Introduction
Ulcerative colitis (UC) is a chronic and relapsing
inflammatory intestinal disease, which is characterized by acute
pain, diarrhea and weight loss (1,2). Despite
progress in previous decades, the etiology and pathogenesis of
ulcerative colitis remain unclear (3). Multiple factors, including dysbiosis of
commensal microbiota, dysfunction of the mucosal barrier and a
defective immune response, may contribute to the etiology of UC
(4,5). At present, the morbidity of UC is
increasing annually (6). Currently,
aminosalicylic acid preparations, glucocorticoids and
immunosuppressants are the three major classes of drugs for the
clinical treatment of UC (7).
However, the side effects and sometimes serious adverse reactions
to these drugs greatly limit their use in the treatment of UC
(7,8). Therefore, more effective therapeutic
strategies for treating UC are required.
Plant-derived natural products exhibit distinctive
chemical diversity (9). Previous
studies have indicated that medicinal plant-derived extracts
including polysaccharides and flavonoids have anti-UC activity, by
regulating the levels of inflammatory mediators (10,11).
Astragaloside IV (ASI) is a monomeric compound
isolated from the traditional Chinese herb Astragalus
membranaceus (12). Previous
studies indicated that ASI possesses extensive pharmacological
activities, including regulation of intestinal microbiota, and
anti-tumor and anti-inflammatory activities (13–15). A
previous study indicated that ASI may alleviate heat-induced
inflammation by regulating endoplasmic reticulum stress (15). ASI also improves renal fibrosis
through the inhibition of the NF-κB signaling pathway (16). Kang et al (17) indicated that Wasabia japonica
prevented UC via inhibiting the NF-κB signaling pathway. In
addition, ASI may attenuate lipopolysaccharide (LPS)-induced
cardiac dysfunction and inflammation by inhibiting the levels of
tumor necrosis factor-α (TNF-α), interleukin (IL)-1β and IL-6 in
mice (18). Jiang et al
(19) identified that ASI alleviated
colonic mucosal injury in a mouse model of colitis (19). However, the specific effect of ASI on
experimental UC in mice remains unclear. Therefore, the present
study aimed to explore the role of ASI in experimental UC in
vivo and in vitro.
Materials and methods
Cell line and cell culture
Human colon fibroblast CCD-18Co cells were purchased
from The American Type Culture Collection. Cells were cultured in
Dulbecco's modified Eagle's medium (Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.),
penicillin and streptomycin (100 U/ml) in a humidified 5%
CO2 incubator at 37°C.
Cell viability detection
A Cell Counting Kit-8 (CCK-8; Beyotime Institute of
Biotechnology) assay was used to evaluate the cell viability,
according to the manufacturer's protocols. CCD-18Co cells were
seeded into a 96-well plate (5×103 cells/well) and
cultured overnight. Then, CCD-18Co cells were pretreated with ASI
(0, 10 or 50 µM) for 24 h, followed by treatment with LPS (0 or 1
µg/ml) for an additional 2 h. After 24 h incubation, 10 µl CCK-8
reagent was added to each well. The absorbance values were measured
at a wavelength of 450 nm using a microplate reader (Bio-Rad
Laboratories, Inc.). LPS was provided by Sigma-Aldrich; Merck KGaA.
ASI was purchased from MedChemExpress.
ELISA for detection of
pro-inflammatory factors
CCD-18Co cells were seeded into a 24-well plate
(2×106 cells/well) and cultured overnight. Cells were
pretreated with ASI (0, 10 or 50 µM) for 24 h, followed by
treatment with LPS (0 or 1 µg/ml) for an additional 2 h. Then,
after 24 h incubation, samples of the supernatant were collected
from each well to measure TNF-α, IL-1β and IL-6 levels by ELISA.
TNF-α (cat. no. H052), IL-1β (cat. no. H002) and IL-6 (cat. no.
H007) ELISA kits were purchased from Nanjing Jiancheng
Bioengineering Institute.
Western blot analysis
CCD-18Co cells and segments of mouse colon were
collected and homogenized using radioimmunoprecipitation assay
lysis buffer (Thermo Fisher Scientific, Inc.) supplemented with
protease and phosphatase inhibitors. A Bradford protein assay
(Beyotime Institute of Biotechnology) was used to quantify the
protein concentration. Equal amounts of proteins (30 µg) in the
lysate were then separated via 10% SDS-PAGE and transferred onto a
polyvinylidene fluoride (PVDF) membrane (Thermo Fisher Scientific,
Inc.) for 2 h. The PVDF membranes were blocked in TBST containing
5% skim milk at room temperature for 1 h, and then incubated at 4°C
with primary antibodies overnight. The primary antibodies were as
follows: Rabbit anti-NF-κB transcription factor p65 (p65; 1:1,000;
cat. no. ab16502), rabbit anti-phosphorylated NF-κB transcription
factor p65 (p-p65; 1:1,000; cat. no. ab86299), rabbit
anti-inhibitor of NF-κB (IκB; 1:1,000; cat. no. ab32518), rabbit
anti-phosphorylated inhibitor of NF-κB (p-IκB; 1:1,000; cat. no.
ab133462), rabbit anti-claudin-1 (1:1,000; cat. no. ab15098),
rabbit anti-tight junction protein ZO-1 (ZO-1; 1:1,000; cat. no.
ab96587) and anti-β-actin (1:1,000; cat. no. ab8227). The PVDF
membranes were washed 3 times with TBST, then incubated with
horseradish peroxidase-conjugated goat anti-rabbit IgG H&L
secondary antibody (1:5,000; cat. no. ab7090) at room temperature
for 1 h. All antibodies were purchased from Abcam. The protein
blots were detected with an enhanced chemiluminescence system
(Bio-Rad Laboratories, Inc.) and visualized using a Fluor Chem E
imager on a FluorChem E Imaging System 46688 (ProteinSimple). The
density of the blots for target proteins were normalized to
β-actin.
