Introduction
Pathological neovascularization (NV) is a common
cause of blindness globally and occurs in retinal/choroidal
vascular diseases, including retinopathy of prematurity (ROP),
diabetic retinopathy and age-related macular degeneration. Aberrant
neovascularization occurs due to the demand for oxygen and energy
substrates (1). Although effective
therapeutic methods for ocular NV are currently available, they are
associated with certain disadvantages. For example, laser
photocoagulation preserves central vision but leads to the loss of
peripheral vision (2), whilst
vitreous injection of anti-angiogenic agents, such as vascular
endothelial growth factor (VEGF) inhibitors, have become the
preferred therapy option for the inhibition of NV. However,
anti-VEGF-treatment has little or no efficacy in certain patients
because of secondary factors, including fibrosis of NV (3) and enhancement of vascular stability by
pericytes (4). Therefore, efforts
are being made to develop novel therapeutic strategies in addition
to the classical therapies mentioned above.
Platelet-derived growth factor (PDGF) is a family of
chemokines and mitogens that consist of five members: PDGF-AA,
PDGF-BB, PDGF-AB, PDGF-C and PDGF-D. PDGF is involved in vascular
homeostasis by activating their corresponding tyrosine kinase
receptors, PDGFR-α and PDGFR-β. In the eyes, PDGF participates in
NV processes of the choroid, retina, and cornea by binding PDGFR,
and also serves a pro-angiogenic role in a VEGF-dependent or
-independent manner (5,6). The PDGF/PDGFR axis has a wide range of
cellular targets and may serve a greater role compared with the
VEGF/VEGFR axis. Consequently, inhibition of the PDGF/PDGFR axis
may be a potential therapeutic target for diseases associated with
angiogenesis.
In microvessels, PDGF-BB is secreted by endothelial
cells, and binds to PDGFR-β on the surfaces of pericytes. The
PDGF-BB/PDGFR-β signaling pathway has essential effects on the
formation and maturation of the blood-retinal barrier through the
recruitment of pericytes onto new capillaries (7). Pericytes are involved in angiogenic
cascades, including the formation and maturation of NV. In a
laser-induced model of choroidal neovascularization (CNV), a
PDGFRβ+ scaffold that limits the extent of NV formed before the
formation of CNV lesions (8). PDGF
inhibitor (E10030) combined with ranibizumab (an anti-VEGF agent)
was demonstrated to be superior to anti-VEGF monotherapy in a
previous phase IIb clinical study (9). Although superiority was not shown in a
phase III trial, anti-PDGF agents could yet serve a role in
reducing the injection burden and improving outcome for patients
(10).
Stromal cell-derived factor-1α (SDF-1α; or C-X-C
motif chemokine 12) is a chemokine that exerts its biological
function by binding to its receptors, chemokine (C-X-C motif)
receptors 4 (CXCR4) and 7 (CXCR7) (11). The SDF-1α pathway recruits
endothelial precursor cells or hematopoietic stem cells to
neoangiogenic niches to participate in the formation of CNV
(12). In addition, in a previous
study performed in our laboratory, it was demonstrated that the
SDF-1α pathway is critical for pathological angiogenesis in the rat
choroidal NV model (13) and there
may be crosslinks between SDF-1α and other molecules in the
pathogenesis of CNV (14,15). When the SDF-1α/CXCR4 axis was impeded
during tumor NV, PDGF-B expression was reduced and bone
marrow-derived pericyte differentiation was inhibited (16). Indeed, tumor-derived PDGF-B induces
SDF-1α expression in endothelial cells, which is consistent with
PDGF-B-induced pericyte recruitment during angiogenesis (17). Nonetheless, the effect of
PDGF-BB/SDF-1α on retinal microvascular pericytes, which is an
essential component of angiogenesis, remains unclear and require
further study.
In the present study, evidence is provided for the
first time that CXCR4 and CXCR7 are expressed on retinal
microvascular pericytes. In addition, PDGF-BB treatment increased
CXCR4 and CXCR7 expression, which subsequently potentiated
SDF-1α-induced proliferation and migration in retinal microvascular
pericytes.
Materials and methods
Cell culture and treatment
Primary human retinal microvascular pericytes were
purchased from Angio-Proteomie (cat. no. CAP-0025) and maintained
in Dulbecco's modified Eagle's medium (DMEM) (Sigma-Aldrich; Merck
KGaA) supplemented with 4,500 mg/l glucose, L-glutamine, sodium
pyruvate, sodium bicarbonate and 10% fetal bovine serum (FBS;
Gibco, Thermo Fisher Scientific, Inc.) in a humidified atmosphere
at 37°C with 5% CO2.
Materials
Human PDGF-BB protein was purchased from Novus
Biologicals (cat. no. NBP2-35203), LLC. Recombinant human SDF-1α
(CXCL12) was obtained from PeproTech (cat. no. 300-28A), Inc.
AMD3100 (a CXCR4 inhibitor, cat. no. S8030) and niclosamide (a
STAT3 inhibitor, cat. no. S3030) were purchased from Selleck
Chemicals. Anti-CXCR4 (cat. no. ab124824, 1:100), anti-CXCR7 (cat.
no. ab72100, at 6 µg/ml), and anti-PDGFR-β (cat. no. ab69506, at 1
µg/ml) were obtained from Abcam. Anti-β-tubulin was obtained from
Absin Biotechnology Co., Ltd (cat. no. abs830032, at 0.5 µg/ml).
Anti-STAT3 (cat. no. ET1605-45, 1:1,000) and anti-phosphorylated
(p)-STAT3 (cat. no. ET1603-40, 1:1,000) were obtained from Hangzhou
Hua'An Biotechnology Co., Ltd. Anti-ERK-1/2 (cat. no. 9102,
1:1,000), anti-p-ERK1/2 (cat. no. 9106, 1:2,000), anti-AKT (cat.
no. 9272, 1:1,000), and anti-p-AKT (cat. no. 9611, 1:1,000) were
obtained from Cell Signaling Technology, Inc. Anti-mouse
horseradish peroxidase (HRP)-conjugated secondary antibodies (cat.
no. A0216, 1:1,000) and anti-rabbit HRP-conjugated secondary
antibodies (cat. no. A0208, 1:1,000) were obtained from Beyotime
Institute of Biotechnology. Primary Antibody Dilution Buffer (cat.
no. P0023A) and Secondary Antibody Dilution Buffer (cat. no.
P0023D) were also obtained from Beyotime Institute of
Biotechnology.
