Introduction
Osteoarthritis (OA) is a joint disease characterized
by the degeneration of articular cartilage and modifications in
subchondral bone. Importantly, OA has a complex pathophysiology and
its presentation is associated with pathologies of manifold joint
tissues. Primary OA is usually a result of the interaction of
genetic and epigenetic factors that remain to be fully defined
(1,2). However, joint inflammation, obesity,
hormonal imbalance and a low calcium concentration are strongly
associated with secondary OA (3,4).
Importantly, OA and osteoporosis (OP) are two skeletal pathologies
which are closely associated and have as a distinct feature the
abnormal reconstruction of subchondral bone (5).
OP is one of the most common diseases affecting
elderly individuals worldwide (6).
It is characterized by reduced bone mineral density (BMD) and the
microarchitectural deterioration of bone tissue. Based on its
etiology, OP is categorized into two distinct types, namely type I
(postmenopausal) and type II (senile) (7,8). In type
I OP, the pathology generally develops with the estrogen reduction
following the onset of menopause which causes bone loss. During the
progression of type I disease, pathological changes of the
trabecular bone are the most common. On the other hand, type II OP,
which usually occurs after the age of 70, involves the thinning of
both trabecular and cortical bone (7,8). Factors
associated with the presentation of OP include the absence of
physical exercise, malnutrition, poor protein synthesis and the
lack of vitamin C, as well as low menopausal and postmenopausal
estrogen secretion (6). Currently,
various pharmacological options are available for the treatment of
OP; however, in OA, the management of patients is mostly limited to
pain reduction and diverse modifications of lifestyle (9). Patients with advanced OA may receive
non-steroidal anti-inflammatory drugs (NSAIDs), undergo physical
therapy or occupational therapy, as well as surgical procedures,
including cortisone and lubrication injections. At the final stages
of the disease, total joint replacement or osteotomy are common and
are usually the only treatment options (10).
Aging in OP and OA
Even though OP and OA are clinically distinct
pathologies, they have many similarities as regards the hallmarks
of aging. Thus, both pathologies have been characterized as
age-related diseases, determining the lifestyle of affected
patients more challenging as compared to a healthy elderly
population (6). In particular, OA
decreases the mobility, productivity and quality of life of
individuals and leads to an increase in morbidity, as well as the
use of medication and social welfare expenditures that contribute
to a substantial socioeconomic burden (8).
The key feature of OA is the presentation of
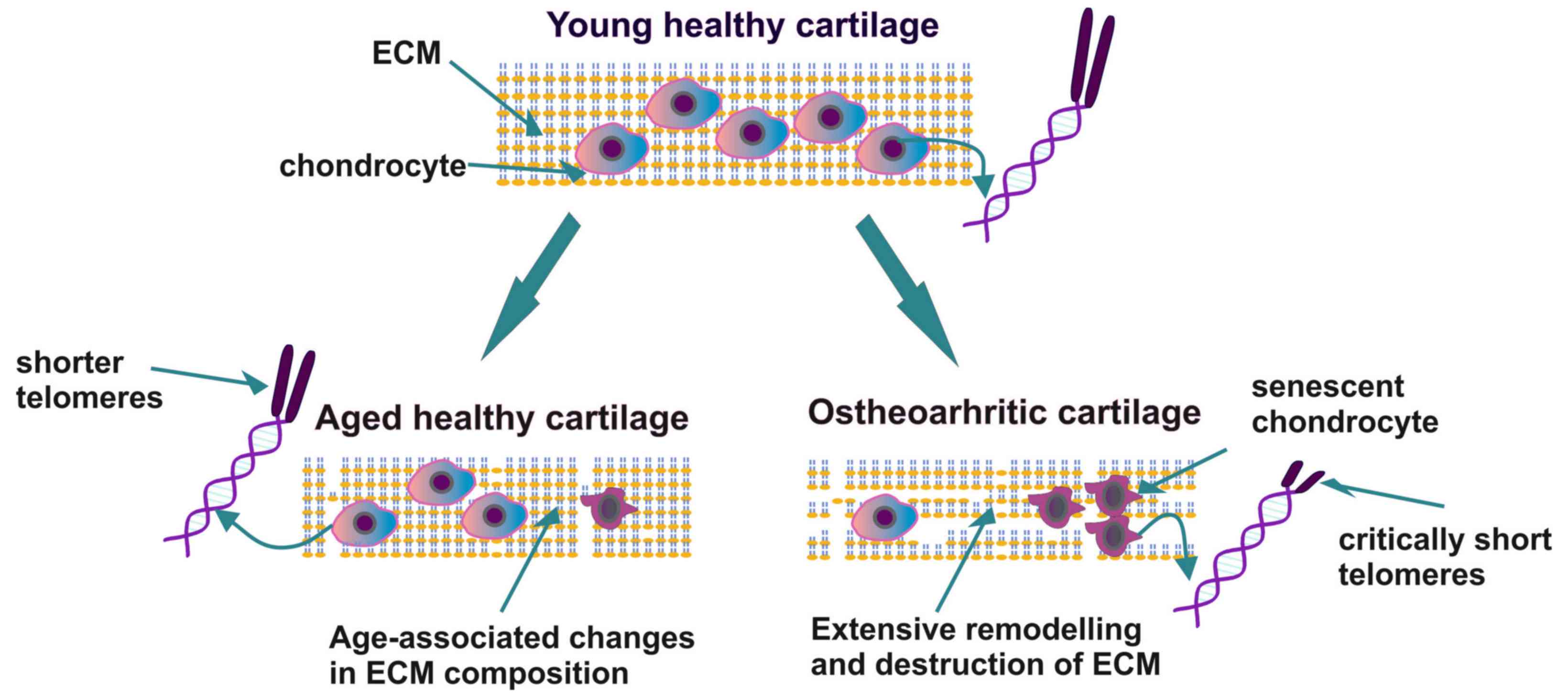
senescent cartilage with different histopathological
characteristics compared to the aging cartilage tissue that is
normally present in a healthy elderly individual. In aging
cartilage, chondrocytes exhibit a decreased number and a low
ability to proliferate. Simultaneously, the synthesis and
deposition of extracellular matrix (ECM) components, that plays an
important role in bone homeostasis and pathophysiology (11), is diminished, resulting in the
gradual thinning of the cartilage layers (12). Indeed, it has been demonstrated that
alterations in the function of chondrocytes and the deposition of
ECM components may weaken the structural properties of articular
cartilage and render the joint susceptible to OA. Furthermore, the
tissues of patients with OA exhibit clusters of chondrocytes at the
site of the lesion with an altered metabolism and an increased
ability to produce pro-inflammatory cytokines and matrix-degrading
enzymes (12). Thus, the
pathogenesis of OA is linked to aging through several mechanisms,
including inflammation related to aging, obesity, senescence,
oxidative stress, alterations in metabolism, as well as cell
signaling due to epigenetic mechanisms. Importantly, chondrocyte
senescence contributes to the decreased ability of chondrocytes to
repair articular cartilage tissue (12–14).