Dextran sulfate sodium (DSS)-induced
ulcerative colitis in mice
Male C57BL/6 mice (21–23 g, aged 8–10 weeks) were
provided by Shanghai SLAC Laboratory Animal Co., Ltd. The animals
were housed at a temperature of 24±1°C, a 12 h light: Dark schedule
starting at 08:00, and a humidity of 50–60% for 4 days prior to the
initiation of the experiment. The animals were provided with water
and food ad libitum.
Following acclimation, the 30 mice were randomly
divided into five experimental groups (n=6): Control (water); model
group (DSS); DSS + 50 mg/kg ASI (DSS/ASI); DSS + 200 mg/kg ASI
(DSS/ASI); and 200 mg/kg ASI treatment (ASI). The mice were orally
administered 50 or 200 mg/kg ASI once daily for 3 days (from day 0
to day 3). On day 3, UC was induced in the animals via oral
administration of 3% (w/v) DSS (ad libitum; MP Biomedicals)
in fresh drinking water for 5 days. From day 8, animals were orally
administered 50 or 200 mg/kg ASI alone until the end of the
experiment. The body weight of the mice was monitored every other
day. The disease activity index (DAI) score was determined as
described previously (20). At the
end of the experiments, the mice were sacrificed using
CO2 at a displacement rate of 20% of the chamber
volume/min (CO2 flow rate, 2.5 l/min). The length of the
colon between the ileocecal junction and the proximal rectum in the
mice was measured. The colon tissues were immediately fixed in 4%
formalin overnight at room temperature, embedded in paraffin wax
for histological analysis and stored at −80°C for other
experiments. All experimental procedures were approved by the
Ethical Committee of Suzhou Integrated Traditional Chinese and
Western Medicine Hospital. The National Institutes of Health Guide
for the Care and Use of laboratory animals was followed (21).
Histological evaluation
Colon specimens from the different groups were
embedded in paraffin and fixed with 4% formalin at room temperature
for 2 weeks, and sliced at a 5-mm thickness. These fixed tissues
were stained via hematoxylin-eosin staining, as described
previously (22). A BH22 light
microscope (Olympus Corporation; magnification, ×400) was used to
observe the histological damage and inflammation of the colon
tissues. Severity of histological inflammation was graded as
described previously (23,24): Ulceration (0, none; 1, erosion; 2,
submucosa ulceration); crypt abscesses (0, none; 1, mild; 2,
severe), degree of mononuclear cell infiltration (MNCI) (0, no
infiltration; 1, <25%; 2, 25–50%; 3, 50–75%; 4, >75%);
segmental distribution of MNCI (0, continuous; 1, mildly segmental;
2, markedly segmental); and eosinophil infiltration (0, none or
minimal; 1, mild; 2, severe).
Nitric oxide (NO) detection
The colon tissues were freeze-thawed in liquid
nitrogen and levigated. The NO concentration in the colon tissues
was detected by Griess assay (Sigma Aldrich; Merck KGaA) as
