Introduction
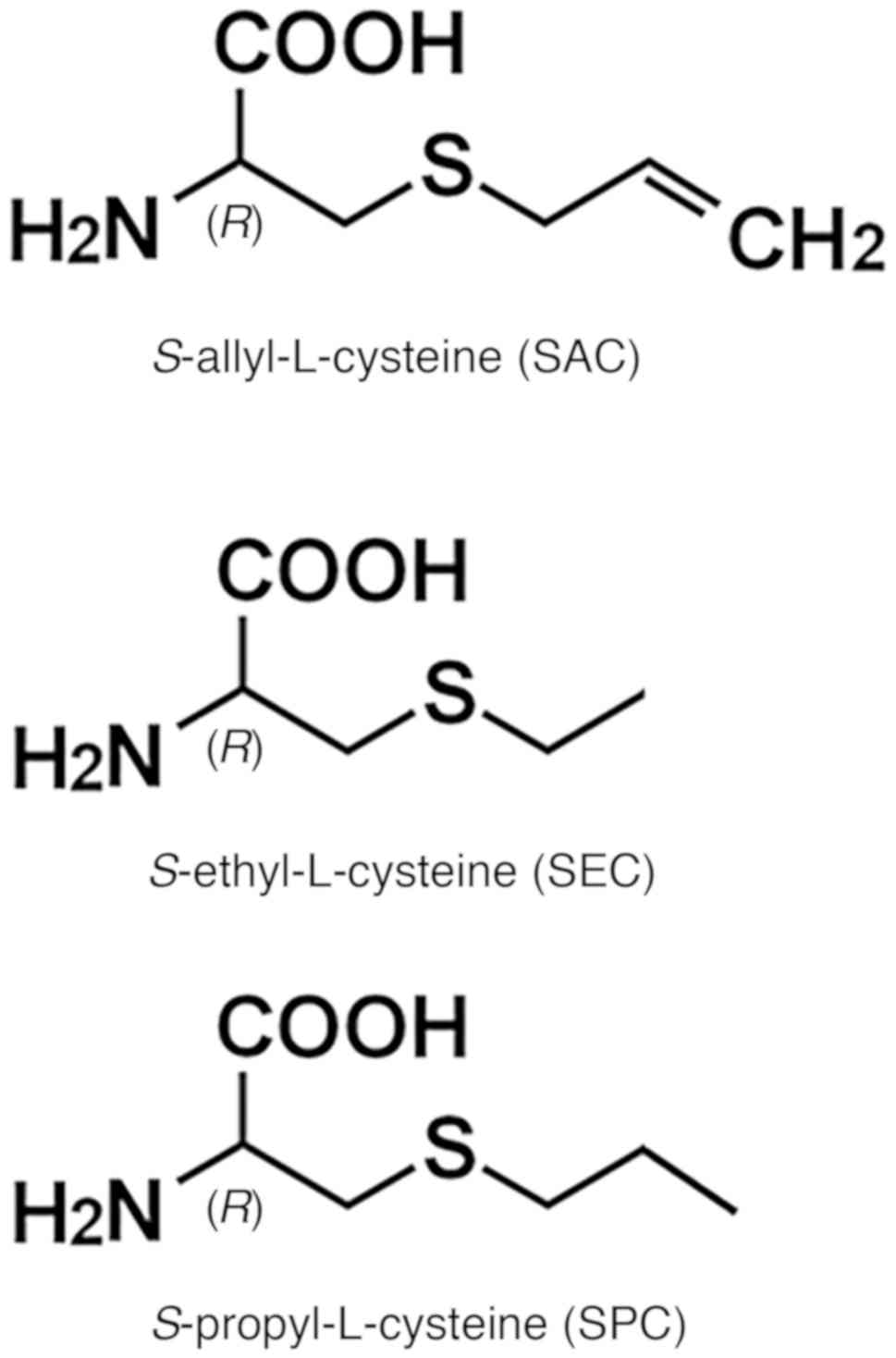
S-allyl-L-cysteine (SAC) is the most abundant
organosulfur compound derived from garlic (Allium sativum)
and is biosynthesized by hydrolysis of
γ-glutamyl-S-allyl-cysteine (GSAC) by the enzyme γ-glutamyl
transpeptidase (γGTP). SAC is known as a water soluble bioactive
compound of extremely high antioxidant capacity, and has long been
used as a common dietary supplement and in traditional medicine
(1). Moreover, SAC has been reported
to exert multiple biological effects, such as anti-diabetic
(2), cholesterol-lowering (3), anticancer (4) and anti-hepatotoxic (5) effects. SAC and other organosulfur
compounds are found in aged garlic extract (AGE), which is produced
by natural extraction from fresh garlic followed by incubation for
>10 months in aqueous ethanol. In cultures of dissociated rat
hippocampal neurons, these compounds have been shown to promote
neuronal survival and to increase the number of branching points
per axon (6). The chronic dietary
intake of a low dose of SAC (40 mg/kg) has been reported to
ameliorate deficits in learning performance and memory
consolidation normally observed in the senescence-accelerated mouse
strains, SAMP8 and SAMP10 (7).
