXW7 siRNA or DKK3 siRNA, respectively.
RT-qPCR data showed that the mRNA levels of FBXW7 and DKK3 were
significantly reduced in the FBXW7 siRNA and DKK3 siRNA groups,
when compared with those in the NC siRNA group. Thus, FBXW7 and
DKK3 expression levels were successfully downregulated in the U251
and T98G cells (Fig. 5H).
FBXW7 and DKK3 are involved in
miR-25-mediated glioma cell proliferation and migration
To investigate whether FBXW7 and DKK3 are involved
in miR-25-mediated glioma cell proliferation and migration, U251
and T98G cells were co-transfected with miR-25 inhibitor together
with FBXW7 siRNA or DKK3 siRNA. The FBXW7 mRNA and protein
expression levels were downregulated in each miR-25 in + siFBXW7
group compared with the respective miR-25 in + siNC group (Fig. 6A and B). In addition, CKK-8 and
wound-healing assays demonstrated that glioma cell proliferation
and migration were increased in the miR-25 in + siFBXW7 group
compared with the respective miR-25 in + siNC group in both cell
lines (Fig. 6C-F). Similarly, the
relative DKK3 mRNA and protein expression levels were downregulated
in the miR-25 in + siDKK3 group compared with the respective miR-25
in + siNC group in both cell lines (Fig.
7A and B). In addition, glioma cell proliferation and migration
were increased in the miR-25 in + siDKK3 group compared with the
miR-25 in + siNC group (Fig. 7C-F).
Taken together, these results suggest that FBXW7 and DKK3 may be
