Introduction
Breast cancer exhibits the highest incidence and
mortality rates among women worldwide. According to a statistical
analysis of cancer cases in 2012 (1), the number of new breast cancer cases
and cases of mortality are ~1,700,000 and 521,900, respectively.
Breast cancer accounts for 25% of all cancer cases and 15% of
cancer-associated cases of mortality (2).
Gemcitabine (2′,2′-difluoro-deoxycytidine; dFdC) is
a nucleoside analog that prevents DNA replication and transcription
leading to apoptosis (3).
Preliminary clinical observation indicated that the effectiveness
of gemcitabine in the treatment of advanced breast cancer was
14–27% (4,5). Advanced breast cancer chemotherapeutic
drugs recommended by the Chinese breast cancer diagnosis and
treatment guide (2017) include gemcitabine (6). Gemcitabine has good therapeutic effects
in a variety of patients with cancer, including the elderly
(7). However, gemcitabine resistance
is common, and the underlying molecular mechanisms remain to be
elucidated (8).
The endoplasmic reticulum (ER) protein 29 (ERp29) is
a member of the ER chaperones that does not contain an active-site
double-cysteine motif with oxidoreductase activity, observed in
other ER chaperones (9). The
structural variation of ERp29 suggests a varying function in cells,
particularly cancer cells (9).
Recent studies demonstrated that ERp29 overexpression resulted in
G0/G1 arrest in MDA-MB-231 cells, leading to a marked delay in
tumorigenesis onset in vivo (10). ERp29 upregulated heat shock protein
27 (HSP27) expression in cancer cells (11,12) and
attenuated apoptosis caused by chemotherapy drugs (13). Another study revealed that ERp29
served a key role in the development of gemcitabine resistance in
cancer cells (14).
microRNAs (miRNAs or miRs) are small non-coding RNA
species containing 22–25 ribonucleic acids, which are major
elements of gene expression control, causing translation inhibition
or mRNA degradation (15,16). It is widely reported that miRNAs
serve important roles in tumorigenesis (17,18).
miR-205-5p is a highly conserved miRNA that is expressed in mammary
gland progenitor cells and in stratified squamous
epithelium-derived tissues (19,20). A
previous study suggested that miR-205-5p is downregulated in breast
cancer (21,22). As one of the most researched miRNAs
in breast cancer, it is involved in cellular differentiation,
migration and proliferation (18,19).
However, the role of miR-205-5p in the regulation of gemcitabine
sensitivity has not been elucidated in breast cancer.
In this study, miR-205-5p expression was
downregulated and cell viability was decreased in a
gemcitabine-resistant breast cancer cell line (MDA-MB-231/GEM)
compared with the parental cell line (MDA-MB-231/P). miR-205-5p
bound to the 3′ untranslated region (3′-UTR) of ERp29 directly and
negatively regulated its expression. Taken together, this study
revealed that miR-205-5p may mediate gemcitabine resistance in
breast cancer by targeting ERp29.
Materials and methods
Clinical tissue samples
A total of 25 breast cancer and matched adjacent
non-tumor tissue samples were collected at the Tianjin Baodi
People's Hospital between July 2014 and February 2017 (Tianjin,
China). All tumor tissue samples were obtained from women aged
30–65 years (mean age, 46 years). Tissue samples were immediately
frozen in liquid nitrogen after surgery and stored at −80°C until
use. The present study was approved by the Ethics Committee of
Tianjin Baodi People's Hospital. Written informed consent was
provided by all participants.
Cell culture and reagents
MDA-MB-231 and BT549 human breast cancer cell lines
were purchased from the American Type Culture Collection (ATCC).
MDA-MB-231 and BT549 cells were cultured in Dulbecco's modified
Eagle's medium (DMEM; Thermo Fisher Scientific, Inc.) supplemented
with 10% fetal bovine serum (HyClone; GE Healthcare Life Sciences)
and 1% penicillin-streptomycin solution (Thermo Fisher Scientific,
Inc.) in a humidified atmosphere containing 5% CO2 at
37°C. MDA-MB-231/GEM cells were established through continuous
exposure of MDA-MB-231 cells to increasing concentrations of
gemcitabine (initial concentration, 12 nM; increased to 6 µM over 6
months; Sigma-Aldrich; Merck KGaA) at 37°C. After 6 months,
gemcitabine resistance was confirmed by comparison of cell
viability between MDA-MB-231/GEM and MDA-MB-231 cells in response
to 6 µM gemcitabine. The immortal breast cell line MCF10A was
purchased from ATCC and cultured in DMEM/F12 media with 10% horse
serum, 20 ng/ml epidermal growth factor, 100 ng/ml cholera toxin,
0.01 mg/ml insulin, 500 ng/ml hydrocortisone (all from HyClone; GE
Healthcare Life Sciences) and 1% penicillin-streptomycin solution
in a humidified atmosphere containing 5% CO2 at
37°C.
Overexpression and downregulation of
miR-205-5p
miR-205-5p mimic, miR-negative control (NC) mimic,
miR-205-5p inhibitor and miR-NC inhibitor were purchased from
Shanghai GenePharma Co., Ltd. The sequences were: miR-205-5p mimic,
5′-UCCUUCAUUCCACCGGAGUCUG-3′; miR-NC mimic,
5′-UCGCUUGGUGCAGGUCGGGAA-3′; miR-205-5p inhibitor,
5′-CAGACUCCGGUGGAAUGAAGGA-3′; miR-NC inhibitor,
5′-CAGUACUUUUGUGUAGUACAA-3′. A total of 50 nM miRNA was transfected
into MDA-MB-231 and BT549 cells using Lipofectamine®
2000 reagent (Thermo Fisher Scientific, Inc.) according to
manufacturer's protocol for 48 h at 37°C. Cells were then subjected
to the following experiments.
Overexpression of ERp29
The full-length opening reading frame of ERp29 was
amplified from MCF10A cells, non-tumorigenic epithelial cell line
expressing the normal sequence for ERp29 and ligated into the
pcDNA3.1 plasmid (Invitrogen; Thermo Fisher Scientific, Inc.) by
the restriction sites of HindIII and XhoI using the
following primer sequences: ERp29 forward,
5′-AAGCTTACTATCGCTTACCTA-3′ and reverse,
5′-CTCGAGTGTTGGCACAAGTGCT-3′. For the overexpression of ERp29, 2 µg
pcDNA3.1-ERp29 was transfected into cells using the Lipofectamine
2000 reagent (Thermo Fisher Scientific, Inc.) according to
manufacturer's protocol and incubated for 48 h at 37°C. Cells were
then subjected to subsequent experiments.