OP is a chronic skeletal disease with a high
frequency worldwide, which has as a main characteristic, the
deteriorated bone microarchitecture associated with the
co-presentation of low BMD. However, the pathology is asymptomatic,
and is associated in the majority of cases with a high risk of bone
fracture, resulting in significant morbidity and mortality. OP
related to aging is the most common form of the disease. There are
several pathways involved in the etiopathogenesis of OP, including
metabolic, endocrine and mechanical factors, whereas chronic
inflammation has also been shown to play an important role
(15). Indeed, the well-established
estrogen deficiency in postmenopausal women has been shown to
enhance the release of inflammatory mediators, leading to
postmenopausal OP. Moreover, the ability of estrogens to
downregulate receptor activator of nuclear factor-κΒ ligand (RANKL)
synthesis has been shown in osteoblasts and likewise in T- and
B-cells (16). Indeed, it is the
lack of estrogen that, through the enhancement in T-cell
activities, induces the increased secretion of pro-osteoclastogenic
cytokines and subsequent osteoclastogenesis (17). Concomitantly, immune cells have been
recognized as factors contributing to the development of
osteoporosis (18). Moreover, it has
been suggested that chronic antigenic load and oxidative stress
that accumulate with aging cause a low-grade inflammation
associated with a decrease in bone formation, as well as in bone
resorption. These processes together lead to an imbalance in bone
remodeling and the increased prevalence of OP (18).
Oxidative stress is intimately associated with the
mechanisms of aging and together with other aging-related factors,
including inflammation, an altered metabolism, cellular senescence
and mitochondrial dysfunction, induce gradual OA-dependent joint
destruction and increased bone fragility in OP (19). Reactive oxygen species (ROS) are
generated under physiological conditions during mitochondria
respiratory chain activities or through the actions of oxidative
enzymes. However, upon biological, mechanical or chemical
stimulation, an imbalance between ROS formation and ROS elimination
occurs, favoring oxidative damage. This imbalance leads to an
uncontrolled ROS production that favors pro-oxidant processes and
oxidative damage (20,21). Importantly, previous studies have
demonstrated that oxygen free radicals can directly cause DNA
damage (22), particularly at the
guanine-rich end parts of chromosomes termed telomeres, the
association of which with OA, OP and aging is discussed below
(23).
Role of telomeres and telomerase in
aging
Telomeres are specific structures positioned at the
end of chromosomes that together with specific protein complexes
bound to them, provide DNA protection (24), ensuring genomic stability.
Specifically, the repetitive 5′-TTAGGG-3′ sequences of 70 to 100
nucleotides bound to telomeric interacting proteins provide a
protective cap of the chromosomal DNA, resembling the end of a
shoelace. Due to the inability of the enzyme DNA polymerase to
preserve the length of the 3′ overhang (the so-called ‘end
replication problem’), telomeres are deprived of a small number of
nucleotides during each mitotic cycle, practically becoming shorter
and shorter (25). Indeed, the
‘telomere hypothesis of cellular aging’ was postulated in 1992 by
Harley et al (26), and the
over the past few years, accumulating evidence has indicated that
telomere length (TL), which can be affected by various lifestyle
factors, is associated with aging and the onset of age-related
diseases (26,27). In particular, stem cell dysfunction
caused by telomere shortening may be one of the mechanisms
responsible for aging in both humans and mice (9). Of note, a recent study revealed that
the administration of nutraceutical supplements to healthy
individuals was implicated in TL maintenance (28).
Telomeres are replicated by a specialized
ribonucleoprotein complex, known as telomerase, that consists of a
protein component entitled telomerase reverse transcriptase (TERT)
that serves as a catalytic subunit (29) and an essential telomerase RNA
component (TERC or RT) (30).
Importantly, the enzyme telomerase can reverse telomere shortening,
as it contributes to sustaining TL. However, it exhibits a high
activity only in a subgroup of highly proliferating adult somatic
cells, e.g., stem and progenitor cells, activated lymphocytes, as
well as germline cells (31), while
in the majority of adult human somatic cells, the expression and
activity of telomerase is undetectable.