involved in miR-25-mediated glioma cell proliferation and
migration.
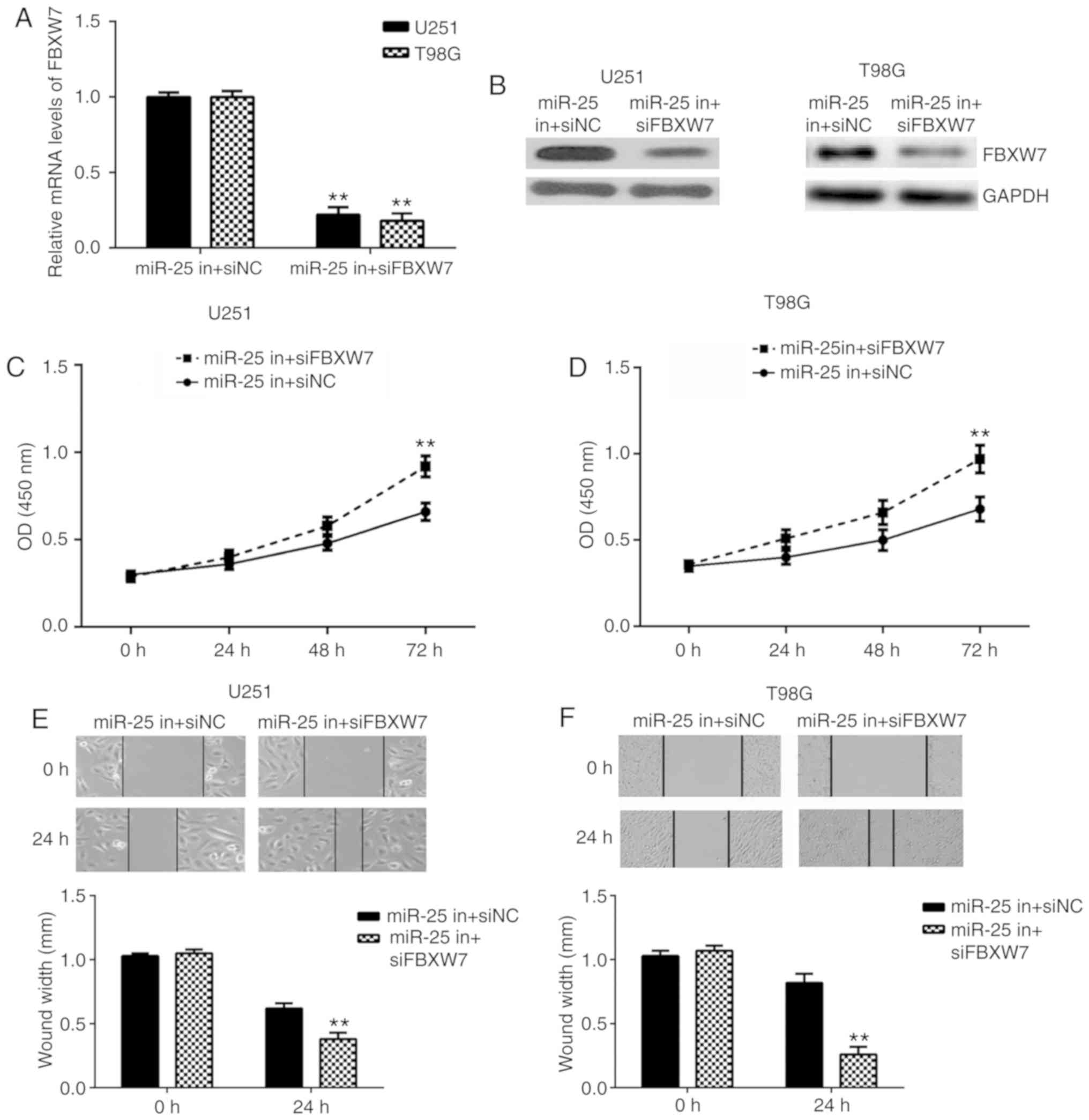 | Figure 6.Knockdown of FBXW7 reversed the
inhibitory effects of miR-25 downregulation on glioma cell
proliferation and migration. The (A) mRNA and (B) protein FBXW7
expression levels were determined by RT-qPCR and western blotting,
respectively, in U251 and T98G cells following co-transfection with
miR-25 inhibitor together with FBXW7 siRNA or NC siRNA. Cell
proliferation was examined by CCK-8 assays in (C) U251 and (D) T98G
cells following transfection with miR-25 inhibitor and FBXW7 siRNA
or NC siRNA. Cell migration was examined using wound-healing assays
in (E) U251 (magnification, ×40) and (F) T98G cells (magnification,
×200) following transfection with miR-25 inhibitor and FBXW7 siRNA
or NC siRNA. **P<0.01 vs. miR-25 in + siNC. FBXW7, F-box and WD
repeat domain containing 7; miR, microRNA; RT-qPCR, reverse
transcription-quantitative PCR; siRNA, small interfering RNA; NC,
negative control; CCK-8, Cell Counting Kit-8. |
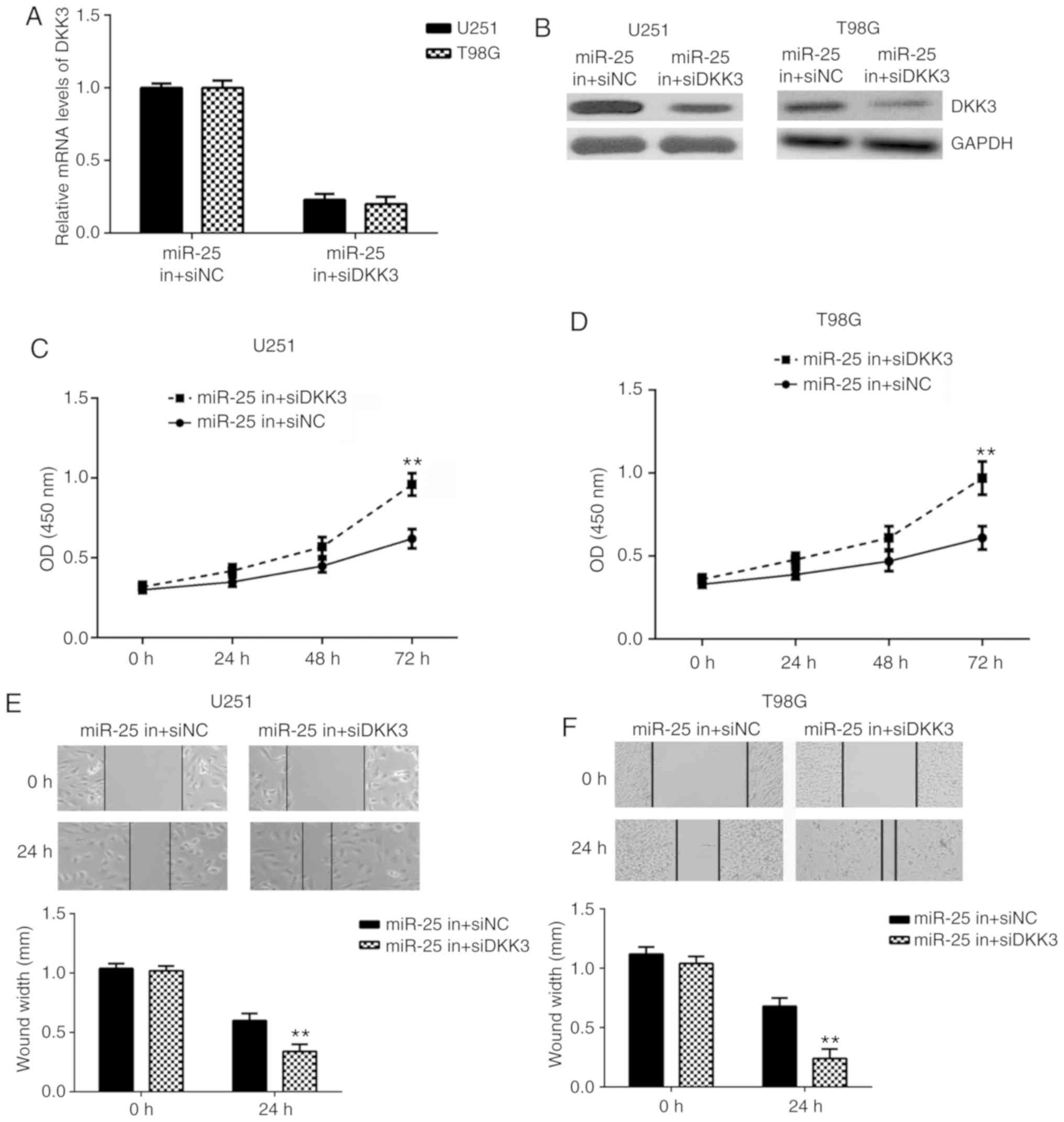 | Figure 7.DKK3 knockdown reversed the inhibitory
effects of miR-25 downregulation on glioma cell proliferation and
migration. The (A) mRNA and (B) protein expression levels of DKK3
were determined by RT-qPCR and western blotting, respectively, in
U251 and T98G cells following co-transfection with miR-25 inhibitor
