Introduction
In recent years, obesity has received increasing
attention because it leads to various metabolic diseases, such as
hyperlipidemia, hypertension, atherosclerosis and type II diabetes
mellitus (1). The liver is an
important organ associated with lipid metabolism. Notably,
increases in lipid metabolism is mainly observed in the liver,
which can lead to chronic diseases related to hepatocytes (2). If left untreated, this increase in
metabolism can lead to hepatocellular carcinoma therefore, it is
necessary to investigate the underlying molecular mechanism
(3). Adipogenesis is regulated via a
complex process involving coordinated changes in hormone
sensitivity and gene expression regulated by various transcription
factors (4). The sterol regulatory
element-binding protein (SREBP) family of transcription factors
have pivotal roles in the regulation of cellular lipid metabolism
(5). SREBP-1 mainly regulates the
metabolism of fatty acids and triglycerides, whilst SREBP-2
regulates the biosynthesis of cholesterol (6). Fatty acid synthase (FAS), a protein
related to fat synthesis, is regulated by SREBP-l (7). SREBP-2 regulates
3-hydroxy-3-methylglutaryl CoA reductase (HMGCR) expression and
thereby catalyzes the rate-limiting step of cholesterol
biosynthesis (8). Furthermore,
AMP-activated protein kinase (AMPK) is an important protein
involved in energy metabolism and has a significant role in the
regulation of fatty acid metabolism. An in vitro study
determined that alliin attenuates adipogenesis induced by
1,3-dichloropropan-2ol in HepG2 cells by activating the AMPK/SREBP
signaling pathway (9). Therefore, it
is reasonable to speculate that the AMPK/SREBP pathway might be
important for the amelioration of lipogenesis during palmitic acid
treatment.
For the development of novel antilipidemic drugs
with high efficacy and few adverse effects, attention has gradually
shifted to the natural plant flavonoids in recent years. Apigenin,
a type of plant flavonoid with the chemical name
4′,5,7-dihydroxyflavone, is recognized as a bioactive flavonoid
that has antioxidant, anticancer and anti-inflammatory properties
that can also lower blood pressure (10–12). In
particular, a previous study has demonstrated that apigenin can
alleviate myocardial hypertrophy and the hypertrophy-induced
glucose and lipid metabolism abnormalities in rats (13). In addition, apigenin alleviates
glucose metabolic disorder induced by a high-fat diet in
20-week-old mice (14). It still
remains unclear whether apigenin has a hypolipidemic effect, or
whether the AMPK/SREBP pathway is the major determinant of
anti-adipogenic activity. Palmitic acid is a saturated high-grade
fatty acid that is widely distributed in animal and vegetable oils
in the form of glycerides, and is commonly used to establish
lipotoxic in vitro and in vivo models (15).
The present study established a common
hyperlipidemia model by using appropriate doses of palmitic acid to
determine whether apigenin lowered lipid levels and, to explore the
underlying mechanism in depth.
Materials and methods
Reagents
Palmitic acid and dimethyl sulfoxide (DMSO) were
purchased from Sigma-Aldrich; Merck KGaA. The apigenin standard was
purchased from Beijing Solarbio Technology Co. The AMPK inhibitor
compound C was purchased from Selleck Chemicals LLC.
Anti-phospho-AMPK (P-AMPK; Thr172; cat. no. 2535) was purchased
from Cell Signaling Technology, Inc., anti-AMPKα1 (cat. no.
10929-2-AP), SREBP-2 (cat. no. 14508-1-AP), FAS (cat. no.
13098-1-AP), HMGCR (cat. no. 13533-1-AP) and β-actin (cat. no.
66009-1-lg) were purchased from Proteintech Group Inc., SREBP-1
(cat. no. ab3259) was purchased from Abcam. Horseradish peroxidase
(HRP)-conjugated goat anti-mouse IgG (H + L; cat. no. SA00001-1)
and HRP-conjugated goat anti-rabbit IgG (H + L; cat. no. SA00001-2)
secondary antibodies were obtained from Proteintech Group Inc.
Cell culture and treatment
Liver cancer HepG2 cells, purchased from the
American Type Culture Collection, were cultured in high-glucose
DMEM (GE Healthcare Life Sciences) with 10% fetal bovine serum
(Clark Bioscience) and 1% penicillin/streptomycin at 37°C in an
atmosphere containing 5% CO2. The cells were dissociated
with 0.25% trypsin (w/v) and 0.52 mM EDTA [M&C Gene Technology
(Bejing) Ltd.] and routinely sub-cultured at 80% confluency.
Cell cultures were treated with various
concentrations of apigenin in DMSO as the carrier solvent. Palmitic
acid binds to fatty-acid-free BSA (Beijing Solarbio Science &
Technology Co., Ltd.). In brief, palmitic acid was dissolved in 1X
PBS and a 250 mM stock solution was obtained following various
cycles of incubation in a water bath at 70°C and vortexing. The
stock solution was then added to serum-free DMEM containing 5%
fatty-acid-free BSA to obtain a 250 µM palmitic acid solution, and
the resulting diluted solution was used for the cell treatments
(16,17). AMPK inhibitor compound C was diluted
with DMSO to a final concentration of 10 µM and was used to verify
the signaling pathway.
Cell viability assays
Cells were seeded in 96-well plates at a
concentration of 1×104 cells/well (Corning Inc.) and
allowed to adhere overnight at 37°C. Cells were then treated with
different concentrations of apigenin (0, 20, 40, 80, 160, 320, 640
and 1,280 µM) or palmitic acid (0, 62.5, 125, 250, 500 and 1,000
µM) for 24 h, and 20 µl of MTT (Sigma-Aldrich; Merck KGaA) solution
(5 mg/ml in 1% PBS) was added to each well and incubated for 4 h at
37°C prior to the end of the culture. The culture medium was
subsequently removed and the formazan crystals were dissolved using
150 µl DMSO/well. Absorbance values at 570 nm were determined using
a microplate reader (BioTek Instruments, Inc.).
