Introduction
Ovarian cancer is one of the most common causes of
gynecological reproductive system tumors (1). There are approximately 299,000 new
cases of ovarian cancer and 152,000 related deaths each year,
accounting for the seventh highest incidence among female malignant
tumors (2). Most patients are
already at an advanced stage by the time of diagnosis, and the
5-year survival rate of advanced ovarian cancer is less than 30%
(3). Only 15% of patients are
diagnosed at an early stage (stage 1) with a 5-year survival rate
of 92% (4). In recent years, much
research has focused on the development of new potential ovarian
cancer targeted therapies (5–7).
Long non-coding RNAs (lncRNAs) are a class of RNAs
with a length of more than 200 nucleotides that lack a meaningful
open reading frame (8). LncRNAs
participate extensively in embryonic development, apoptosis, cell
cycle regulation, tumor formation and other biological processes
(9–12). Previous studies have found that
lncRNA plays an important role in tumor metastasis, and abnormal
expression of many lncRNAs is directly related to tumor metastasis
or recurrence (13–15). LncRNA activated by transforming
growth factor-β (lncRNA-ATB) is part of a family of lncRNAs, which
can regulate the invasion and metastasis of various tumor cells,
such as intestinal cancer and liver cancer, by regulating cell
phenotype switching (16–18). However, there have been no reports on
the role of lncRNA-ATB in ovarian cancer. This study therefore
aimed to investigate the role of lncRNA-ATB in ovarian cancer cells
and to further explore the underlying mechanisms of its action.
Materials and methods
Cell culture and treatments
Ovarian cancer cell line SKOV3 (cat no.
ATCC® HTB-77™) was obtained from American Type Culture
Collection. Non-tumorous human ovarian surface epithelial cells
(HOSEpiC; cat no. BNCC340096) were purchased from the BeNa Culture
Collection (Suzhou Bei Na Chuanglian Biotechnology Co., Ltd.).
SKOV3 and HOSEpiC cells were cultured in complete Dulbecco's
Modified Eagle Medium (DMEM)/nutrient mixture F12 (Gibco, Thermo
Fisher Scientific, Inc.) including 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin-streptomycin solution at 37°C
in a 5% CO2 incubator.
Cell transfection
LncRNA-ATB-shRNA and the negative control of
lncRNA-ATB-shRNA (lncRNA-ATB-NC) were purchased from Biomics
(Biomics Biotechnologies, Co., Ltd.). A total of 1 µg LncRNA-ATB-NC
or 1 µg lncRNA-ATB-shRNA were transfected into SKOV3 cells using
Lipofectamine 2000 (Invitrogen, Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Total RNA was
extracted from the cells 48 h after transfection, and the
expression of lncRNA-ATB was detected using reverse
transcription-quantitative PCR (RT-qPCR) to determine the
transfection efficiency. Cells without any treatment Were
considered as the control.
CCK-8 assay
The CCK-8 method was used to determine SKOV3 cell
proliferative ability. A cell suspension at a concentration of
1×104/ml was prepared and 100 µl of this suspension was
added to each well of a 96-well plate. Cells were then cultured for
12, 24 or 48 h, respectively, before 10 µl CCK-8 reagent
(Sigma-Aldrich; Merck KGaA) was added to each well. After 2 h of
incubation absorbance at a wavelength of 450 nm was measured using
the FLUOstar® Omega Microplate Reader (BMG Labtech
GmbH). This experiment was repeated three times.
Flow cytometry analysis
SKOV3 cells of different groups (control: Cells
without any treatment, lncRNA-ATB-NC and lncRNA-ATB-shRNA
respectively) were harvested using 0.2% trypsin, washed with
phosphate buffered saline (PBS) and then fixed with 70% ethanol
overnight at 4°C. The number of apoptotic cells was quantified
using the AnnexinV-FITC/PI kit (cat no. 70-AP101-100;
MultiSciences) according to the manufacturer's instructions. Cell
apoptosis rate was measured using a FACS Calibur flow cytometer (BD
Biosciences) and the data were analyzed using FlowJo software
(version 7.6.1; FlowJo LLC). The assay was performed in
triplicate.
Transwell assay
A transwell invasion assay was used to determine
cell invasion ability. Polycarbonate filters (8-µm pore size;
Corning Inc.) with Matrigel (BD Biosciences) were used in the
transwell migration assay. A total of 200 µl DMEM (supplemented
with 0.1% FBS) containing 1×105 SKOV3 cells was added to
the upper chamber while 600 µl DMEM supplemented with 10% FBS was
added to the lower chamber. After incubation for 48 h, any SKOV3
cells that had invaded into the lower chamber were fixed with 100%
methanol at room temperature for 20 min, stained with 0.1% crystal
violet at 37°C for 20 min, and counted using an upright microscope
(magnification, ×200; five randomly-selected fields per chamber).
Each transwell assay was repeated in five independent
experiments.
Scratch wound healing assay
SKOV3 cells from different treatment groups
(control, lncRNA-ATB-NC and lncRNA-ATB-shRNA, respectively) were
seeded onto six-well plates (5×105 cells/well) and
cultured until they reached 80–90% confluence. Scratch wounds were
created in the monolayer of confluent SKOV3 cells using a 200 µl
pipette tip. The wounded cells were then incubated at 37°C with 5%
CO2 for 24 h. Wound healing was measured using ImageJ
software version 1.46 (National Institutes of Heath) and images
were captured using a phase-contrast microscope at two time points,
immediately after scratching and after 24-h incubation. Data were
presented as wound width (at 24 h)/wound width (at 0 h) ×100%.