Recently, Baluchnejadmojarad et al demonstrated that chronic
oral treatment with SAC (150 mg/kg) ameliorated cognitive deficits
in a rat model of streptozotocin-induced diabetes by suppressing
oxidative stress and neuroinflammation (8). Despite the accumulating knowledge of
the properties of SAC in the presence of oxidative stress, little
is known as to whether SAC ameliorates stress conditions other than
oxidative stress, as well as the underlying mechanisms.
The endoplasmic reticulum (ER) is the largest
organelle of most eukaryotic cells and plays crucial roles in
various cellular processes, such as the synthesis, transport and
correct folding of proteins, as well as lipid synthesis,
carbohydrate metabolism and intracellular Ca2+ storage.
Various physiological and pathological conditions, such as glucose
deprivation, Ca2+ depletion and exposure to free
radicals lead to the accumulation of misfolded or unfolded proteins
in the ER, a condition known as ER stress (9). ER stress has been implicated in various
neurological diseases, including brain ischemia, traumatic brain
injury, and age-associated neurodegeneration. ER stress-induced
cell death has been shown to be involved in various
neurodegenerative diseases, such as Alzheimer's disease,
Parkinson's disease, amyotrophic lateral sclerosis and Huntington's
disease (10). Thus, it is possible
that a pharmacological agent targeting the intracellular signaling
of ER stress could provide a potential therapeutic approach for
neurodegeneration and brain damage. The purpose of the present
review was to summarize the neuroprotective mechanisms of SAC in
relation to ER stress and to discuss the prospects of SAC becoming
the prototype of a new type of therapeutic drug for
neurodegenerative diseases linked to ER stress.
Protective effects of SAC against neuronal
death due to ER stress
ER stress can be activated by various pathological
and physiological conditions. The accumulation of misfolded or
unfolded proteins in the ER activates a set of signaling pathways
termed the unfolded protein response (UPR). The UPR can promote
cellular repair and survival by reducing the load of unfolded
proteins through the upregulation of chaperones and the attenuation
of protein synthesis (11). However,
when adaptive responses fail to resolve ER stress, the UPR
ultimately initiates multiple, ER-specific pathways to induce
apoptosis. In recent years, several lines of evidence have
implicated several proteins in ER stress-induced cell death, such
as C/EBP homologous protein (CHOP), also known as growth arrest and
DNA damage-inducible protein 153 (GADD153), caspase-12 and
apoptosis-signal-regulating kinase 1 (ASK-1) (12). Among these stress-related factors,
caspase-12 is localized specifically on the cytoplasmic side of the
ER and has been shown to be proteolytically activated under
conditions of ER stress in rodents (13,14).
Mutant mice lacking the caspase-12 gene are resistant to chemical
ER stress inducers such as tunicamycin (an inhibitor of protein
glycosylation), thapsigargin (an inhibitor of ER-associated
Ca2+ ATPase) and brefeldin A (an inhibitor of
ER-to-Golgi transport) (14).
Previous studies by our group have demonstrated that neuronal death
is mediated by caspase-12 when death is induced either by amyloid
β-peptide (abbreviated Aβ, and a key player in the pathology of
Alzheimer's disease), or by tunicamycin, in both rat cultured
hippocampal neurons (15,16) and rat organotypic hippocampal slice
cultures (17,18). We have also previously demonstrated
that the cell-permeable caspase-12-selective inhibitor, z-ATAD-fmk,
significantly suppresses cell death induced by tunicamycin in
organotypic hippocampal slice cultures (17,18).
Moreover, previous studies conducted in our laboratory have
revealed that SAC protects against Aβ- and tunicamycin-induced cell
death in 3 cell lines: PC12 cells differentiated by nerve growth
factor (NGF) (19), cultured
hippocampal neurons (15,16,20) and
organotypic hippocampal slice cultures (17,18).