Cell viability and apoptosis
assays
Cell viability assays were performed using the Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.)
according to the manufacturer's instruction. A total of
5×103 cells/well were seeded into 96-well plates. After
treatment of MDA-MB-231, BT549 and MDA-MB-231/GEM cells with
different concentrations of gemcitabine (10, 20, 40, 80 and 160 nM)
for 24 h at 37°C, cells were transfected with miRNA and/or cDNA
using procedures described as follows: MDA-MB-231 cells and BT549
cells were transfected with miR-205-5p mimic or miR-NC mimic; and
MDA-MB-231/GEM cells were co-transfected with miR-205-5p mimic or
miR-NC mimic and pcDNA3.1-ERp29 or pcDNA3.1 plasmid. Following 48 h
incubation, 10 µl CCK-8 solution was added into each well. Cells
were incubated for 2 h at 37°C and the absorbance was measured at a
wavelength of 450 nm.
Cell apoptosis assays were performed using the
Annexin-V/Dead Cell Apoptosis kit (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. After
treatment with gemcitabine (40 nM) or corresponding concentrations
of dimethylsulfoxide (DMSO) followed by co-transfection with
miR-205-5p mimic or miR-NC mimic and pcDNA3.1-ERp29 or pcDNA3.1
plasmid for 48 h, MDA-MB-231/GEM cells were harvested and washed in
cold PBS. Cells were then diluted in 1X Annexin-binding buffer to
1×106 cells/ml; 100-µl samples per assay were prepared.
Alexa Fluor 488 annexin V (5 µl) and propidium iodide working
solution (1 µl; 100 µg/ml) were added to the 100-µl cell
suspensions. Cells were incubated at room temperature for 15 min
and annexin-binding buffer (400 µl) was added. Subsequently,
stained cells were analyzed using a BD FACSCalibur flow cytometer
(BD Biosciences). The results were quantified using Cell Quest
software (version 5.1; BD Biosciences).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) for
miRNA and mRNA assays
Total RNA was extracted from cultured cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and cDNA was synthetized using PrimeScript™ RT reagent kit
(Takara Bio, Inc.) according to the manufacturer's protocol.
RT-qPCR reactions were performed in triplicate using
SYBR® Premix Ex Taq (Takara Bio, Inc.) in a Bio-Rad
CFX96 Real-Time PCR system (Bio-Rad Laboratories, Inc.). The
thermocycling conditions used for qPCR were described as follows:
Initial denaturation at 95°C for 30 sec, followed by 40 cycles of
95°C for 5 sec and 60°C for 30 sec. Levels of miR-205-5p and ERp29
were normalized to GAPDH and U6, respectively. The
2−ΔΔCq method was used to quantify gene expression
levels (23). The primer sequences
used in the present study were listed as follows: miR-205-5p
forward, 5′-TCCTTCATTCCACCGGAGTCTG-3′ and reverse,
5′-GCGAGCACAGAATTAATACGAC-3′; U6 forward,
5′-ATTGGAACGATACAGAGAAGATT-3′ and reverse,
5′-GGAACGCTTCACGAATTTG-3′; ERp29 forward,
5′-AAAGCAAGTTCGTCTTGGTGA-3′ and reverse,
5′-CGCCATAGTCTGAGATCCCCA-3′; GAPDH forward,
5′-TTGGTATCGTGGAAGGACTCA-3′ and reverse,
5′-TGTCATCATATTTGGCAGGTT-3′.
Western blot analysis
ERp29 antibody (cat. no. 37555; 1:1,000) and HSP27
(cat. no., 41043; 1:1,000) were purchased from Signalway Antibody
LLC, and GAPDH mouse monoclonal antibody (cat. no. ab8245;
1:10,000) was obtained from Abcam. Anti-mouse (cat. no. CW0221S;
1:10,000) and anti-rabbit (cat. no. CW0234S; 1:10,000) secondary
antibodies were provided by CWBiotech. Western blotting was
performed as follows: Harvested cells were washed twice with cold
PBS and incubated with a cold RIPA buffer (Beyotime Institute of
Biotechnology) containing protease inhibitor cocktail
(Sigma-Aldrich; Merck KGaA) for lysis on ice for 30 min.
Subsequently, lysates were centrifuged at 12,000 × g for 15 min at
4°C. A bicinchoninic acid protein assay kit (Thermo Fisher
Scientific, Inc.) was used to determine the protein concentration
in the supernatant. Equal amounts of protein (20 µg/well) were
separated by SDS-PAGE (8% gel), transferred to polyvinylidene
fluoride membranes (EMD Millipore; Merck KGaA) and incubated with
the aforementioned primary antibodies at 4°C overnight and
corresponding secondary antibodies at room temperature for 2 h.
Blots were developed using SuperSignal West Femto Maximum
Sensitivity substrate (Thermo Fisher Scientific, Inc.) and images
were recorded using ImageQuant LAS 4000 (GE Healthcare Life
Sciences). Densitometry was quantified with ImageJ software
(version. 1.8.0; National Institutes of Health).
Dual-luciferase reporter gene
assays
Using the TargetScan (release 7.1, www.targetscan.org) miRNA target prediction database,
a putative binding site of miR-205-5p was predicted within the
3′-UTR of ERp29. The ERp29 3′-UTR region was amplified using cDNA
from MCF10A cells and cloned into the pGL3 vector [wild-type
(WT)-ERp29-3′-UTR; Promega Corporation]. Two site mutations were
introduced to WT-ERp29-3′-UTR to construct the mutant (Mut)
ERp29-3′-UTR. MDA-MB-231 cells were co-transfected with 100 nM of
WT-ERp29-3′-UTR or Mut-ERp29-3′-UTR plasmid and 100 nM of
miR-205-5p mimic or negative control using Lipofectamine 2000
reagent. Luciferase activity was evaluated 48 h following
transfection using the Dual-Glo Luciferase assay system (Promega
Corporation), with all luciferase activity normalized to that of
Renilla luciferase activity.
Statistical analysis
All experiments were performed in triplicate. Data
were analyzed using GraphPad Prism 5.0 (GraphPad Software, Inc.)