Oil Red O Staining
HepG2 cells were inoculated at a density of
4×105 cells per well in 6-well plates and cultured with
different concentrations of apigenin (10, 20 and 40 µM) and 250 µM
palmitic acid for 24 h. Cells were then washed twice with cold 1X
PBS and fixed with 4% paraformaldehyde for 40 min at 37°C.
Subsequently, the residual paraformaldehyde was washed with
double-distilled water, and cells were incubated with 60%
isopropanol for 3–5 sec. Cells were then stained with freshly
prepared 0.5% Oil Red O solution in 60% isopropanol for 1 h at 37°C
in the dark. Lipid droplets in the cells were observed and imaged
using a bright-field microscope (Olympus Corporation) at a
magnification of ×400. In addition, the accumulated lipids were
extracted with 100% isopropanol for 3–5 min at room temperature,
and the absorbance at 570 nm was determined using a microplate
reader.
Triglyceride (TG) and total
cholesterol (TC) assays
HepG2 cells were inoculated at a density of
4×105 cells/well in 6-well plates and pretreated with 10
µM AMPK inhibitor compound C for 1 h at 37°C then cultured with
different concentrations of apigenin (10, 20 and 40 µM) and 250 µM
palmitic acid for 24 h at 37°C in a culture chamber with 5%
CO2. The intracellular TC and TG contents were measured
using a TC assay kit (cat. no. E1015; Applygen Technologies Inc.)
and a TG assay kit (cat. no. E1013; Applygen Technologies Inc.)
according to the manufacturer's instructions. The absorbance values
were measured at 570 nm. Bicinchoninic acid (BCA) protein
quantitative kit (Beyotime Institute of Biotechnology) was used to
determine the protein concentration of the samples, and the
absorbance value of the proteins was measured at 562 nm. The
intracellular TC and TG contents are presented as µM/mg cellular
protein.
Western blot analysis
HepG2 cells were seeded in 6-well plates at a
concentration of 6×105 cells per well and pretreated
with 10 µM AMPK inhibitor compound C for 1 h and then cultured with
different concentrations of apigenin (10, 20 and 40 µM) and 250 µM
palmitic acid (16,17). The cell pellets were lysed on ice for
30 min in RIPA buffer (Beyotime Institute of Biotechnology)
supplemented with 1:100 PMSF (Beijing Solarbio Science &
Technology Co., Ltd.) according to the manufacturer's
specifications and centrifuged at 14,000 × g at 4°C for 8 min. The
supernatant was then collected and the protein concentration was
determined using a BCA protein concentration assay kit (Beyotime
Institute of Biotechnology). An equal amount of protein extract (80
µg) was denatured in the sample buffer (Bio-Rad Laboratories, Inc.)
at 98°C for 10 min and subsequently separated via 8% SDS-PAGE and
then transferred to polyvinylidene fluoride membranes. The
membranes were blocked with 3% BSA in Tris-buffered saline
containing 0.1% Tween 20 (TBST) buffer for 4 h at room temperature.
Subsequently, the membranes were incubated overnight at 4°C with
primary antibodies anti-phosphorylated (P)-AMPK (1:500), anti-AMPK
(1:500) anti-SREBP-1 (1:300), anti-FAS (1:800), anti-SREBP-2
(1:500), anti-HMGCR (1:800) and anti-β-actin (1:800 dilution).
Following incubation with the primary antibodies, the membranes
were subjected to three 15-min washes with TBST and then incubated
with horseradish peroxidase-conjugated secondary anti-mouse
antibody and anti-rabbit antibody (1:3,000 dilution) at room
temperature for 2 h. Protein bands were visualized using the
Enhanced Chemiluminescence Plus substrate (Azure Biosystems, Inc.).
All Western blot assays were performed at least three times. The
resulting images were analyzed using the ImageJ software (version
1.51; National Institutes of Health) with β-actin as the loading
control.
Statistical analysis
Data were processed using SPSS v18.0 statistical
software (SPSS Inc.), and the measurement data were expressed as
the mean ± standard deviation. Statistical analysis was performed
by one-way analysis of variance with Tukey's test and Dunnett's
test used for post hoc analysis. Histograms were plotted by Prism
software v5.0 (GraphPad Software, Inc.). P<0.05 was considered
to indicate a statistically significant difference.
Results
Effects of apigenin and palmitic acid
on cell viability
To analyze the effect of apigenin and palmitic acid
on the viability of HepG2 cells, the cells were exposed to
different concentrations of apigenin or palmitic acid for 24 h.
Results demonstrated that apigenin significantly decreased the
viability of HepG2 cells at concentrations from 320 to 1,280 µM
(P<0.01; Fig. 1A), and palmitic
acid significantly decreased the viability of HepG2 cells at
concentrations from 500 to 1,000 µM (P<0.01; Fig. 1B).
