Introduction
Gestational diabetes mellitus (GDM) is a common
metabolic condition during pregnancy that is characterized by
glucose intolerance, leading to varying degrees of hyperglycemia
(1,2). GDM accounts for >80% of pregnant
women with diabetes, whilst pregnancies complicated by
non-gestational diabetes are <20% (3). The incidence of GDM varies around the
world due to the use of varying diagnostic criteria and
environmental factors (4,5). However, incidences of GDM in China had
been increasing annually with economic development and changes in
lifestyle (6,7). In 2017, 21.3 million or 16.2% of live
births had some form of hyperglycaemia in pregnancy with ~85.1% due
to GDM. (8). The adverse effects of
GDM on pregnant women and infants should not be underestimated as
hyperglycemia during pregnancy increases the risk of adverse
outcomes in the mother and fetus (9). In particular, infants born from
pregnant women with GDM are more susceptible to obesity, metabolic
and cardiovascular complications during childhood and adulthood
(10,11). In addition, ~50% of women with a
history of GDM go on to develop type 2 diabetes within five to ten
years post-delivery (8). Therefore,
GDM research has become a subject of interest in recent years
globally.
Currently, oral glucose tolerance test is a commonly
used clinical diagnostic tool for GDM. However, this test is
usually performed between 24 and 28 or 32 weeks of gestation
(12). This timeframe restricts
pregnant women from altering diet and exercise regimens, or to
access medical interventions; hence increasing the risk of GDM in
late pregnancy. Therefore, there is an urgent demand for developing
novel diagnostic approaches for the accurate and rapid prediction
of GDM.
Long non-coding RNA (lncRNAs) is a type of
functional RNA molecule consisting of >200 nucleotides, which
lacks the ability to encode proteins (13). LncRNAs can regulate protein-coding
genes in a number of ways, including forming complexes with target
genes, as well as promoting the degradation and inhibiting the
translation and modification of target microRNAs (miRNA). LncRNAs
can regulate the expression of genes at epigenetic, transcriptional
and post-transcriptional levels, and are widely involved in most
physiological and pathological processes in the body (14). Studies have suggested that the
aberrant expression of lncRNAs is closely associated with the
development of complex diseases, including cancer, cardiovascular
diseases, nervous system diseases and diabetes (15–18).
LncRNA maternally expressed gene 3 (MEG3) is a known
imprinted gene that is ~1.6 kbp in length and located on human
chromosome 14q32.2 (19). Imprinting
of this gene is controlled by upstream intergenic differentially
methylated region (20). In recent
years, a number of studies have demonstrated that lncRNA MEG3 is
involved in the development and progression of diseases, including
retinopathy, osteoarthritis and cardiovascular disease (21–23). It
was previously found that lncRNA MEG3 may serve a role in
micro-vascular dysfunction that is associated with diabetes
(24). In addition, the upregulation
of lncRNA MEG3 promotes insulin resistance in the liver by
increasing forkhead box O1 expression (25). In mouse pancreatic β-cells, lncRNA
MEG3 promotes the expression of v-maf musculoaponeurotic
fibrosarcoma oncogene family protein A and affects insulin
production by inhibiting the expression of RAD21 cohesin complex
component, structural maintenance of chromosomes 3 or
transcriptional regulator SIN3A (26). These studies indicate that lncRNA
MEG3 is closely associated with insulin resistance. However, the
role of lncRNA MEG3 in the development of GDM and its potential
molecular mechanism remain unclear. Therefore, the present study
aimed to investigate the expression and role of lncRNA MEG3 in GDM,
and to explore any potential underlying mechanism.
Materials and methods
Clinical samples
A total of 20 paired blood samples and matched
placental villous tissues were collected from pregnant women
between 23 and 37 years old with or without GDM at Weifang People's
Hospital (Weifang, China) between May 2015 and May 2017. Fasting
peripheral blood was collected from all pregnant women following 28
weeks of gestation, whilst the placental tissues were obtained
after delivery. Pregnant women with the following conditions were
excluded from the present study: i) Abnormal blood lipid
(triglyceride, low-density lipoprotein cholesterol and high-density
lipoprotein cholesterol) levels, hypertension, and chronic liver
and kidney diseases; ii) endocrine diseases, including, thyroid
disease, adrenal cortical disease, obesity, osteoporosis, diabetes
and hyperthyroidism prior to pregnancy; iii) pregnant women
currently undergoing long-term drug treatments (such as
sodiumlevothyroxine) that affect the metabolism of carbohydrates;
and iv) other pregnancy complications (pregnancy-induced
hypertension, pre-eclampsia, pregnancy with chronic nephritis). GDM
was diagnosed if the fasting plasma glucose (FPG) level was ≥5.1
mmol/l. GDM was excluded if the FPG level was ≤4.4 mmol/l. Women
with an FPG level ≥4.4 mmol/l but ≤5.1 mmol/l underwent a 75 g oral
glucose tolerance test (OGTT). In such cases, a diagnosis of GDM
was made when at least one glucose value was elevated (FPG ≥5.1
mmol/l, 1-h OGTT ≥10.0 mmol/l or 2-h OGTT ≥8.5 mmol/l). Inclusion
criteria: Women with DGM who did not fulfill any of the exclusion
criteria were involved in the present study. Written informed
consent was obtained from each patient and the present study was
approved by the Ethics Committee of Weifang People's Hospital.
Cell culture
HTR-8/SVneo cells, a human chorionic trophoblast
cell line, were purchased from Shanghai Huzhen Industrial Co., Ltd.
(cat. no. HZ-CC337655). Cells were cultured in RPMI 1640 medium
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) and 1%
streptomycin-penicillin solution (Beyotime Institute of
Biotechnolohy), and incubated in a humidified atmosphere under 5%
CO2 at 37°C.
Cell transfection
The MEG3 sequence was synthesized based on the MEG3
sequence and then sub-cloned into the pcDNA3.1 vector (pcDNA-MEG3;
Shanghai GeneChem Co., Ltd.). The empty pcDNA3.1 vector was used as
a control (pcDNA-control). For MEG3 knockdown, the
MEG3-shRNA plasmid (sh-MEG3 sequence,
5′-GAGAGGTTGTTTCACTGGTATCTATTGCA-3′; pGFP-C-shLenti Vector; cat.
no. TL 320132C) and the scrambled shRNA plasmid (sh-control;
pGFP-C-shLenti Vector; cat. no. TR30021) were purchased from
OriGene. HTR-8/SVneo cells were seeded into 6-well plates
(1×106 cells/well) and cultured at 37°C for 24 h. The
cells were subsequently transfected with 100 ng pcDNA-control, 100
ng pcDNA-MEG3, 100 ng sh-control, 100 ng sh-MEG3, 100 nM miR-345-3p
inhibitor (cat. no. HmiR-AN0437-AM01; GeneCopoeia, Inc.), 100 nM
inhibitor control (cat. no. CmiR-AN0001-SN; GeneCopoeia, Inc.) or
100 ng sh-MEG3 + 100 nM miR-345-3p inhibitor using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocols.