RT-qPCR
Total RNA was isolated from cells using an EASYspin
Plus tissue/cell RNA extraction kit (Aidlab Biotechnologies, Co.
Ltd.) according to the manufacturer's instructions. RNA was
reverse-transcribed into cDNA using a Transcriptor First Strand
cDNA Synthesis kit (Roche Diagnostics) following the manufacturer's
instructions. qPCR was performed on a LightCycler 480 system (Roche
Diagnostics) using Fast SYBR Green Master Mix (Roche Diagnostics)
as per the manufacturer's instructions. The sequences of RT-qPCR
primers were obtained as required and listed as following: GAPDH
forward, 5′-CTTTGGTATCGTGGAAGGACTC-3′; reverse,
5′-GTAGAGGCAGGGATGATGTTCT-3′; lncRNA- ATB forward,
5′-TCTGGCTGAGGCTGGTTGAC-3′; reverse, 5′-ATCTCTGGGTGCTGGTGAAGG-3′;
STAT3 forward, 5′-ACAGCAGGATGGCCAGGTTGC-3′; reverse,
5′-TCTGTCTGGTGGCTGCTGCCT-3′; E-Cadherin forward,
5′-AGCCATGTACGTTGCTATCC-3′; reverse, 5′-CGTAGCACAGCTTCTCCTTAAT-3′;
Vimentin forward, 5′-GCTGCAGGCCCAGATTCA-3′; reverse,
5′-TTCATACTGCTGGCGCACAT-3′. The following thermocycling conditions
were used for PCR: Initial denaturation at 95°C for 45 sec,
followed by 40 cycles of 95°C for 10 sec and 52°C for 35 sec. The
relative expression ratio of target genes was calculated using the
2−∆∆Cq method (19).
Western blotting
Cells were washed twice with PBS, then lysed in RIPA
buffer (cat. no. P0013B; Beyotime Institute of Biotechnology).
Protein concentration was measured using a BCA assay kit (Thermo
Fisher Scientific, Inc.). The cell lysates (30 µg/lane) were
separated on 12% SDS polyacrylamide gels and transferred onto PVDF
membranes (Bio-Rad Laboratory, Inc.). After blocking of
non-specific binding with TBS-T (0.1% Tween) containing 5% non-fat
milk for 1 h at room temperature, the membranes were incubated with
the primary antibodies at 4°C overnight. Then, the membranes were
incubated with horseradish peroxidase (HRP)-conjugated goat
anti-rabbit secondary antibody (cat no. 7074; 1:2,000; Cell