Western blotting
Cells were lysed using radioimmunoprecipitation
assay lysis buffer (Beyotime Institute of Biotechnology)
supplemented with protease inhibitor. Total protein (40 µg/lane)
was separated by 6 or 8% sodium dodecyl sulfate polyacrylamide gel
electrophoresis and transferred onto polyvinylidene fluoride
membranes. Membranes were then blocked with 10% skimmed milk
diluted in TBS-T solution (0.1% Tween-20) for 1 h at room
temperature and incubated overnight at 4°C with their respective
primary antibodies. Proteins of interest were visualized with
HRP-conjugated secondary antibodies at 1:5,000 dilutions at room
temperature for 1 h and subsequently ECL reagents (Beyotime
Institute of Biotechnology). Densitometric analysis was performed
using Tanon 5200 Image System (Tanon Science and Technology Co.,
Ltd.).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA from retinal microvascular pericytes was
extracted using TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Total RNA was converted
into complementary DNA (cDNA) using PrimeScript™ RT Master Mix
(Takara Bio, Inc.) according to manufacturer's protocol. The
reaction mixture was incubated under the following condition: 37°C
for 15 min, 85°C for 5 sec and 4°C for 5 min. cDNA was subsequently
used for qPCR according to the manufacturer's protocols. Expression
levels of CXCR4 and CXCR7 were analyzed using SYBR®
Premix Ex Taq™ kit (Takara Bio, Inc.) in a Mastercycler®
ep realplex machine (Eppendorf). The qPCR thermocycler conditions
were as follows: Initial denaturation at 95°C for 10 sec, followed
by 40 cycles of 95°C for 5 sec and 60°C for 34 sec. β-Tubulin were
used as internal reference. Relative mRNA expression levels were
determined using the 2−ΔΔCq method (18). The primers used for the qPCR
experiments are shown in Table
I.
 | Table I.Primer pairs used for reverse
transcription-quantitative PCR. |
Table I.
Primer pairs used for reverse
transcription-quantitative PCR.
| Gene | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| Human
β-Tubulin |
AGCGGGAAATCGTGCGTG |
CAGGGTACATGGTGGTGCC |
| Human CXCR4 |
GGCAATGGATTGGTCATCCT |
CATCTTGAACCTGGCCATTG |
| Human CXCR7 |
AGGTGTCAGGCAGAGACACG |
AGGTGTCAGGCAGAGACACG |
| Human PDGFR |
ATGGACATGAGCAAGGACGA |
CCAGCTTGCCTTCACAGATG |
Small interfering (si)-RNA-mediated
knockdown of PDGFR-β gene expression
siRNA for PDGFR-β were purchased from Shanghai
GenePharma Co., Ltd. The sequence for siRNA-PDGFR-β was
5′-GACGUCAAAUAUGCAGACATT-3′. The sequence for scrambled
siRNA-negative control was 5′-UUCUCCGAACGUGUCACGUTT-3′. Cells were
seeded into 6-well plates at a density of 2×106
cells/well and incubated at 37°C. At 70% confluency, cells were
transfected with siRNA-PDGFR-β using Lipofectamine® 2000
transfection reagent (Thermo Fisher Scientific Inc.) according to
manufacturer's protocol. Briefly, 100 pmol siRNA was first mixed
with 5 µl Lipofectamine 2000 and incubation for 25 min at room
temperature before this mixture was added to the cells. Further
experiments were performed 24 h after transfection.
Cell proliferation
Cell Counting Kit-8 (CCK-8) assay (Beyotime
Institute of Biotechnology) was performed to assess pericyte cell
viability, in which 5×103 retinal microvascular
pericytes were first seeded into 96-well plates. A total of five
groups were designated: Negative control (NC) group, SDF-1α group,
PDGF-BB + SDF-1α group, PDGF-BB + SDF-1α + AMD3100 group and
PDGF-BB + SDF-1α + niclosamide group. With the exception of the NC
group, SDF-1α (100 ng/ml) was added to all groups. PDGF-BB (10
ng/ml) was added to the PDGF-BB + SDF-1α, PDGF-BB + SDF-1α +
AMD3100 and PDGF-BB + SDF-1α + niclosamide groups. The
concentration of AMD3100 and niclosamide used was 1 µM. Fresh
serum-free medium was added with a final volume of 100 µl to every
well. Cells were first incubated for 2 h at 37°C, following which
10 µl CCK-8 reagent was added to each well at 24, 48, and 72 h.
Finally, absorbance was measured at 450 nm using a microplate
reader (Thermo Fisher Scientific, Inc.). All experiments were
performed in triplicates.
Cell migration
Cell migration was measured in 24-well plates with
8.0 µm pore size polycarbonate membrane inserts (Corning Inc.). As
aforementioned, five groups were designated. With the exception of
the NC and SDF-1α groups, pericytes (4×104 cells)
diluted in 200 µl serum-free medium were treated with PDGF-BB (10
ng/ml) and seeded into the upper chambers. Each well contained 500
µl DMEM supplemented with 10% FBS supplemented with (100 ng/ml) or
without SDF-1α (NC group). In the PDGF-BB + SDF-1α + AMD3100 and
PDGF-BB + SDF-1α + niclosamide groups, pericytes were treated with
AMD3100 (1 µM) or niclosamide (1 µM) in the lower chambers. After
incubation at 37°C for 24 h, inserts were fixed with 4%
paraformaldehyde at room temperature for 30 min and dyed with
crystal violet at room temperature for 20 min (Beyotime Institute
of Biotechnology). The numbers of migrated cells across the
membrane were counted from five random fields of view/insert using
a light microscope (magnification, ×40).
Apoptosis assay using flow
cytometry
Retinal microvascular pericytes were plated into
6-well plates at 3×105 cells/well and incubated for 24
h. Following treatment with 0.25% ethylenediaminetetraacetic acid
(EDTA)-free trypsin (Gibco; Thermo Fisher Scientific, Inc.), cells
were adjusted to a density of 1×106/ml and centrifuged
at 100 × g for 5 min at room temperature. The cell supernatant was
then discarded and 2 ml phosphate-buffered saline (PBS) was added
to the cell pellet to wash the cells. After two-time wash with PBS,
the cell pellet was then resuspended in 100 µl binding buffer
solution, following which 5 µl propidium iodide (PI) and 5 µl
Annexin V-fluorescein isothiocyanate (FITC) dyes were added to the
cells with this mixture subsequently incubated for 15 min at room
temperature in the dark. After incubation, a total of 400 µl
binding buffer was added to the cells. The FITC Annexin V apoptosis
detection kit I was purchased from BD Pharmingen™ (BD Biosciences).
Flow cytometry (BD FACSCalibur™; BD Biosciences) and BD CELLQuest™
Pro software (version 5.1; BD Biosciences) were used to measure
cell apoptosis at an excitation wavelength of 488 nm.