Recently, however, Astragalus membranaceus
root and its active component, cycloastragenol, has been shown to
activate telomerase in human somatic cells, in vitro and
in vivo (32). In agreement
with this, de Jesus et al previously demonstrated that the
TA-65 component from Astragalus membranaceus root induced
the elongation of short telomeres and the neutralization of
associated DNA damage, in a telomerase-dependent manner (33). In addition to these studies, it has
recently been demonstrated that specific natural compounds can
significantly activate telomerase in human peripheral blood
mononuclear cells in vitro (28).
TL and telomerase activity are strongly associated
with human health as they have been linked to several age-related
diseases, such as cancer, cardiovascular disease (CAD), diabetes,
rheumatoid arthritis and psychiatric disorders (34–38).
Moreover, previous studies have suggested that female human
fertility decreases with an increased maternal age and that various
adverse factors, including reduced telomerase activity, can
contribute to age-associated infertility in women (39,40).
Furthermore, Vakonaki et al recently demonstrated the
existence of a link between TL and drug abuse, which ultimately
results in premature biological aging (41).
Telomere shortening, which is the main cause of
age-related diseases, can be perpetrated through two distinctive
mechanisms (42). According to the
first mechanism, telomeres physiologically shorten with each cell
division, due to the end replication problem. Since cells have a
pre-defined number of cell divisions, when they reach the stage of
critically short telomeres, they become senescent (43). As regards the second mechanism,
imbalanced ROS production and associated oxidative stress can cause
DNA damage to the guanine residues of telomeres, inducing the
erosion of single telomeres (44).
In general, DNA damage at the site of telomeres caused by various
environmental factors triggers a DNA-damage response that protects
them from instability and shortening (44,45).
However, if this protective mechanism is dysfunctional, telomeres
are exposed to several damaging agents, leading to their critical
shortening.
From all the above, it is clear that the
determination of TL and its maintenance through intervention, can
highly contribute to the delay of the aging process and the
treatment of several age-related diseases, leading to longevity. In
that context, a distinction between short telomeres and critically
short telomeres has been achieved, considering the pivotal role
that the critically short telomeres play in cell homeostasis. Based
on these findings and developments, the ‘BIOTEL’ database was
recently created, that is able to calculate a wide range of TL
statistics, biological age and applications telomere biology
research (46). The utilization of
BIOTEL and similar tools will facilitate the analysis and
assessment of telomere biology data and their application in health
care.
The current review focuses on the role of telomeres
in OA and OP pathologies and discusses the usability of TL and the
rate of telomere shortening as potential disease biomarkers.
Association between telomere length, OP and
OA
In an early, milestone study, Oreffo et al
demonstrated that the number of osteoblast progenitors in the bone
marrow (BM) of patients with OP, compared to that of age-matched
controls, was decreased (47). In
women, this has been partly attributed to age-related alterations
in hormone levels, regarding sex hormones. In the context of
elucidating the association between the aging of BM-mesenchymal
cells and the development of OP, telomere shortening has gained
increasing attention. Previous studies have demonstrated that the
proliferative and osteogenic capacity of cultured mesenchymal
stromal cells (MSCs) isolated from patients with OP was
significantly decreased, which can be a possible marker of
premature aging (48). It has been
suggested that the decreased proliferative ability of MSCs may be
due to the overexpression of osteogenic inhibitors in these cells
in the case of OP (49,50). More specifically, differences have
been observed between the mesenchymal cell transcriptomes in OP and
non-OP aging populations, which may be due to epigenetic changes
reflecting a specific OP-associated aging process (49,50).
These findings are further supported by in vivo mouse
models, which demonstrated that the proliferation-independent
dysfunction of telomeres can induce an attenuation of osteoblast
differentiation in mice with accelerated aging (51).
However, even though various studies had examined TL
and its association with the pathology of OP, there is a great deal
of inconsistency among them. For instance, in a large cohort of
unselected women, the blood leukocyte TL was shown to be associated
with BMD, whereas clinical OP was associated with shorter telomeres
(52). Another study estimated the
TL of female patients with OP using the telomere restriction
fragment (TRF) approach, which revealed a stable decrease in TL
among the different age groups of patients. Moreover, telomere
shortening in leukocytes was associated with BMD or bone loss, but
only after correcting for age, where TL was found to be associated
with longitudinal bone loss, regarding sites in the distal forearm
region (53). On the other hand, an
early study comparing TRFs from peripheral blood length (PBL) DNA
from female patients with OP and age-matched controls did not find
any significant alterations (54).
Likewise, in a separate study, even though age was found to be
associated with both TL and BMD, no significant association was
observed between TL and BMD. These inconsistencies among different
studies could be explained by the results of a recent study that
was performed in a cohort of elderly Chinese female and male
patients with OP. According to that study, in the case of the
female patients, age affected the association of TL with BMD and
OP, but not in the case of the male patients, strongly suggesting
that the TL predictive role may be sex-specific (55).
In addition to the above, accumulating evidence
demonstrates that HIV is a significant risk factor for a low BMD
and fractures due to bone fragility (56). Recently, it was determined that in a
cohort of women with HIV, there was an association between
premature spinal bone loss and a shorter TL. In summary, further
focused studies are required to evaluate the association of TL and
OP, as well as the feasibility of utilizing TL as an OP prognostic
index.