and are presented as the mean ± standard deviation. Paired
Student's t-test was used to analyze paired samples from patients
with breast cancer. Unpaired Student's t-test was used to analyze
differences between two independent groups. Pearson's correlation
analysis was used to analyze the correlation between miR-205-5p and
ERp29 mRNA expression levels in tumor tissues. One-way analysis of
variance followed by Newman Keuls test was performed for
comparisons among multiple groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
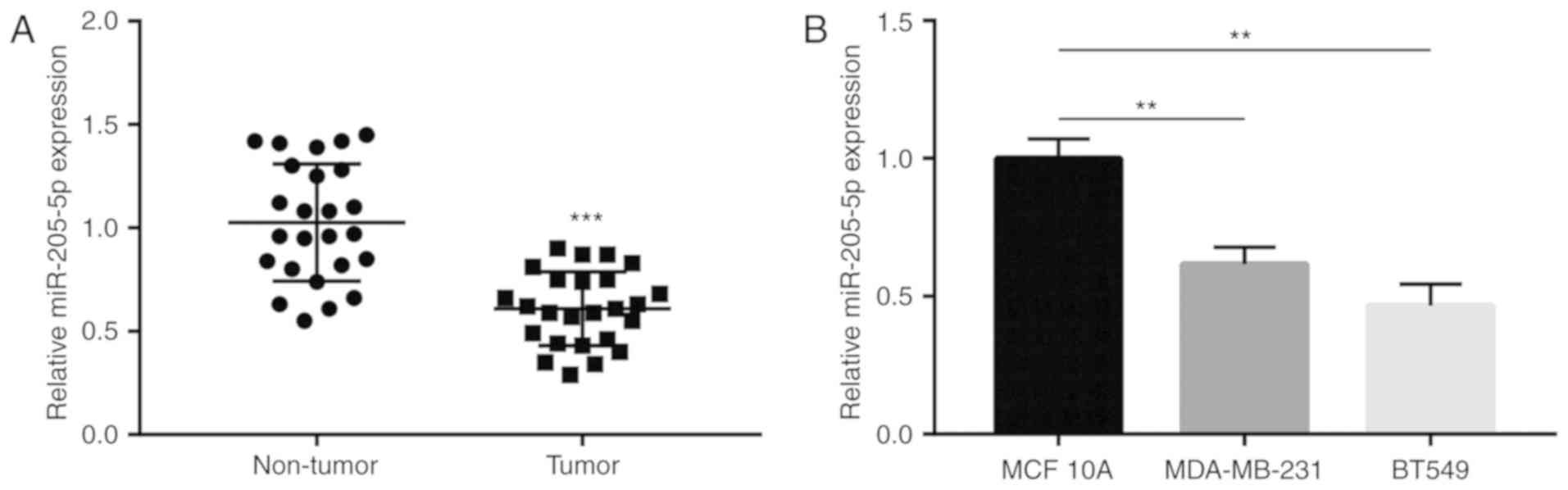
Low miR-205-5p expression is detected
in breast cancer tissues and cells
miR-205-5p expression in breast cancer and matched
normal tissues collected from 25 patients with breast cancer was
compared using RT-qPCR. It was observed that miR-205-5p levels were
significantly decreased in tumor compared with non-tumor tissues
(Fig. 1A). Subsequently, relative
miR-205-5p expression was determined in MDA-MB-231 and BT549 breast
cancer cells and in MCF10A immortalized normal breast cells. The
results revealed that miR-205-5p expression levels in MDA-MB-231
and BT549 were lower compared with MCF10A cells (Fig. 1B). Collectively, these finding
suggested that miR-205-5p expression decreased in breast cancer
tissues and cells compared with normal controls.
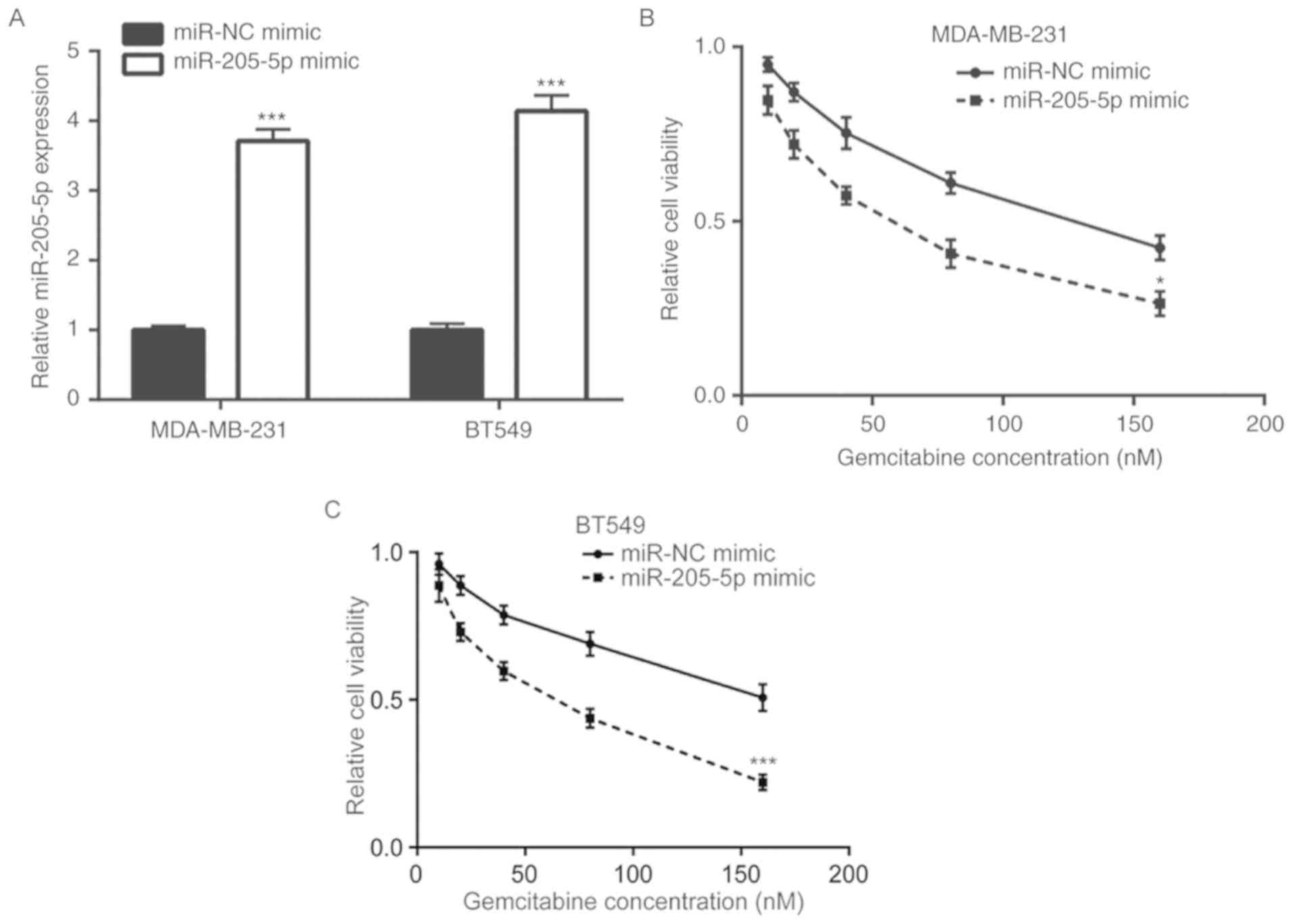
miR-205-5p overexpression increases
gemcitabine sensitivity of breast cancer cells
To investigate the role of miR-205-5p in breast
cancer gemcitabine resistance, a miR-205-5p mimic was transfected
into MDA-MB-231 cells and RT-qPCR was performed to detect
miR-205-5p mRNA levels. The results demonstrated that the
miR-205-5p mimic increased miR-205-5p expression in MDA-MB-231 and
BT549 cells compared with the miR-NC mimic group (Fig. 2A). Overexpression of miR-205-5p
enhanced gemcitabine-induced cell viability reduction in MDA-MB-231
and BT549 cells (Fig. 2B and C).