Statistical analysis
All data are presented as mean ± SD. Student's
t-test and one-way analysis of variance (ANOVA) followed by
Bonferroni's or Dunnett's post hoc test were used to calculate
statistical differences. Each experiment was repeated at least 3
times. Prism 6 (GraphPad Software, Inc.) was used perform
statistical analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
PDGF-BB increases pericyte CXCR4 and
CXCR7 expression
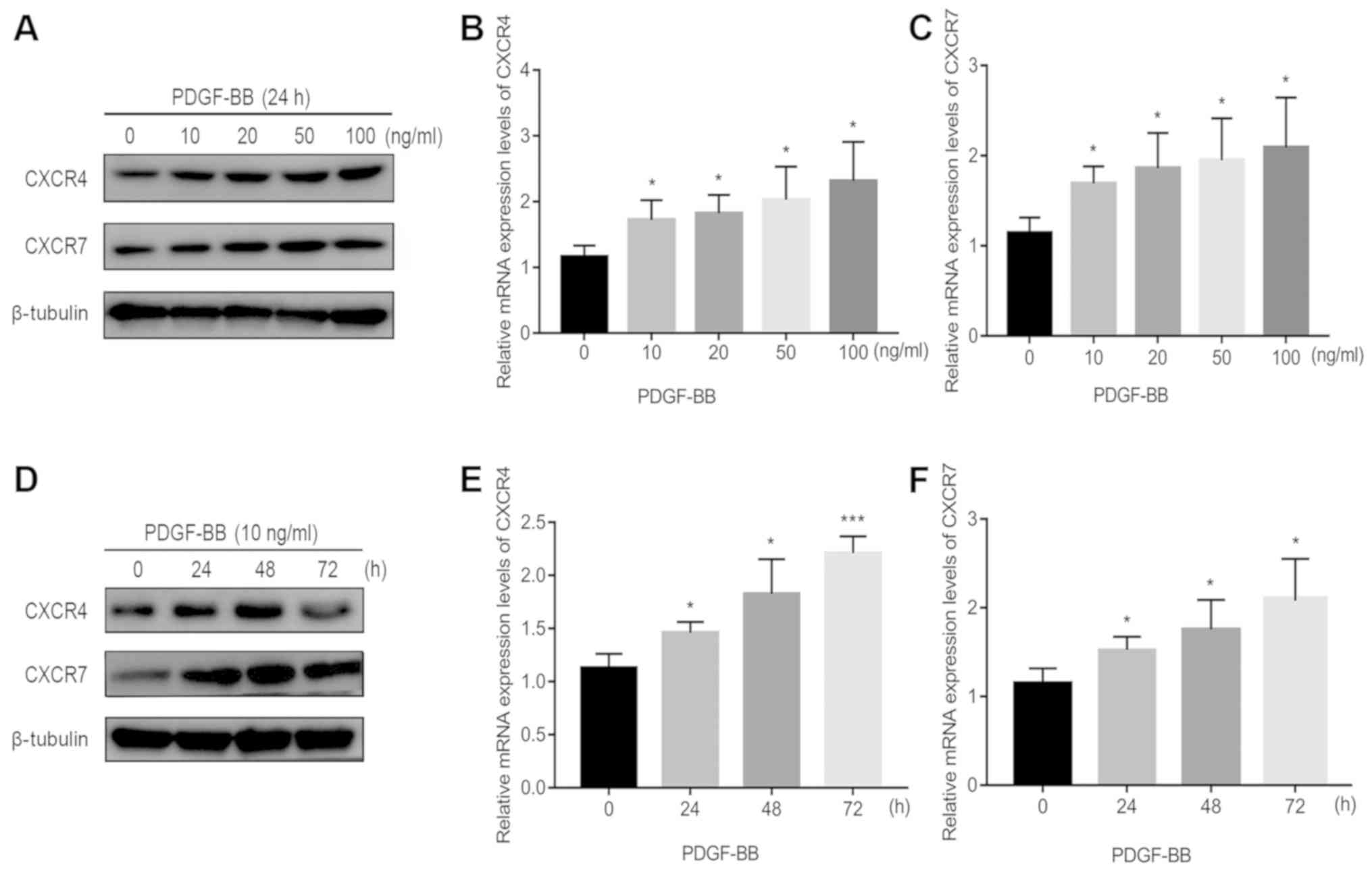
The effect of increasing concentrations of PDGF-BB
at a number of incubation times on CXCR4 and CXCR7 expression in
pericytes is shown in Figs. 1 and
S1. Endogenous CXCR4 and CXCR7
expression was first confirmed in pericytes in the absence of
PDGF-BB treatment (Fig. 1).
Following 24 h of PDGF-BB treatment in Fig. 1B-C, 10 ng/ml PDGF-BB significantly
increased CXCR4/CXCR7 mRNA expression, but further increases in the
PDGF-BB dose only resulted in mild further increases (Fig. 1A-C). Following 24 h of PDGF-BB
treatment in Fig. 1A and S1A, 10 ng/ml PDGF-BB significantly
increased CXCR4/CXCR7 protein expression. However, when the dose of
PDGF-BB was increased to 100 ng/ml, protein levels of CXCR7 reduced
slightly (Figs. 1A and S1A). To evaluate the time dependency of
this effect, PDGF-BB (10 ng/ml) was added to the cell culture
medium at 0, 24, 48 and 72 h. With increasing treatment time, CXCR4
and CXCR7 mRNA and protein expression significantly increased at 24
and 48 h (Figs. 1D-F and S1B). However, at 72 h, the levels of CXCR4
and CXCR7 protein expression reduced slightly (Figs. 1D and S1B). These results suggest that PDGF-BB
treatment significantly upregulated CXCR4 and CXCR7 expression in
pericytes.
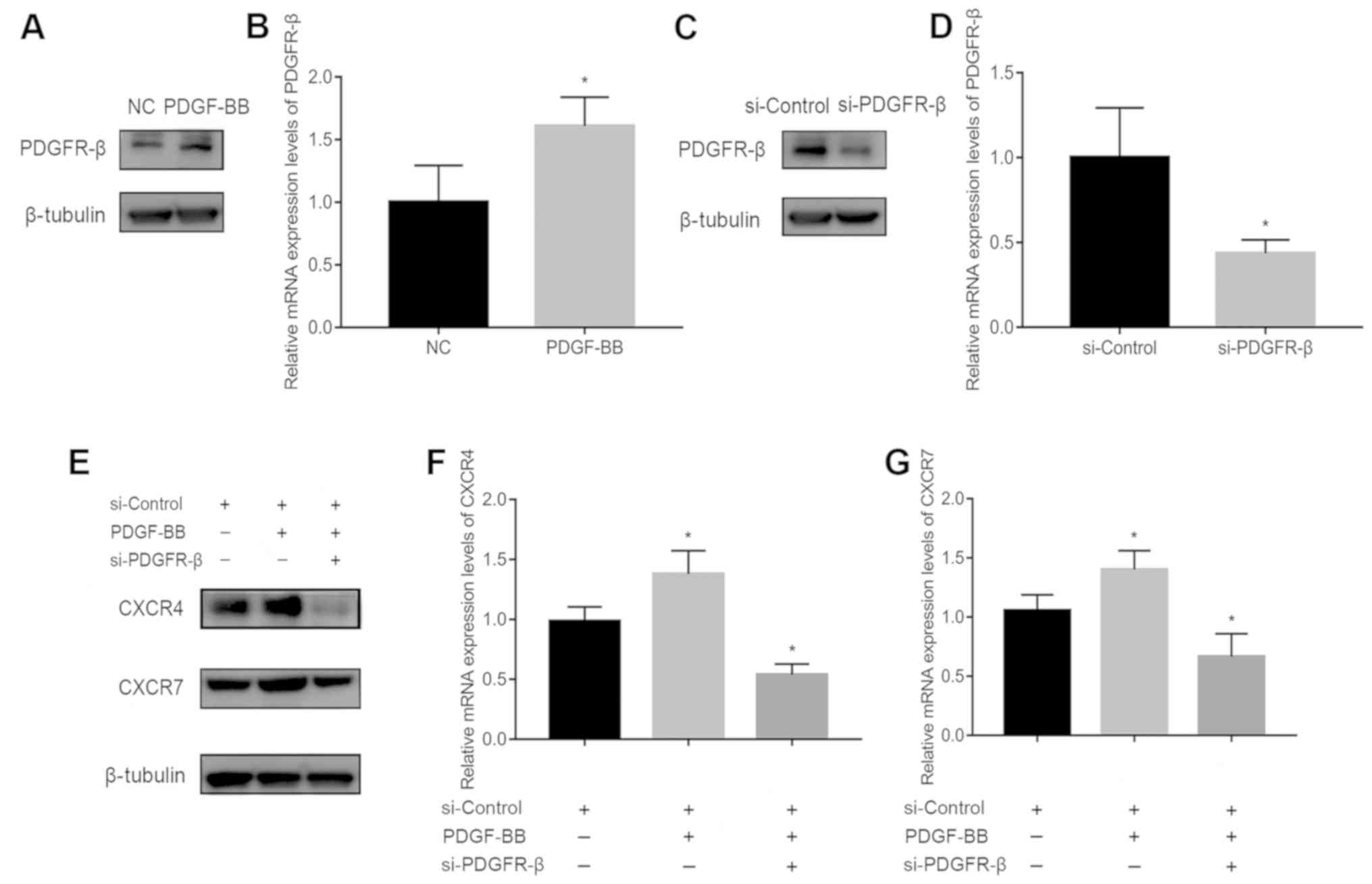
SiRNA transfection downregulates
PDGFR-β, CXCR4, and CXCR7 expression
To assess the involvement of PDGFR-β in
PDGF-BB-stimulated CXCR4 and CXCR7 expression, PDGFR-β expression
was first measured in pericytes treated with or without 10 ng/ml
PDGF-BB. PDGFR-β expression was significantly increased by PDGF-BB
stimulation (Fig. 2A and B). It was
subsequently found that PDGFR-β expression was significantly
decreased following transfection with siRNA-PDGFR-β for 24 h
compared with cells transfected with the siRNA-Control (Fig. 2C and D). In addition, CXCR4 and CXCR7
expression was reduced in PDGF-BB-treated pericytes transfected
with siRNA-PDGFR-β compared with those transfected with negative
siRNA-Control (Fig. 2E-G). These
results suggest that the activation of PDGFR-β signaling in
pericytes is a prerequisite for the upregulation of CXCR4/CXCR7
expression.