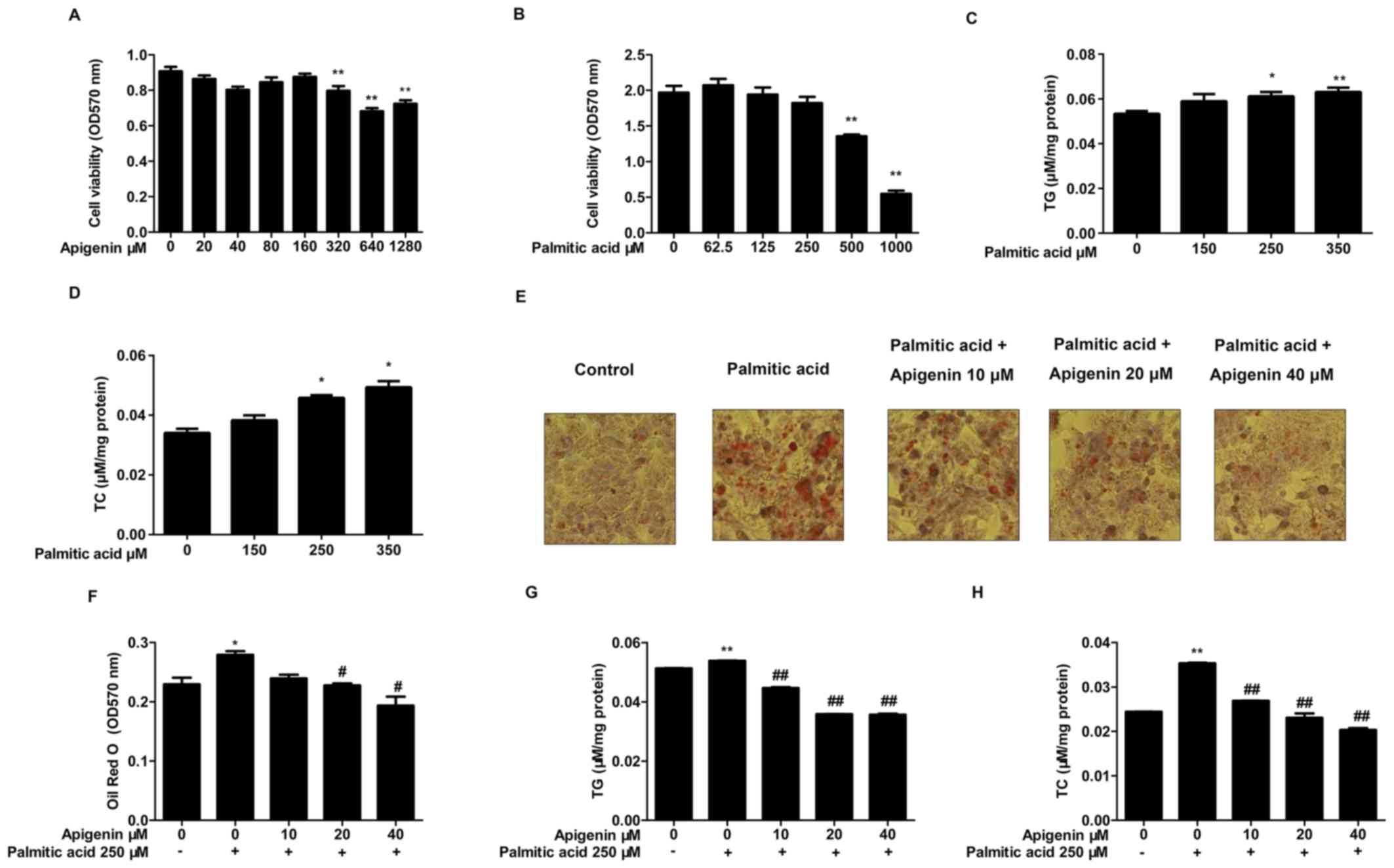 | Figure 1.Effects of apigenin and palmitic acid
on the viability of HepG2 cells, and analysis of apigenin-mediated
inhibition of lipid accumulation in HepG2 cells. (A) Cell viability
following treatment with various concentrations of apigenin (0, 20,
40, 80, 160, 320, 640 and 1,280 µM) and (B) palmitic acid (0, 62.5,
125, 250, 500 and 1,000 µM) (n=6). (C) TG and (D) TC contents
following treatment with various concentrations of palmitic acid
(150, 250 and 350 µm). (E) Photomicrographs of Oil Red O-stained,
intracellular lipids (red staining) were obtained by light
microscopy (magnification, ×400) and (F) quantification of Oil Red
O absorbance. (G) TG and (H) TC contents following treatment with
palmitic acid (0 or 250 mm) various concentrations of apigenin (0,
10, 20 and 40 mm; n=3). *P<0.05 and **P<0.01 vs. control;
#P<0.05 and ##P<0.01 vs. treatment with
palmitic acid alone. TG, triglyceride; TC, total cholesterol. |
Effects of apigenin on lipid
accumulation in palmitic acid-treated HepG2 cells
The cells were exposed to different concentrations
of palmitic acid (150, 250 and 350 µM) for 24 h and cellular TC and
TG contents were subsequently measured. The results demonstrated
that the intracellular TC and TG levels were significantly elevated
when cells were treated with 250 and 350 µM palmitic acid compared
with control cells (P<0.05; Fig. 1C
and D). The accumulation of lipid droplets in cells was
measured by Oil red O staining. The results indicated that the
lipid content in HepG2 cells treated with 250 µM palmitic acid was
increased significantly compared with the control cells. In
addition, 20 and 40 µM apigenin treatment significantly decreased
the level of the lipid content compared with treatment with
palmitic acid alone (P<0.05; Fig. 1E
and F).
To verify the Oil red O staining results, the
cellular TC and TG contents were measured. Results demonstrated
that the intracellular TC and TG levels were significantly elevated
when cells were treated with 250 µM palmitic acid compared with the
control cells (P<0.01; Fig. 1G and
H). However, treatment with 250 µM palmitic acid and different
concentrations of apigenin (10, 20 and 40 µM) significantly
suppressed intracellular TC and TG levels in a
concentration-dependent manner (P<0.01; Fig. 1G and H).
Effects of apigenin on the AMPK and
P-AMPK proteins in palmitic acid-induced HepG2 cells
The activity of AMPK is closely associated with
phosphorylation of the Thr172 site in the α subunit (P-AMPKα1).
Therefore, the activity of AMPK can be evaluated by measuring the
levels of P-AMPKα protein. Results demonstrated that the
P-AMPK/AMPK ratio in the cells treated with 250 µM palmitic acid
was significantly lower than in the normal group (P<0.01;
Fig. 2A). However, the treatment of
HepG2 cells with 250 µM palmitic acid and different concentrations
of apigenin (10, 20 and 40 µM) significantly increased AMPK
phosphorylation (P<0.01; Fig.