These data indicated that miR-205-5p overexpression may increase
gemcitabine sensitivity of breast cancer cells.
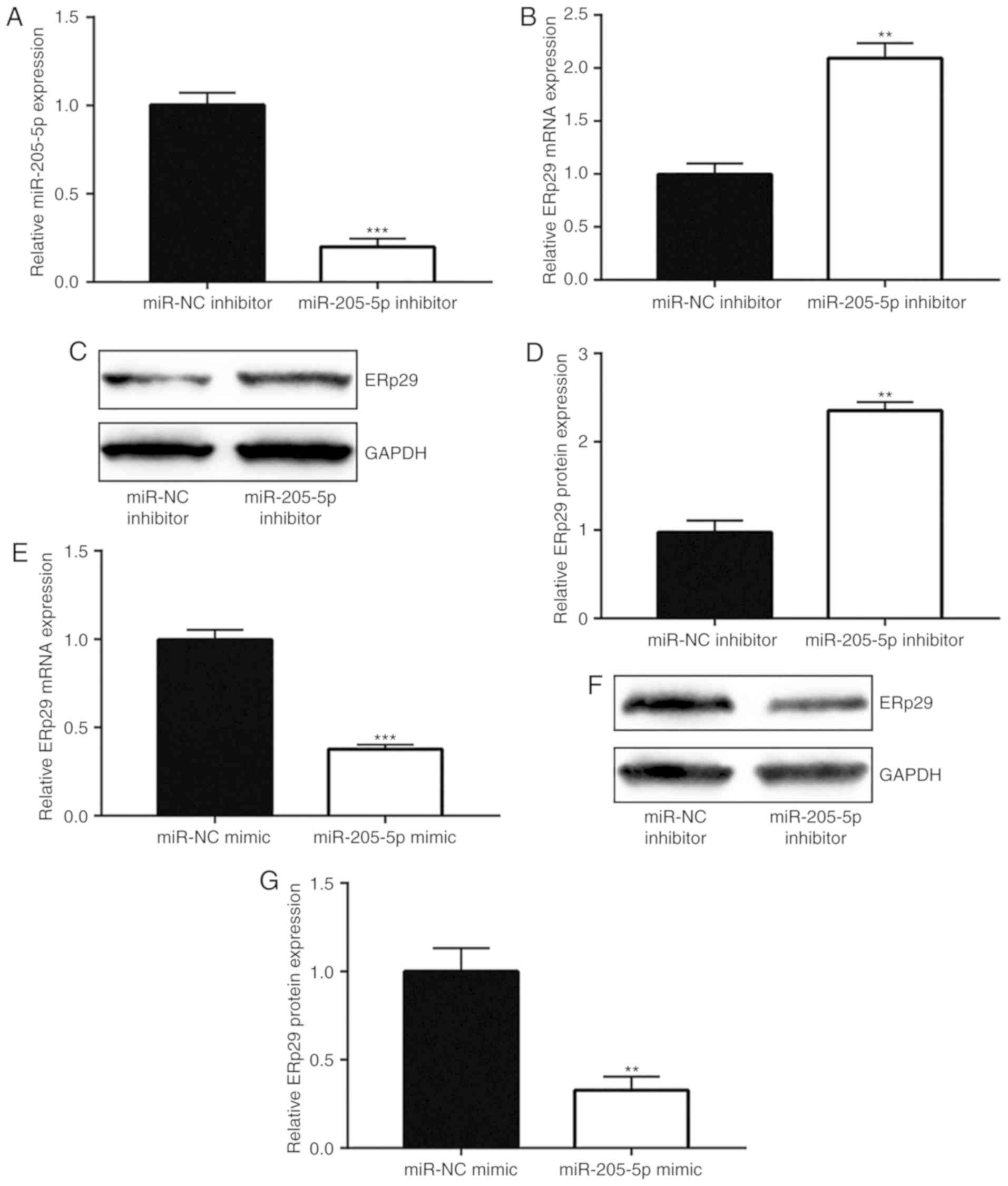
miR-205-5p downregulates ERp29 in
breast cancer cells
The current study explored the association between
miR-205-5p and ERp29 in breast cancer. RT-qPCR was used to confirm
that miR-205-5p inhibitor significantly downregulated miR-205-5p
expression in MDA-MB-231 cells compared with the miR-NC inhibitor
group (Fig. 3A). Inhibition of
miR-205-5p markedly enhanced ERp29 mRNA (Fig. 3B) and protein (Fig. 3C and D) levels. miR-205-5p
overexpression decreased ERp29 mRNA (Fig. 3E) and protein (Fig. 3F and G) levels. Taken together, these
data indicated that miR-205-5p negatively regulated ERp29
expression levels.
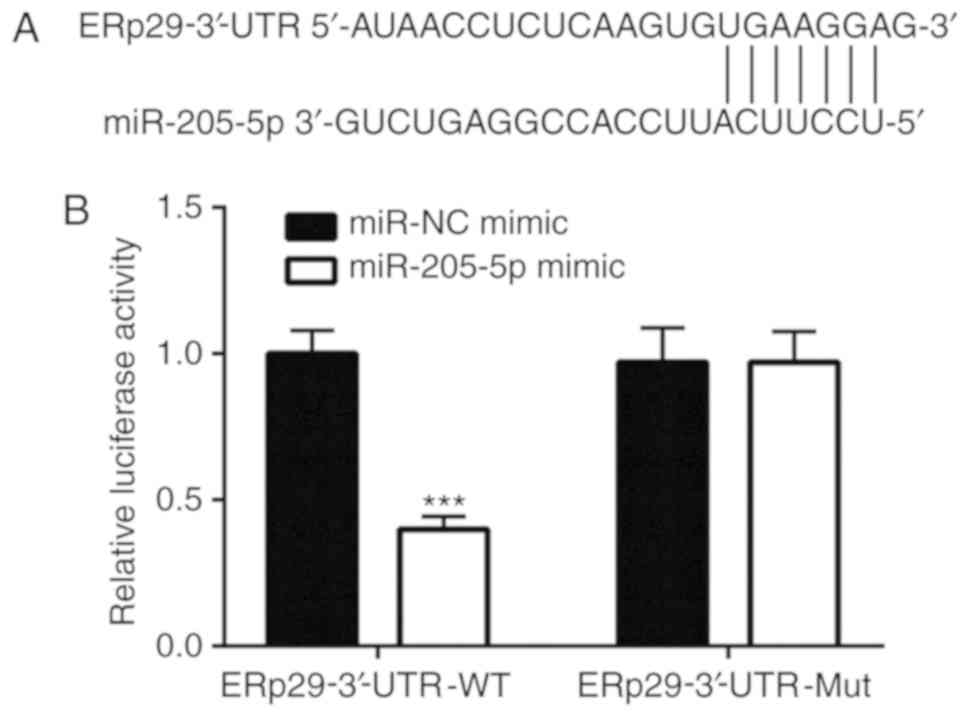
miR-205-5p regulates ERp29 by binding
to the 3′-UTR
According to TargetScan, ERp29 is a potential target
gene of miR-205-5p. A putative binding site for miR-205-5p was
predicted within the 3′-UTR of ERp29 (Fig. 4A). ERp29 WT and Mut 3′-UTR luciferase
reporter gene plasmids were constructed to verify this potential
binding. Dual-luciferase reporter gene assay results revealed that
the miR-205-5p mimic significantly decreased the luciferase
activity in the WT-ERp29-3′-UTR co-transfection system in
MDA-MB-231 cells compared with the miR-NC mimic transfection
(Fig. 4B).