previously described (2). The data
are presented as the mean (nitrite) in mM/g colon tissue.
Measurement of cytokines by ELISA in
vivo
The TNF-α (cat. no. H052), IL-1β (cat. no. H002) and
IL-6 (cat. no. H007) content in the colon tissues of the mice was
measured with ELISA kits (Nanjing Jiancheng Bioengineering
Institute) according to the manufacturer's protocol.
Measurement of neutrophil
infiltration
The infiltration of neutrophils in the colon samples
was assessed via myeloperoxidase (MPO) activity. Colon tissues were
first homogenized in PBS (w/v 1/9), then the MPO activity assay kit
(Nanjing Jiancheng Bioengineering Institute) was used to detect the
MPO activity according to the manufacturer's protocol. The results
are presented as U/g wet tissue.
Statistical analysis
Each group was analyzed in at least three
independent experiments and all data are presented as the mean ±
standard deviation. SPSS 17.0 software (SPSS, Inc.) was used for
all statistical analyses. Comparisons among multiple groups were
performed with one-way analysis of variance followed by Dunnett's
post-hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
ASI decreases the production of TNF-α,
IL-β and IL-6 in LPS-stimulated CCD-18Co cells in vitro
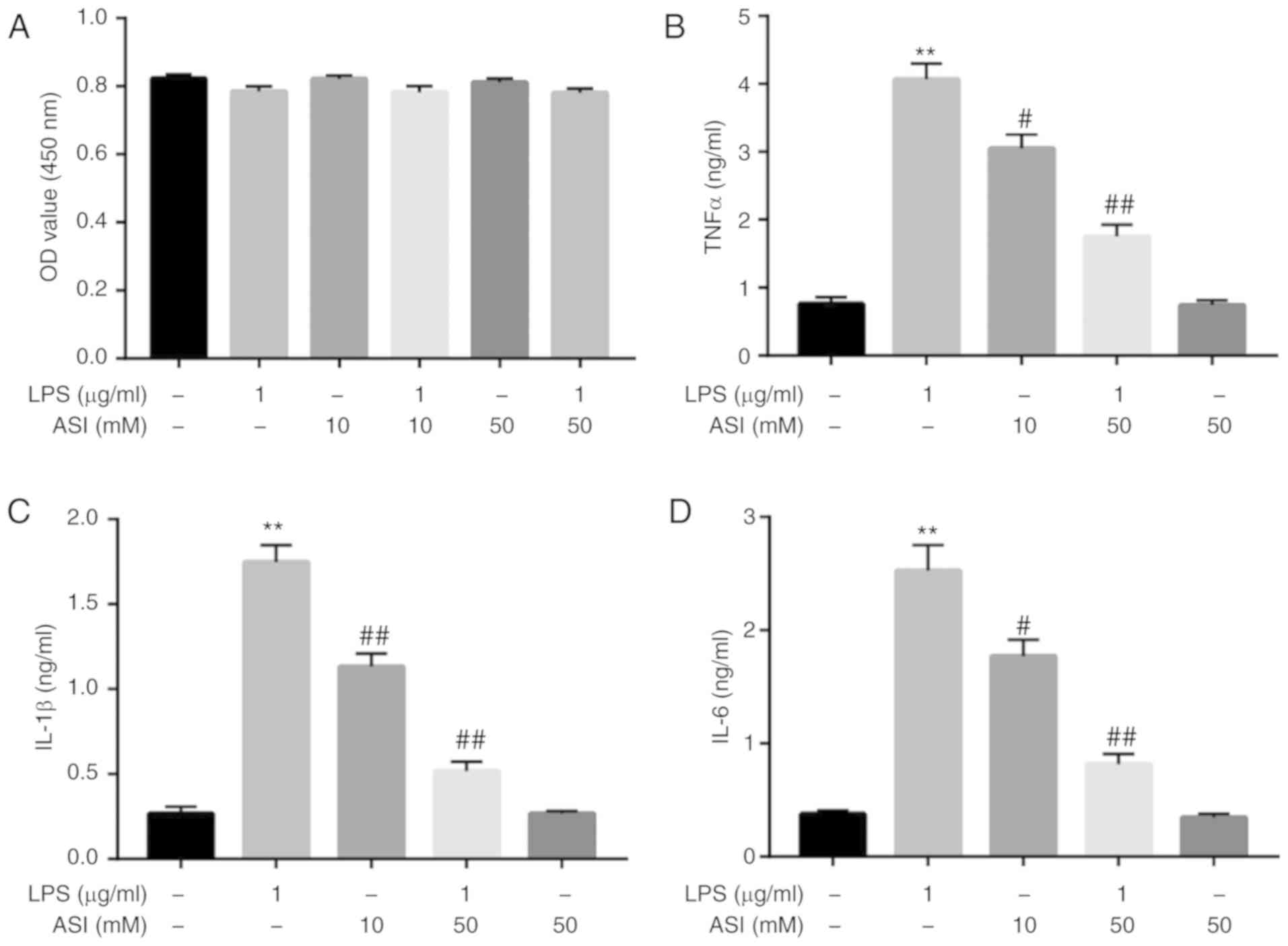
The potential cytotoxic effects of ASI and/or LPS on
CCD-18Co cells were initially assessed using a CCK-8 assay. As
indicated in Fig. 1A, neither ASI
nor LPS affected the cell viability of CCD-18Co cells. A previous
study indicated that ASI exhibited gastro-protective effects
against acute gastric lesions in rats by alleviating inflammation
(25). The anti-inflammatory action
of ASI in LPS-treated CCD-18Co cells was subsequently investigated.
As demonstrated in Fig. 1B-D,
compared with control treatment, LPS induced a significant increase
in TNF-α, IL-β and IL-6 levels in cells. By contrast, LPS-induced
TNF-α, IL-β and IL-6 upregulation were significantly reversed by 50
µM ASI. These data demonstrated that ASI may decrease the
production of TNF-α, IL-β and IL-6 in LPS-stimulated CCD-18Co cells
in vitro.
ASI inhibits the phosphorylation of
NF-κB p65 and IκB in LPS-treated CCD-18Co cells in vitro
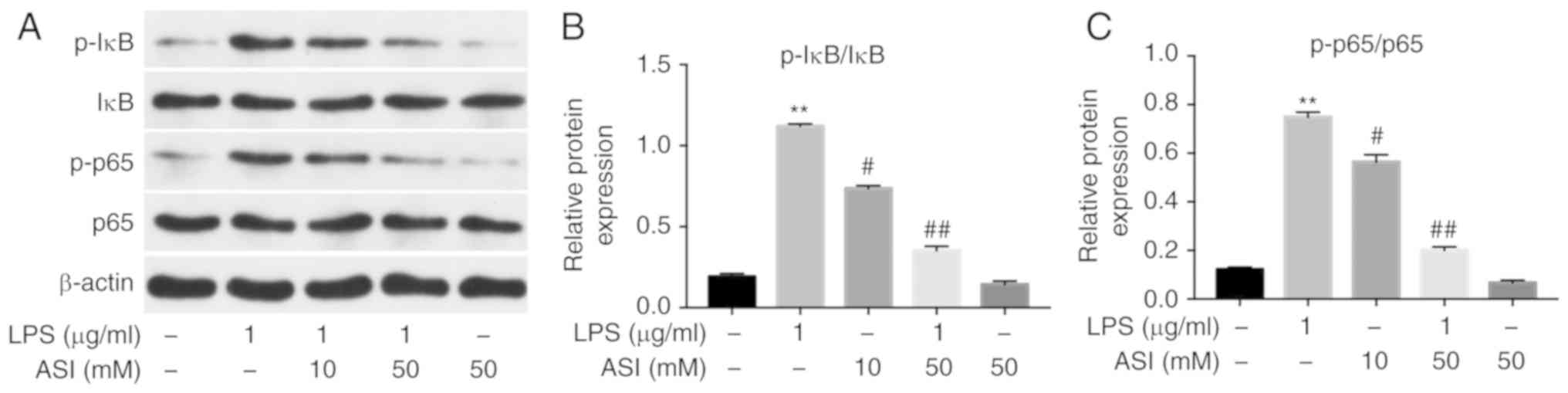
It has been suggested that NF-κB serves as a key
factor in the mediation of inflammation, and phosphorylated NF-κB
p65 (p-p65) serves a vital role in the activation of NF-κB
(26). Therefore, the present study
investigated whether ASI exerted anti-inflammatory effects on
CCD-18Co cells via the inhibition of NF-κB activation. As indicated
in Fig. 2, the expression of p-p65
and p-IκB in cells was significantly increased by LPS. By contrast,
the LPS-induced increases in p-p65 and p-IκB protein levels were
notably reduced by 50 µM ASI. These data demonstrated that ASI
inhibited the phosphorylation of NF-κB p65 and IκB in LPS-treated
CCD-18Co cells in vitro.