2A). The P-AMPK/β-actin ratio in the cells treated with 250 µM
palmitic acid and different concentrations of apigenin (10, 20 and
40 µM) produced the same results as aforementioned (P<0.01;
Fig. 2B).
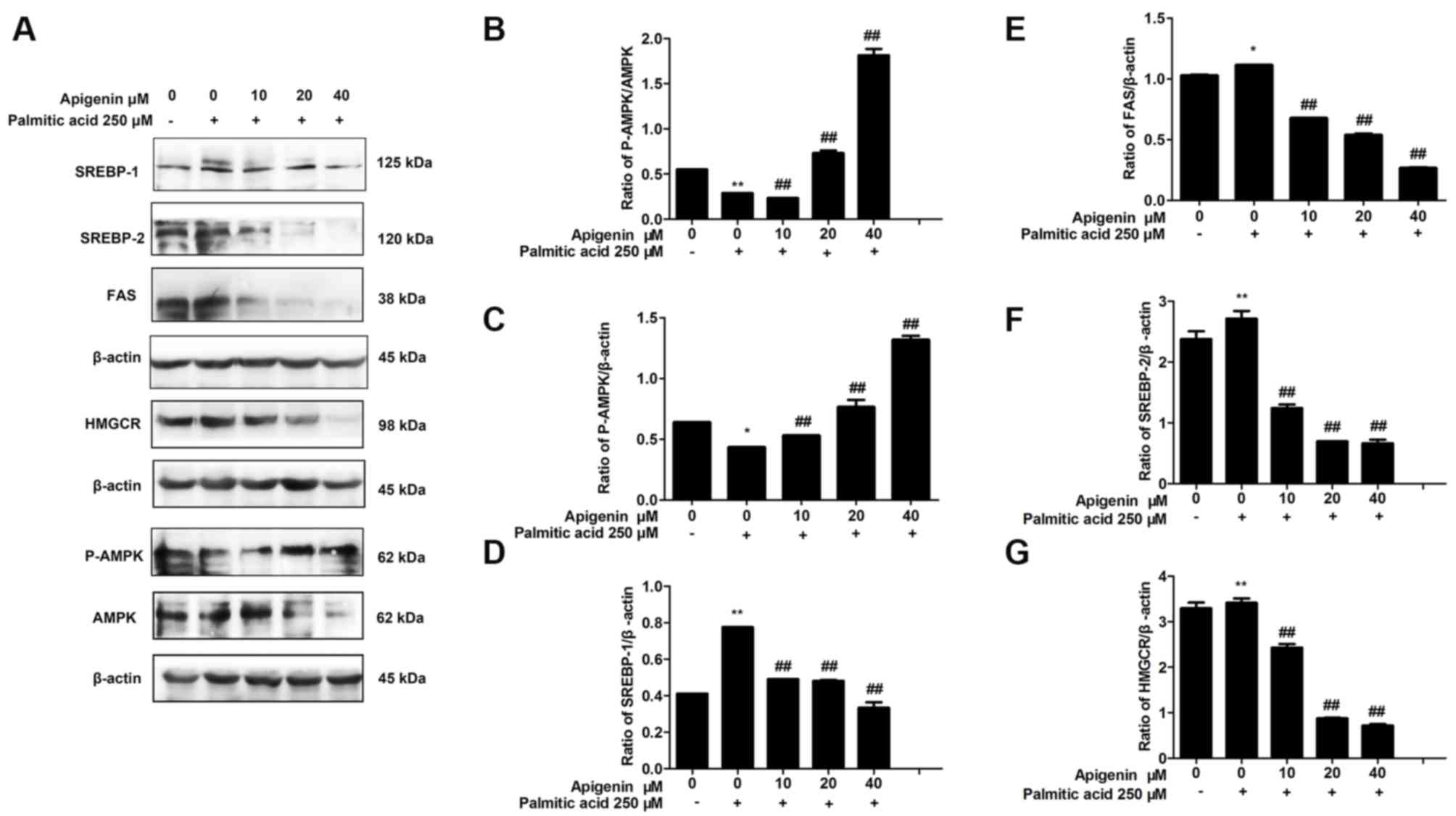 | Figure 2.Effect of apigenin on the expression
of AMPK, P-AMPK, SREBP-1, FAS, SREBP-2 and HMGCR proteins in HepG2
cells. (A) Representative western blots and (B) quantification of
P-AMPK/AMPK, (C) P-AMPK/β-actin, (D) SREBP-1, (E) FAS, (F) SREBP-2
and (G) HMGCR protein expression measured by western blot analysis
(n=3). *P<0.05 and **P<0.01 vs. control;
##P<0.01 vs. treatment with palmitic acid alone.
AMPK, AMP-activated protein kinase; P-AMPK, phosphorylated AMPK;
SREBP, sterol regulatory element-binding protein; FAS, fatty acid
synthase; HMGCR, 3-hydroxy-3-methylglutaryl CoA reductase. |
Effects of apigenin on the AMPK/SREBP
signaling pathway in palmitic acid-treated HepG2 cells
To investigate the mechanism underlying the
apigenin-mediated inhibition of lipid production and regulation of
the AMPK signaling pathway in HepG2 cells, the expression levels of
proteins associated with fatty acid synthesis and cholesterol
synthesis were determined.
Results demonstrated that compared with the control
cells, the levels of SREBP-1 (P<0.01; Fig. 2B) and FAS (P<0.05; Fig. 2C) increased significantly following
treatment with 250 µM palmitic acid, whereas the levels of SREBP-1
and FAS were decreased by apigenin in a concentration-dependent
manner (P<0.01; Fig. 2B and C).