Transfection efficiency was assessed using reverse
transcription-quantitative PCR (RT-qPCR) 48 h after
transfection.
Bioinformatics analyses
Bioinformatics analysis (http://starbase.sysu.edu.cn/index.php) (27) was performed to predict potential
binding sites between lnc-MEG3 and miR-345-3p using the
‘miRNA-lncRNA’ search function.
Luciferase reporter assay
The putative binding sequences of MEG3-wild type
(WT; 5′-CCAGAGCCTGGTTCAGGG-3′) and MEG3-mutant (MUT;
5′-AACCCGAAGGCGGACAAA-3′) were respectively cloned into a pmirGLO
vector (Promega Corporation). HTR-8/SVneo cells were co-transfected
with 50 nM either miR-345-3p mimics (HmiR0210-MR03; GeneCopoeia,
Inc.) or mimic control (cat. no. CmiR0001-MR03; GeneCopoeia, Inc.)
and 100 ng either MEG3-WT or MEG3-MUT using
Lipofectamine® 2000. Following 48 h transfection,
Dual-Luciferase Reporter Assay System (Promega Corporation) was
used to detect luciferase activity, according to the manufacturer's
protocols. All firefly luciferase activities were normalized to
Renilla luciferase activity.
Cell viability assay
HTR-8/SVneo cell viability (1×104
cells/well) was measured using MTT assay. Following 48 h
transfection, 20 µl MTT solution (0.5 mg/ml) was added into each
well. After 4 h incubation at 37°C, 150 µl dimethyl sulfoxide
(DMSO) (Sigma-Aldrich; Merck KGaA) was used to dissolve the
formazan crystals. Finally, cell viability was detected by
determining absorbance values at 570 nm using a
FLUOstar® Omega Microplate Reader (BMG Labtech
GmbH).
Apoptosis analysis
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) apoptosis detection kit [cat. no.
70-AP101-100; Multisciences (Lianke) Biotech Co., Ltd.] was used to
assess cell apoptosis. Briefly, 48 h after cell transfection,
HTR-8/SVneo cells (1×106) were collected though
centrifugation (1,000 × g) at 4°C for 5 min, then the cells were
stained using 5 µl Annexin V-FITC and 5 µl PI for 30 min at room
temperature in the dark. Lastly, apoptotic cells were analyzed
using a flow cytometer (BD Biosciences) with WinMDI soft-ware
(version 2.5; Purdue University Cytometry Laboratories; www.cyto.purdue.edu/flowcyt/software/Catalog.htm).
Transwell assay
The invasive and migratory abilities of HTR-8/SVneo
cells were measured using Transwell inserts (Corning Inc.) with or
without Matrigel (BD Biosciences), respectively. For cell invasion
assay, Transwell inserts were pre-coated with Matrigel and
incubated at 37°C for 5 h. Following 48 h transfection, HTR-8/SVneo
cells (2×104) suspended in 100 µl serum-free medium were
seeded into the upper chamber, whilst 500 µl medium supplemented
with 20% FBS was added into the lower chamber. The cells were then
incubated for 48-h at 37°C. The migratory or invasive cells were
subsequently fixed with 100% methanol at room temperature for 20
min, stained with 0.1% crystal violet at 37°C for 20 min and
counted under an inverted light microscope (magnification, ×100) in
five randomly-selected fields.
RNA extraction and RT-qPCR
Total RNA was extracted from blood, tissues and
cells using the TRIzol reagent (Thermo Fisher Scientific, Inc.)
according to manufacturer's protocols and the cDNA of miR-345-3p
was synthesized using the TaqMan™ miRNA reverse transcription kit
(Thermo Fisher Scientific, Inc.) according to manufacturer's
protocols. The temperature protocol for the reverse transcription
reaction consisted of primer annealing at 25°C for 5 min, cDNA
synthesis at 42°C for 60 min and termination at 80°C for 2 min.
miR-345-3p levels were quantified using TaqMan™ human MiRNA assay
kit (Thermo Fisher Scientific, Inc.) according to manufacturer's
protocols. cDNA synthesis and subsequent qPCR of lncRNA MEG3 were
performed using SYBR® Premix Ex Taq™ (Tli RNaseH Plus;
Takara Biotechnology Co., Ltd.) according to manufacturer's
protocols. The following thermocycling conditions were used for
PCR: Initial denaturation at 95°C for 45 sec, followed by 40 cycles
of 95°C for 10 sec and 52°C for 35 sec. U6 and GAPDH were used as
the endogenous controls for miR-345-3p and lncRNA MEG3,
respectively. The primer sequences used for qPCR were as follows:
lncRNA MEG3 forward, 5′-CTGCCCATCTACACCTCACG-3′ and reverse,
5′-CTCTCCGCCGTCTGCGCTAGGGGCT-3′; GAPDH forward,
5′-CTTTGGTATCGTGGAAGGACTC-3′ and reverse,
5′-GTAGAGGCAGGGATGATGTTCT-3′; U6 forward,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; miR-345-3p forward,
5′-GGTTTTTGGATTGGGTTGTAGAGTG-3′ and reverse,
5′-AACCAAAACAATCCCTTACCACTAC-3′. Relative gene expression was
calculated using the 2−ΔΔCq method (28).
Western blotting
Total protein was extracted from cells using the
radioimmunoprecipitation assay buffer kit (Thermo Fisher
Scientific, Inc.). Protein concentration was quantified using a
bicinchoninic acid assay kit (Thermo Fisher Scientific, Inc.) prior
to separation via SDS-PAGE on a 10% gel at 30 µg protein/lane. The
separated proteins were then transferred onto polyvinylidene
fluoride membranes and blocked with 5% non-fat milk at room
temperature for 1 h. Following blocking, the membranes were then
incubated with primary antibodies against Bcl-2 (cat no. 4223), Bax
(cat no. 5023) and β-actin (cat no. 4970; all dilution: 1:1,000;
Cell Signaling Technology, Inc.) at 4°C overnight. The membranes
were washed with phosphate buffer saline (PBS)-0.05% Tween 20 5
times and then incubated with a horseradish peroxidase-conjugated
anti-rabbit immunoglobulin G secondary antibody (cat no. 7074;
dilution: 1:2,000; Cell Signaling Technology, Inc.) at room
temperature for 2 h. The protein bands were visualized using an
enhanced chemiluminescence kit (Applygen Technologies, Inc.)
according to manufacturer's protocols. Densitometry was performed
using the ImageJ software (version 1.38X; National Institutes of
Health).