As regards OA and its association with TL, patients
with OA also appear to acquire several abnormalities indicative of
premature aging. Indeed, OA-affected chondrocytes tend to obtain a
senescence-like phenotype (57), a
fact that has initiated several attempts to evaluate the putative
association of TL with the progression of OA. In that context, in a
recent study, leukocyte relative telomere length (RTL) in patients
with knee OA was compared to that of healthy controls (58). Additionally, possible associations
between plasma angiogenic cytokine concentrations and leukocyte RTL
were examined, indicating that TL was shorter in patients with knee
OA compared to age-matched healthy controls. Notably, plasma
hepatocyte growth factor (HGF), vascular endothelial growth factor
(VEGF) and granulocyte-colony stimulating factor (G-CSF) levels
were found to be negatively associated with leukocyte RTL.
Therefore, that study indicated that high circulating angiogenic
cytokine levels in the knees of patients with OA may reflect high
oxidative stress and chronic inflammation, leading to subsequent
telomere shortening (58). Indeed,
overall oxidative stress can expedite telomere shortening either
indirectly through increase in cell division, or directly acting on
DNA telomere repeats, suggesting that redox balance is a prominent
factor that regulates chondrocytes lifespan (59). In 2001, Martin and Buckwalter
suggested that age-related changes in human cartilage chondrocytes
may lead to cartilage erosion and osteoarthritis (60). Moreover, Price et al,
utilizing Southern blot analysis, had demonstrated that patients
with OA had shorter telomeres compared to unaffected chondrocytes
in a group of 15 patients with hip OA, 30 patients with knee OA and
a control group of 11 patients with no joint diseases (61). Additionally, Zhai et al
measured relative TL in 160 patients with hand OA and in 926
patients without hand OA (62,63).
According to this study, the affected patients had shorter
telomeres compared to the control group, suggesting that oxidative
stress and inflammation within the affected joints led to telomere
shortening due to accelerated DNA replication (61). In a separate study, Tamayo et
al measured the average TL by qPCR in 34 patients with OA, in
35 patients with OP and 130 controls, and did not detect any
differences between the patient and control groups (64). In 2011, these authors had estimated
TL in human chondrocytes and peripheral blood leukocytes in 20
controls and 39 patients with knee and hip OA, and it was found
that chondrocytes from patients with OA exhibited a significantly
shorter TL when compared to chondrocytes from healthy individuals.
Moreover, in patients with OA, telomeres were 1.6-fold longer in
chondrocytes compared to leukocytes, indicating the existence of a
cell-type specificity regarding TL. This hypotheses was
corroborated by data from control subjects, where telomeres in
chondrocytes were found to be even twice as long as telomeres from
leukocytes (65). On the other hand,
no difference was detected between the leukocytes and chondrocytes
of controls, suggesting that the shorter telomeres in leukocytes
may result from higher frequency of divisions of leukocytes
compared to chondrocytes. Further supporting this theory, articular
cartilage is a post-mitotic tissue, indicating that chondrocytes do
not replicate often and in addition, according to a PCR-based assay
decreased peripheral blood TL is markedly associated with the
presentation of hand OA (66,67).
Importantly, the shortening of chondrocyte telomeres
caused by oxidative stress is a common aging-related process
strongly associated with the incidence of cellular senescence, a
finding that may have vast clinical importance in the early
diagnosis and prognosis of OA. Harbo et al estimated the
mean TL, the number of short telomeres (<1,500 bp) and the sites
with senescence relative to the distance of the OA lesion, and
concluded that all examined markers were highly associated with the
distance from the lesion site (68).
Importantly, the short telomere load was found to be a more
significant marker for OA presentation, compared to the mean TL
(68). Even though the respective
study included only 3 patients with OA, it provides evidence that
short telomere load of chondrocytes could be an important marker
for OA diagnosis and prognosis. Based on these results, further
studies focusing on the measurement of short telomere load in a
larger dataset are warranted.
Notably, Sibille et al suggested that
OA-associated pain was a severe stress factor that could affect TL.
They measured TL in 136 women, aged between 45 and 85 years with or
without symptomatic OA and categorized the participants into 5
groups according to the pain severity. This approach revealed that
patients with chronic severe pain had shorter telomeres, although
long TL did not correspond to low pain severity in this cohort
(69).
Therefore, aging, OA and OP are independent
processes; however, age is an important factor contributing to the
progression of OA (4). Importantly,
Ganguly et al among others, suggested that a decline in the
number and ‘fitness’ of MSCs in the BM may be one of the main
factors contributing to bone abnormalities in OP and OA (9) (Table
I).
 | Table I.Association of TL with the
progression of OA and OP pathologies. |
Table I.
Association of TL with the
progression of OA and OP pathologies.