Signaling Technology, Inc.) for 2 h at room temperature. Finally,
the protein bands were detected using the ChemiDOC™ system
(Bio-Rad, Hercules, CA) following the manufacturer's protocol. The
primary antibodies: Anti-STAT3 (1:1,000; cat no. 12640),
anti-p-STAT3 (1:1,000; cat no. 9145), anti-E-cadherin (1:1,000; cat
no. 3195), anti-vimentin (1:1,000; cat no. 5741) and anti-GAPDH
(cat no. 5174) were purchased from Cell Signaling Technology, Inc.
Protein bands were analyzed using ImageJ version 1.49 software
(National Institute of Health).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism version 5.0 (GraphPad Software, Inc.). The tests performed
were one-way ANOVA followed by Tukey's post-hoc test or a Student's
t-test as appropriate. Data are presented as the mean ± SD.
P<0.05 was considered to indicate a statistically significant
difference.
Results
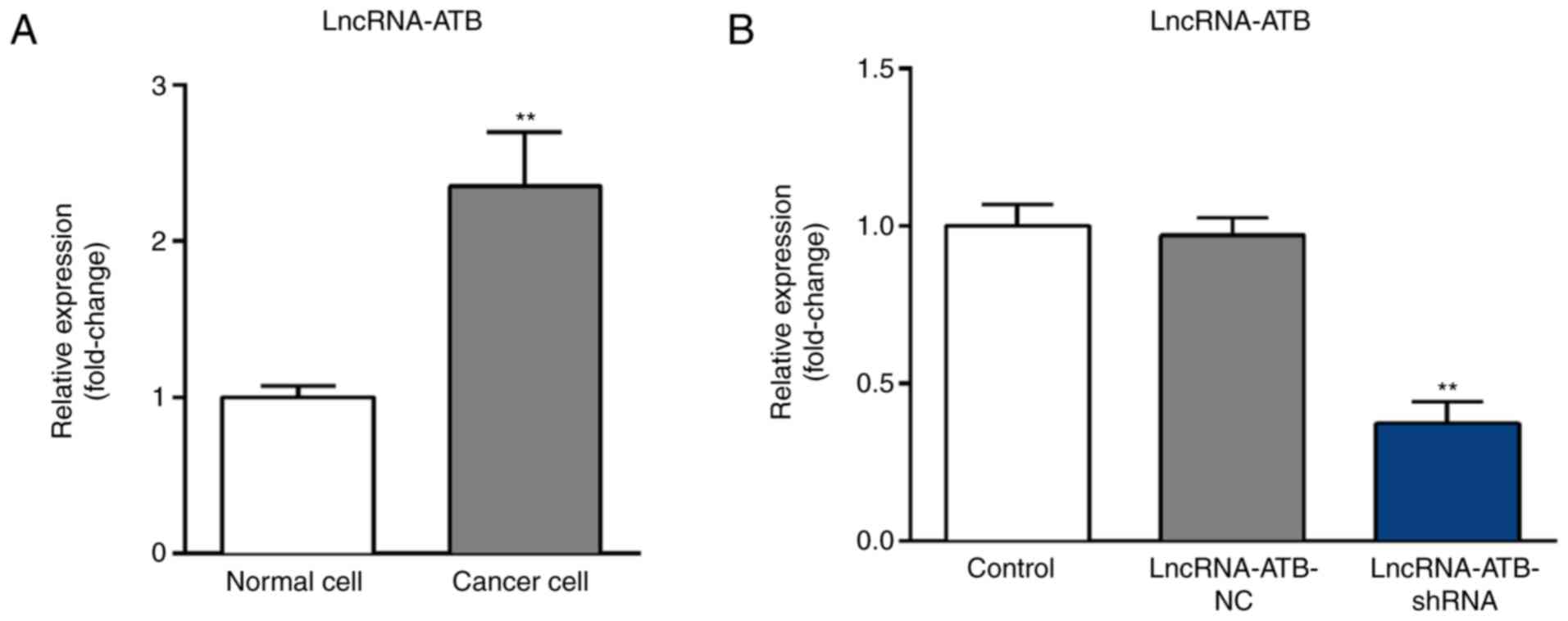
LncRNA-ATB is highly expressed in
ovarian cancer cell line SKOV3
Expression levels of lncRNA-ATB in SKOV3 and HOSEpiC
cells were measured using RT-qPCR. The results indicated that when
compared with HOSEpiC cells, lncRNA-ATB was significantly
upregulated in SKOV3 cells (Fig.
1A). To further investigate the role of lncRNA-ATB in ovarian
cancer, lncRNA-ATB-shRNA or lncRNA-ATB-NC were transfected into
SKOV3 cells. RT-qPCR results revealed that compared with the
control group, lncRNA-ATB-shRNA significantly decreased the
expression of lncRNA-ATB in SKOV3 cells (Fig. 1B).
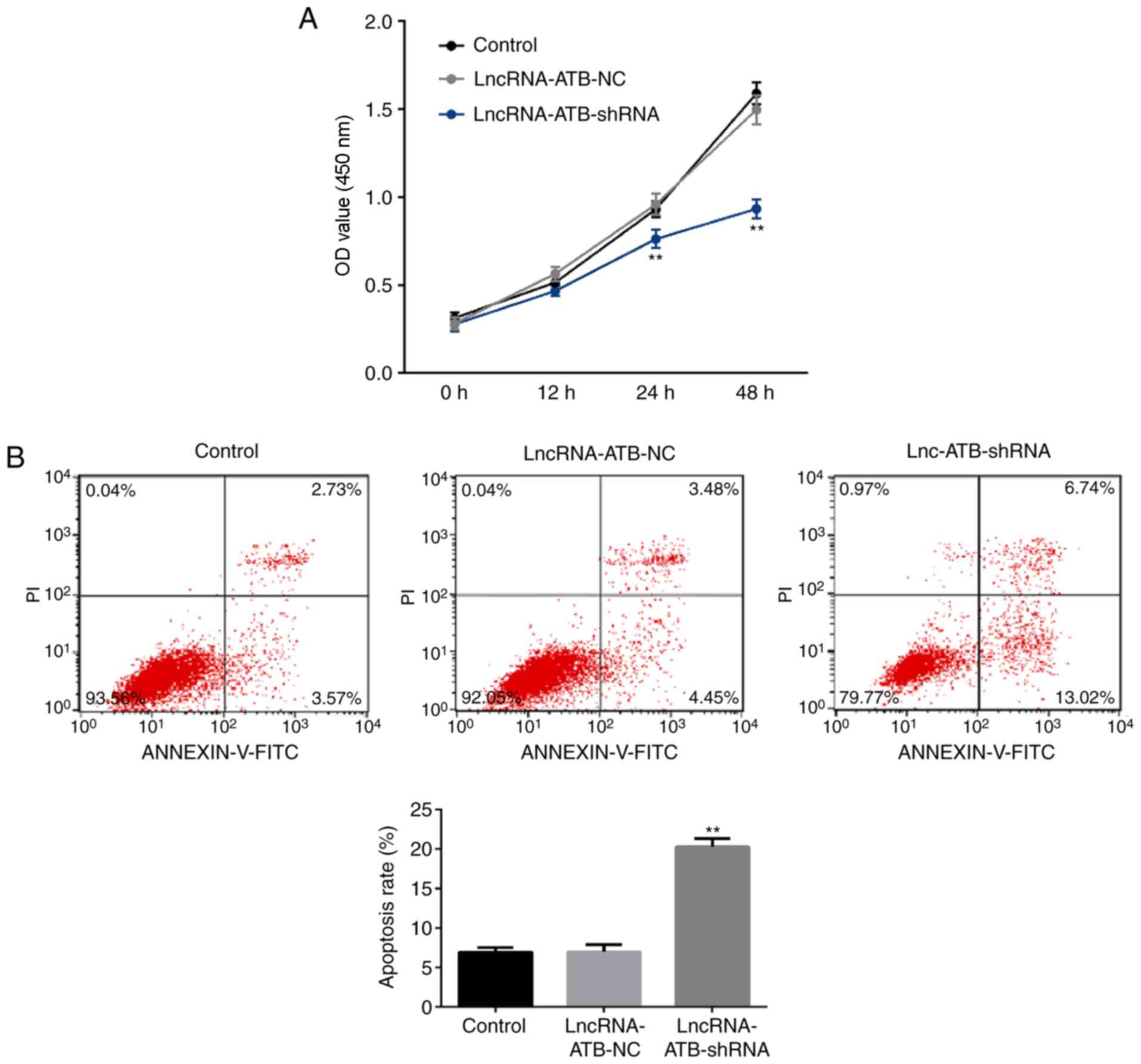
LncRNA-ATB downregulation reduces
proliferation and induces apoptosis in SKOV3 cells
A CCK-8 assay was used to analyze cell proliferation
and flow cytometry was used to analyze apoptosis. The results
revealed that when compared with the control group,
lncRNA-ATB-shRNA significantly reduced the proliferative ability of
SKOV3 cells at the 24 and 48 h time points (Fig. 2A) and promoted apoptosis (Fig. 2B).