miR-205-5p expression is decreased in
MDA-MB-231/GEM cells
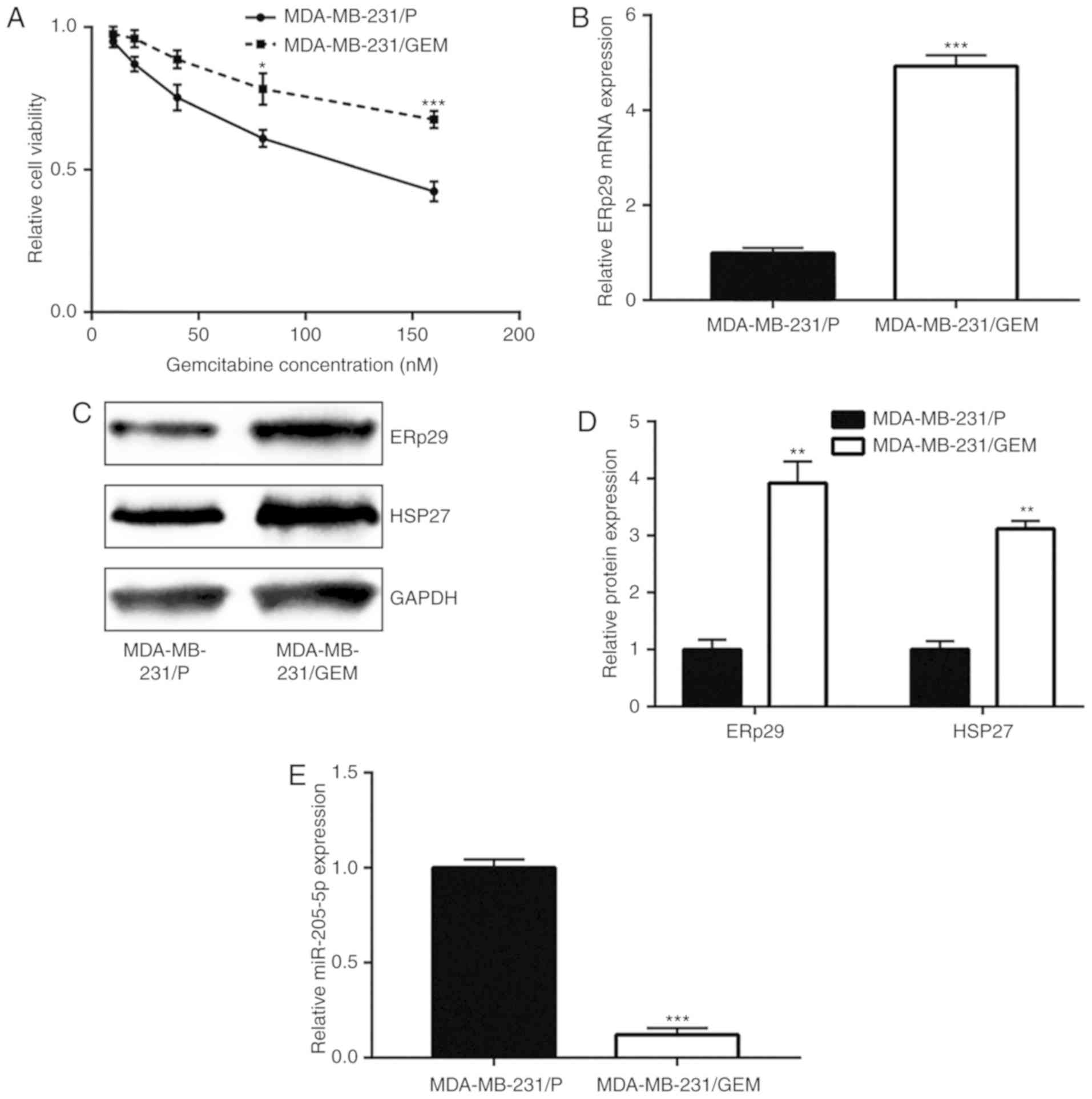
MDA-MB-231/GEM cells were established to study the
mechanisms of gemcitabine resistance. With increasing gemcitabine
concentration, MDA-MB-231/GEM viability was significantly greater
compared with MDA-MB-231/P cells, suggesting MDA-MB-231/GEM cells
were comparatively less insensitive towards gemcitabine treatment
(Fig. 5A). ERp29 mRNA and protein
expression levels were notably increased in MDA-MB-231/GEM cells
compared with the MDA-MB-231/P group (Fig. 5B-D). HSP27 is regulated by ERp29 and
involved in breast cancer drug resistance (12,24). In
the current study, it was observed that HSP27 protein levels
significantly increased in MDA-MB-231/GEM cells compared with
MDA-MB-231/P cells (Fig. 5C and D).
In addition, miR-205-5p expression significantly decreased in
MDA-MB-231/GEM cells compared with MDA-MB-231/P cells (Fig. 5E). miR-205-5p downregulation and
ERp29 upregulation in MDA-MB-231/GEM cells suggested that
miR-205-5p/ERp29 may be involved in the development of gemcitabine
resistance in breast cancer cells.