| Authors/(Refs.),
year | Sample
demographics | Main endpoint | TL assay type |
|---|
| Price et al
(61), 2002 | OA hip patients
(n=15) vs. OA knee patients (n=30) vs. healthy (n=11) | Shorter TL in OA
patients | Southern blot
analysis |
| Zhai et al
(62), 2006; Li et al
(63), 2012 | OA hand patients
(n=160) vs. healthy (n=926) | Shorter TL in OA
patients | Southern blot
analysis |
| Valdes et al
(52), 2007 | OP and BMD in
females (n=2,150) aged 18–79 years | Shorter leukocyte
TL is not associated with decreased BMD or OP | TRF |
| Sanders et
al (71), 2009 | Individuals
(n=2,750) aged 70–79 years with OP or fractures | TL is not
associated with BMD, OP, or fractures in older men or women | qPCR |
| Tang et al
(72), 2010 | BMD in hip in
elderly individuals (n=1,876) | TL was not
associated with either baseline BMD or bone loss over a period of 4
years | qPCR |
| Tamayo et al
(64), 2010 | OA (n=34) and OP
(n=35) vs. healthy individuals (n=130) | No differences
observed in OA, but a decreased observed TL in OP | qPCR |
| Tamayo et al
(65), 2011 | OA patients (n=39)
and in healthy (n=20) individuals in leukocytes and in
chondrocytes | OA longer TL in
chondrocytes vs. leukocytes; no differences between TL in
chondrocytes and leukocytes in healthy individuals | qPCR |
| Nielsen et
al (73), 2015 | Samples of the
lumbar spine (LS), femoral neck (FN) and total hip (TH) were
evaluated in 460 healthy women | TL and BMD were not
associated, but a shorter TL could predict a lower BMD | qPCR |
| Sibille et
al (69), 2017 | Women without OA
but pain severity (n=136) | Shorter TL women
with chronic pain severity | Southern blot
analysis |
| Poonpet et
al (58), 2018 | Patients with knee
OA vs. healthy controls (n=140) | Negative
associations of angiogenetic cytokines with RTL | qPCR |
Therapeutic implications
To date, oxidative stress and chronic inflammation
in patients with OA and OP are considered to be the main reasons
leading to chondrocyte cellular senescence and apoptosis.
Putatively, targeted antioxidant treatment protects chondrocytes
and MSCs against oxidative stress-induced injury and associated
inflammation. In that context, Hudita et al, using an in
vitro scaffold-free 3-dimensional MSC culture model of
chondrogenesis, demonstrated that acetylated fatty acids mixture
from Celadrin reduced the secretion of inflammatory mediators and
facilitated the chondrogenic differentiation process of human
adipose-derived stem cells (70).
Moreover, the estimation of TL in chondrocytes and/or PBL is a
promising marker for the diagnosis and prognosis of OA.
Conclusions and future perspectives
OA and OP are two of the most common chronic
diseases affecting the aging population with significant associated
morbidity and mortality. The currently available therapies and
disease progression markers do not meet the needs of these
patients. Accumulating data indicate that telomere shortening may
contribute to OA and OP as an epigenetic factor. Consequently, the
measurement of TL of chondrocytes and/or PBL may prove to be
appropriate markers for the evaluation of the progression of these
diseases. It is important to identify the common mechanisms and
etiologies among these pathologies and the aging process (Fig. 1). Indeed, options preventing the
premature aging of mesenchymal cells could lead to novel therapies
which specifically target altered bone formation in OP and OA.
However, further studies with larger cohorts are required, in order
to obtain objective results and to enhance our understanding of the
association between TL, inflammation and aging. This may in turn
provide further insight into the pathophysiology of degenerative
joint diseases.
Acknowledgements
Not applicable.
Funding
This study was funded by Spin-Off Toxplus S.A. and
supported by the Special Research Account of University of Crete
(ELKE nos. 4602, 4920 and 3963).
Availability of data and materials
Not applicable.
Authors' contributions
All the authors (PR, DN, KK, ES, MT, PDS, CN, DAS,
TT and AT) contributed to the conception and design of the study.
PF, CN and KK searched the literature for inclusion in the study
that was then examined and reviewed by DN, ES and MT. PF and MT
drafted and wrote the manuscript. AT and TT provided advice and
critically revised the manuscript. All authors have read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
DAS is the Editor in Chief for the journal, but had
no personal involvement in the reviewing process, or any influence
in terms of adjudicating on the final decision, for this article.
The other authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
OA
|
osteoarthritis
|
|
OP
|
osteoporosis
|
|
BMD
|
bone mineral density
|
|
NSAIDs
|
non-steroidal anti-inflammatory
drugs
|
|
ROS
|
reactive oxygen species
|
|
ECM
|
extracellular matrix
|
|
TL
|
telomere length
|
|
TERT
|
telomerase reverse transcriptase
|
|
TERC
|
telomerase RNA component
|
|
CAD
|
cardiovascular disease
|
|
TRF
|
telomere restriction fragment
|
|
RTL
|
leukocyte relative telomere length
|
|
MSCs
|
mesenchymal stromal cells
|
|
BM
|
bone marrow
|
|
PBL
|
peripheral blood length
|
References
|
1
|
Panoutsopoulou K and Zeggini E: Advances
in osteoarthritis genetics. J Med Genet. 50:715–724. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yamasaki K, Nakasa T, Miyaki S, Ishikawa
M, Deie M, Adachi N, Yasunaga Y, Asahara H and Ochi M: Expression
of MicroRNA-146a in osteoarthritis cartilage. Arthritis Rheum.
60:1035–1041. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li H, Zeng C, Wei J, Yang T, Gao SG, Li
YS, Luo W, Xiao WF, Xiong YL and Lei GH: Serum Calcium
Concentration Is Inversely Associated With Radiographic Knee
Osteoarthritis: A Cross-Sectional Study. Medicine (Baltimore).