Moreover, the increases in cleaved, activated caspase-12 induced by
Aβ and tunicamycin have been shown to be prevented by the
simultaneous application of SAC (15). The lipid peroxidation product,
4-hydroxynonenal, associated with oxidative stress, has been shown
to play a pivotal role in the pathogenesis of a number of
neurodegenerative disorders. However, SAC was shown to not prevent
4-hydroxynonenal-induced cell death in NGF-differentiated PC12
cells (19) or in cultured
hippocampal neurons (15). Taken
together, these results strongly suggest that SAC exerts a
significant neuroprotective effect against ER stress-induced
neuronal death by attenuating the activation of caspase-12.
Calpain is a probable target molecule for
SAC
Three main mechanisms for caspase-12 activation by
ER stress signals have been discovered thus far. First, caspase-12
forms a stable complex in the ER membrane with inositol-requiring
enzyme 1 (IRE1) and the adapter protein tumor necrosis factor
receptor-associated factor 2 (TRAF2) (21). The dissociation of TRAF2 from
caspase-12 is reportedly a trigger for the activation of caspase-12
under ER stress conditions (21).
Second, Rao et al have reported that in the 293T cell line,
caspase-7 translocates from the cytosol to the ER membrane in
response to ER stress, which leads to the processing of
procaspase-12 and activation of caspase-12 (22). Third and most importantly, calpain, a
cytoplasmic cysteine protease, has also been shown to cleave and
activate caspase-12, which it does in response to Ca2+
release from the ER during ER stress (13). Furthermore, embryonic fibroblasts
derived from conditional calpain knockout mice have been shown to
be resistant to ER stress-induced cell death, which is due to
resistance to caspase-12 activation (23). Moreover, the calpain inhibitor,
PD150606, has been shown to inhibit tunicamycin-induced cell death
in the kidney cell line, LLC-PK1 (24). These results suggest that the calpain
activation pathway dominates in caspase-12-dependent, ER
stress-induced cell death.
To further support this concept, we previously
evaluated the effects of SAC on ER stress-induced calpain
activation in cultured hippocampal neurons (20). Calpain activity can be measured by
the quantitative analysis of full-length α-spectrin, a
neuron-specific cytoskeletal protein, which undergoes proteolysis
by activated calpain. The exposure of cultured hippocampal neurons
to tunicamycin for 24 h was shown to result in a decrease in the
levels of full-length α-spectrin. The degradation of α-spectrin was
significantly prevented by simultaneous treatment with SAC
(20). We have also previously
examined the effects of SAC on activation of recombinant calpain in
a cell-free assay system containing a synthetic substrate for
calpain, Suc-LLVY-Glo (20). The
calpain activity was inhibited by SAC in a concentration-dependent
manner, as observed with typical synthetic calpain inhibitors such
as calpeptin, ALLN and PD150606 (20). At the higher concentrations tested
(ALLN, 10 nM; PD150606, 100 mM), these synthetic calpain inhibitors
completely inhibited calpain activity in our in vitro assay
system. SAC partially inhibited calpain activity at still higher
concentrations (10 mM) (20). These
findings suggest that, at least in the hippocampus, the
neuroprotective effects of SAC against ER stress are partly
attributable to the direct inhibition of calpain activity.