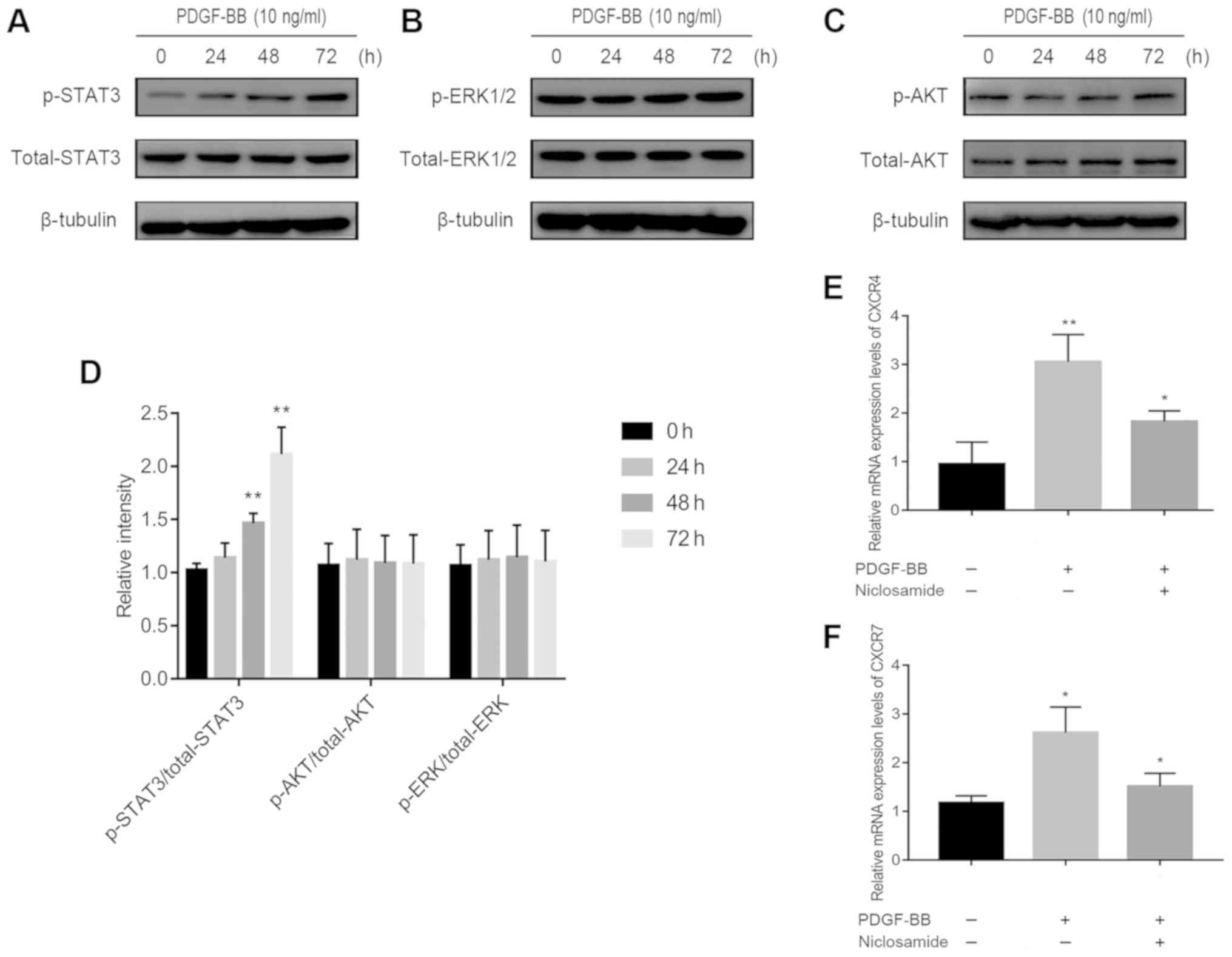
The STAT3 signaling pathway is
involved in PDGF-BB-induced up-regulation of CXCR4 and CXCR7
expression
The potential mechanism of PDGF-BB-induced
CXCR4/CXCR7 upregulation was subsequently investigated. PDGF-BB has
been previously reported to activate phosphorylation of ERK-1/2 in
human brain pericytes (19) and AKT
in brain pericytes after ischemic stroke and in HT29 cells
(20,21). In addition, PDGF-BB has been
demonstrated to activate janus kinase 2 (Jak2)/STAT3 signaling
(22), resulting in increased CXCR4
expression (23). Significantly
increased ratio of p-STAT3/total STAT3 expression (Fig. 3A), but not of p-ERK-1/2/total ERK-1/2
(Fig. 3B) or p-AKT/total AKT
(Fig. 3C), was observed in
PDGF-BB-treated pericytes (Fig.
3A-D). This suggests that PDGF-BB regulated CXCR4 and CXCR7 by
signaling through the Jak2/STAT3 pathway and not the ERK1/2 or AKT
pathway. Supporting this, pre-treatment with niclosamide (1 µM), a
STAT3 inhibitor, significantly reversed PDGF-BB-induced CXCR4 and
CXCR7 upregulation, in pericytes (Fig.
3E and F).