together with DKK3 siRNA or NC siRNA. Cell proliferation was
examined by CCK-8 assays in (C) U251 and (D) T98G cells following
transfection with miR-25 inhibitor and DKK3 siRNA or NC siRNA. Cell
migration was examined using wound-healing assays in (E) U251
(magnification, ×200) and (F) T98G cells (magnification, ×40)
following transfection with miR-25 inhibitor and DKK3 siRNA or NC
siRNA. **P<0.01 vs. miR-25 in + siNC. DKK3, dickkopf WNT
signaling pathway inhibitor 3; miR, microRNA; RT-qPCR, reverse
transcription-quantitative PCR; siRNA, small interfering RNA; NC,
negative control; CCK-8, Cell Counting Kit-8. |
Discussion
Since glioma is a malignant brain tumour, it is
important to understand the molecular mechanism underlying glioma
progression. In the present study, it was observed that miR-25
expression levels were significantly increased in glioma tissue
samples and cell lines, and miR-25 upregulation was associated with
glioma progression. Knockdown of miR-25 significantly inhibited
glioma cell proliferation and migration. FBXW7 and DKK3 were
identified as target genes of miR-25. FBXW7 and DKK3 expression
levels were significantly downregulated in glioma tissue samples
and cells lines, and their expression levels were negatively
regulated by miR-25 in glioma cells. Furthermore, knockdown of
FBXW7 and DKK3 impaired the miR-25-induced effects on glioma cell
proliferation and migration.