ASI attenuates DSS-induced UC in
mice
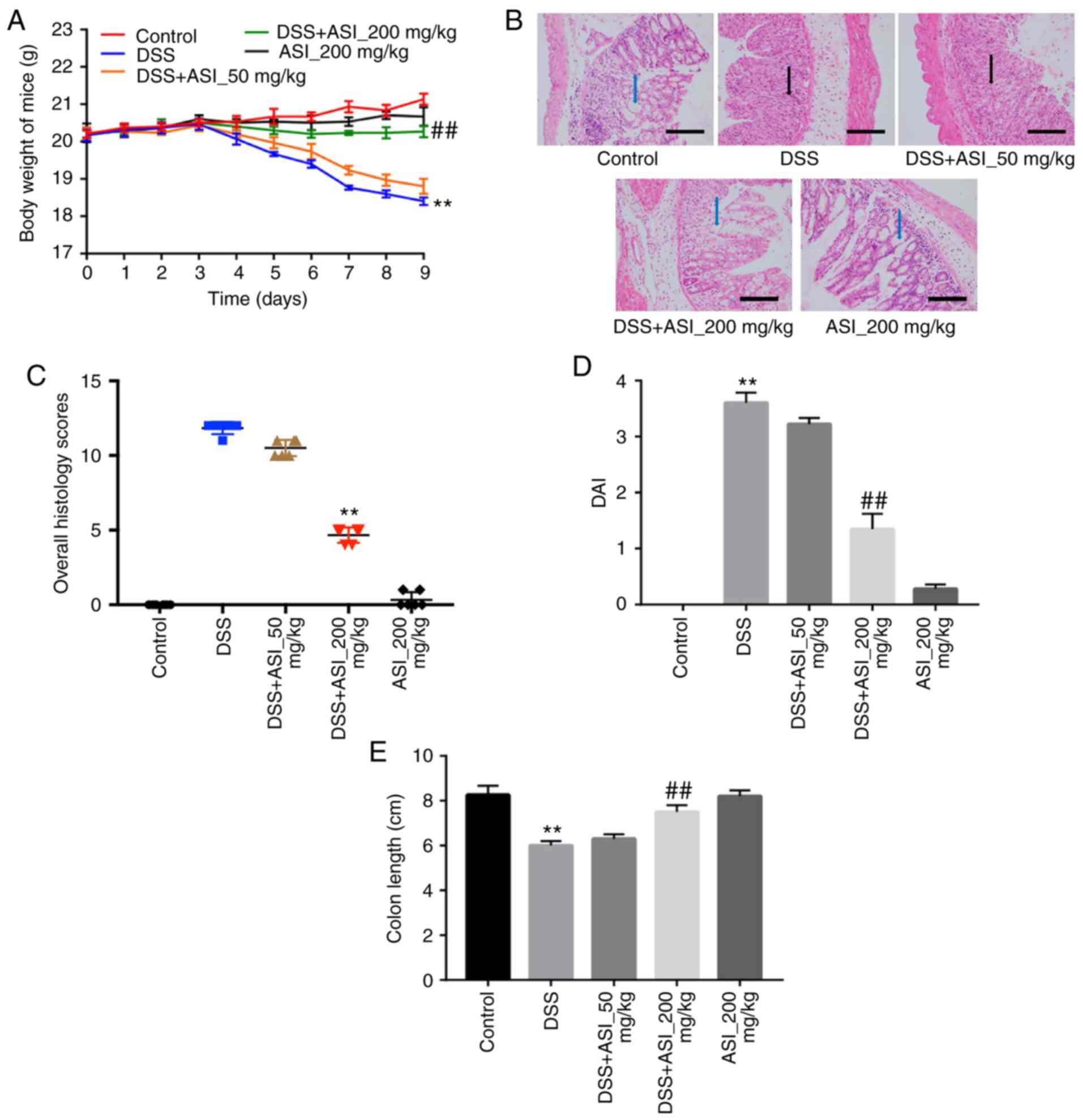
In order to confirm the anti-inflammatory effects of
ASI in vivo, a DSS-induced mouse UC model was established.
As indicated in Fig. 3A, compared
with the control group, the body weights of the mice were markedly
decreased in the DSS group. However, ASI (200 mg/kg) notably
prevented body weight loss during the progression of experimental
UC in mice (Fig. 3A). In addition,
H&E staining was used to observe the infiltration of
inflammatory cells in sections of mouse colon segments. As
indicated in Fig. 3B, colons in the
control group were healthy, exhibiting intact surface epithelia and
submucosa. However, DSS resulted in distortion of the crypt
epithelium, and ulceration and damage to the surface epithelium.
Compared with the DSS group, colons in the ASI (200 mg/kg) group
exhibited well-preserved intact surface epithelia and cryptal
glands (Fig. 3B). The histological
damage scores in the ASI (200 mg/kg) group were significantly
decreased compared with the DSS group (Fig. 3C). In addition, 200 mg/kg ASI
significantly decreased the DAI during the progression of
experimental UC in mice, which reflects the health status of the
mice (Fig. 3D). Furthermore, ASI
(200 mg/kg) significantly alleviated colon shortening caused by DSS
in mice (Fig. 3E). All these data
demonstrated that ASI attenuated UC induced by DSS in
vivo.