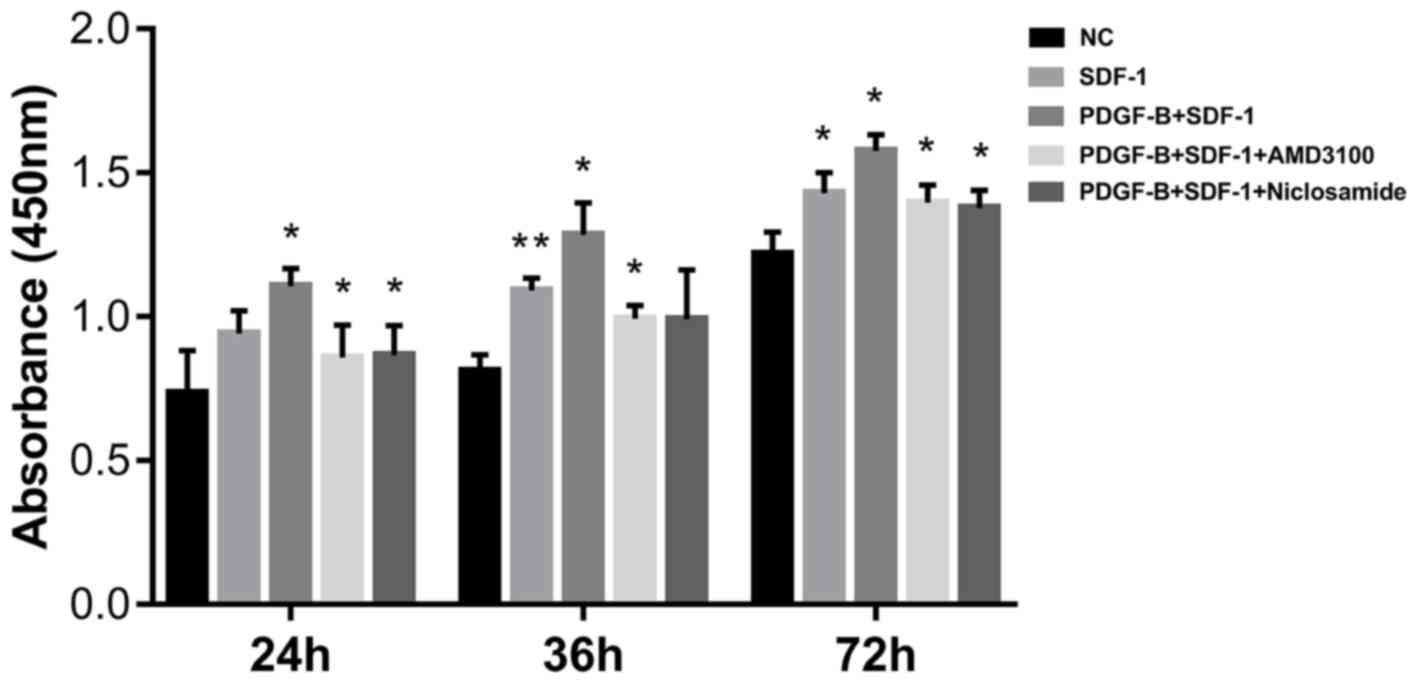
PDGF-BB promotes SDF-1α-treated
pericyte cell viability
CCK-8 assay was next applied to measure the effect
of PDGF-BB treatment on pericyte viability. Pericytes were first
pretreated with 10 ng/ml PDGF-BB (with or without 1 µΜ AMD3100 or 1
µΜ niclosamide for 24 h), followed by stimulation with SDF-1α (10
ng/ml, 24 h). Compared with the NC group, SDF-1α treatment
significantly enhanced pericyte proliferation, especially at 36 and
72 h. Compared with SDF-1α group, additive PDGF-BB treatment
increased pericyte proliferation further (Fig. 4). CXCR4 inhibition (AMD3100) and
STAT3 inhibition (Niclosamide) significantly reversed the
stimulatory effects of SDF-1α and PDGF-BB (Fig. 4). These observations suggest that
upregulation of CXCR4 by PDGF-BB enhances SDF-1α-induced pericyte
cell viability.
PDGF-BB promotes SDF-1α-induced
pericyte migration, which is reversed by niclosamide and
AMD130
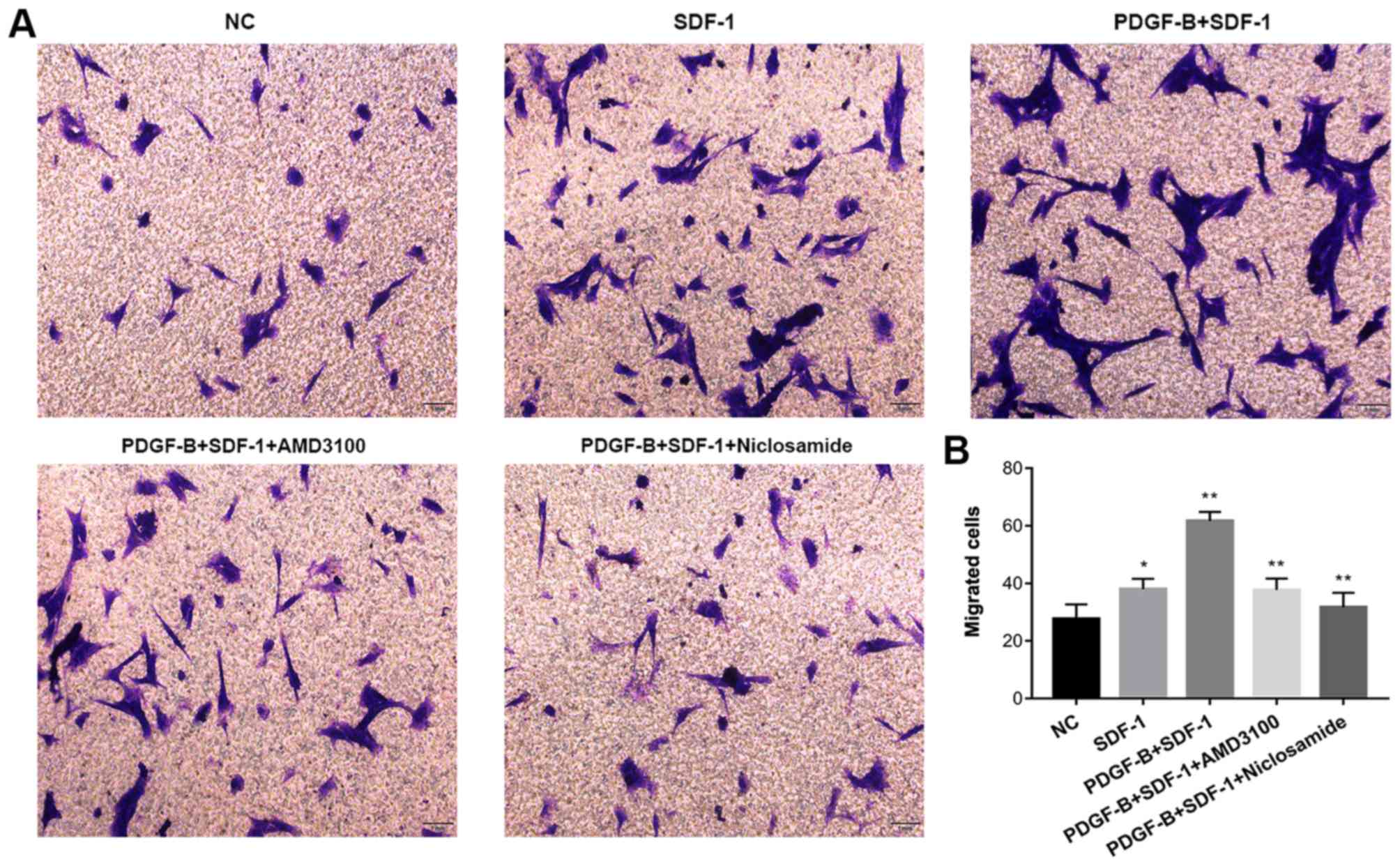
The Transwell migration assay was used to determine
whether PDGF-BB treatment influences SDF-1α-treated migratory
capability of pericytes in vitro. Pericytes were first
pretreated with 10 ng/ml PDGF-BB (with or without 1 µM AMD3100 or 1
µM niclosamide for 24 h), followed by stimulation with SDF-1α (10
ng/ml, 24 h). Compared with NC group, SDF-1α treatment increased
pericyte migration. Compared with the SDF-1α group, PDGF-BB +
SDF-1α group exhibited significantly enhanced pericyte migration.