Statistical analysis
Data were presented as the mean ± SD. Statistical
analysis was performed using the SPSS software (version 18.0; SPSS,
Inc.). Differences between multiple groups were analyzed using
one-way ANOVA followed by Tukey's post-hoc test, and differences
between two groups were analyzed using Student's t-test. P<0.05
was considered to indicate a statistically significant
difference.
Results
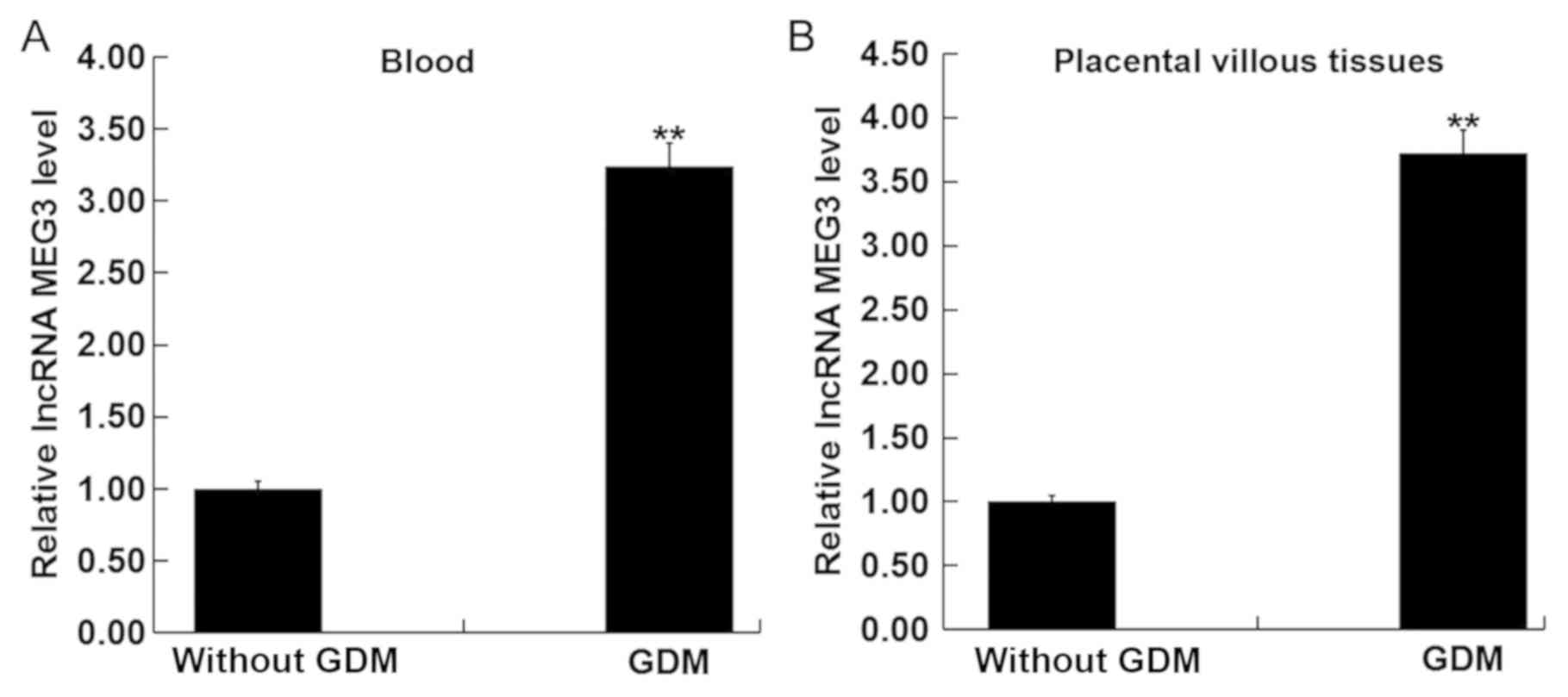
LncRNA MEG3 is highly expressed in
blood and placental villous tissues of pregnant women with GMD
The lncRNA MEG3 levels in the blood and placental
villous tissues of pregnant women were measured using RT-qPCR.
LncRNA MEG3 levels in the blood and placental villous tissues from
pregnant women with GDM were significantly upregulated compared
with those without GDM (Fig. 1A and
B). This suggest that lncRNA MEG3 may serve a role during
GDM.