A previous study investigated the expression
profiles of miRs in four patients with primary gliomas (grade II)
that spontaneously progressed to secondary gliomas (grade IV), and
demonstrated that miR-25 expression levels increased following
progression (24). In the current
study, miR-25 was significantly upregulated in glioma tissue
compared with normal brain tissue samples as well as in glioma cell
lines compared with NHAs. In addition, high miR-25 expression in
glioma tissue samples was associated with advanced clinical stage,
which suggests that upregulation of miR-25 may be associated with
glioma progression. To further clarify the function of miR-25 in
glioma progression, U251 and T98G cell lines were transfected with
miR-25 inhibitor. The results demonstrated that the knockdown of
miR-25 inhibits glioma cell proliferation and migration.
It is well established that miRs function by
regulating the expression of their target genes (25,26).
TargetScan bioinformatics software was used in the present study to
predict target genes of miR-25. FBXW7 and DKK3 were identified as
two potential target genes of miR-25 and so were subsequently
examined. Luciferase reporter gene assays confirmed that FBXW7 and
DKK3 are direct target genes of miR-25, and their expression was
negatively regulated by miR-25 in glioma cell lines. FBXW7 is a
well-known tumour suppressor; Hagedorn et al (27) reported that FBXW7 inhibits glioma
cell proliferation and is a prognostic marker for survival in
patients with glioblastoma, while the loss of FBXW7 serves an
important role in glioma malignancy by allowing the accumulation of
multiple oncoproteins. Cao et al (28) reported a tumour-suppressive role of
the long noncoding RNA metastasis associated lung adenocarcinoma
transcript 1 in glioma cells via the suppression of miR-155
expression and activation of FBXW7. The current study demonstrated
that FBXW7 inhibition attenuated the suppressive effects on glioma
cell proliferation and migration induced by miR-25 downregulation,
which suggests that FBXW7 may be involved in miR-25-mediated glioma
cell proliferation and migration.
DKK3 also functions as a tumour suppressor in
glioma; Mizobuchi et al (21)
reported that DKK3 was significantly downregulated in glioma tissue
compared with normal brain tissue samples, which is consistent with
the current study. In addition, overexpression of DKK3 has been
reported to inhibit glioma cell proliferation and induce cell
apoptosis via the activation of JUN phosphorylation, caspase-9 and
caspase-3, and the reduction of β-catenin (21). Li et al (20) demonstrated that miR-92b increased
glioma cell proliferation and inhibited cell apoptosis by targeting
DKK3. In the current study, FBXW7 inhibition attenuated the
miR-25-induced suppressive effects on glioma cell proliferation and
migration, which suggests that DKK3 may also be involved in
miR-25-mediated glioma cell proliferation and migration.
An association between miR-25 and FBXW7 has been
previously reported in several other types of cancer, including
oesophageal squamous cell carcinoma (29), lung cancer (30) and gastric cancer (31). In addition, miR-25 has been found to
promote the migration of melanoma cells by targeting DKK3 (32). Thus, the current study increases our
understanding of the importance of the miR-25/FBXW7 axis and
miR-25/DKK3 axis in human cancer.
To the best of our knowledge, the current study is
the first to demonstrate that miR-25 plays a promoting role in
glioma cell proliferation and migration, at least in part through
directly targeting FBXW7 and DKK3. Therefore, miR-25 may serve as a
potential molecular target for the treatment of glioma.