95:e28382016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Roman-Blas JA, Castañeda S, Largo R and
Herrero-Beaumont G: Osteoarthritis associated with estrogen
deficiency. Arthritis Res Ther. 11:2412009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dequeker J, Aerssens J and Luyten FP:
Osteoarthritis and osteoporosis: Clinical and research evidence of
inverse relationship. Aging Clin Exp Res. 15:426–439. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Franceschi C, Garagnani P, Morsiani C,
Conte M, Santoro A, Grignolio A, Monti D, Capri M and Salvioli S:
The continuum of aging and age-related diseases: common mechanisms
but different rates. Front Med (Lausanne). 5:612018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cosman F, de Beur SJ, LeBoff MS, Lewiecki
EM, Tanner B, Randall S and Lindsay R; National Osteoporosis
Foundation, : Clinician's Guide to Prevention and Treatment of
Osteoporosis. Osteoporos Int. 25:2359–2381. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sözen T, Özışık L and Başaran NC: An
overview and management of osteoporosis. Eur J Rheumatol. 4:46–56.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ganguly P, El-Jawhari JJ, Giannoudis PV,
Burska AN, Ponchel F and Jones EA: Age-related changes in bone
marrow mesenchymal stromal cells: A potential impact on
osteoporosis and osteoarthritis development. Cell Transplant.
26:1520–1529. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nelson AE: Osteoarthritis year in review
2017: Clinical. Osteoarthritis Cartilage. 26:319–325. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nikitovic D, Aggelidakis J, Young MF,
Iozzo RV, Karamanos NK and Tzanakakis GN: The biology of small
leucine-rich proteoglycans in bone pathophysiology. J Biol Chem.
287:33926–33933. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lotz M and Loeser RF: Effects of aging on
articular cartilage homeostasis. Bone. 51:241–248. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang M, Theleman JL, Lygrisse KA and Wang
J: Epigenetic mechanisms underlying the aging of articular
cartilage and osteoarthritis. Gerontology. 65:387–396. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Martin JA and Buckwalter JA: Roles of
articular cartilage aging and chondrocyte senescence in the
pathogenesis of osteoarthritis. Iowa Orthop J. 21:1–7.
2001.PubMed/NCBI
|
|
15
|
Ginaldi L, Di Benedetto MC and De Martinis
M: Osteoporosis, inflammation and ageing. Immun Ageing. 2:142005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Eghbali-Fatourechi G, Khosla S, Sanyal A,
Boyle WJ, Lacey DL and Riggs BL: Role of RANK ligand in mediating
increased bone resorption in early postmenopausal women. J Clin
Invest. 111:1221–1230. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
D'Amelio P, Grimaldi A, Di Bella S,
Brianza SZM, Cristofaro MA, Tamone C, Giribaldi G, Ulliers D,
Pescarmona GP and Isaia G: Estrogen deficiency increases
osteoclastogenesis up-regulating T cells activity: A key mechanism
in osteoporosis. Bone. 43:92–100. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ponzetti M and Rucci N: Updates on
Osteoimmunology: What's New on the Cross-Talk Between Bone and
Immune System. Front Endocrinol. 10:2362019. View Article : Google Scholar
|
|
19
|
Wauquier F, Leotoing L, Coxam V, Guicheux
J and Wittrant Y: Oxidative stress in bone remodelling and disease.
Trends Mol Med. 15:468–477. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sies H, Cadenas E, Symons MCR and Scott G:
Oxidative stress: Damage to intact cells and organs. Philos Trans R
Soc Lond B Biol Sci. 311:617–631. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Valko M, Leibfritz D, Moncol J, Cronin MT,
Mazur M and Telser J: Free radicals and antioxidants in normal
physiological functions and human disease. Int J Biochem Cell Biol.
39:44–84. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fischer-Nielsen A, Corcoran GB, Poulsen
HE, Kamendulis LM and Loft S: Menadione-induced DNA fragmentation
without 8-oxo-2′-deoxyguanosine formation in isolated rat
hepatocytes. Biochem Pharmacol. 49:1469–1474. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yermilov V, Rubio J, Becchi M, Friesen MD,
Pignatelli B and Ohshima H: Formation of 8-nitroguanine by the
reaction of guanine with peroxynitrite in vitro. Carcinogenesis.
16:2045–2050. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Masutomi K, Possemato R, Wong JM, Currier
JL, Tothova Z, Manola JB, Ganesan S, Lansdorp PM, Collins K and
Hahn WC: The telomerase reverse transcriptase regulates chromatin
state and DNA damage responses. Proc Natl Acad Sci USA.
102:8222–8227. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aubert G and Lansdorp PM: Telomeres and
aging. Physiol Rev. 88:557–579. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Harley CB, Vaziri H, Counter CM and
Allsopp RC: The telomere hypothesis of cellular aging. Exp
Gerontol. 27:375–382. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Saretzki G: Telomeres, Telomerase and
Ageing. Subcell Biochem. 90:221–308. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tsoukalas D, Fragkiadaki P, Docea AO,
Alegakis AK, Sarandi E, Thanasoula M, Spandidos DA, Tsatsakis A,
Razgonova MP and Calina D: Discovery of potent telomerase
activators: Unfolding new therapeutic and anti-aging perspectives.
Mol Med Rep. 20:3701–3708. 2019.PubMed/NCBI
|
|
29
|
Counter CM, Meyerson M, Eaton EN and
Weinberg RA: The catalytic subunit of yeast telomerase. Proc Natl
Acad Sci USA. 94:9202–9207. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Blackburn EH: Switching and signaling at
the telomere. Cell. 106:661–673. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kosebent EG, Uysal F and Ozturk S:
Telomere length and telomerase activity during folliculogenesis in
mammals. J Reprod Dev. 64:477–484. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu Y, Zhou L, Yang Y and Liu Y:
Cycloastragenol: An exciting novel candidate for age-associated
diseases. Exp Ther Med. 16:2175–2182. 2018.PubMed/NCBI
|
|
33
|
Bernardes B de Jesus, Schneeberger K, Vera
E, Tejera A, Harley CB and Blasco MA: The telomerase activator
TA-65 elongates short telomeres and increases health span of
adult/old mice without increasing cancer incidence. Aging Cell.