ASI alleviates inflammatory responses
in the colon in DSS-treated mice
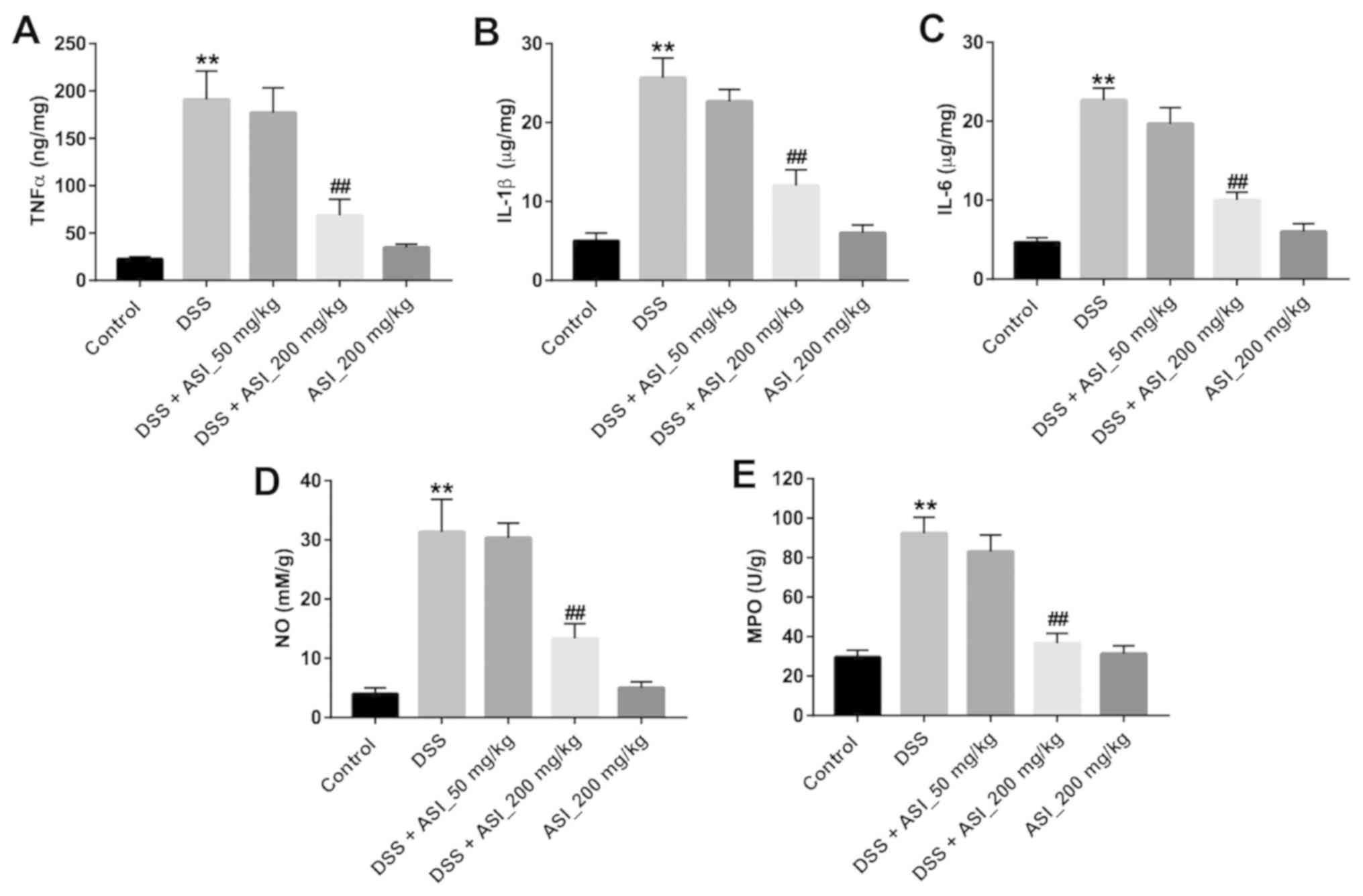
Next, the concentrations of TNF-α, IL-1β and IL-6 in
colon tissues were detected in vivo. As indicated in
Fig. 4A-C, the pro-inflammatory
cytokines TNF-α, IL-1β and IL-6 in the colon tissues were
significantly upregulated in the DSS group, compared with the
control group (P<0.01). However, ASI (200 mg/kg) treatment
markedly suppressed the DSS-induced production of these cytokines
(Fig. 4A-C). Similar to the
pro-inflammatory cytokines, the results for NO and the MPO activity
assay indicated that the levels of NO and MPO in the colon tissues
were significantly upregulated in the DSS group. Nevertheless, ASI
(200 mg/kg) treatment alleviated the DSS-induced production of NO
and MPO in the colon (Fig. 4D and
E). These data demonstrated that ASI may alleviate inflammatory
responses in the colon in the DSS-induced UC mouse model.