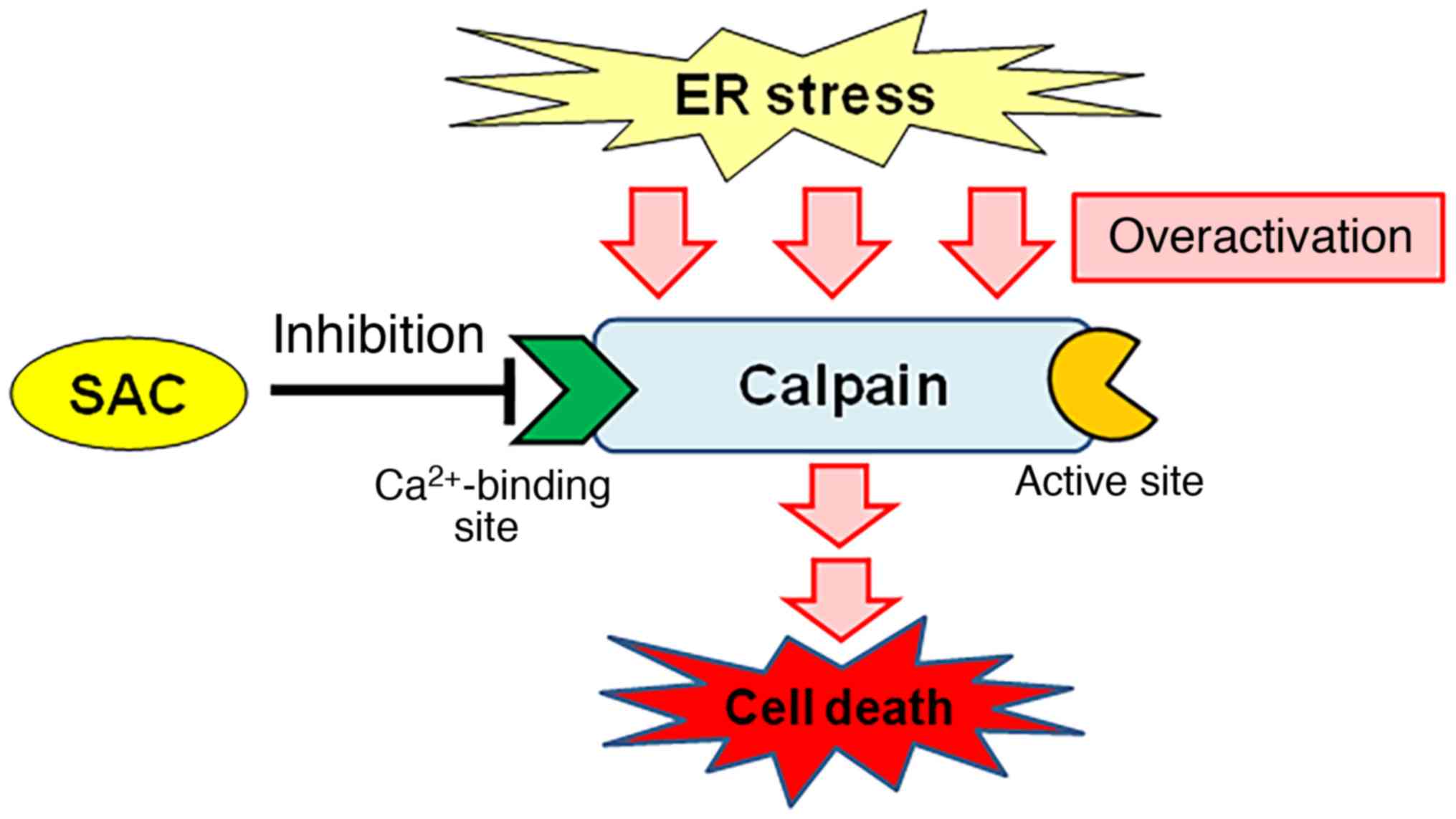
To characterize the site of interaction of SAC with
calpain, we previously examined the interactions of SAC with
synthetic calpain inhibitors that have known calpain interaction
sites: ALLN, which interferes with the active site of calpain, and
PD150606, which interacts with the Ca2+-binding site
(20,25,26).
When submaximal concentrations of synthetic calpain inhibitors were
applied in the in vitro assay system, ALLN (5 pM) in
combination with SAC additively inhibited calpain activity, whereas
the inhibitory effect of PD150606 (25 µM) was unaffected (20). By contrast, at supramaximal
concentrations of synthetic calpain inhibitors, SAC significantly
reversed the inhibitory effects of PD150606 (100 µM), but did not
affect inhibition by ALLN (1 nM) (20). Taken together, these results suggest
that SAC interacts with the Ca2+-binding site of calpain
(Fig. 1). This finding of a novel
target for SAC strongly suggests that the limited concept of SAC as
a mere antioxidant agent must be expanded to include the property
of SAC of inhibiting ER stress. Although further in vivo
studies should be carried out to clarify the underlying mechanisms
responsible for the neuroprotective effects of SAC, the calpain
inhibitory potential of SAC may prove to be useful in deriving
therapeutic agents for neurological disorders associated with ER
stress or the overactivation of calpain.