10:604–621. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fujii H, Shao L, Colmegna I, Goronzy JJ
and Weyand CM: Telomerase insufficiency in rheumatoid arthritis.
Proc Natl Acad Sci USA. 106:4360–4365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Vakonaki E, Tsiminikaki K, Plaitis S,
Fragkiadaki P, Tsoukalas D, Katsikantami I, Vaki G, Tzatzarakis MN,
Spandidos DA and Tsatsakis AM: Common mental disorders and
association with telomere length (Review). Biomed Rep. 8:111–116.
2018.PubMed/NCBI
|
|
36
|
Wu KD, Orme LM, Shaughnessy J Jr, Jacobson
J, Barlogie B and Moore MA: Telomerase and telomere length in
multiple myeloma: Correlations with disease heterogeneity,
cytogenetic status, and overall survival. Blood. 101:4982–4989.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Willeit P, Willeit J, Brandstätter A,
Ehrlenbach S, Mayr A, Gasperi A, Weger S, Oberhollenzer F, Reindl
M, Kronenberg F, et al: Cellular aging reflected by leukocyte
telomere length predicts advanced atherosclerosis and
cardiovascular disease risk. Arterioscler Thromb Vasc Biol.
30:1649–1656. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Calado RT and Young NS: Telomere diseases.
N Engl J Med. 361:2353–2365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fragkiadaki P, Tsoukalas D, Fragkiadoulaki
I, Psycharakis C, Nikitovic D, Spandidos DA and Tsatsakis AM:
Telomerase activity in pregnancy complications (Review). Mol Med
Rep. 14:16–21. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Vasilopoulos E, Fragkiadaki P, Kalliora C,
Fragou D, Docea AO, Vakonaki E, Tsoukalas D, Calina D, Buga AM,
Georgiadis G, et al: The association of female and male infertility
with telomere length (Review). Int J Mol Med. 44:375–389.
2019.PubMed/NCBI
|
|
41
|
Vakonaki E, Tzatzarakis M, Tsiminikai K,
Nathena D, Fragkiadaki P, Kalliantasi K, Kanaki K, Vaki G, Plaitis
S, Tsoukalas D, et al: Effect of chronic and heavy drug abuse on
biological aging. World Acad Sci J. 1:67–73. 2019.
|
|
42
|
Hemann MT, Strong MA, Hao LY and Greider
CW: The shortest telomere, not average telomere length, is critical
for cell viability and chromosome stability. Cell. 107:67–77. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Muraki K, Nyhan K, Han L and Murnane JP:
Mechanisms of telomere loss and their consequences for chromosome
instability. Front Oncol. 2:1352012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Thanasoula M, Escandell JM, Martinez P,
Badie S, Muñoz P, Blasco MA and Tarsounas M: p53 prevents entry
into mitosis with uncapped telomeres. Curr Biol. 20:521–526. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Thanasoula M, Escandell JM, Suwaki N and
Tarsounas M: ATM/ATR checkpoint activation downregulates CDC25C to
prevent mitotic entry with uncapped telomeres. EMBO J.
31:3398–3410. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tsatsakis A, Tsoukalas D, Fragkiadaki P,
Vakonaki E, Tzatzarakis M, Sarandi E, Nikitovic D, Tsilimidos G and
Alegakis AK: Developing BIOTEL: A Semi-Automated Spreadsheet for
Estimating Telomere Length and Biological Age. Front Genet.
10:842019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Oreffo RO, Bennett A, Carr AJ and Triffitt
JT: Patients with primary osteoarthritis show no change with ageing
in the number of osteogenic precursors. Scand J Rheumatol.
27:415–424. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Rodríguez JP, Garat S, Gajardo H, Pino AM
and Seitz G: Abnormal osteogenesis in osteoporotic patients is
reflected by altered mesenchymal stem cells dynamics. J Cell
Biochem. 75:414–423. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Benisch P, Schilling T, Klein-Hitpass L,
Frey SP, Seefried L, Raaijmakers N, Krug M, Regensburger M, Zeck S,
Schinke T, et al: The transcriptional profile of mesenchymal stem
cell populations in primary osteoporosis is distinct and shows
overexpression of osteogenic inhibitors. PLoS One. 7:e451422012.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhou Z, Gao M, Liu Q and Tao MD:
Comprehensive transcriptome analysis of mesenchymal stem cells in
elderly patients with osteoporosis. Aging Clin Exp Res. 27:595–601.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang H, Chen Q, Lee SH, Choi Y, Johnson FB
and Pignolo RJ: Impairment of osteoblast differentiation due to
proliferation-independent telomere dysfunction in mouse models of
accelerated aging. Aging Cell. 11:704–713. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Valdes AM, Richards JB, Gardner JP,
Swaminathan R, Kimura M, Xiaobin L, Aviv A and Spector TD: Telomere
length in leukocytes correlates with bone mineral density and is
shorter in women with osteoporosis. Osteoporos Int. 18:1203–1210.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Bekaert S, Van Pottelbergh I, De Meyer T,
Zmierczak H, Kaufman JM, Van Oostveldt P and Goemaere S: Telomere
length versus hormonal and bone mineral status in healthy elderly
men. Mech Ageing Dev. 126:1115–1122. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kveiborg M, Kassem M, Langdahl B, Eriksen
EF, Clark BFC and Rattan SIS: Telomere shortening during aging of
human osteoblasts in vitro and leukocytes in vivo: Lack of
excessive telomere loss in osteoporotic patients. Mech Ageing Dev.