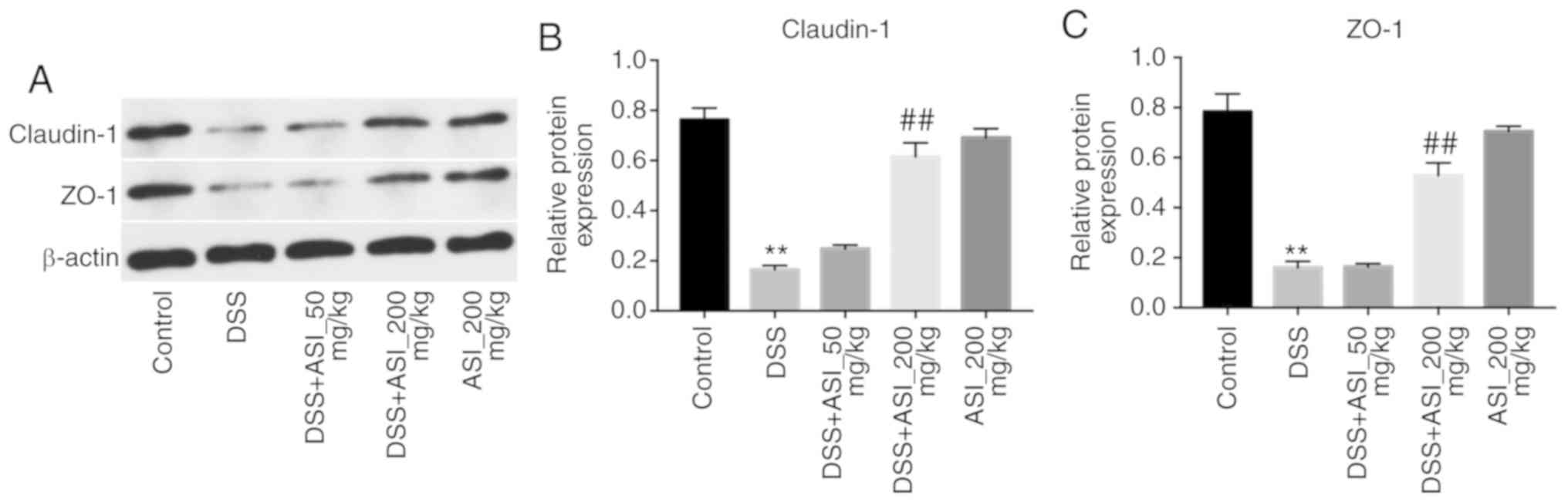
ASI increases the expression of tight
junction (TJ) proteins in colonic tissues
It has been demonstrated that UC is associated with
increased intestinal penetrability and decreased TJ protein
expression, including that of ZO-1 and claudin-1 (27). As presented in Fig. 5, the expression levels of claudin-1
and ZO-1 were markedly decreased in the DSS group. However,
DSS-induced claudin-1 and ZO-1 downregulation was notably reversed
by 200 mg/kg ASI (Fig. 5). These
data demonstrated that ASI increased the expression levels of TJ
proteins in colonic tissues.
Discussion
Several compounds isolated from traditional Chinese
medicines have been identified to have therapeutic effects in UC
(2,28). However, whether ASI has anti-UC
activity remains unknown. Studies addressing this question has been
very limited to date, to the best of our knowledge. The results of
the present study indicated that ASI reduced the production of
inflammatory cytokines and proteins in LPS-stimulated CCD-18Co
cells in vitro. In addition, ASI alleviated the inflammatory
response in the colon in DSS-induced UC in vivo. These data
highlighted that ASI may exert its anti-inflammatory action on
experimental UC in vitro and in vivo. To the best of
our knowledge, this is the first study of the effects of ASI on
experimental UC in vitro and in vivo.
The novel data from the present study is that ASI
reversed LPS-induced TNF-α, IL-β and IL-6 inflammatory cytokine
increases via the inhibition of the NF-κB pathway in vitro.
TNF-α, IL-1β and IL-6 are pro-inflammatory cytokines associated
with both colorectal and colitis-associated disease (29). Once these pro-inflammatory cell
factors are delivered from immune cells, they increase inflammation
and lead to mucosal tissue injury (26). Biologically, the inflammatory process
is strongly mediated by NF-κB, a critical transcription factor for
various pro-inflammatory cytokines including TNF-α, IL-β and IL-6.
NF-κB serves as an inactive heterotrimer, including p50, p65 and
IκB subunits in the cytoplasm (30).
A recent study indicated that ASI ameliorated LPS-induced
neuroinflammation in mice by decreasing the levels of TNF-α and
IL-1β (31). Xu et al
(32) identified that ASI suppressed
synoviocytes and collagen-induced arthritis by inhibiting the
levels of TNF-α, IL-6 and IL-1β. Collectively, these results
demonstrate that ASI has a potent inhibitory effect on
NF-κB-mediated cytokine expression.
The DSS-induced UC model may be used to examine the
pathogenesis of UC in humans, due to its similarity with the
natural history of the disease in humans (1). It is well-known that weight loss and
DAI in mice are commonly used to evaluate severity of UC (1). The results of the present study
demonstrated that ASI (200 mg/kg) markedly reversed DSS-induced
colonic shortening and weight loss in UC model mice, which
indicated an attenuation of clinical UC symptoms. Consistently,
Zhang et al (1) indicated
that Zanthoxylum bungeanum pericarp extract alleviates
DSS-induced colonic shortening and weight loss in an experimental
colitis model in vivo. These results demonstrated that ASI
(200 mg/kg) significantly alleviated the inflammatory response in
the colon in the DSS-induced UC mouse model.
Importantly, MPO released from inflamed enteric
mucosa and bowel tissues also serves a vital role in local bowel
injury (33). In addition, NO is one
of the primary sources of free radicals, which may additionally
cause oxidative damage and intestinal lesions (2). The present study also examined the
effect of ASI on MPO and NO production in colonic tissues. ASI (200
mg/kg) significantly inhibited the generation of MPO compared with
the model group. Meanwhile, ASI (200 mg/kg) markedly decreased NO
production in the colon tissues of UC mice compared with the model
group. Kannan et al (34)
demonstrated that Bauhinia tomentosa attenuated acetic
acid-induced UC by regulating the levels of MPO and NO. Han et
al (35) also identified that
Xiexin decoction improves the inflammation associated with colitis
and inhibits colonic inflammatory damage by decreasing the level of
MPO in UC rats. The data from the present study were consistent
with these results.
Clinical data has indicated that pro-inflammatory
cytokines, including IL-6, TNF-α and IL-1β, also perform important
roles in UC pathogenesis in vivo (36). Jiang et al (19) demonstrated that ASI alleviated
2,4,6-trinitrobenzene sulfonic acid-induced inflammation in
inflammatory bowel disease. Based on these aforementioned results,
the present study investigated whether ASI had an effect on these
pro-inflammatory factors during UC. The present results indicated
that ASI (200 mg/kg) significantly downregulated the production of
TNF-α, IL-1β and IL-6 in colon tissues compared with the model
group. These data suggested that ASI may alleviate inflammation of
the colonic tissues through downregulation of pro-inflammatory
cytokines.