miR-205-5p overexpression reverses
gemcitabine resistance of MDA-MB-231/GEM cells via ERp29
repression
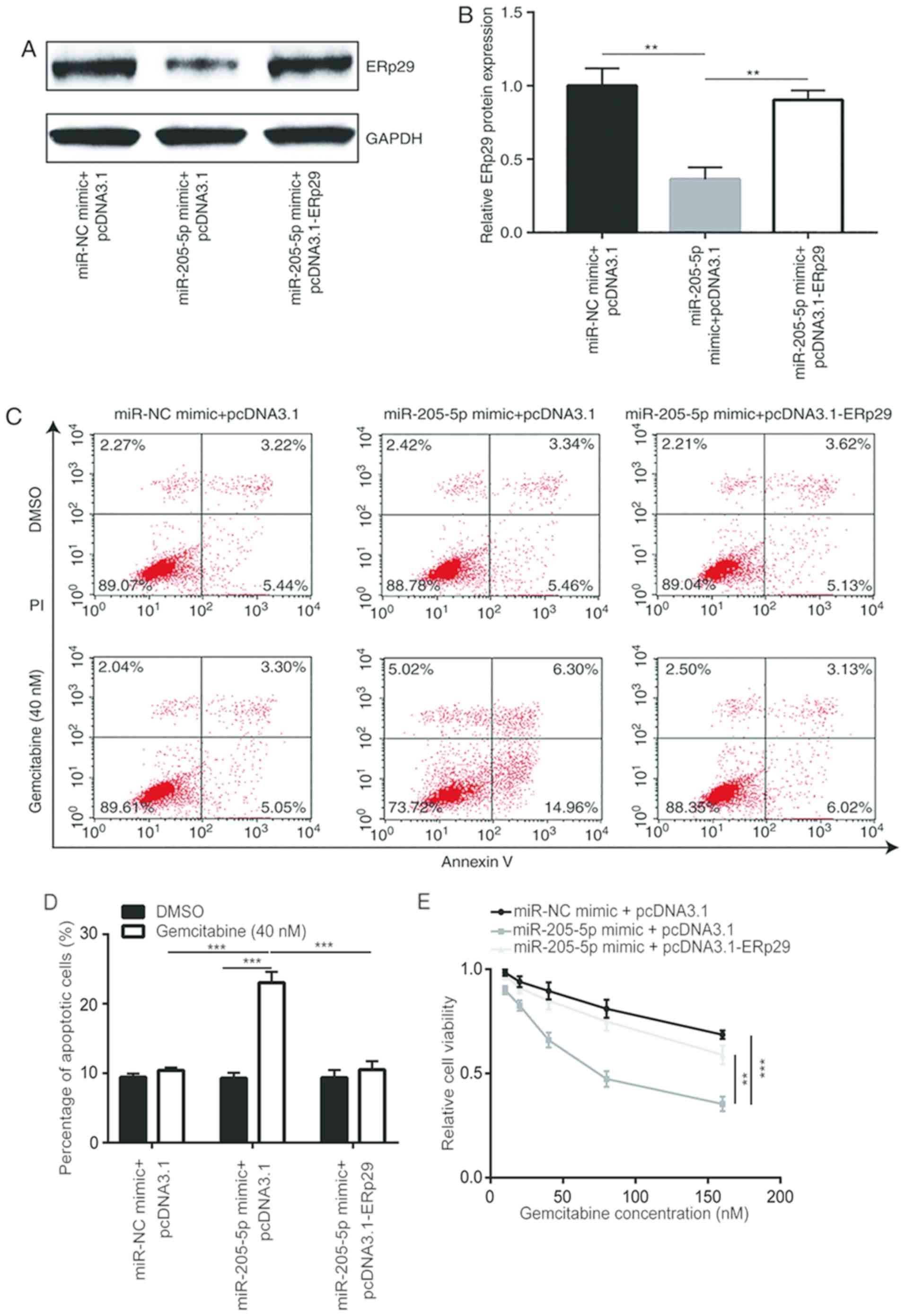
pcDNA3.1-ERp29 recombinant plasmids were constructed
and co-transfected with the miR-205-5p mimic into MDA-MB-231/GEM
cells to confirm the aforementioned results. The results
demonstrated that miR-205-5p overexpression reduced ERp29 protein
expression levels; a phenomenon that was reversed by
co-transfection with pcDNA3.1-ERp29 recombinant plasmid (Fig. 6A and B). To evaluate the function of
miR-205-5p in the development of gemcitabine resistance, cell
apoptosis and viability assays were performed to detect effects of
miR-205-5p and ERp29 overexpression on gemcitabine sensitivity in
breast cancer cells. The results revealed that among cells treated
with gemcitabine, miR-205-5p overexpression significantly enhanced
gemcitabine-induced apoptosis in MDA-MB-231/GEM cells, which was
reversed following transfection with recombinant ERp29. Among cells
treated with DMSO, there were no significant differences between
the 3 different groups. In addition, gemcitabine significantly
increased apoptosis in cells transfected with miR-205-5p mimic +
pcDNA3.1 but not in miR-NC mimic + pcDNA3.1 or miR-205-5p mimic +
pcDNA3.1-ERp29 groups (Fig. 6C and
D). Meanwhile, miR-205-5p overexpression significantly
potentiated the inhibitory effects of gemcitabine on cell viability
in MDA-MB-231/GEM cells, which was significantly reversed by
transfection with recombinant ERp29 (Fig. 6E). These findings indicate that
miR-205-5p overexpression sensitized MDA-MB-231/GEM cells to
gemcitabine by suppressing ERp29 expression.
ERp29 expression is elevated in breast
cancer tissues and negatively correlated with miR-205-5p
expression
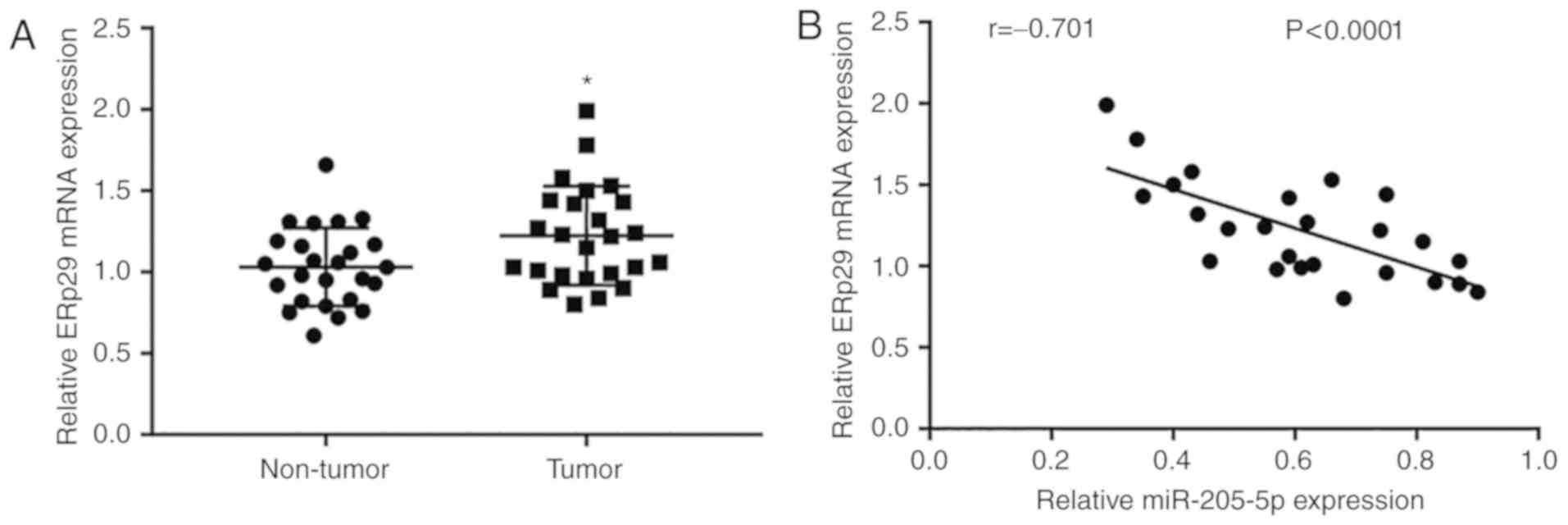
To further study the association between miR-205-5p
and ERp29 in breast cancer tissues, RT-qPCR was used to detect
ERp29 mRNA levels in 25 paired tumor and normal tissues. It was
observed that ERp29 expression was significantly elevated in tumor
tissues compared with non-tumor samples (Fig. 7A). Furthermore, ERp29 mRNA expression
was negatively correlated with the expression of miR-205-5p, as
determined by analyzing the expression data of 25 breast cancer
tissues (Fig. 7B).