SREBP-2 and HMGCR protein expression increased significantly upon
treatment with 250 µM palmitic acid compared with the control
(P<0.01; Fig. 2D and E). However,
HepG2 cells treated with 250 µM palmitic acid and different
concentrations of apigenin (10, 20 and 40 µM) inhibited the
upregulation of SREBP-2 and HMGCR protein expression (P<0.01;
Fig. 2D and E).
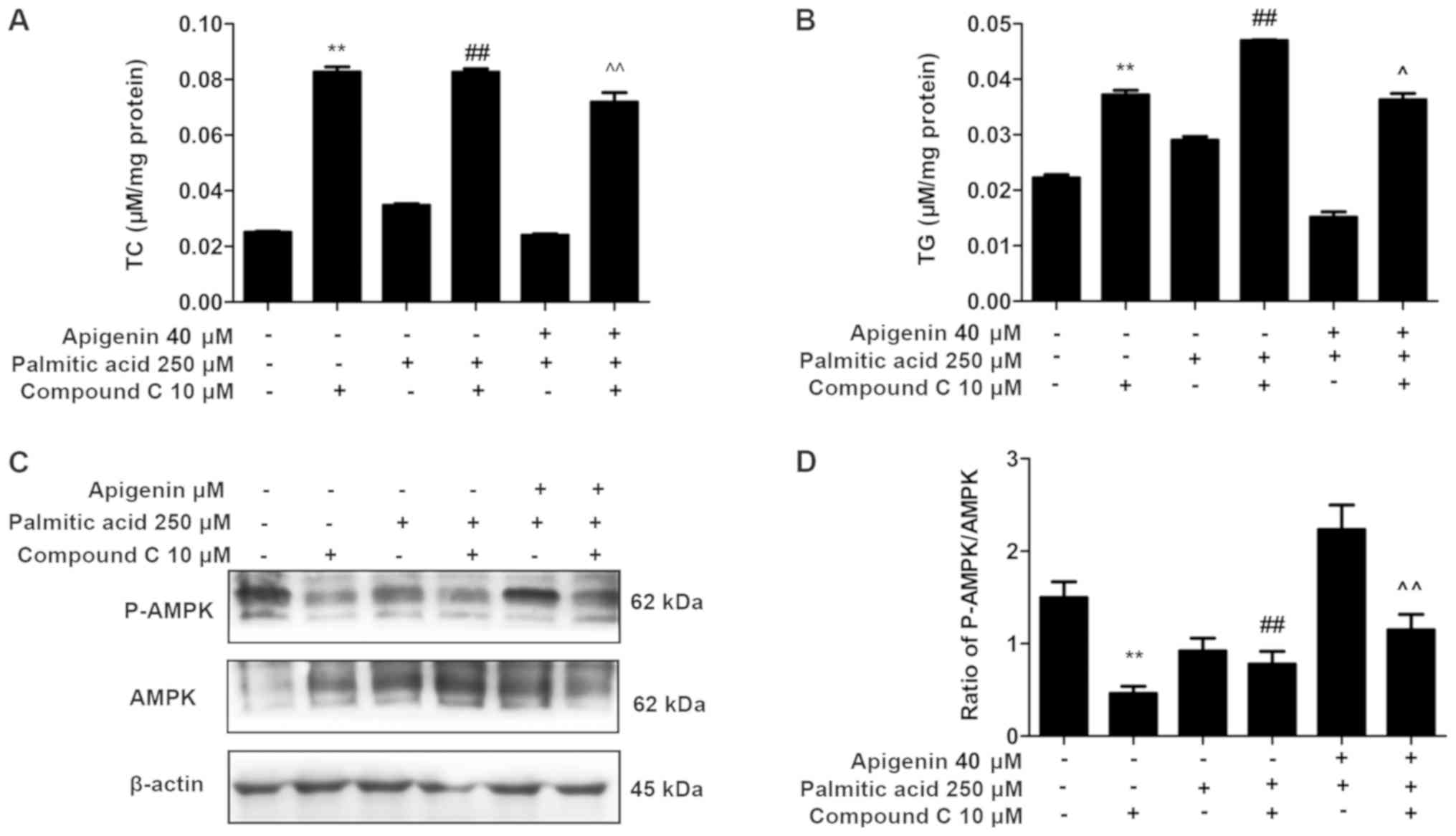
Effects of compound C on lipid
accumulation and AMPK expression in palmitic acid-treated HepG2
cells
To further validate the previous findings, the
intracellular TC and TG contents were measured following
pretreatment with 10 µM compound C for 1 h. The compound C
treatment significantly reversed the apigenin-induced reductions in
lipid accumulation and led to increases in TC and TG contents
(P<0.01; Fig. 3A and B).
In addition, the protein expression of AMPK and
P-AMPK were examined. The increase in P-AMPK/AMPK ratio induced by
apigenin was reversed by pretreatment with compound C in HepG2
cells (P<0.01; Fig. 3C and D).
These results indicated that apigenin activated the AMPK signaling
pathway in HepG2 cells.