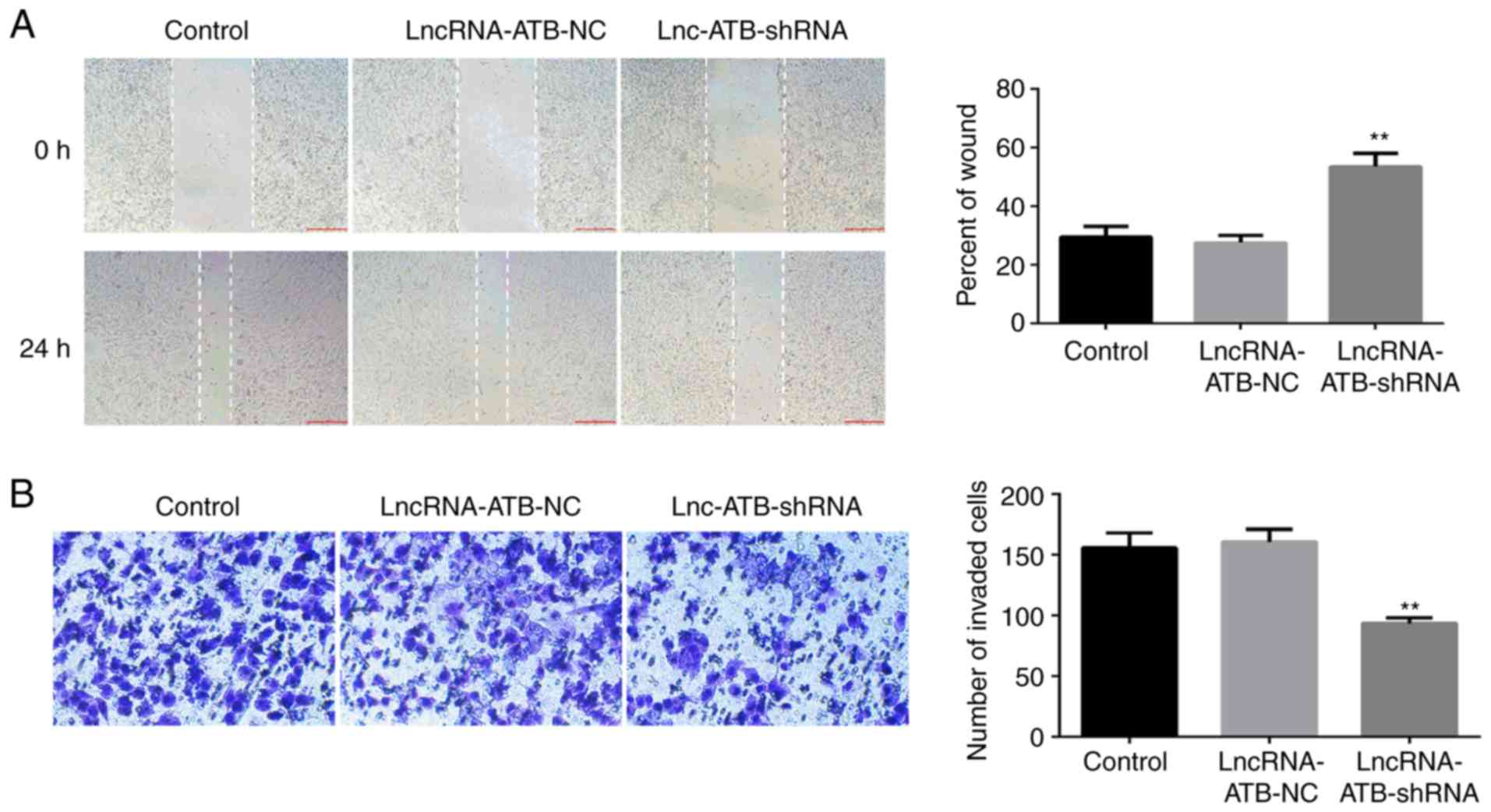
LncRNA-ATB downregulation reduces
migration and invasion of SKOV3 cells
To study the effect of lncRNA-ATB downregulation on
ovarian cancer cell migration and invasion, scratch wound healing
and transwell assays were performed. The results revealed that when
compared with the control group, lncRNA-ATB-shRNA significantly
reduced the migration (Fig. 3A) and
invasion ability (Fig. 3B) of SKOV3
cells.