CXCR4 inhibition (AMD3100) and STAT3 inhibition (Niclosamide)
significantly attenuated the stimulatory effects of SDF-1α and
PDGF-BB (Fig. 5). These findings
suggest that upregulation of CXCR4 expression by PDGF-BB
potentiates SDF-1α-induced pericyte migration.
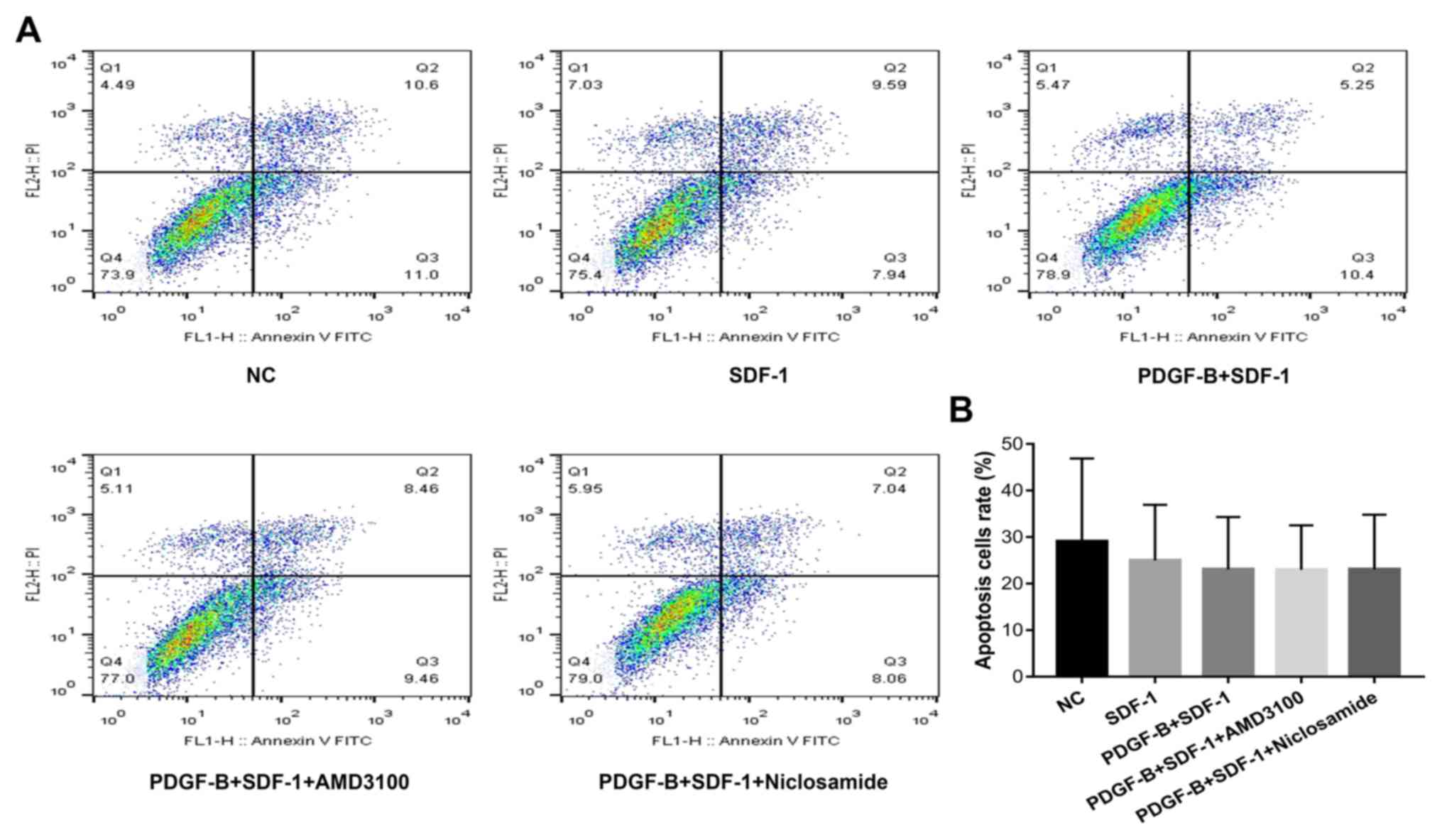
PDGF-BB treatment does not affect
apoptosis in SDF-1α-treated pericytes
Flow cytometry was used to measure cell apoptosis in
PDGF-BB and SDF-1α-treated pericytes (Fig. 6). Compared with the NC group,
apoptosis in the other four groups were slightly decreased, though
there were no statistically significant differences among the
groups (P>0.05).
Discussion
As a common cause of vision loss, pathological NV
occurs in a variety of retinal/choroidal vascular diseases
(24). Retinal microvascular
pericytes are an important component of NV and serve an essential
role in the maintenance of microvascular integrity during
angiogenesis (25). It is widely
hypothesized that endothelial cell-derived PDGF-BB promotes
pericyte proliferation and migration by interacting with PDGFR-β in
pericytes, leading to retinal microvasculature development
(26–28). However, the precise mechanism
underlying the crosslinks between the SDF-1α/CXCR4/CXCR7 and
PDGF-BB/PDGFR-β axes in retinal microvascular pericytes remain
underreported. In the present study, it was first found that CXCR4
and CXCR7 are endogenously expressed by pericytes and demonstrated
further that PDGF-BB increased CXCR4 and CXCR7 expression by
activating STAT3 phosphorylation and subsequently potentiating
SDF-1α-stimulated cell viability and migration in retinal
microvascular pericytes.
Previously, Hamdan et al (16) found that suppression of the
SDF-1/CXCR4 axis reduced PDGF and inhibited tumor vascular
expansion in Ewing's sarcoma, whilst Song et al (17) discovered that PDGF overexpression
promoted pericyte content in A549 and MCF-7 tumor-bearing mice
through activation of the SDF-1/CXCR4 axis. In the present study,
CXCR4 and CXCR7 expression was studied in normal retinal
microvascular pericytes for the first time, which found that
PDGF-BB increased CXCR4 expression, consistent with Song's results.
In addition, PDGF-BB increased the expression of CXCR7, another
receptor for SDF-1α, in a similar manner to CXCR4.