106:261–271. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tao L, Huang Q, Yang R, Dai Y, Zeng Y, Li
C, Li X, Zeng J and Wang Q: The age modification to leukocyte
telomere length effect on bone mineral density and osteoporosis
among Chinese elderly women. J Bone Miner Metab. 37:1004–1012.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Calmy A, Chevalley T, Delhumeau C,
Toutous-Trellu L, Spycher-Elbes R, Ratib O, Zawadynski S and
Rizzoli R: Long-term HIV infection and antiretroviral therapy are
associated with bone microstructure alterations in premenopausal
women. Osteoporos Int. 24:1843–1852. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
McCulloch K, Litherland GJ and Rai TS:
Cellular senescence in osteoarthritis pathology. Aging Cell.
16:210–218. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Poonpet T, Saetan N, Tanavalee A,
Wilairatana V, Yuktanandana P and Honsawek S: Association between
leukocyte telomere length and angiogenic cytokines in knee
osteoarthritis. Int J Rheum Dis. 21:118–125. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yudoh K, Nguyen vT, Nakamura H,
Hongo-Masuko K, Kato T and Nishioka K: Potential involvement of
oxidative stress in cartilage senescence and development of
osteoarthritis: oxidative stress induces chondrocyte telomere
instability and downregulation of chondrocyte function. Arthritis
Res Ther. 7:R380–R391. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
60
|
Martin JA and Buckwalter JA: Aging,
articular cartilage chondrocyte senescence and osteoarthritis.
Biogerontology. 3:257–264. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Price JS, Waters JG, Darrah C, Pennington
C, Edwards DR, Donell ST and Clark IM: The role of chondrocyte
senescence in osteoarthritis. Aging Cell. 1:57–65. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhai G, Aviv A, Hunter DJ, Hart DJ,
Gardner JP, Kimura M, Lu X, Valdes AM and Spector TD: Reduction of
leucocyte telomere length in radiographic hand osteoarthritis: A
population-based study. Ann Rheum Dis. 65:1444–1448. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li J, Huang J, Dai L, Yu D, Chen Q, Zhang
X and Dai K: miR-146a, an IL-1β responsive miRNA, induces vascular
endothelial growth factor and chondrocyte apoptosis by targeting
Smad4. Arthritis Res Ther. 14:R752012. View
Article : Google Scholar : PubMed/NCBI
|
|
64
|
Tamayo M, Mosquera A, Rego JI,
Fernández-Sueiro JL, Blanco FJ and Fernández JL: Differing patterns
of peripheral blood leukocyte telomere length in rheumatologic
diseases. Mutat Res. 683:68–73. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Tamayo M, Mosquera A, Rego I, Blanco FJ,
Gosálvez J and Fernández JL: Decreased length of telomeric DNA
sequences and increased numerical chromosome aberrations in human
osteoarthritic chondrocytes. Mutat Res. 708:50–58. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Aigner T, Haag J, Martin J and Buckwalter
J: Osteoarthritis: Aging of matrix and cells--going for a remedy.
Curr Drug Targets. 8:325–331. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
McAlindon T, Roberts M, Driban J, Schaefer
L, Haugen IK, Smith SE, Duryea J, Cunha D, Blanco F,
Fernández-Garcia JL, et al: Incident hand OA is strongly associated
with reduced peripheral blood leukocyte telomere length.
Osteoarthritis Cartilage. 26:1651–1657. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Harbo M, Delaisse JM, Kjaersgaard-Andersen
P, Soerensen FB, Koelvraa S and Bendix L: The relationship between
ultra-short telomeres, aging of articular cartilage and the
development of human hip osteoarthritis. Mech Ageing Dev.
134:367–372. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Sibille KT, Chenc H, Bartley EJ, Riley J
III, Gloverd TL, King CD, Zhang H, Cruz-Almeid Y, Goodin BR,
Sotolongo A, et al: Accelerated aging in adults with knee
osteoarthritis pain: consideration for frequency, intensity, time,
and total pain sites. Pain Rep. 2:e5912017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Hudita A, Galateanu B, Dinescu S, Costache
M, Dinischiotu A, Negrei C, Stan M, Tsatsakis A, Nikitovic D,
Lupuliasa D, et al: In Vitro Effects of Cetylated Fatty Acids
Mixture from Celadrin on Chondrogenesis and Inflammation with
Impact on Osteoarthritis. Cartilage. 11:88–97. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Sanders JL, Cauley JA, Boudreau RM, Zmuda
JM, Strotmeyer ES, Opresko PL, Hsueh WC, Cawthon RM, Li R, Harris
TB, et al Health ABC Study, : Leukocyte Telomere Length Is Not
Associated With BMD, Osteoporosis, or Fracture in Older Adults:
Results From the Health, Aging and Body Composition Study. J Bone
Miner Res. 24:1531–1536. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Tang NL, Woo J, Suen EW, Liao CD, Leung JC
and Leung PC: The effect of telomere length, a marker of biological
aging, on bone mineral density in elderly population. Osteoporos
Int. 21:89–97. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Nielsen BR, Linneberg A, Bendix L, Harboe
M, Christensen K and Schwarz P: Association between leukocyte
telomere length and bone mineral density in women 25–93 years of
age. Exp Gerontol. 66:25–31. 2015. View Article : Google Scholar : PubMed/NCBI
|