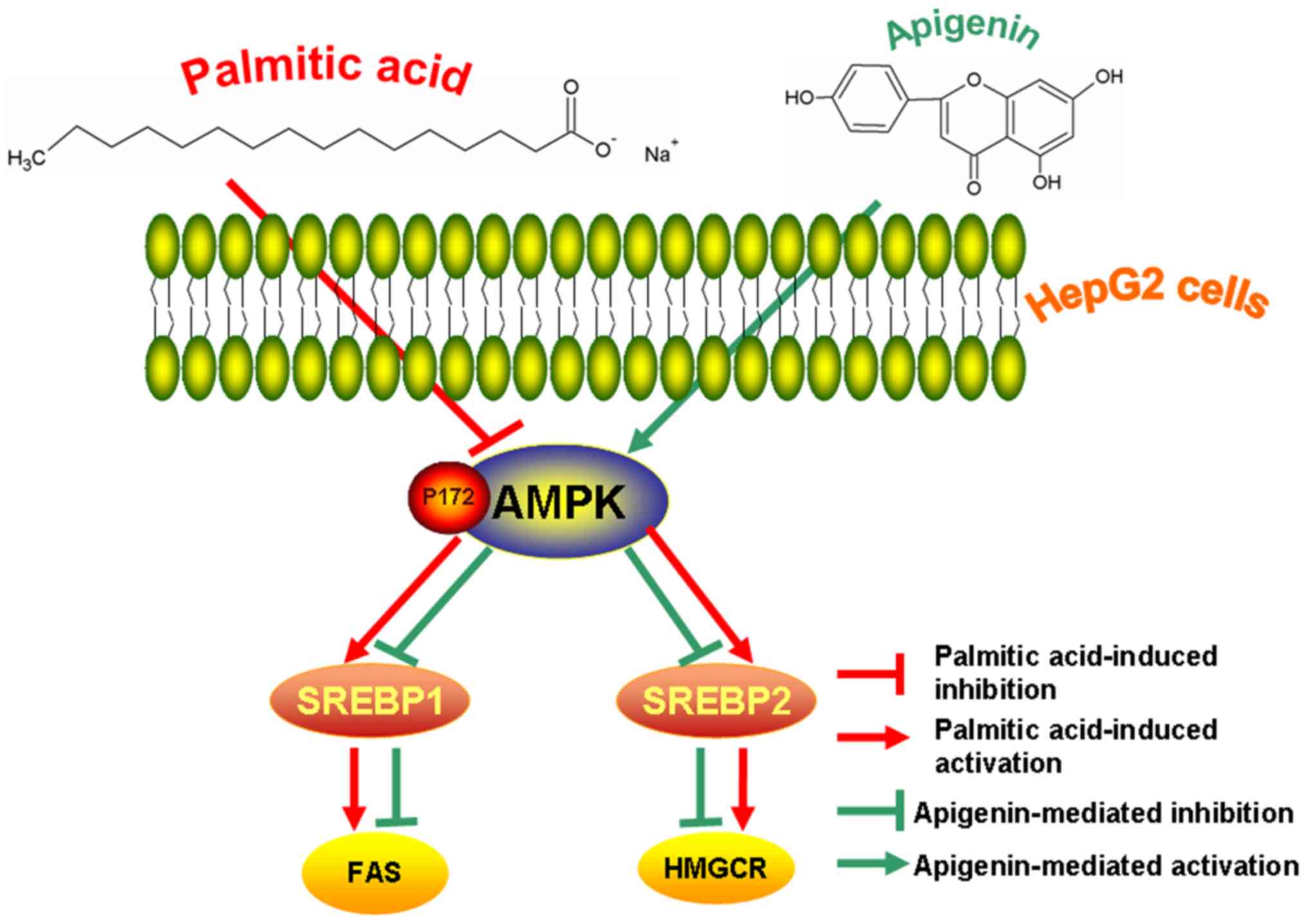
The present findings indicated that apigenin
effectively inhibited the effects of palmitic acid-induced lipid
accumulation via the AMPK/SREBP signaling pathway in HepG2 cells
(Fig. 4).
Discussion
Numerous flavonoids display anti-obesity and
hypolipidemic effects in animals and humans. It has been
demonstrated that apigenin exerts anticancer, anti-inflammatory and
hypotensive effects (10–12). Previous studies have determined that
apigenin reduces the accumulation of intracellular lipids in 3T3-L1
cells (18), and also attenuates
atherogenesis via upregulating ATP binding cassette subfamily A
member 1-mediated cholesterol efflux and by inhibiting inflammation
(19). Apigenin suppresses
adipogenesis through the downregulation of peroxisome proliferator
activated receptor-γ function by activating AMPK in 3T3-L1 cells
(4). Nonetheless, the regulatory
mechanism underlying the apigenin-mediated suppression of lipid
accumulation has yet to be fully identified.
The present study identified that apigenin served an
essential role in the reduction of excessive lipid accumulation
induced by palmitic acid in HepG2 cells. Results demonstrated that
apigenin was not toxic to HepG2 cells at concentrations of <320
µM, whilst palmitic acid exhibited no apparent toxicity to HepG2
cells at concentrations <250 µM. Of note, apigenin has been
determined to be cytotoxic to 3T3-L1 cells at concentrations as low
as 10 µM (4), whilst palmitic acid
is cytotoxic to HepG2 cells at concentrations as low as 200 µM,
which is in line with the present study (20). The discrepancies might be due to
differences between cell lines or assay systems with further
investigation required to elucidate a clear explanation. A previous
study demonstrated that apigenin is a poorly soluble bioactive
compound/nutraceutical with a poor bioavailability upon oral
administration (21). The oral
bioavailability of apigenin is relatively poor due to its low lipid
(0.001–1.63 mg/ml in nonpolar solvents) and water (2.16 µg/ml in
water) solubility, which has severely limited its development for
clinical applications (22,23). However, there are a number of methods
in which the bioavailability of apigenin can be improved, including
utilizing spray-drying techniques and apigenin-loaded polymeric
micelles (24,25). Therefore, future research into
apigenin should not only take into consideration the lower reported
toxic dose but also try to improve the oral bioavailability of
apigenin.
To further elucidate the molecular mechanism
underlying the apigenin-mediated suppression of adipogenesis, the
present study investigated the activation of AMPK, which is an
important kinase that regulates energy homeostasis. This key
protein is involved in various cell signal transduction pathways,
senses the intracellular energy state and has an important role in
the regulation of glucose and lipid metabolism (26,27).
AMPK activation reportedly leads to changes in obesity and has been
linked to obesity prevention (28,29).