The binding of PDGF-B to PDGFR activates a series of
intracellular signaling cascades that internalize the receptors,
targeting them for lysosomal degradation. Receptor endocytosis
provides a mechanism in which downstream signaling pathways,
including the ERK signaling (19),
AKT signaling pathway (20,21) and mitogen-activated protein kinase
(MAPK) signaling pathways (29) are
regulated. Endocytic transport of PDGFR-β has also been reported to
contribute to PDGF-induced STAT3 signal transduction (22,30–33).
STAT3 is an important class of cytoplasmic transcription factors.
The STAT3 pathway is involved in many physiological and
pathological processes, including cell proliferation, survival,
invasion and angiogenesis (34).
PDGFR is an epidermal and hepatocyte growth factor receptor, whilst
other growth factor receptors with tyrosine kinase activity can
also stimulate STAT3 phosphorylation (35–37). In
the present study, it was found that PDGF-BB increased
SDF-1α-treated up-regulation of CXCR4 and CXCR7 expression in
pericytes by activating the JAK/STAT3 signaling pathway rather than
the AKT or ERK signaling pathways.
The SDF-1α/CXCR4 axis participates in the
proliferation and migration of a number of different cell types. A
previous study in our laboratory found that the SDF-1α/CXCR4/CXCR7
axis is involved in regulating the proliferation and migration of
choroid-retinal endothelial cells (38). Zhao et al (39) also found that endothelial progenitor
cell proliferation was regulated by the SDF-1α/CXCR4 axis, whilst
Białopiotrowicz et al (40)
suggested that SDF-1/CXCR4 is involved in the migration of chronic
lymphocytic leukemia cells. In the present study, the SDF-1α/CXCR4
axis was found to participate in retinal microvascular pericyte
migration through interaction with the PDGF-BB/PDGFR-β axis.
However, Burns et al (41)
reported that CXCR4, instead of CXCR7, is involved in umbilical
vein endothelial cell migration because SDF-1 did not induce
Ca2+ influx through CXCR7. These discrepancies could be
explained by differing cell types and tumor microenvironments,
which require further investigation.
Further experiments are required to identify the
possible effects of PDGF-BB in regulating CXCR4 and CXCR7
expression in a hypoxia-induced ROP mouse model. Although the
present study supports a role for PDGF-BB-mediated regulation of
CXCR4 and CXCR7 expression in pericytes via the JAK/STAT3 signaling
pathway, there may be other pathways involved and many mechanisms
between endothelial cells and pericytes that do not involve CXCR4
or CXCR7 signaling. Indeed, previous studies from our laboratory
have suggested that transforming growth factor (TGF)-β and
lipopolysaccharide (LPS) promote CNV by upregulating CXCR4 and
CXCR7 levels in endothelial cells (14,15).
Therefore, whether there are crosslinks between signaling pathways
such as TGF-β or LPS and the PDGF-BB signaling pathway warrants
further investigation.
Altogether, findings from the present study show
that the PDGF-BB/PDGFR-β signaling pathway has a stimulatory effect
on the proliferative and migratory ability on SDF-1α-treated
retinal microvascular pericytes through upregulation of CXCR4 and
CXCR7 expression. Therefore, targeting CXCR4/CXCR7 signaling in
retinal microvascular pericytes may represent a potential therapy
avenue for restraining NV in fundus diseases.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
Beijing Bethune Charitable Foundation (grant no.
BJ-LM2015004J) and the National Nature Science Foundation of China
(grant nos. 81470637, 81600735 and 81873680) supported this
study.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
DX, YY and YF conceived and designed the experiments
of the current study. DX, XZ and RZ performed the experiments. DX,
JW and JS analyzed the data. DX and JS drafted the manuscript. XZ,
YF and YY revised it critically for important intellectual
content.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Joyal JS, Sun Y, Gantner ML, Shao Z, Evans
LP, Saba N, Fredrick T, Burnim S, Kim J, Patel G, et al: Retinal
lipid and glucose metabolism dictates angiogenesis through the
lipid sensor Ffar1. Nat Med. 22:439–445. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ciulla TA, Amador AG and Zinman B:
Diabetic retinopathy and diabetic macular edema: Pathophysiology,
screening, and novel therapies. Diabetes Care. 26:2653–2664. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang M, Chu S, Zeng F and Xu H:
Bevacizumab modulates the process of fibrosis in vitro. Clin Exp
Ophthalmol. 43:173–179. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Amoaku WM, Chakravarthy U, Gale R, Gavin
M, Ghanchi F, Gibson J, Harding S, Johnston RL, Kelly SP, Lotery A,
et al: Defining response to anti-VEGF therapies in neovascular AMD.
Eye (Lond). 29:721–731. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cao R, Brakenhielm E, Li X, Pietras K,
Widenfalk J, Ostman A, Eriksson U and Cao Y: Angiogenesis
stimulated by PDGF-CC, a novel member in the PDGF family, involves
activation of PDGFR-alphaalpha and -alphabeta receptors. FASEB J.
16:1575–1583. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hou X, Kumar A, Lee C, Wang B, Arjunan P,
Dong L, Maminishkis A, Tang Z, Li Y, Zhang F, et al: PDGF-CC
blockade inhibits pathological angiogenesis by acting on multiple
cellular and molecular targets. Proc Natl Acad Sci USA.
107:12216–12221. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Park DY, Lee J, Kim J, Kim K, Hong S, Han
S, Kubota Y, Augustin HG, Ding L, Kim JW, et al: Plastic roles of
pericytes in the blood-retinal barrier. Nat Commun. 8:152962017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Strittmatter K, Pomeroy H and Marneros AG:
Targeting platelet-derived growth factor receptor β(+) scaffold
formation inhibits choroidal neovascularization. Am J Pathol.
186:1890–1899. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jaffe GJ, Ciulla TA, Ciardella AP, Devin
F, Dugel PU, Eandi CM, Masonson H, Mones J, Pearlman JA,
Quaranta-El Maftouhi M, et al: Dual antagonism of PDGF and VEGF in
neovascular age-related macular degeneration: A phase Iib,
multicenter, randomized controlled trial. Ophthalmology.
124:224–234. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dunn EN, Hariprasad SM and Sheth VS: An
overview of the fovista and rinucumab trials and the fate of
anti-PDGF medications. Ophthalmic Surg Lasers Imaging Retina.
48:100–104. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee E and Rewolinski D: Evaluation of
CXCR4 inhibition in the prevention and intervention model of
laser-induced choroidal neovascularization. Invest Ophthalmol Vis
Sci. 51:3666–3672. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sengupta N, Caballero S, Mames RN, Timmers
AM, Saban D and Grant MB: Preventing stem cell incorporation into
choroidal neovascularization by targeting homing and attachment
factors. Invest Ophthalmol Vis Sci. 46:343–348. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Feng Y, Wang J, Yuan Y, Zhang X, Shen M
and Yuan F: miR-539-5p inhibits experimental choroidal
neovascularization by targeting CXCR7. Faseb J. 32:1626–1639. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Feng YF, Yuan F, Guo H and Wu WZ: TGF-β1
enhances SDF-1-induced migration and tube formation of
choroid-retinal endothelial cells by up-regulating CXCR4 and CXCR7
expression. Mol Cell Biochem. 397:131–138. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feng YF, Guo H, Yuan F and Shen MQ:
Lipopolysaccharide promotes choroidal neovascularization by
up-regulation of CXCR4 and CXCR7 expression in choroid endothelial
cell. PLoS One. 10:e01361752015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hamdan R, Zhou Z and Kleinerman ES:
Blocking SDF-1α/CXCR4 downregulates PDGF-B and inhibits bone
marrow-derived pericyte differentiation and tumor vascular
expansion in Ewing tumors. Mol Cancer Ther. 13:483–491. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Song N, Huang Y, Shi H, Yuan S, Ding Y,
Song X, Fu Y and Luo Y: Overexpression of platelet-derived growth
factor-BB increases tumor pericyte content via stromal-derived
factor-1alpha/CXCR4 axis. Cancer Res. 69:6057–6064. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak K and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gaceb A, Ozen I, Padel T, Barbariga M and
Paul G: Pericytes secrete pro-regenerative molecules in response to
platelet-derived growth factor-BB. J Cereb Blood Flow Metab.