LncRNA-ATB downregulation
significantly reduces the phosphorylation of STAT3, increases
E-cadherin expression and decreases vimentin expression in SKOV3
cells
The effect of lncRNA-ATB downregulation on ovarian
cancer cell epithelial to mesenchymal transition (EMT) was examined
through analysis of the expression of EMT-related proteins
(E-cadherin and vimentin) in SKOV3 cells. Transfection with
lncRNA-ATB-shRNA significantly increased E-cadherin protein and
mRNA expression (Fig 4A, B and D),
and decreased vimentin protein and mRNA expression (Fig. 4A, B and E).
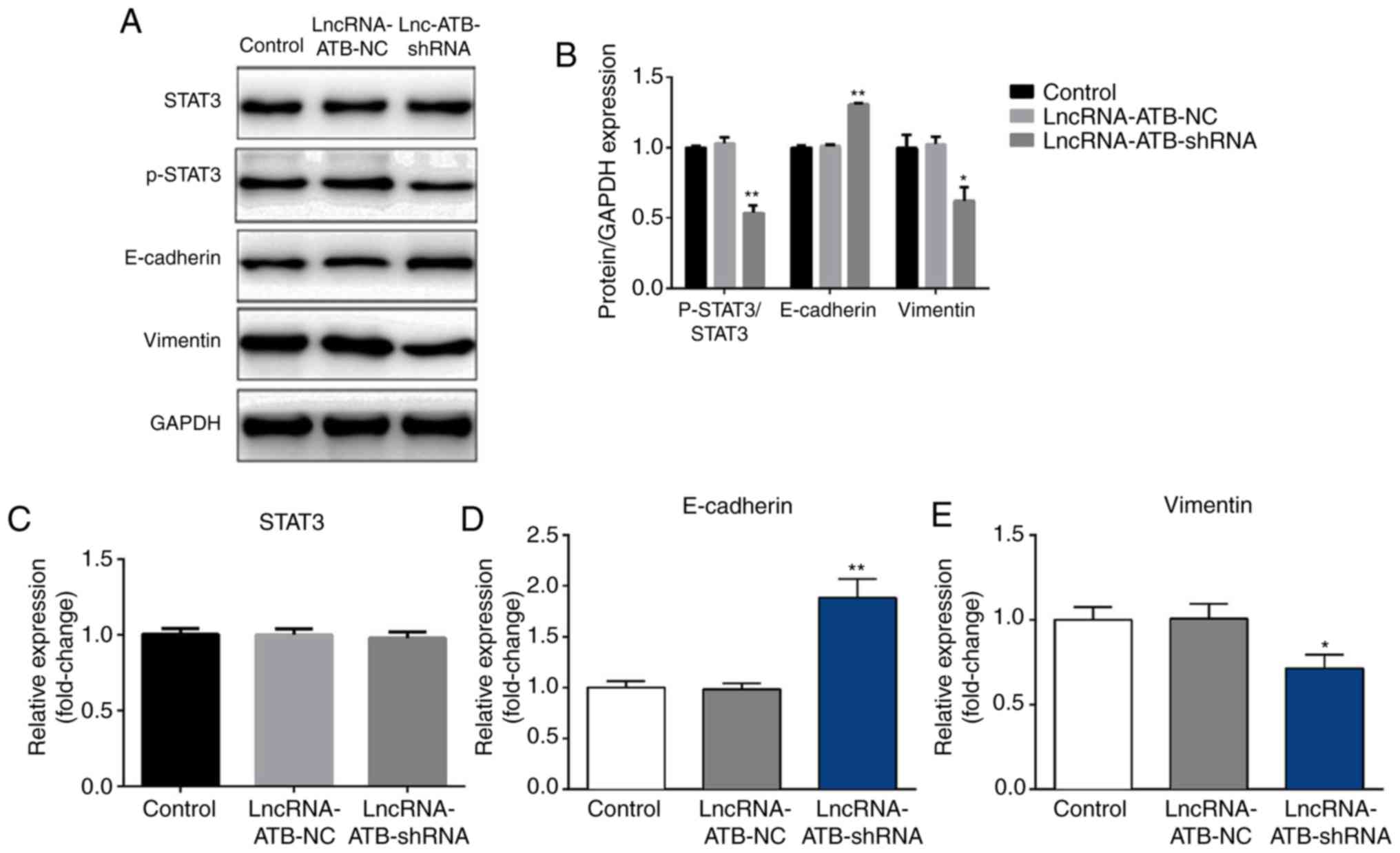 | Figure 4.Effect of lncRNA-ATB downregulation
on the expression of epithelial to mesenchymal transition-related
proteins and the STAT3 pathway activity in SKOV3 cells. (A)
LncRNA-ATB-NC or lncRNA-ATB-shRNA was transfected into SKOV3 cells
for 48 h, then western blotting was used to detect the protein
level of p-STAT3, STAT3, E-cadherin and vimentin in SKOV3 cells.