Analogs of SAC
In addition to SAC, various other sulfur-substituted
compounds are abundant in the genus, Allium (27). Among these are
S-methyl-L-cysteine (SMC), S-ethyl-L-cysteine (SEC)
and S-propyl-L-cysteine (SPC), three typical hydrophilic
cysteine-containing compounds naturally found in garlic (27). More recently, we synthesized various
structurally related derivatives of SAC and examined their
neuroprotective effects in a model of ER stress based on
tunicamycin-induced neurotoxicity in rat cultured hippocampal
neurons (28). The following 20 SAC
derivatives were tested: SAC itself, S-allyl-D-cysteine
(ent-SAC), S-allyl-L-cysteine amide (SACA),
S-benzyl-L-cysteine (SBC), S-benzyl-D-cysteine
(ent-SBC), S-carboxymethyl-L-cysteine (SCMC), SEC,
S-allyl-DL-homocysteine (rac-SAHC), SMC, SPC,
S-propynyl-L-cysteine (SPNC), S-trityl-L-cysteine
(STC), methyl S-allyl-L-cysteinate (MSAC), methyl
O-allyl-L-serinate (MOAS), methyl
O-allyl-N-boc-L-serinate (MOANBS),
N-acetyl-S-allyl-L-cysteine (NASAC),
N-acetyl-S-allyl-L-cysteine amide (NASACA),
O-allyl-L-serine (OAS) and O-allyl-L-serine amide
(OASA). Cultured hippocampal neurons were treated with tunicamycin
(10 µg/ml) in the absence or presence of 1 µM of a SAC derivative.
These experiments demonstrated that the neuroprotective effects of
the side-chain substituted SAC derivatives, SEC and SPC, against ER
stress-induced cytotoxicity were more potent than those of SAC
(Fig. 2). By contrast, compounds
with bulky substituents (SBC, ent-SBC, SCMC, SPNC and STC)
exerted neurotoxic effects. We have also previously demonstrated
that several compounds with modified carboxyl or amino groups
(SACA, rac-SAHC and NASAC) are as effective as SAC. Unlike
SAC, SEC and SPC did not block calpain activity in the cell-free
assay system, suggesting that the mechanism underlying the
protective activity of SEC and SPC differs from that of SAC. We
have also demonstrated that tunicamycin-induced cell death in
cultured hippocampal neurons is not mediated by the oxidative
stress cascade (20). Thus, we are
left with the possibility that in hippocampal neurons, the
stimulation of survival cascades other than the antioxidant
defensive system underlies the neuroprotective action of SAC
derivatives, such as SEC and SPC. Although further detailed studies
are required to clarify the underlying mechanisms, it now appears
that the number of carbon atoms in the linear 3-alkylthio groups of
some SAC derivatives is key in potentiating neuroprotective effects
against ER stress.
Conclusions and future perspectives
The pharmacokinetics of SAC are characterized by
high oral absorption, limited metabolism, and extensive renal
reabsorption, all of which potentially contribute to its high oral
bioavailability (29). Moreover, SAC
has already been used in the treatment of patients with
hypertension without any obvious signs of toxicity (30). These results support the hypothesis
that SAC may be a useful therapeutic agent with few harmful
effects. Our group, as well as others have shown that SAC has
neuroprotective potency in in vivo and in vitro in
models of brain injury. SAC has been shown to ameliorate the
neuronal damage usually observed in a rat model of transient middle
cerebral artery occlusion (MCAO) (31). In a transgenic mouse model of
Alzheimer's disease, SAC was shown to prevent synaptic degeneration
(32) and the abnormal
phosphorylation of tau protein (33). The dietary intake of SAC, SEC and SPC
has been shown to decrease the production of Aβ in the brains of
mice with D-galactose-induced aging (34). Similar cysteine-containing compounds
exert neuroprotective effects against the loss of dopaminergic
neurons in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)
mouse model of Parkinson's disease (35,36).
Oxidative stress plays a critical role in the pathogenesis of the
neurodegenerative diseases modeled above, including Alzheimer's
disease, Parkinson's disease and cerebral ischemic insults.