38:45–57. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Arimura K, Ago T, Kamouchi M, Nakamura K,
Ishitsuka K, Kuroda J, Sugimori H, Ooboshi H, Sasaki T and Kitazono
T: PDGF receptor β signaling in pericytes following ischemic brain
injury. Curr Neurovasc Res. 9:1–9. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Moench R, Grimmig T, Kannen V, Tripathi S,
Faber M, Moll EM, Chandraker A, Lissner R, Germer CT Waaga-Gasser
AM and Gasser M: Exclusive inhibition of PI3K/Akt/mTOR signaling is
not sufficient to prevent PDGF-mediated effects on glycolysis and
proliferation in colorectal cancer. Oncotarget. 7:68749–68767.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang L, Shao J, Zhou Y, Chen H, Qi H,
Wang Y, Chen L, Zhu Y, Zhang M, Chen L, et al: Inhibition of
PDGF-BB-induced proliferation and migration in VSMCs by
proanthocyanidin A2: Involvement of KDR and Jak-2/STAT-3/cPLA2
signaling pathways. Biomed Pharmacother. 98:847–855. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lim R, Li L, Chew N and Yong EL: The
prenylflavonoid Icaritin enhances osteoblast proliferation and
function by signal transducer and activator of transcription factor
3 (STAT-3) regulation of C-X-C chemokine receptor type 4 (CXCR4)
expression. Bone. 105:122–133. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu CH, Wang Z, Sun Y and Chen J: Animal
models of ocular angiogenesis: From development to pathologies.
FASEB J. 31:4665–4681. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Motiejūnaite R and Kazlauskas A: Pericytes
and ocular diseases. Exp Eye Res. 86:171–177. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Genové G, Mollick T and Johansson K:
Photoreceptor degeneration, structural remodeling and glial
activation: A morphological study on a genetic mouse model for
pericyte deficiency. Neuroscience. 279:269–284. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chantrain CF, Henriet P, Jodele S, Emonard
H, Feron O, Courtoy PJ, DeClerck YA and Marbaix E: Mechanisms of
pericyte recruitment in tumour angiogenesis: A new role for
metalloproteinases. Eur J Cancer. 42:310–318. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tomkowicz B, Rybinski K, Sebeck D, Sass P,
Nicolaides NC, Grasso L and Zhou Y: Endosialin/TEM-1/CD248
regulates pericyte proliferation through PDGF receptor signaling.
Cancer Biol Ther. 9:908–915. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yokota J, Chosa N, Sawada S, Okubo N,
Takahashi N, Hasegawa T, Kondo H and Ishisaki A: PDGF-induced
PI3K-mediated signaling enhances the TGF-β-induced osteogenic
differentiation of human mesenchymal stem cells in a
TGF-β-activated MEK-dependent manner. Int J Mol Med. 33:534–542.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li L, Xu M, Li X, Lv C, Zhang X, Yu H,
Zhang M, Fu Y, Meng H and Zhou J: Platelet-derived growth factor-B
(PDGF-B) induced by hypoxia promotes the survival of pulmonary
arterial endothelial cells through the PI3K/Akt/Stat3 pathway. Cell
Physiol Biochem. 35:441–451. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lennartsson J, Ma H, Wardega P, Pelka K,
Engstrom U, Hellberg C and Heldin CH: The Fer tyrosine kinase is
important for platelet-derived growth factor-BB-induced signal
transducer and activator of transcription 3 (STAT3) protein
phosphorylation, colony formation in soft agar, and tumor growth in
vivo. J Biol Chem. 288:15736–15744. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Simeone-Penney MC, Severgnini M, Rozo L,
Takahashi S, Cochran BH and Simon AR: PDGF-induced human airway
smooth muscle cell proliferation requires STAT3 and the small
GTPase Rac1. Am J Physiol Lung Cell Mol Physiol. 294:L698–L704.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vij N, Sharma A, Thakkar M, Sinha S and
Mohan RR: PDGF-driven proliferation, migration, and IL8 chemokine
secretion in human corneal fibroblasts involve JAK2-STAT3 signaling
pathway. Mol Vis. 14:1020–1027. 2008.PubMed/NCBI
|
|
34
|
Furtek SL, Backos DS, Matheson CJ and
Reigan P: Strategies and approaches of targeting STAT3 for cancer
treatment. ACS Chem Biol. 11:308–318. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lo HW, Hsu SC, Ali-Seyed M, Gunduz M, Xia
W, Wei Y, Bartholomeusz G, Shih JY and Hung MC: Nuclear interaction
of EGFR and STAT3 in the activation of the iNOS/NO pathway. Cancer
Cell. 7:575–589. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yu YC, Yang PM, Chuah QY, Huang YH, Peng
CW, Lee YJ and Chiu SJ: Radiation-induced senescence in
securin-deficient cancer cells promotes cell invasion involving the
IL-6/STAT3 and PDGF-BB/PDGFR pathways. Sci Rep. 3:16752013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Levy DE and Darnell JE Jr: Stats:
Transcriptional control and biological impact. Nat Rev Mol Cell
Bio. 3:651–662. 2002. View
Article : Google Scholar
|
|
38
|
Jin J, Zhao WC and Yuan F:
CXCR7/CXCR4/CXCL12 axis regulates the proliferation, migration,
survival and tube formation of choroid-retinal endothelial cells.
Ophthalmic Res. 50:6–12. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhao Z, Ma X, Ma J, Sun X, Li F and Lv J:
Naringin enhances endothelial progenitor cell (EPC) proliferation
and tube formation capacity through the CXCL12/CXCR4/PI3K/Akt
signaling pathway. Chem Biol Interact. 286:45–51. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bialopiotrowicz E, Gorniak P,
Noyszewska-Kania M, Pula B, Makuch-Lasica H, Nowak G, Bluszcz A,
Szydlowski M, Jabłonska E, Piechna K, et al:
Microenvironment-induced PIM kinases promote CXCR4-triggered mTOR
pathway required for chronic lymphocytic leukaemia cell migration.
J Cell Mol Med. 22:3548–3559. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Burns JM, Summers BC, Wang Y, Melikian A,
Berahovich R, Miao Z, Penfold ME, Sunshine MJ, Littman DR, Kuo CJ,
et al: A novel chemokine receptor for SDF-1 and I-TAC involved in
cell survival, cell adhesion, and tumor development. J Exp Med.
203:2201–2213. 2006. View Article : Google Scholar : PubMed/NCBI
|