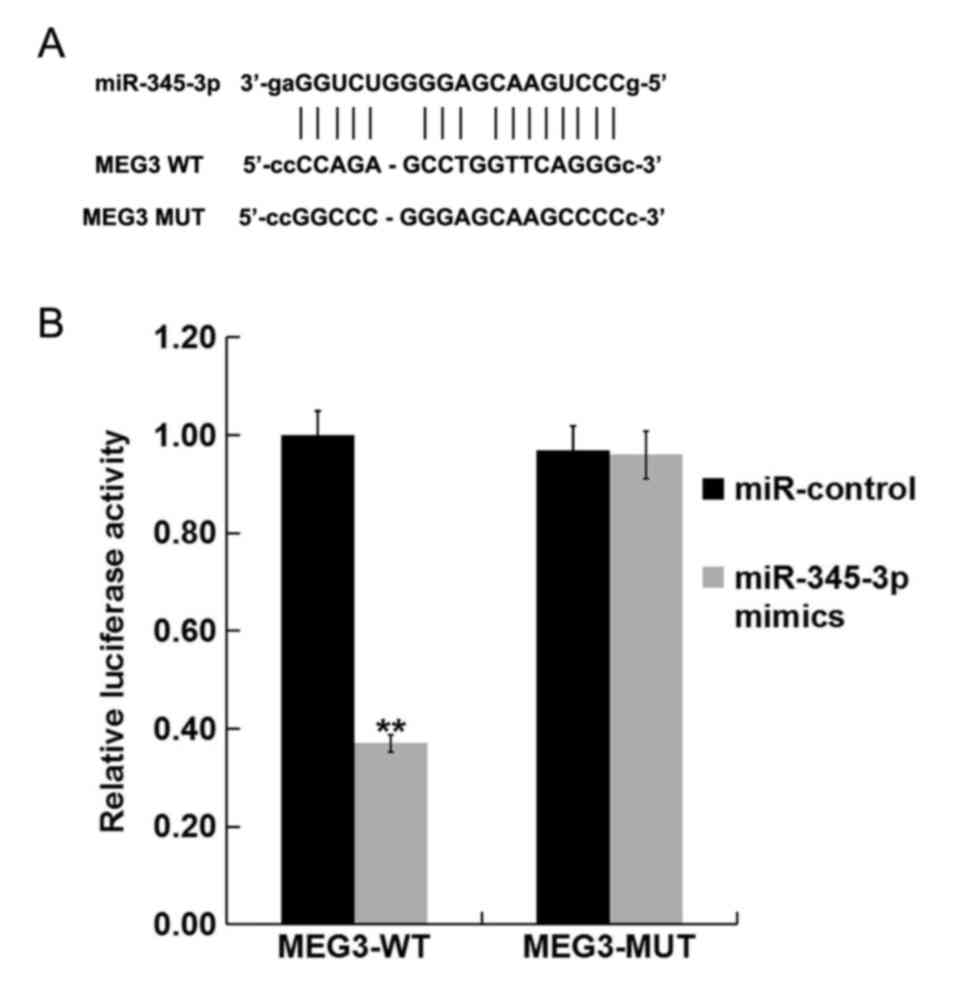
miR-345-3p is a target of lncRNA
MEG3
Bioinformatics analysis (http://starbase.sysu.edu.cn/index.php) showed
miR-345-3p may be a target of lncRNA MEG3 (Fig. 2A). Luciferase assay was used to
confirm whether lncRNA MEG3 could regulate miR-345-3p expression by
acting as molecular sponge. Luciferase activity was significantly
reduced in cells co-transfected with the miR-345-3p mimic and
MEG3-WT compared with cells co-transfected with mimic control
(miR-control) and MEG3-WT. However, no significant differences were
observed between the luciferase activity of cells co-transfected
with mimic control (miR-control) and MEG3-MUT and that in cells
co-transfected with the miR-345-3p mimic and MEG3-MUT (Fig. 2B). This finding suggests that
miR-345-3p is a direct target of lncRNA MEG3.
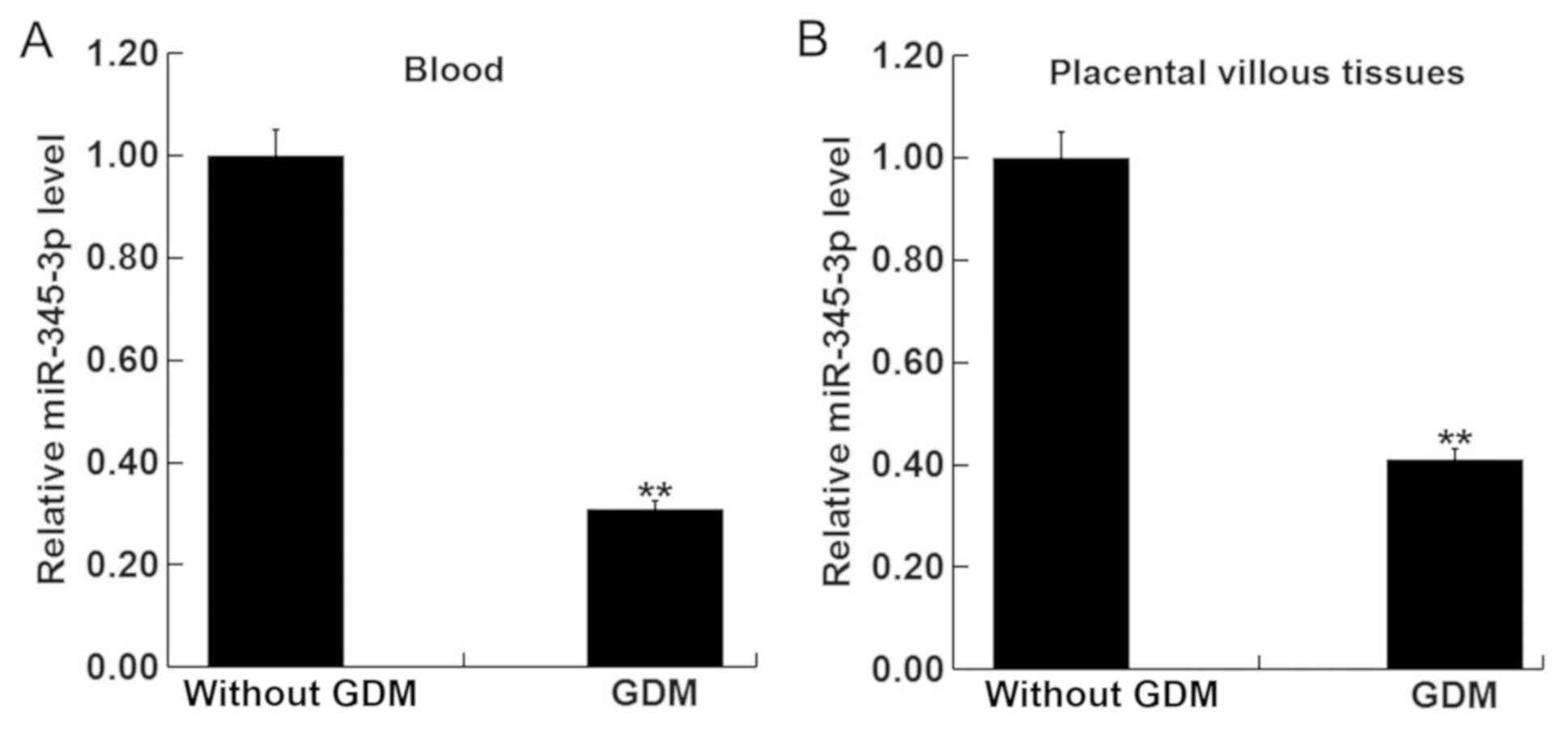
miR-345-3p is downregulated in the
blood and placental villous tissues of pregnant women with GMD
The levels of miR-345-3p in the blood and placental
villous tissues of pregnant women with or without GMD were next
measured using RT-qPCR. miR-345-3p levels in the blood (Fig. 3A) and placental villous tissues
(Fig. 3B) of women with GMD was
significantly reduced compared with pregnant women without GDM.