(B) Western blotting results were quantified. RT-qPCR was used to
detect the mRNA expression of (C) STAT3, (D) E-cadherin and (E)
vimentin in SKOV3 cells. Data are presented as the mean ± SD. Each
experiment was repeated three times. *P<0.05 and **P<0.01 vs.
control group. E-cadherin, epithelial cadherin; lncRNA-ATB, long
non-coding RNA-activated by transforming growth factor-β;
lncRNA-ATB-NC, long non-coding RNA-activated by transforming growth
factor-β-negative control; SKOV3, ovarian cancer cell line; STAT-3,
signal transducer and activator of transcription 3; p-STAT-3,
phosphorylated signal transducer and activator of transcription
3. |
Finally, to explore the molecular mechanisms behind
the role of lncRNA-ATB in ovarian cancer, the STAT3 pathway was
studied via analysis of the phosphorylation of STAT3 using western
blotting. The results suggested that lncRNA-ATB-shRNA significantly
inhibited the phosphorylation of STAT3 (Fig. 4A and B), but had no significant
effect on STAT3 expression (Fig. 4A and
C).
Discussion
Ovarian cancer, the most lethal of all gynecological
malignancies, ranks as the fifth leading cause of cancer deaths in
women (1,2). The prognosis of ovarian cancer is
generally poor due to the lack of clear symptoms and effective
screening and diagnostic methods, especially for identifying early
stages of the disease. For this reason the identification of better
tumor bio-markers to predict the prognosis of ovarian cancer has
attracted the attention of oncologists (20–25).
Increasing evidence suggests that lncRNAs play vital roles in
tumorigenesis and the invasion and metastasis of cancer (26–28).
Several studies have evaluated the prognostic value of lncRNA-ATB
expression in human cancer (29,30).
These previous studies (26–30) indicated that lncRNA-ATB may serve
important roles in ovarian cancer. In the present study, lncRNA-ATB
was significantly upregulated in ovarian cancer cells compared with
healthy ovarian cells, suggesting the possible involvement of
lncRNA-ATB in ovarian cancer development. Furthermore, lncRNA-ATB
downregulation effectively reduced the proliferative, invasive and
migratory capabilities of ovarian cancer cells and significantly
increased apoptosis. These data indicate that lncRNA-ATB may
promote cell proliferation, invasion and migration, and inhibit
cell apoptosis in ovarian cancer cells.
The discovery of EMT, as a mechanism that allows
cancer cells to dedifferentiate and acquire enhanced migratory and
invasive properties, has had a significant impact on cancer
research (31,32). EMT describes the process whereby
epithelial cells undergo multiple biological changes and transform
into interstitial tissues with strongly invasive characteristics
(33,34). Current research indicates that there
are three types of EMT, EMT associated with embryo implantation,
EMT associated with tissue fibrosis and remodeling characteristics,
and EMT associated with strong carcinogenic effects (35). Tumor cells contribute to the
formation of fibrosis through the EMT process and affect the
secretion pattern of cytokines to promote tumorigenesis (36). Moreover, in EMT, initially inactive
polarized epithelial cells dissolve their own intercellular
junctions to form independent, depolarized and active metastatic
mesenchymal cells (37,38). For example, the expression and
function of E-cadherin linked to epithelial cells can be lost,
while N-cadherin, which induces adhesion to mesenchymal cells is
induced (39). EMT is an important
process for cancer cell migration and metastasis and can be induced
by various transcription factors and signal transduction factors
(37–39). In the current study, the effect of
lncRNA-ATB downregulation on ovarian cancer cell EMT was examined
through assessment of the expression of EMT-related proteins
(E-cadherin and vimentin) in SKOV3 cells. This study showed that
lncRNA-ATB downregulation may inhibit ovarian cancer cell EMT.
STAT3 enhances various signaling pathways involved
in the regulation of cell growth, invasion and EMT, and is
recognized as a key target for cancer treatment (40–42). The
STAT3 signaling pathway plays an important role in ovarian cancer
progression (43–45). In order to explore the molecular
mechanism behind the role of lncRNA-ATB in ovarian cancer, the
STAT3 pathway was analyzed. lncRNA-ATBA downregulation
significantly reduced STAT3 activation, indicating that lncRNA-ATB
may act on ovarian cancer cells at least partly through influencing
the STAT3 signaling pathway activation.
In conclusion, this study demonstrated that
lncRNA-ATB was highly expressed in ovarian cancer cells, and that
its downregulation effectively inhibited ovarian cancer cell
proliferation, invasion, and migration and induced cell apoptosis.
These effects might be mediated by the regulation of the STAT3
signaling pathway. Therefore, lncRNA-ATB may be a new therapeutic
target for the treatment of ovarian cancer. However, this study
only includes a preliminary analysis. To clarify the role of
lncRNA-ATB in ovarian cancer, further research is needed. The role
of lncRNA-ATB in other ovarian cancer cell lines and in ovarian
cancer in vivo should be studied. These issues will be
addressed in future studies.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data sets used and/or generated during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
DY and XZ contributed to study design, data
collection, statistical analysis, data interpretation and
manuscript preparation. YZ, HQ, HW, CH, XL, TG and ML contributed
to data collection and statistical analysis. HY and JY contributed
to study design, data interpretation and manuscript
preparation.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Webb PM and Jordan SJ: Epidemiology of
epithelial ovarian cancer. Best Pract Res Clin Obstet Gynaecol.