Acknowledgements
Not applicable.
Funding
The present study was supported by Scientific
Research Project of the Commission of Finance of Hunan province
(grant no. 62.2).
Availability of data and materials
The datasets used and/or analysed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
CS designed the study and revised the manuscript. GP
collected tissue samples, conducted statistical analysis and wrote
the manuscript. YL and CY performed all the experiments.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Affiliated Hospital of Xiangya Hospital of Central
South University (Changsha, China). All participants provided
written informed consent.
Patient consent for publication
All participants provided written informed
consent.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brower JV, Clark PA, Lyon W and Kuo JS:
MicroRNAs in cancer: Glioblastoma and glioblastoma cancer stem
cells. Neurochem Int. 77:68–77. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Murphy AM and Rabkin SD: Current status of
gene therapy for brain tumors. Transl Res. 161:339–354. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang Z, Li X, Xiao Q and Wang Z:
MiR-574-5p mediates the cell cycle and apoptosis in thyroid cancer
cells via Wnt/beta-catenin signaling by repressing the expression
of Quaking proteins. Oncol Lett. 15:5841–5848. 2018.PubMed/NCBI
|
|
6
|
Luo T, Yan Y, He Q, Ma X and Wang W:
miR-328-5p inhibits MDA-MB-231 breast cancer cell proliferation by
targeting RAGE. Oncol Rep. 39:2906–2914. 2018.PubMed/NCBI
|
|
7
|
Li Z, Guo J, Ma Y, Lin Z and Zhang L:
Oncogenic role of MicroRNA-30b-5p in glioblastoma through targeting
proline-rich transmembrane protein 2. Oncol Res. 26:219–230. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu R, Zhu X, Chen F, Huang C, Ai K, Wu H,
Zhang L and Zhao X: LncRNA XIST/miR-200c regulates the stemness
properties and tumourigenicity of human bladder cancer stem
cell-like cells. Cancer Cell Int. 18:412018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu Y, Zhao H, Rao M and Xu S:
MicroRNA-365 inhibits proliferation, migration and invasion of
glioma by targeting PIK3R3. Oncol Rep. 37:2185–2192. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang T, Ma G, Zhang Y, Huo H and Zhao Y:
miR-599 inhibits proliferation and invasion of glioma by targeting
periostin. Biotechnol Lett. 39:1325–1333. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang L, Wang C, Lei F, Zhang L, Zhang X,
Liu A, Wu G, Zhu J and Song L: miR-93 promotes cell proliferation
in gliomas through activation of PI3K/Akt signaling pathway.
Oncotarget. 6:8286–8299. 2015.PubMed/NCBI
|
|
12
|
Chen H, Pan H, Qian Y, Zhou W and Liu X:
MiR-25-3p promotes the proliferation of triple negative breast
cancer by targeting BTG2. Mol Cancer. 17:42018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Casadei L, Calore F, Creighton CJ,
Guescini M, Batte K, Iwenofu OH, Zewdu A, Braggio DA, Bill KL,
Fadda P, et al: Exosome-derived miR-25-3p and miR-92a-3p stimulate
liposarcoma progression. Cancer Res. 77:3846–3856. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang J, Gong X, Tian K, Chen D, Sun J,
Wang G and Guo M: miR-25 promotes glioma cell proliferation by
targeting CDKN1C. Biomed Pharmacother. 71:7–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Peng G, Yuan X, Yuan J, Liu Q, Dai M, Shen
C, Ma J, Liao Y and Jiang W: miR-25 promotes glioblastoma cell
proliferation and invasion by directly targeting NEFL. Mol Cell
Biochem. 409:103–111. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cao J, Ge MH and Ling ZQ: Fbxw7 tumor
suppressor: A vital regulator contributes to human tumorigenesis.