TJ proteins, including ZO-1 and claudin-1, form
physiologically active regions in colonic tissues that may become
disorganized in UC (27). For
example, a previous study revealed that levels of key TJ proteins
ZO-1 and claudin-1 were decreased in UC experimental models
(37). Consistently, the present
results indicated that ASI upregulated the expression of claudin-1
and ZO-1 proteins in DSS-treated mice.
The results of the present study were limited in
that the therapeutic strategy was not adjusted according to the
disease stage of UC in the animal studies. Future studies will be
required to explore the effect of ASI on UC in different disease
stages.
In summary, ASI was effective in ameliorating
experimental UC in vitro and in vivo via the
inhibition of inflammatory molecule production and the
downregulation of NF-κB signaling. These results suggest that ASI
may serve as a potential therapeutic agent for the treatment of
UC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SW and ZC analyzed and interpreted the experimental
data, and were major contributors to the development of the first
draft of the present manuscript. ZC reviewed and approved the final
draft of the manuscript prior to submission.
Ethics approval and consent to
participate
Ethics approval for the present study was approved
by the Ethical Committee of Suzhou Integrated Traditional Chinese
and Western Medicine Hospital (Suzhou, China).
Patient consent for publication
Not applicable.
Conflicts of interest
The authors declare that they have no competing
interests.
References
|
1
|
Zhang Z, Liu J, Shen P, Cao Y, Lu X, Gao
X, Fu Y, Liu B and Zhang N: Zanthoxylum bungeanum pericarp
extract prevents dextran sulfate sodium-induced experimental
colitis in mice via the regulation of TLR4 and TLR4-related
signaling pathways. Int Immunopharmacol. 41:127–135. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang H, Deng A, Zhang Z, Yu Z, Liu Y,
Peng S, Wu L, Qin H and Wang W: The protective effect of
epicatechin on experimental ulcerative colitis in mice is mediated
by increasing antioxidation and by the inhibition of NF-κB pathway.
Pharmacol Rep. 68:514–520. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Harris KG and Chang EB: The intestinal
microbiota in the pathogenesis of inflammatory bowel diseases: New
insights into complex disease. Clin Sci (Lond). 132:2013–2028.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yadav V, Varum F, Bravo R, Furrer E, Bojic
D and Basit AW: Inflammatory bowel disease: Exploring gut
pathophysiology for novel therapeutic targets. Transl Res.
176:38–68. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ordás I, Eckmann L, Talamini M, Baumgart
DC and Sandborn WJ: Ulcerative colitis. Lancet. 380:1606–1619.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thia KT, Loftus EV Jr, Sandborn WJ and
Yang SK: An update on the epidemiology of inflammatory bowel
disease in Asia. Am J Gastroenterol. 103:3167–3182. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peyrin-Biroulet L, Deltenre P, Ardizzone
S, D'Haens G, Hanauer SB, Herfarth H, Lémann M and Colombel JF:
Azathioprine and 6-mercaptopurine for the prevention of
postoperative recurrence in Crohn's disease: A meta-analysis. Am J
Gastroenterol. 104:2089–2096. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kemp R, Dunn E and Schultz M:
Immunomodulators in inflammatory bowel disease: An emerging role
for biologic agents. BioDrugs. 27:585–590. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Atanasov AG, Waltenberger B,
Pferschy-Wenzig EM, Linder T, Wawrosch C, Uhrin P, Temml V, Wang L,
Schwaiger S, Heiss EH, et al: Discovery and resupply of
pharmacologically active plant-derived natural products: A review.
Biotechnol Adv. 33:1582–1614. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maria-Ferreira D, Nascimento AM, Cipriani
TR, Santana-Filho AP, Watanabe PDS, Sant Ana DMG, Luciano FB,
Bocate KCP, van den Wijngaard RM, Werner MFP and Baggio CH:
Rhamnogalacturonan, a chemically-defined polysaccharide, improves
intestinal barrier function in DSS-induced colitis in mice and
human Caco-2 cells. Sci Rep. 8:122612018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun Y, Zhao Y, Yao J, Zhao L, Wu Z, Wang
Y, Pan D, Miao H, Guo Q and Lu N: Wogonoside protects against
dextran sulfate sodium-induced experimental colitis in mice by
inhibiting NF-κB and NLRP3 inflammasome activation. Biochem
Pharmacol. 94:142–154. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang L, Xing F, Han X, Li Q, Wu H, Shi H,
Wang Z, Huang F and Wu X: Astragaloside IV regulates
differentiation and induces apoptosis of activated CD4+ T cells in
the pathogenesis of experimental autoimmune encephalomyelitis.
Toxicol Appl Pharmacol. 362:105–115. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xu N, Kan P, Yao X, Yang P, Wang J, Xiang
L and Zhu Y: Astragaloside IV reversed the autophagy and oxidative
stress induced by the intestinal microbiota of AIS in mice. J
Microbiol. 56:838–846. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dai PC, Liu DL, Zhang L, Ye J, Wang Q,
Zhang HW, Lin XH and Lai GX: Astragaloside IV sensitizes non-small
cell lung cancer cells to gefitinib potentially via regulation of
SIRT6. Tumour Biol. 39:10104283176975552017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dong Z, Zhou J, Zhang Y, Chen Y, Yang Z,
Huang G, Chen Y, Yuan Z, Peng Y and Cao T: Astragaloside-IV
alleviates heat-induced inflammation by inhibiting endoplasmic
reticulum stress and autophagy. Cell Physiol Biochem. 42:824–837.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou X, Sun X, Gong X, Yang Y, Chen C,
Shan G and Yao Q: Astragaloside IV from Astragalus
membranaceus ameliorates renal interstitial fibrosis by
inhibiting inflammation via TLR4/NF-кB in vivo and in vitro. Int
Immunopharmacol. 42:18–24. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kang JH, Choi S, Jang JE, Ramalingam P, Ko
YT, Kim SY and Oh SH: Wasabia japonica is a potential
functional food to prevent colitis via inhibiting the NF-κB
signaling pathway. Food Funct. 8:2865–2874. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao P, Wang Y, Zeng S, Lu J, Jiang TM and
Li YM: Protective effect of astragaloside IV on
lipopolysaccharide-induced cardiac dysfunction via downregulation
of inflammatory signaling in mice. Immunopharmacol Immunotoxicol.