41:3–14. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reid BM, Permuth JB and Sellers TA:
Epidemiology of ovarian cancer: A review. Cancer Biol Med. 14:9–32.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pierredon S, Ribaux P, Tille JC, Petignat
P and Cohen M: Comparative secretome of ovarian serous carcinoma:
Gelsolin in the spotlight. Oncol Lett. 13:4965–4973. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Howlader N, Noone A, Krapcho M, Neyman N,
Aminou R, Waldron W, Altekruse SF, Kosary CL, Ruhl J, Tatalovich Z,
et al: SEER Cancer Statistics Review, 1975–2008, National Cancer
Institute.
|
|
5
|
Grunewald T and Ledermann JA: Targeted
Therapies for Ovarian Cancer. Best Pract Res Clin Obstet Gynaecol.
41:139–152. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim JY, Cho CH and Song HS: Targeted
therapy of ovarian cancer including immune check point inhibitor.
Korean J Intern Med. 32:798–804. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brasseur K, Gévry N and Asselin E:
Chemoresistance and targeted therapies in ovarian and endometrial
cancers. Oncotarget. 8:4008–4042. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lintner NG and Cate J: Regulating the
ribosome: A spotlight on RNA dark matter. Mol Cell. 54:1–2. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ponting CP, Oliver PL and Rei W: Evolution
and functions of long noncoding RNAs. Cell. 136:629–641. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cesana M, Cacchiarelli D, Legnini I,
Santini T, Sthandier O, Chinappi M, Tramontano A and Bozzoni I: A
long noncoding RNA controls muscle differentiation by functioning
as a competing endogenous RNA. Cell. 147:358–369. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kitagawa M, Kitagawa K, Kotake Y, Niida H
and Ohhata T: Cell cycle regulation by long non-coding RNAs. Cell
Mol Life Sci. 70:4785–4794. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang XM, Yang GL, Le XH, et al:
Expression profiles of long non-coding RNA in HCC patients
suffering hepatitis B liver cirrhosis. J Zhengzhou Univ (Med Sci).
50:194–198. 2015.
|
|
13
|
Yu T, Zhao Y, Hu Z, Li J, Chu D, Zhang J,
Li Z, Chen B, Zhang X, Pan H, et al: MetaLnc9 facilitates lung
cancer metastasis via a PGK1-activated AKT/mTOR pathway. Cancer
Res. 77:5782–5794. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Su W, Xu M, Chen X, Chen N, Gong J, Nie L,
Li L, Li X, Zhang M and Zhou Q: Long noncoding RNA ZEB1-AS1
epigenetically regulates the expressions of ZEB1 and downstream
molecules in prostate cancer. Mol Cancer. 16:1422017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dey BK, Mueller AC and Dutta A: Long
non-coding RNAs as emerging regulators of differentiation,
development, and disease. Transcription. 5:e9440142014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Iguchi T, Uchi R, Nambara S, Saito T,
Komatsu H, Hirata H, Ueda M, Sakimura S, Takano Y, Kurashige J, et
al: A long noncoding RNA, lncRNA-ATB, is involved in the
progression and prognosis of colorectal cancer. Anticancer Res.
35:1385–1388. 2015.PubMed/NCBI
|
|
17
|
Fu N, Zhao SX, Kong LB, Du JH, Ren WG, Han
F, Zhang QS, Li WC, Cui P, Wang RQ, et al:
LncRNA-ATB/microRNA-200a/β-catenin regulatory axis involved in the
progression of HCV-related hepatic fibrosis. Gene. 618:1–7. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ke L, Xu SB, Wang J, Jiang XL and Xu MQ:
High expression of long non-coding RNA ATB indicates a poor
prognosis and regulates cell proliferation and metastasis in
non-small cell lung cancer. Clin Transl Oncol. 19:599–605. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen S, Zhang L, Yan G, Cheng S, Fathy AH,
Yan N and Zhao Y: Neutrophil-to-lymphocyte ratio is a potential
prognostic biomarker in patients with ovarian cancer: A
Meta-analysis. Biomed Res Int. 2017:79434672014.
|
|
21
|
Wang J, Zhao Y, Qi R, Zhu X, Huang C,
Cheng S, Wang S and Qi X: Prognostic role of podocalyxin-like
protein expression in various cancers: A systematic review and
meta-analysis. Oncotarget. 8:52457–52464. 2017.PubMed/NCBI
|
|
22
|
Zhou Y, Cheng S, Chen S and Zhao Y:
Prognostic and clinicopathological value of SIRT3 expression in
various cancers: A systematic review and meta-analysis. OncoTargets
Ther. 11:2157–2167. 2018. View Article : Google Scholar
|
|
23
|
Zhou Y, Cheng S, Fathy AH, Qian H and Zhao
Y: Prognostic value of platelet-tolymphocyte ratio in pancreatic
cancer: A comprehensive meta-analysis of 17 cohort studies.
OncoTargets Ther. 11:1899–1908. 2018. View Article : Google Scholar
|
|
24
|
Zhao Y, Si G, Zhu F, Hui J, Cai S, Huang
C, Cheng S, Fathy AH, Xiang Y and Li J: Prognostic role of platelet
to lymphocyte ratio in hepatocellular carcinoma: A systematic
review and meta-analysis. Oncotarget. 8:22854–22862. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Song X, Yao H, Liu J and Wang Q: The
prognostic value of long noncoding RNA Sox2ot expression in various
cancers: A systematic review and meta-analysis. Clin Chim Acta.