Therefore, the pharmacological activity of AMPK is expected to have
an important role in the study of apigenin-induced liver lipid
accumulation reduction. This study identified that cotreatment of
apigenin (10, 20 and 40 µM) and 250 µM palmitic acid led to a
dose-dependent upregulation of the P-AMPK/AMPK ratio compared with
the group treated with 250 µM palmitic acid alone. This indicated
that apigenin activated AMPK phosphorylation in the palmitic
acid-induced HepG2 cells. Thus, to further validate the
experimental results, cells were treated with the AMPK inhibitor
compound C. The increase in the P-AMPK/AMPK ratio induced by
apigenin in HepG2 cells was inhibited by pretreatment with compound
C in HepG2 cells. Therefore, apigenin inhibited excessive lipid
accumulation induced by palmitic acid by activating AMPK
phosphorylation.
Changes in proteins associated with the AMPK/SREBP
pathway were investigated to fully elucidate the hypolipidemic
mechanism of apigenin. Apigenin treatment activates AMPK thereby
further inhibiting the synthesis of fatty acids and cholesterol
(8). It has been demonstrated that
AMPK inhibits SREBP-1 and SREBP-2 and thereby reduces the synthesis
of cholesterol, fatty acids, and triglycerides in the liver
(30). Lipid production in the liver
is mainly regulated by SREBP-1, which is a crucial regulatory
factor in fatty acid synthesis. SREBP-1 regulates the expression of
other genes involved in TG synthesis, such as FAS (31). SREBP-1c inhibition reduces the
expression of FAS and alleviates fat-containing liver lesions in
mice (32). Similarly, SREBP-2 is
mainly involved in the regulation of cholesterol synthesis, and
HMGCR is a target gene of SREBP-2 (33). Decreased SREBP-2 activity can lead to
reduced HMGCR expression. In addition, studies have identified that
the presence of free cholesterol significantly increases the levels
of SREBP-2 and its target gene HMGCR in the livers of
hyperlipidemic mice (34,35). These studies indicate that the
changes in the levels of proteins in the AMPK/SREBP pathway are
correlated with the TC and TG levels. In the present study,
compared with palmitic acid treatment alone, cotreatment with
different concentrations of apigenin (10, 20 and 40 µM) and
palmitic acid significantly suppressed TC and TG levels, as well as
SREBP-1, FAS, SREBP-2, and HMGCR protein expression. Taken
together, these results demonstrated that apigenin decreased lipid
levels and alleviated the palmitic acid-induced accumulation of
lipids in HepG2 cells by activating the AMPK/SREBP signaling
pathway.
In summary, apigenin alleviated the palmitic
acid-induced increases in lipid accumulation in HepG2 cells thereby
reducing the lipid content. The underlying mechanism might be
associated with the activation of the intracellular AMPK/SREBP
signaling pathway. These findings provide a novel approach for the
development of natural hypolipidemic drugs to prevent lipid
accumulation.
Acknowledgements
Not applicable.
Funding
This study was funded by the Graduate Innovation
Fund of Jilin University, China (grant no. 419020201348).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SG, JL, ZM and BC conceived the study and
participated in its design. ZM and BC performed the experiments.
ZM, BC, ML and ST participated in the acquisition and
interpretation of the data. SG, JL, ZM and BC participated in
manuscript drafting. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Williams JA, Manley S and Ding WX: New
advances in molecular mechanisms and emerging therapeutic targets
in alcoholic liver diseases. World J Gastroenterol. 20:12908–12933.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wan Y, Liu LY, Hong ZF and Peng J: Ethanol
extract of Cirsium japonicum attenuates hepatic lipid
accumulation via AMPK activation in human HepG2 cells. Exp Ther
Med. 8:79–84. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang M, Yuan Y, Wang Q, Li X, Men J and
Lin M: The Chinese medicine Chai Hu Li Zhong Tang protects against
non-alcoholic fatty liver disease by activating AMPKα. Biosci Rep.
38:BSR201806442018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ono M and Fujimori K: Antiadipogenic
effect of dietary apigenin through activation of AMPK in 3T3-L1
cells. J Agric Food Chem. 59:13346–13352. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xiao X and Song BL: SREBP: A novel
therapeutic target. Acta Biochim Biophys Sin. 45:2–10. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shimano H: Sterol regulatory element
binding proteins (SREBPs): Transcriptional regulators of lipid
synthetic genes. Prog Lipid Res. 40:439–452. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wong TY, Lin SM and Leung LK: The flavone
luteolin suppresses SREBP-2 expression and post-translational
activation in hepatic cells. PLoS One. 10:e01356372015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mottillo EP, Desjardins EM, Crane JD,
Smith BK, Green AE, Ducommun S, Henriksen TI, Rebalka IA, Razi A,
Sakamoto K, et al: Lack of adipocyte AMPK exacerbates insulin
resistance and hepatic steatosis through brown and beige adipose
tissue function. Cell Metab. 24:118–129. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu J, Cheng B, Meng Z, Sun M, Fang B, Li
T, Sun M, Liu M and Guan S: Alliin attenuates
1,3-dichloro-2-propanol-induced lipogenesis in HepG2 cells through
activation of the AMP-activated protein kinase-dependent pathway.