Discussion
Breast cancer is one of the leading causes of
cancer-associated mortality among women worldwide (1,2). Despite
advances in breast cancer chemotherapy (7), the development of chemotherapy
resistance remains a major obstacle in successful breast cancer
treatment (8). Gemcitabine is a
widely used chemotherapy drug that serves an important role in the
treatment of advanced breast cancer (6). However, its therapeutic use in cancer
chemotherapy is impeded, at least in part, by drug resistance
(25). Over the past decade,
increasing research efforts focused on exploring the underlying
mechanisms of drug resistance. Recently, various studies have
suggested that miRNAs serve an important role in chemotherapy
resistance (26,27). miRNAs are tumor suppressors or
oncogenes and are able to modulate cancer progression, including
cell proliferation and apoptosis (16,28).
Certain miRNAs, including let-7, miR-122, miR-152 and miR-1246,
have been reported to modulate liver cancer stem cells by directly
or indirectly binding to specific target genes that are involved in
signal pathways, including the Wnt/β-catenin signaling, TGF-β
signaling, JAK/STAT signaling and epithelial-mesenchymal transition
(EMT) pathway (27). Yu et al
(29) reported that miR-200a was
overexpressed in gemcitabine-resistant breast cancer cells and that
miR-200a inhibition restored sensitivity. In this study, miR-200a
promoted DNA damage resistance by inhibiting DNA damage-induced
apoptosis via Yes associated protein 1 and tumor protein p53
inducible nuclear protein 1. Previous studies revealed that
miR-205-5p is one of the most researched miRNAs in breast cancer
and it regulates cell proliferation and invasion (21,30). Li
and Li (31) reported that
miR-205-5p inhibited cell migration and invasion in prostate
carcinoma by targeting zinc finger E-box binding homeobox (ZEB) 1.
Previous studies further reported that compared with differentiated
tumor cells, patient-derived breast cancer stem cells expressed
higher levels of miR-205-5p (22).
miR-205-5p upregulation controls cancer stem cell phenotype by
targeting erb-b2 receptor tyrosine kinase 2, tumor protein p63 and
epidermal growth factor receptor contributing to targeted therapy
resistance (24). miR-205
accumulation confers gemcitabine sensitivity and in
gemcitabine-resistant cancer stem cells it targets and suppresses
ZEB1/2, E2F transcriptional factor 1, erb-b2 receptor tyrosine
kinase 3 and vascular endothelial growth factor A (32). However, the effects of miR-205-5p on
gemcitabine resistance in breast cancer cells remain to be
elucidated. In the current study, using cell viability assays,
miR-205-5p overexpression in combination with gemcitabine treatment
at varying concentration was used to investigate
gemcitabine-sensitivity of breast cancer cells. miR-205-5p
overexpression increased gemcitabine sensitivity in breast cancer
cells. Additionally, miR-205-5p was downregulated in the
established MDA-MB-231/GEM cells and induced miR-205-5p
overexpression reversed gemcitabine resistance of MDA-MB-231/GEM
cells. Furthermore, in the present study, miR-205-5p overexpression
increased apoptosis in MDA-MB-231/GEM cells following treatment
with 40 nM gemcitabine. The results revealed the role of miR-205-5p
in breast cancer gemcitabine resistance.
The current study reported that ERp29 was predicted
as target gene of miR-205-5p according to the TargetScan database.
Transfection with WT and Mut ERp29-3′-UTR luciferase reporter gene
plasmids verified that miR-205-5p regulated ERp29 by binding to the
3′-UTR. ERp29 is negatively correlated with breast cancer
development (33,34) and functions as a tumor suppressor
(35,36). ERp29 overexpression aids tumor cell
survival by inhibiting genotoxic effects caused by chemotherapy
(37). Zhang and Putti (12) investigated the effects of breast
cancer cell chemotherapeutic agents on ERp29 expression. ERp29
expression increased doxorubicin resistance and decreased
doxorubicin-induced apoptosis in MDA-MB-231 cells, and ERp29
knockdown in MCF-7 cells increased the cytotoxicity of doxorubicin.
In the current study, overexpression and inhibition of miR-205-5p
suggested that ERp29 mRNA and protein levels were negatively
regulated by miR-205-5p in breast cancer cells. In addition,
miR-205-5p overexpression-induced apoptosis and decreased cell
viability were reversed by increased ERp29 expression.
In conclusion, it was revealed that miR-205-5p
decreased breast cancer gemcitabine sensitivity in vitro via
regulation of ERp29, suggesting that miR-205-5p may serve as a
potential treatment target in breast cancer.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author's contribution
CM and GW contributed to study design and
supervision. CM, XS and WG were responsible for acquisition of
data. GW performed interpretation of data. Clinical sample
collection and analysis was performed by WG and FF. CM and GW
prepared the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Tianjin Baodi People's Hospital. All patients provided
written consent before being enrolled into the study.
Patient consent for publication
Written informed consent for publication was
provided by all participants.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Smith IE, Dowsett M, Ebbs SR, Dixon JM,
Skene A, Blohmer JU, Ashley SE, Francis S, Boeddinghaus I and Walsh
G; IMPACT Trialists Group, : Neoadjuvant treatment of
postmenopausal breast cancer with anastrozole, tamoxifen, or both
in combination: The immediate preoperative anastrozole, tamoxifen,
or combined with tamoxifen (IMPACT) multicenter double-blind
randomized trial. J Clin Oncol. 23:5108–5116. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gralow JR, Burstein HJ, Wood W, Hortobagyi
GN, Gianni L, von Minckwitz G, Buzdar AU, Smith IE, Symmans WF,
Singh B and Winer EP: Preoperative therapy in invasive breast
cancer: Pathologic assessment and systemic therapy issues in
operable disease. J Clin Oncol. 26:814–819. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tripathy D: Gemcitabine as single-agent
therapy for advanced breast cancer. Clin Breast Cancer. 1
(Suppl):S8–S11. 2002. View Article : Google Scholar
|
|
4
|
Spielmann M: Single-agent gemcitabine as
first line therpy in patient with metastatic breast cancer.
Oncology. 62:2–8. 2011.
|
|
5
|
Blackstein M, Vogel CL, Ambinder R, Cowan
J, Iglesias J and Melemed A: Gemcitabin as first-line therapy in
patient with metastatic breast cancer: A phase II trail. Oncology.