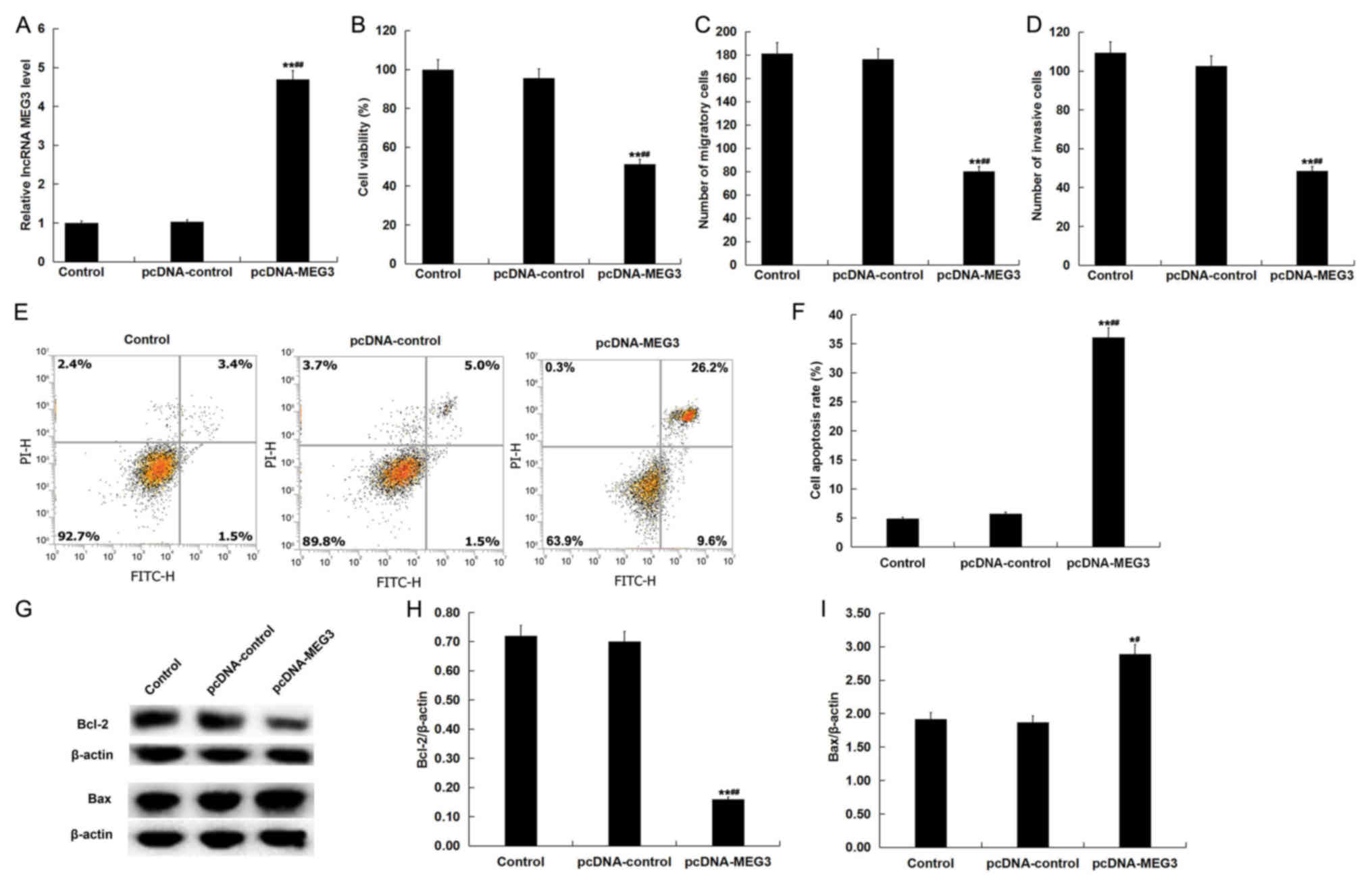
LncRNA MEG3 overexpression inhibits
viability, migration and invasion of placental trophoblast cells,
and induces apoptosis
The effects of lncRNA MEG3 overexpression on human
chorionic trophoblast cells (HTR-8/SVneo cells) was next
investigated by transfecting HTR-8/SVneo cells with pcDNA-control
or pcDNA-MEG3. Following 48-h transfection, efficiency was
evaluated using RT-qPCR. pcDNA-MEG3 transfection significantly
increased the levels of lncRNA MEG3 in HTR-8/SVneo cells compared
with cells in the control group and pcDNA control group (Fig. 4A). HTR-8/SVneo cell viability,
apoptosis, migratory and invasive ability were subsequently
analyzed using MTT, flow cytometry and Transwell assays,
respectively. LncRNA MEG3 overexpression significantly inhibited
HTR-8/SVneo cell viability (Fig.
4B), and prevented cell migration (Fig. 4C) and invasion (Fig. 4D), in addition to significantly
inducing cell apoptosis (Fig. 4E and
F) compared with cells in the control group and pcDNA control
group. Supporting this, it was also found that lncRNA MEG3
overexpression significantly decreased protein levels of Bcl-2
whilst increasing levels of Bax compared with the control group and
pcDNA control group (Fig. 4G-I).