484:52–59. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hu Y, Chen HY, Yu CY, Xu J, Wang JL, Qian
J, Zhang X and Fang JY: A long non-coding RNA signature to improve
prognosis prediction of colorectal cancer. Oncotarget. 5:2230–2242.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Beckedorff FC, Amaral MS,
Deocesano-Pereira C and Verjovski-Almeida S: Long non-coding RNAs
and their implications in cancer epigenetics. Biosci Rep. 33(pii):
e000612013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gutschner T and Diederichs S: The
hallmarks of cancer: A long non-coding RNA point of view. RNA Biol.
9:703–719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen Y, Wei G, Xia H, Tang Q and Bi F:
Long noncoding RNAATB promotes cell proliferation, migration and
invasion in gastric cancer. Mol Med Rep. 17:1940–1946.
2018.PubMed/NCBI
|
|
30
|
Fu XM, Guo W, Li N, Liu HZ, Liu J, Qiu SQ,
Zhang Q, Wang LC, Li F and Li CL: The expression and function of
long noncoding RNA lncRNA-ATB in papillary thyroid cancer. Eur Rev
Med Pharmacol Sci. 21:3239–3246. 2017.PubMed/NCBI
|
|
31
|
Karlsson MC, Gonzalez SF, Welin J and Fuxe
J: Epithelial-mesenchymal transition in cancer metastasis through
the lymphatic system. Mol Oncol. 11:781–791. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liao TT and Yang MH: Revisiting
epithelial-mesenchymal transition in cancer metastasis: The
connection between epithelial plasticity and stemness. Mol Oncol.
11:792–804. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Davis FM, Azimi I, Faville RA, Peters AA,
Jalink K, Putney JW Jr, Goodhill GJ, Thompson EW, Roberts-Thomson
SJ, Monteith GR, et al: Induction of epithelial-mesenchymal
transition (EMT) in breast cancer cells is calcium signal
dependent. Oncogene. 33:2307–2316. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yoshida T, Ozawa Y, Kimura T, Sato Y,
Kuznetsov G, Xu S, Uesugi M, Agoulnik S, Taylor N, Funahashi Y and
Matsui J: Eribulin mesilate suppresses experimental metastasis of
breast cancer cells by reversing phenotype from
epithelial-mesenchymal transition (EMT) to mesenchymal-epithelial
transition (MET) states. Br J Cancer. 110:1497–1505. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qu C, Zhang W, Zheng G, Zhang Z, Yin J and
He Z: Metformin reverses multidrug resistance and
epithelial-mesenchymal transition (EMT) via activating
AMP-activated protein kinase (AMPK) in human breast cancer cells.
Mol Cell Biochem. 386:63–71. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vetuschi A, Pompili S, Flati V, Berardinis
LD, Di Gregorio J, Latella G and Sferra R:
Epithelial-to-Mesenchimal Transition (EMT) in experimental
intestinal fibrosis. Italian J Anatomy Embryology. 120:582015.
|
|
37
|
Yang Y, Gao M, Lin Z, Chen L, Jin Y, Zhu
G, Wang Y and Jin T: DEK promoted EMT and angiogenesis through
regulating PI3K/AKT/mTOR pathway in triple-negative breast cancer.
Oncotarget. 8:98708–98722. 2017.PubMed/NCBI
|
|
38
|
Ma Z, Xin Z, Hu W, Jiang S, Yang Z, Yan X,
Li X, Yang Y and Chen F: Forkhead box O proteins: Crucial
regulators of cancer EMT. Semin Cancer Biol. 50:21–31. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zuo L, Zhao H, Yang R, Wang L, Ma H, Xu X,
Zhou P and Kong L: Lamin A/C might be involved in the EMT
signalling pathway. Gene. 663:51–64. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhao J, Du P, Cui P, Qin Y, Hu C, Wu J,
Zhou Z, Zhang W, Qin L and Huang G: LncRNA PVT1 promotes
angiogenesis via activating the STAT3/VEGFA axis in gastric cancer.
Oncogene. 37:4094–4109. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Egusquiaguirre SP, Yeh JE, Walker SR, Liu
S and Frank DA: The STAT3 Target Gene TNFRSF1A Modulates the NF-κB
Pathway in Breast Cancer Cells. Neoplasia. 20:489–498. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bournazou E and Bromberg J: Targeting the
tumor microenvironment: JAK-STAT3 signaling. JAKSTAT.
2:e238282013.PubMed/NCBI
|
|
43
|
Chen L, Wang J, Wu J, Zheng Q and Hu J:
Indirubin suppresses ovarian cancer cell viabilities through the
STAT3 signaling pathway. Drug Des Devel Ther. 12:3335–3342. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jia ZH, Jia Y, Guo FJ, Chen J, Zhang XW
and Cui MH: Phosphorylation of STAT3 at Tyr705 regulates MMP-9
production in epithelial ovarian cancer. PLoS One. 12:e01836222017.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Saini U, Naidu S, ElNaggar AC, Bid HK,
Wallbillich JJ, Bixel K, Bolyard C, Suarez AA, Kaur B, Kuppusamy P,
et al: Elevated STAT3 expression in ovarian cancer ascites promotes
invasion and metastasis: A potential therapeutic target. Oncogene.
36:168–181. 2017. View Article : Google Scholar : PubMed/NCBI
|