62:2–8. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
CACA-CBCS, Chinese Anti-Cancer
Association, Committee of Breast Cancer Society, . Guidelines for
diagnosis and treatment of breast cancer in China (2017). China
Oncol. 27:1007–3639. 2017.
|
|
7
|
Vincenzi B, Santini D, Spoto S, Finolezzi
E, D'Angelillo RM, La Cesa A and Tonini G: The antineoplastic
treatment in the elderly. Clin Ter. 153:207–215. 2002.PubMed/NCBI
|
|
8
|
Wu ZH, Lin C, Liu MM, Zhang J, Tao Z and
Hu XC: Src inhibition can synergize with gemcitabine and reverse
resistance in triple negative breast cancer cells via the AKT/c-Jun
pathway. PLoS One. 11:e01692302016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang D and Richardson DR: Endoplasmic
reticulum protein 29 (ERp29): An emerging role in cancer. Int J
Biochem Cell Biol. 43:33–36. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Martin C, Ardizzoni A and Rosso R:
Gemcitabine: Safety profile and efficacy in non-small cell lung
cancer unaffected by age. Aging (Milano). 9:297–303.
1997.PubMed/NCBI
|
|
11
|
Wu P, Zhang H, Qi L, Tang Q, Tang Y, Xie
Z, Lv Y, Zhao S and Jiang W: Identification of ERp29 as a biomarker
for predicting nasopharyngeal carcinoma response to radiotherapy.
Oncol Rep. 27:987–994. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang D and Putti TC: Over-expression of
ERp29 attenuates doxorubicin-induced cell apoptosis through
up-regulation of Hsp27 in breast cancer cells. Exp Cell Res Dec.
316:3522–3531. 2010. View Article : Google Scholar
|
|
13
|
Mori-Iwamoto S, Kuramitsu Y, Ryozawa S,
Mikuria K, Fujimoto M, Maehara S, Maehara Y, Okita K, Nakamura K
and Sakaida I: Proteomics finding heat shock protein 27 as a
biomarker for resistance of pancreatic cancer cells to gemcitabine.
Int J Oncol. 31:1345–1350. 2007.PubMed/NCBI
|
|
14
|
Zhang Y, Hu Y, Wang JL, Yao H, Wang H,
Liang L, Li C, Shi H, Chen Y, Fang JY and Xu J: Proteomic
identification of ERP29 as a key chemoresistant factor activated by
the aggregating p53 mutant Arg282Trp. Oncogene. 36:5473–5483. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hammond SM: An overview of microRNAs. Adv
Drug Deliv Rev. 87:3–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Calin GA and Konopleva M: Small gene, big
number, many effects. Blood. 120:240–241. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Farmer DT, Shariat N, Park CY, Liu HJ,
Mavropoulos A and McManus MT: Partially penetrant postnatal
lethality of an epithelial specific MicroRNA in a mouse knockout.
PLoS One. 8:e766342013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Greene SB, Gunaratne PH, Hammond SM and
Rosen JM: A putative role for microRNA-205 in mammary epithelial
cell progenitors. J Cell Sci. 123:606–618. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu H, Zhu S and Mo YY: Suppression of cell
growth and invasion by miR-205 in breast cancer. Cell Res.
19:439–448. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
De Cola A, Volpe S, Budani MC, Ferracin M,
Lattanzio R, Turdo A, D'Agostino D, Capone E, Stassi G, Todaro M,
et al: MIR-205-5p-mediated downregulation of ErbB/HER receptors in
breast cancer stem cells results in targeted therapy resistance.
Cell Death. 6:e18232015. View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qi L, Wu P, Zhang X, Qiu Y, Jiang W, Huang
D, Liu Y, Tan P and Tian Y: Inhibiting ERp29 expression enhances
radiosensitivity in human nasopharyngeal carcinoma cell lines. Med
Oncol. 29:721–728. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dyawanapelly S, Kumar A and Chourasia MK:
Lessons learned from gemcitabine: Impact of therapeutic carrier
systems and Gemcitabine's drug conjugates on cancer therapy. Crit
Rev Ther Drug Carrier Syst. 34:63–96. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ying SY, Chang DC and Lin SL: The microRNA
(miRNA): Overview of the RNA genes that modulate gene function. Mol
Biotechnol. 38:257–268. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lou W, Liu J, Gao Y, Zhong G, Ding B, Xu L
and Fan W: MicroRNA regulation of liver cancer stem cells. Am J
Cancer Res. 8:1126–1141. 2018.PubMed/NCBI
|
|
28
|
Zhang W, Liu J and Wang G: The role of
microRNAs in human breast cancer progression. Tumour Biol.
35:6235–6244. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yu SJ, Yang L, Hong Q, Kuang XY, Di GH and
Shao ZM: MicroRNA-200a confers chemoresistance by antagonizing
TP53INP1 and YAP1 in human breast cancer. BMC Cancer. 18:742018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gregory PA, Bracken CP, Bert AG and
Goodall GJ: MicroRNAs as regulators of epithelial-mesenchymal
transition. Cell Cycle. 7:3112–3118. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li L and Li S: miR-205-5p inhibits cell
migration and invasion in prostatic carcinoma by targeting ZEB1.
Oncol Lett. 16:1715–1721. 2018.PubMed/NCBI
|
|
32
|
Okamoto K, Miyoshi K and Murawaki Y:
miR-29b, miR-205 and miR-221 enhance chemosensitivity to
gemcitabine in HuH28 human cholangiocarcinoma cells. PLoS One.
8:e776232013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bambang IF, Lee YK, Richardson DR and
Zhang D: Endoplasmic reticulum protein 29 regulates epithelial cell
integrity during the mesenchymal-epithelial transition in breast
cancer cells. Oncogene. 32:1240–1251. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Deng YJ, Tang N, Liu C, Zhang JY, An SL,
Peng YL, Ma LL, Li GQ, Jiang Q, Hu CT, et al: CLIC4, ERp29, and
Smac/DIABLO derived from metastatic cancer stem-like cells stratify
prognostic risks of colorectal cancer. Clin Cancer Res.
20:3809–3817. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bambang IF, Xu S, Zhou J, Salto-Tellez M,
Sethi SK and Zhang D: Overexpression of endoplasmic reticulum
protein 29 regulates mesenchymal-epithelial transition and
suppresses xenograft tumor growth of invasive breast cancer cells.
Lab Invest. 89:1229–1242. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shnyder SD, Mangum JE and Hubbard MJ:
Triplex profiling of functionally distinct chaperones
(ERp29/PDI/BiP) reveals marked heterogeneity of the endoplasmic
reticulum proteome in cancer. J Proteome Res. 7:3364–3372. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Farmaki E, Mkrtchian S, Papazian I,
Papavassiliou AG and Kiaris H: ERp29 regulates response to
doxorubicin by a PERK-mediated mechanism. Biochim Biophys Acta.
1813:1165–1171. 2011. View Article : Google Scholar : PubMed/NCBI
|