37:428–433. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jiang XG, Sun K, Liu YY, Yan L, Wang MX,
Fan JY, Mu HN, Li C, Chen YY, Wang CS and Han JY: Astragaloside IV
ameliorates 2,4,6-trinitrobenzene sulfonic acid (TNBS)-induced
colitis implicating regulation of energy metabolism. Sci Rep.
7:418322017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
He X, Zheng Z, Yang X, Lu Y, Chen N and
Chen W: Tetramethylpyrazine attenuates PPAR-γ
antagonist-deteriorated oxazolone-induced colitis in mice. Mol Med
Rep. 5:645–650. 2012.PubMed/NCBI
|
|
21
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . Guide for the Care and Use of Laboratory Animals. 8th.
National Academies Press (US); Washington, DC: 2011
|
|
22
|
Zhou Q, Gong X, Kuang G, Jiang R, Xie T,
Tie H, Chen X, Li K, Wan J and Wang B: Ferulic acid protected from
kidney ischemia reperfusion injury in mice: Possible mechanism
through increasing adenosine generation via HIF-1a. Inflammation.
41:2068–2078. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zheng B, van Bergenhenegouwen J, Overbeek
S, van de Kant HJ, Garssen J, Folkerts G, Vos P, Morgan ME and
Kraneveld AD: Bifidobacterium breve attenuates murine dextran
sodium sulfate-induced colitis and increases regulatory T cell
responses. PLoS One. 9:e954412014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Araki T, Hashimoto K, Okita Y, Fujikawa H,
Kondo S, Kobayashi M, Ohi M, Toiyama Y, Inoue Y, Uchida K, et al:
Colonic histological criteria predict development of pouchitis
after Ileal Pouch: Anal anastomosis for patients with ulcerative
colitis. Dig Surg. 35:138–143. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mao S, Yang G, Li W, Zhang J, Liang H, Li
J and Zhang M: Gastroprotective effects of Astragaloside IV against
acute gastric lesion in rats. PLoS One. 11:e01481462016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ge H, Tang H, Liang Y, Wu J, Yang Q, Zeng
L and Ma Z: Rhein attenuates inflammation through inhibition of
NF-κB and NALP3 inflammasome in vivo and in vitro. Drug Des Devel
Ther. 11:1663–1671. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim Y, Wu AG, Jaja-Chimedza A, Graf BL,
Waterman C, Verzi MP and Raskin I: Isothiocyanate-enriched moringa
seed extract alleviates ulcerative colitis symptoms in mice. PLoS
One. 12:e01847092017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Qu C, Yuan ZW, Yu XT, Huang YF, Yang GH,
Chen JN, Lai XP, Su ZR, Zeng HF, Xie Y and Zhang XJ: Patchouli
alcohol ameliorates dextran sodium sulfate-induced experimental
colitis and suppresses tryptophan catabolism. Pharmacol Res.
121:70–82. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Venancio VP, Cipriano PA, Kim H, Antunes
LM, Talcott ST and Mertens-Talcott SU: Cocoplum (Chrysobalanus
icaco L.) anthocyanins exert anti-inflammatory activity in
human colon cancer and non-malignant colon cells. Food Funct.
8:307–314. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Buss H, Handschick K, Jurrmann N, Pekkonen
P, Beuerlein K, Muller H, Wait R, Saklatvala J, Ojala PM, Schmitz
ML, et al: Cyclin-dependent kinase 6 phosphorylates NF-κB P65 at
serine 536 and contributes to the regulation of inflammatory gene
expression. PLoS One. 7:e518472012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Song MT, Ruan J, Zhang RY, Deng J, Ma ZQ
and Ma SP: Astragaloside IV ameliorates neuroinflammation-induced
depressive-like behaviors in mice via the PPARγ/NF-κB/NLRP3
inflammasome axis. Acta Pharmacol Sin. 39:1559–1570. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xu H, Wang CY, Zhang HN, Lv CY and Wang
YZ: Astragaloside IV suppresses inflammatory mediator production in
synoviocytes and collagen-induced arthritic rats. Mol Med Rep.
13:3289–3296. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chiu CT, Kuo SN, Hung SW and Yang CY:
Combined treatment with hyaluronic acid and mesalamine protects
rats from inflammatory bowel disease induced by intracolonic
administration of trinitrobenzenesulfonic acid. Molecules. 22(pii):
E9042017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kannan N and Guruvayoorappan C: Protective
effect of Bauhinia tomentosa on acetic acid induced
ulcerative colitis by regulating antioxidant and inflammatory
mediators. Int Immunopharmacol. 16:57–66. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Han XH, Zhong J, Guo JY, Shi R, Wang XH,
Wang CH, Wang K, Du GL, Shen YH and Ma YM: Relationships between
pharmacokinetics and efficacy of Xie-xin decoction in rats with
experimental ulcerative colitis. J Ethnopharmacol. 148:182–189.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Neurath MF: Cytokines in inflammatory
bowel disease. Nat Rev Immunol. 14:329–342. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li Q, Zhang Q, Zhang M, Wang C, Zhu Z, Li
N and Li J: Effect of n-3 polyunsaturated fatty acids on membrane
microdomain localization of tight junction proteins in experimental
colitis. FEBS J. 275:411–420. 2008. View Article : Google Scholar : PubMed/NCBI
|