LncRNA MEG3 knockdown promotes the
viability, migration and invasion of placental trophoblast cells,
and reduces apoptosis
The effects of lncRNA MEG3 knockdown on HTR-8/SVneo
cells was next investigated. HTR-8/SVneo cells were transfected
with sh-control, sh-MEG3, inhibitor control, miR-345-3p inhibitor
or sh-MEG3 + miR-345-3p inhibitor for 48 h prior to RT-qPCR
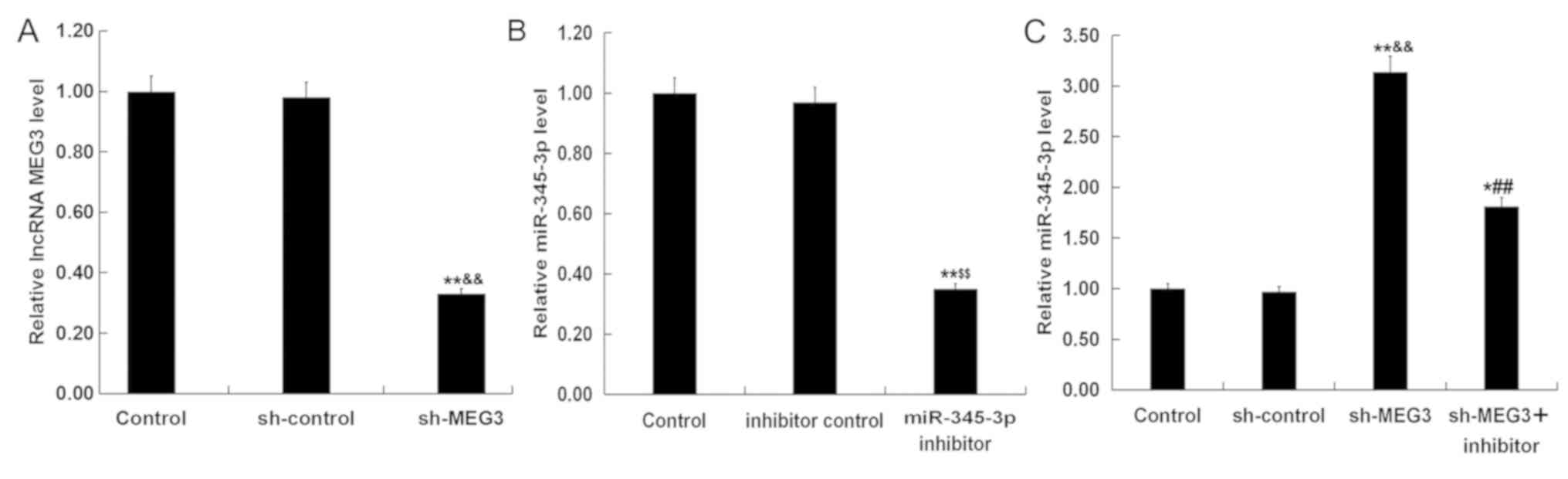
analysis to assess transfection efficiency. Compared with the
control group and sh-control group, sh-MEG3 significantly reduced
the levels of lncRNA MEG3 in HTR-8/SVneo cells (Fig. 5A), whereas the miR-345-3p inhibitor
significantly reduced miR-345-3p levels in HTR-8/SVneo cells
compared with the control group and inhibitor control group
(Fig. 5B). sh-MEG3 significantly
increased the levels of miR-345-3p in the HTR-8/SVneo cells
compared with the control group and sh-control group, which was
partially reversed by transfection with the miR-345-3p inhibitor
(Fig. 5C).
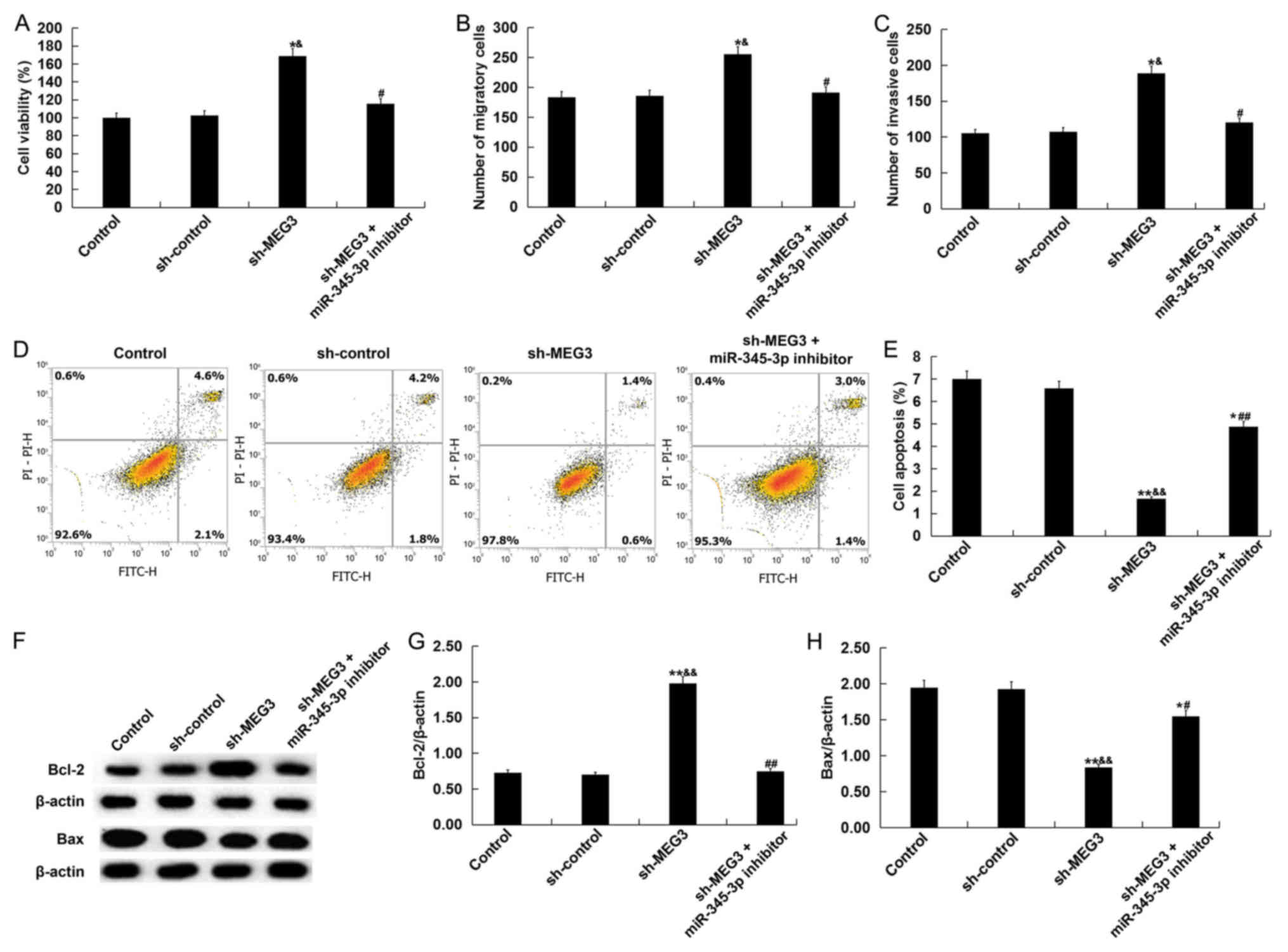
Results from MTT assay, Transwell assay, and flow
cytometry demonstrated that lncRNA MEG3 knockdown significantly
promoted HTR-8/SVneo cell viability (Fig. 6A), enhanced cell migration and
invasion (Fig. 6B and C) and reduced
cell apoptosis (Fig. 6D and E)
compared with the control group and sh-control group. In addition,
compared with the control group and sh-control group, lncRNA MEG3
knockdown significantly increased Bcl-2 protein expression whilst
reducing Bax protein expression (Fig.