Life Sci. 195:19–24. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lordan S, O'Neill C and O'Brien NM:
Effects of apigenin, lycopene and astaxanthin on 7
beta-hydroxycholesterol-induced apoptosis and Akt phosphorylation
in U937 cells. Br J Nutr. 100:287–296. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tong X and Pelling J: Targeting the
PI3K/Akt/mTOR axis by apigenin for cancer prevention. Anticancer
Agents Med Chem. 13:971–978. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhu ZY, Gao T, Huang Y, Xue J and Xie ML:
Apigenin ameliorates hypertension-induced cardiac hypertrophy and
down-regulates cardiac hypoxia inducible factor-lα in rats. Food
Funct. 7:1992–1998. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ren B, Qin W, Wu F, Wang S, Pan C, Wang L,
Zeng B, Ma S and Liang J: Apigenin and naringenin regulate glucose
and lipid metabolism, and ameliorate vascular dysfunction in type 2
diabetic rats. Eur J Pharmacol. 773:13–23. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Escande C, Nin V, Price NL, Capellini V,
Gomes AP, Barbosa MT, O'Neil L, White TA, Sinclair DA and Chini EN:
Flavonoid apigenin is an inhibitor of the NAD+ ase CD38:
Implications for cellular NAD+ metabolism, protein acetylation, and
treatment of metabolic syndrome. Diabetes. 62:1084–1093. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Poitout V and Robertson RP:
Glucolipotoxicity: Fuel excess and beta-cell dysfunction. Endocr
Rev. 29:351–366. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mayer CM and Belsham DD: Palmitate acid
attenuates insulin signaling and induces endoplasmic reticulum
stress and apoptosis in hypothalamic neurons: The rescue of
resistance and apoptosis through adenosine 5′
monophosphate-activated protein kinase activation. Endocrinology.
151:576–585. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Park JY, Kim Y, Im JA and Lee H: Oligonol
suppresses lipid accumulation and improves insulin resistance in a
palmitate-induced in HepG2 hepatocytes as a cellular steatosis
model. BMC Complement Altern Med. 15:1852015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim J, Lee I, Seo J, Jung M, Kim Y, Yim N
and Bae K: Vitexin, orientin and other flavonoids from Spirodela
polyrhiza inhibit adipogenesis in 3T3-L1 cells. Phytother Res.
24:1543–1548. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ren K, Jiang T, Zhou HF, Liang Y and Zhao
GJ: Apigenin retards atherogenesis by promoting ABCA1-mediated
cholesterol efflux and suppressing inflammation. Cell Physiol
Biochem. 47:2170–2184. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vock C, Gleissner M, Klapper M and Döring
F: Identification of palmitate-regulated genes in HepG2 cells by
applying microarray analysis. Biochim Biophys Acta. 1770:1283–1288.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zechner R, Zimmermann R, Eichmann TO,
Kohlwein SD, Haemmerle G, Lass A and Madeo F: FAT SIGNALS-lipases
and lipolysis in lipid metabolism and signaling. Cell Metab.
15:279–291. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Block MJ: American chemical society.
Chemcycolpedia. 2000:106–107. 2001.
|
|
23
|
Zhang J, Liu D, Huang Y, Gao Y and Qian S:
Biopharmaceutics classification and intestinal absorption study of
apigenin. Int J Pharm. 436:311–317. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Altamimi MA, Elzayat EM, Alshehri SM,
Mohsin K, Ibrahim MA, Al Meanazel OT, Shakeel F, Alanazi FK and
Alsarra IA: Utilizing spray drying technique to improve oral
bioavailability of apigenin. Adv Powder Technol. 29:1676–1684.
2018. View Article : Google Scholar
|
|
25
|
Zhai YJ, Guo SS, Liu CH, Yang CF, Dou JF,
Li LB and Zhai GX: Preparation and in vitro evaluation of
apigenin-loaded polymeric micelles. Colloids Surfaces A:
Physicochemical Engineering Aspects. 429:24–30. 2013. View Article : Google Scholar
|
|
26
|
Hardie DG: Minireview: The AMP-activated
protein kinase cascade: The key sensor of cellular energy status.
Endocrinology. 144:5179–5183. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Carling D: The AMP-activated protein
kinase cascade-a unifying system for energy control. Trends Biochem
Sci. 29:18–24. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kang MC, Ding Y, Kim EA, Choi YK, de
Araujo T, Heo SJ and Lee SH: Indole derivatives isolated from Brown
alga Sargassum thunbergii inhibit Adipogenesis through AMPK
activation in 3T3-L1 preadipocytes. Mar Drugs. 15:E1192017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Long YC and Zierath JR: AMP-activated
protein kinase signaling in metabolic regulation. J Clin Invest.
116:1776–1783. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ruderman N and Prentki M: AMP kinase and
malonyl-CoA: Targets for therapy of the metabolic syndrome. Nat Rev
Drug Discov. 3:340–351. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sun X, Duan X, Wang C, Liu Z, Sun P, Huo
X, Ma X, Sun H, Liu K and Meng Q: Protective effects of
glycyrrhizic acid against non-alcoholic fatty liver disease in
mice. Eur J Pharmacol. 5:75–82. 2017. View Article : Google Scholar
|
|
32
|
Frederico MJ, Vitto MF, Cesconetto PA,
Engelmann J, De Souza DR, Luz G, Pinho RA, Ropelle ER, Cintra DE
and De Souza CT: Short-term inbibition of SREBP-1c expression
reversed dietinduced nonalcoholic fatty liver disease in mice.
Scand J Gastroenterol. 46:1381–1388. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sakakura Y, Shimano H, Sone H, Takahashi
A, Inoue N, Toyoshima H, Suzuki S and Yamada N: Sterol regulatory
element-binding proteins induce an entire pathway of cholesterol
synthesis. Biochem Biophys Res Commun. 286:176–183. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Caballero F, Fernandez A, De-Lacy AM,
Fernandez-Checa JC, Caballeria J and Garcia-Ruiz C: Enhanced free
cholesterol, SREBP-2 and StAR expression in human NASH. J Hepatol.
50:789–796. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sun JH, Liu X, Cong LX, Li H, Zhang CY,
Chen JG and Wang CM: Metabolomics study of therapeutic mechanism of
Schisandra Chinensislignans in diet-induced hyperlipidemia mice.
Lipids Health Dis. 16:1452017. View Article : Google Scholar : PubMed/NCBI
|