Medicine (Baltimore). 95:e24962016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou C, Shen L, Mao L, Wang B, Li Y and Yu
H: miR-92a is upregulated in cervical cancer and promotes cell
proliferation and invasion by targeting FBXW7. Biochem Biophys Res
Commun. 458:63–69. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin J, Ji A, Qiu G, Feng H, Li J, Li S,
Zou Y, Cui Y, Song C, He H and Lu Y: FBW7 is associated with
prognosis, inhibits malignancies and enhances temozolomide
sensitivity in glioblastoma cells. Cancer Sci. 109:1001–1011. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee EJ, Jo M, Rho SB, Park K, Yoo YN, Park
J, Chae M, Zhang W and Lee JH: Dkk3, downregulated in cervical
cancer, functions as a negative regulator of beta-catenin. Int J
Cancer. 124:287–297. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Q, Shen K, Zhao Y, Ma C, Liu J and Ma
J: MiR-92b inhibitor promoted glioma cell apoptosis via targeting
DKK3 and blocking the Wnt/beta-catenin signaling pathway. J Transl
Med. 11:3022013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mizobuchi Y, Matsuzaki K, Kuwayama K,
Kitazato K, Mure H, Kageji T and Nagahiro S: REIC/Dkk-3 induces
cell death in human malignant glioma. Neuro Oncol. 10:244–253.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pisapia DJ: The updated world health
organization glioma classification: Cellular and molecular origins
of adult infiltrating gliomas. Arch Pathol Lab Med. 141:1633–1645.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Malzkorn B, Wolter M, Liesenberg F,
Grzendowski M, Stühler K, Meyer HE and Reifenberger G:
Identification and functional characterization of microRNAs
involved in the malignant progression of gliomas. Brain Pathol.
20:539–550. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xiao F, Li Y, Wan Y and Xue M:
MircroRNA-139 sensitizes ovarian cancer cell to cisplatin-based
chemotherapy through regulation of ATP7A/B. Cancer Chemother
Pharmacol. 81:935–947. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ruan L, Chen J, Ruan L, Yang T and Wang P:
MicroRNA-186 suppresses lung cancer progression by targeting SIRT6.
Cancer Biomark. 21:415–423. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hagedorn M, Delugin M, Abraldes I, Allain
N, Belaud-Rotureau MA, Turmo M, Prigent C, Loiseau H, Bikfalvi A
and Javerzat S: FBXW7/hCDC4 controls glioma cell proliferation in
vitro and is a prognostic marker for survival in glioblastoma
patients. Cell Div. 2:92007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cao S, Wang Y, Li J, Lv M, Niu H and Tian
Y: Tumor-suppressive function of long noncoding RNA MALAT1 in
glioma cells by suppressing miR-155 expression and activating FBXW7
function. Am J Cancer Res. 6:2561–2574. 2016.PubMed/NCBI
|
|
29
|
Hua Y, Zhao K, Tao G, Dai C and Su Y:
miR-25 promotes metastasis via targeting FBXW7 in esophageal
squamous cell carcinoma. Oncol Rep. 38:3030–3038. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xiang J, Hang JB, Che JM and Li HC: MiR-25
is up-regulated in non-small cell lung cancer and promotes cell
proliferation and motility by targeting FBXW7. Int J Clin Exp
Pathol. 8:9147–9153. 2015.PubMed/NCBI
|
|
31
|
Gong J, Cui Z, Li L, Ma Q, Wang Q, Gao Y
and Sun H: MicroRNA-25 promotes gastric cancer proliferation,
invasion, and migration by directly targeting F-box and WD-40
domain protein 7, FBXW7. Tumour Biol. 36:7831–7840. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huo J, Zhang Y, Li R, Wang Y, Wu J and
Zhang D: Upregulated MicroRNA-25 mediates the migration of melanoma
cells by targeting DKK3 through the WNT/beta-Catenin Pathway. Int J
Mol Sci. 17:E11242016. View Article : Google Scholar : PubMed/NCBI
|