6F-H). All the observed effects that lncRNA MEG3 knockdown
exerted on HTR-8/SVneo cells were negated by transfection with the
miR-345-3p inhibitor (Fig. 6).
Discussion
The present study demonstrated that the levels of
lncRNA MEG3 in the blood and placental villous tissues from
pregnant women with GDM was significantly upregulated compared with
the pregnant women without GDM. miR-345-3p was found to be a target
of lncRNA MEG3, which was downregulated in the blood and placental
villous tissues from pregnant women with GMD. LncRNA MEG3
overexpression inhibited the cell viability, migration and invasion
in placental trophoblast cells, and induced apoptosis, whilst
suppression of lncRNA MEG3 expression exhibited the opposite
effects. In addition, all the effects of lncRNA MEG3 knockdown
exerted on HTR-8/SVneo cells were reversed by the inhibition of
miR-345-3p expression. These findings suggest that lncRNA MEG3 may
be a novel diagnostic biomarker and therapeutic target for GDM.
Aberrant expression of lncRNAs is closely associated
with the development of complex diseases, including cancer,
cardiovascular diseases, nervous system diseases and diabetes
(16–19). lncRNA MEG3 has been previously
reported to participate in the development and progression of
diseases, including retinopathy, osteoarthritis and cardiovascular
disease (21–23). In particular, inhibition of DNA
methyltransferase 3b (DNMT3B) has been shown to upregulate MEG3
expression in HCC cells (29). In
recent years, a number of studies have suggested that lncRNAs
participate in the occurrence and development of GDM (30,31).
LncRNA MEG3, which is closely associated with insulin resistance in
the body, is involved in the occurrence and development of diabetes
(21–23). The pathogenesis of GDM is highly
analogous to that of type 2 diabetes (32,33).
Therefore, it was hypothesized in the present study that lncRNA
MEG3 may serve an important role in the development of GDM.
Firstly, the expression of lncRNA MEG3 in the blood
and placental tissues from pregnant women with GDM and non-GDM was
examined. LncRNA MEG3 was demonstrated to be highly expressed in
the blood and placental villous tissues from pregnant women with
GDM. It was subsequently found that miR-345-3p was a target of
lncRNA MEG3. miR-345-3p expression has been reported to be
downregulated in diabetes mellitus (34) and diabetic cardiomyopathy (35). In the present study, it was found
that miR-345-3p was downregulated in the blood and placental
villous tissues from pregnant women with GMD. These observations
suggested an important role for lncRNA MEG3 in GDM.
Placental trophoblasts serve an important role in
the process of blastocyst implantation in early pregnancy (36,37).
Trophoblast proliferation, apoptosis, invasion and migration are
key to the establishment, maintenance and finally timely
termination of physiological pregnancy (37,38). In
particular, placental trophoblast function has become the focus of
intense research on GDM pathogenesis (39). Therefore, the effect of lncRNA MEG3
on human chorionic trophoblast cell physiology was next
investigated using the HTR-8/SVneo cell line as model. LncRNA MEG3
overexpression inhibited HTR-8/SVneo cell viability, migration and
invasion in addition to inducing apoptosis; whilst the suppression
of lncRNA MEG3 expression exerted the opposite effects. Notably,
the effects lncRNA MEG3 knockdown exerted on HTR-8/SVneo cells were
negated by miR-345-3p downregulation. However, a limitation of the
present study was that a sh-MEG3 + inhibitor control group was not
evaluated.
Taken together, results from the present study
suggest that the levels of lncRNA MEG3 was significantly
upregulated in GDM, and it participated in the development and
progression of GDM. This was possibly mediated via the regulation
of human chorionic trophoblast cell physiology by targeting
miR-345-3p expression. Therefore, lncRNA MEG3 may be a diagnostic
and therapeutic target for GDM. However, the present study is only
a preliminary study on the role of lncRNA MEG3 in GDM in
vitro. To enhance the scientific significance of the results
from the present study, the relationship between the levels of
lncRNA MEG3 and blood glucose levels should be determined; in
addition to in vivo studies on the role of lncRNA MEG3 in
GDM development. These issues should be addressed further in any
future research.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data sets used and/or generated during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
HZ designed the study, performed all the experiments
and sample collection, analyzed the data and prepared the
manuscript.
Ethics approval and consent to
participate
Written informed consent was obtained from each
patient and this study was approved by the Ethics Committee of
Weifang People's Hospital (Weifang, China). Written informed
consent was obtained from each patient.
Patient consent for publication
Not applicable.
Competing interests
The author declares that they have no competing
interests.
References
|
1
|
Surapaneni T, Nikhat I and Nirmalan PK:
Diagnostic effectiveness of 75 g oral glucose tolerance test for
gestational diabetes in India based on the international
association of the diabetes and pregnancy study groups guidelines.
Obstet Med. 6:125–128. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Djelmis J, Pavic M, Mulliqi Kotori V,
Pavlić Renar I, Ivanisevic M and Oreskovic S: Prevalence of
gestational diabetes mellitus according to IADPSG and NICE
criteria. Int J Gynaecol Obstet. 135:250–254. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liao EY: Endocrine and metabolic diseases.
Beijing. People's Medical Publishing House. 2012.5 ISBN:
9787117151153. https://baike.so.com/doc/
|
|
4
|
Egan AM, Vellinga A, Harreiter J, Simmons
D, Desoye G, Corcoy R, Adelantado JM, Devlieger R, Van Assche A,
Galjaard S, et al: Epidemiology of gestational diabetes mellitus
according to IADPSG/WHO 2013 criteria among obese pregnant women in
Europe. Diabetologia. 60:1913–1921. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Song L, Shen L, Li H, Liu B, Zheng X,
Zhang L, Xu S and Wang Y: Socio-economic status and risk of
gestational diabetes mellitus among Chinese women. Diabet Med.
34:1421–1427. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang F, Dong L, Zhang CP, Li B, Wen J,
Gao W, Sun S, Lv F, Tian H, Tuomilehto J, et al: Increasing
prevalence of gestational diabetes mellitus in Chinese women from
1999 to 2008. Diabet Med. 28:652–657. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Leng J, Shao P, Zhang C, Tian H, Zhang F,
Zhang S, Dong L, Li L, Yu Z, Chan JC, et al: Prevalence of
gestational diabetes mellitus and its risk factors in Chinese
pregnant women: A prospective population-based study in Tianjin,
China. PLoS One. 10:e01210292015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cho NH, Whitng D, Forouhi N, et al: IDF
diabetes atlas, 8th edn. Brussels. Internatonal Diabetes Federaton.
2017.
|
|
9
|
Mitanchez D, Yzydorczy C, Siddeek B,
Boubred F, Benahmed M and Simeoni U: The offspring of the diabetic
mother-short- and long-term implications. Best Pract Res Clin
Obstet Gynaecol. 29:256–269. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bowers K, Laughon SK, Kiely M, Brite J,
Chen Z and Zhang C: Gestational diabetes, pre-pregnancy obesity and
pregnancy weight gain in relation to excess fetal growth:
Variations by race/ethnicity. Diabetologia. 56:1263–1271. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kristensen P, Susser E, Irgens LM, Mehlum
IS, Corbett K and Bjerkedal T: The association of high birth weight
with intelligence in young adulthood: A cohort study of male
siblings. Am J Epidemiol. 180:876–884. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tieu J, McPhee AJ, Crowther CA and
Middleton P: Screening and subsequent management for gestational
diabetes for improving maternal and infant health. Cochrane
Database Syst Rev CD007222. 2014. View Article : Google Scholar
|
|
13
|
Kung JT, Colognori D and Lee JT: Long
noncoding RNAs: Past, present, and future. Genetics. 193:651–669.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sanchez Y and Huarte M: Long non-coding
RNAs: Challenges for diagnosis and therapies. Nucleic Acid Ther.
23:15–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wapinski O and Chang HY: Long noncoding
RNAs and human disease. Trends Cell Biol. 21:354–361. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li CH and Chen Y: Targeting long
non-coding RNAs in cancers: Progress and prospects. Int J Biochem
Cell Biol. 45:1895–1910. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Carter G, Miladinovic B, Patel AA, Deland
L, Mastorides S and Patel NA: Circulatng long noncoding RNA GAS5
levels are correlated to prevalence of type 2 diabetes mellitus.
BBA Clin. 4:102–107. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xie X, Tang B, Xiao YF, Xie R, Li BS, Dong
H, Zhou JY and Yang SM: Long non-coding RNAs in colorectal cancer.
Oncotarget. 7:5226–5239. 2016.PubMed/NCBI
|
|
20
|
Lin SP, Youngson N, Takada S, Seitz H,
Reik W, Paulsen M, Cavaille J and Ferguson-Smith AC: Asymmetric
regulation of imprinting on the maternal and paternal chromosomes
at the Dlk1-Gtl2 imprinted cluster on mouse chromosome 12. Nat
Genet. 35:97–102. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu J and Xu Y: The lncRNA MEG3
downregulation leads to osteoarthritis progression via miR-16/SMAD7
axis. Cell Biosci. 7:692017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhan R, Xu K, Pan J, Xu Q, Xu S and Shen
J: Long noncoding RNA MEG3 mediated angiogenesis after cerebral
infarction through regulating p53/NOX4 axis. Biochem Biophys Res
Commun. 490:700–706. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qiu GZ, Tian W, Fu HT, Li CP and Liu B:
Long noncoding RNA-MEG3 is involved in diabetes mellitus-related
microvascular dysfunction. Biochem Biophys Res Commun. 471:135–141.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu X, Wu YB, Zhou J and Kang DM:
Upregulation of lncRNA MEG3 promotes hepatic insulin resistance via
increasing FoxO1 expression. Biochem Biophys Res Commun.
469:319–325. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang N, Zhu Y, Xie M, Wang L, Jin F, Li Y,
Yuan Q and De W: Long noncoding RNA Meg3 regulates mafa expression
in mouse beta cells by inactivating Rad21, Smc3 or Sin3α. Cell
Physiol Biochem. 45:2031–2043. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu J, Wu J, Zhao Z, Wang J and Chen Z:
Circulating LncRNA serve as fingerprint for gestational diabetes
mellitus associated with risk of macrosomia. Cell Physiol Biochem.
48:1012–1018. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Y, Ren M, Zhao Y, Lu X, Wang M, Hu J,
Lu G and He S: MicroRNA-26a inhibits proliferation and metastasis
of human hepatocellular carcinoma by regulating DNMT3B-MEG3 axis.
Oncol Rep. 37:3527–3535. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang Y, Wu H, Wang F, Ye M, Zhu H and Bu
S: Long non-coding RNA MALAT1 expression in pregnant women with
gestational diabetes mellitus. Int J Gynaecol Obstet. 140:164–169.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Buchanan TA, Xiang A, Kjos SL and Watanabe
R: What is gestatonal diabetes? Diabetes Care. 30 (Suppl
2):S105–S111. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
American Diabetes Association (ADA), .
Standards of medical care in diabetes. Diabetes Care. 37:14–80.
2014. View Article : Google Scholar
|
|
33
|
Loegl J, Nussbaumer E, Cvitic S, Huppertz
B, Desoye G and Hiden U: GDM alters paracrine regulation of
feto-placental angiogenesis via the trophoblast. Lab Invest.
97:409–418. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Esteves JV, Yonamine CY, Pinto-Junior DC,
Gerlinger-Romero F, Enguita FJ and Machado UF: Diabetes modulates
MicroRNAs 29b-3p, 29c-3p, 199a-5p and 532-3p expression in muscle:
Possible role in GLUT4 and HK2 repression. Front Endocrinol
(Lausanne). 9:5362018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chavali V, Tyagi SC and Mishra PK:
Differential expression of dicer, miRNAs, and inflammatory markers
in diabetic Ins2+/- Akita hearts. Cell Biochem Biophys. 68:25–35.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ji L, Brkić J, Liu M, Fu G, Peng C and
Wang YL: Placental trophoblast cell differentiation: Physiological
regulation and pathological relevance to preeclampsia. Mol Aspects
Med. 34:981–1023. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Weiss G, Sundl M, Glasner A, Huppertz B
and Moser G: The trophoblast plug during early pregnancy: A deeper
insight. Histochem Cell Biol. 146:749–756. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang L, Li Y, Wang C, Li N, Hou Y, Chang
R, Sun M, Wang R, Zhu L and Qiao C: Overexpression of collapsin
response mediator protein 1 inhibits human trophoblastcells
proliferation, migration, and Invasion. Reprod Sci. 26:954–960.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li G, Lin L, Wang YL and Yang H:
1,25(OH)2D3 protects trophoblasts against insulin resistance and
inflammation via suppressing mTOR signaling. Reprod Sci.
26:223–232. 2019. View Article : Google Scholar : PubMed/NCBI
|