However, a growing body of evidence indicates that oxidative stress
has a strong connection with ER stress (37). Importantly, intracellular
Ca2+ is a prime candidate for mediating the crosstalk
between ER stress and oxidative stress. The surprising finding that
calpain is a therapeutic target for SAC strongly suggests that SAC
is not merely an antioxidant, but is also an inhibitor of ER stress
signaling. Given the probable existence of intracellular
Ca2+-mediated, ER-oxidative stress crosstalk, the fact
that SAC blocks the calpain calcium-sensing site appears
therapeutically advantageous. Moreover, the two-pronged activity
profile appears ideally suited to treating a wide spectrum of
neurodegenerative diseases in which both types of stress play a
role. Finally, the chemical modification of the parent molecule has
potential for achieving desirable changes in the pharmacodynamic
profile. Although further in vivo detailed studies
culminating in a clinical study are required, SAC and its
derivatives, in their roles as both antioxidants and modulators of
ER stress, can now be identified as good candidate drugs for the
treatment of patients with neurodegenerative disorders. This
concise review may provide new insight into the therapeutic potency
of garlic-derived organosulfur compounds and may lead to the
development of novel treatment strategies for patients with
otherwise intractable neurological disorders.
Acknowledgements
The author would like to thank Editage (www.editage.com) for English language editing.
Funding
This study was supported in part by JSPS KAKENHI
[Grant nos. JP26460634 and JP17K08965], by a Nihon University
Chairman of the Board of Trustees Grant, and by a ‘Private
University Research Branding Project’ from MEXT. The funding bodies
had no role in the design of the study or in the writing of the
manuscript.
Availability of data and materials
Not applicable.
Authors' contributions
YK designed the review, collected the information,
drafted the manuscript, and finalized the manuscript. The author
has read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The author declares that he has no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AGE
|
aged garlic extract
|
|
ASK-1
|
apoptosis signal-regulating kinase
1
|
|
Aβ
|
amyloid β-peptide
|
|
boc
|
tert-butyl carbamate group
|
|
CHOP
|
C/EBP homologous protein
|
|
ent
|
enantiomeric
|
|
ent-SAC
|
S-allyl-D-cysteine
|
|
ent-SBC
|
S-benzyl-D-cysteine
|
|
ER
|
endoplasmic reticulum
|
|
GADD153
|
growth arrest and DNA damage-inducible
protein 153
|
|
GSAC
|
γ-glutamyl-S-allyl-cysteine
|
|
IRE1
|
inositol-requiring enzyme 1
|
|
MCAO
|
middle cerebral artery occlusion
|
|
MOANBS
|
methyl
O-allyl-N-boc-L-serinate
|
|
MOAS
|
methyl O-allyl-L-serinate
|
|
MPTP
|
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine
|
|
MSAC
|
methyl S-allyl-L-cysteinate
|
|
NASAC
|
N-acetyl-S-allyl-L-cysteine
|
|
NASACA
|
N-acetyl-S-allyl-L-cysteine amide
|
|
NGF
|
nerve growth factor
|
|
OAS
|
O-allyl-L-serine
|
|
OASA
|
O-allyl-L-serine amide
|
|
rac
|
racemic
|
|
rac-SAHC
|
S-allyl-DL-homocysteine
|
|
SAC
|
S-allyl-L-cysteine
|
|
SACA
|
S-allyl-L-cysteine amide
|
|
SBC
|
S-benzyl-L-cysteine
|
|
SCMC
|
S-carboxymethyl-L-cysteine
|
|
SEC
|
S-ethyl-L-cysteine
|
|
SMC
|
S-methyl-L-cysteine
|
|
SPC
|
S-propyl-L-cysteine
|
|
SPNC
|
S-propynyl-L-cysteine
|
|
STC
|
S-trityl-L-cysteine
|
|
TRAF2
|
tumor necrosis factor
receptor-associated factor 2
|
|
UPR
|
unfolded protein response
|
|
γGTP
|
γ-glutamyl transpeptidase
|
References
|
1
|
Colín-González AL, Santana RA, Silva-Islas
CA, Chánez-Cárdenas ME, Santamaría A and Maldonado PD: The
antioxidant mechanisms underlying the aged garlic extract- and
S-allylcysteine-induced protection. Oxid Med Cell Longev.
2012:9071622012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Saravanan G and Ponmurugan P: Antidiabetic
effect of S-allylcysteine: Effect on thyroid hormone and
circulatory antioxidant system in experimental diabetic rats. J
Diabetes Complications. 26:280–285. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yeh YY and Liu L: Cholesterol-lowering
effect of garlic extracts and organosulfur compounds: Human and
animal studies. J Nutr. 131 (Suppl 3):989S–993S. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thomson M and Ali M: Garlic [Allium
sativum]: A review of its potential use as an anti-cancer
agent. Curr Cancer Drug Targets. 3:67–81. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kodai S, Takemura S, Minamiyama Y, Hai S,
Yamamoto S, Kubo S, Yoshida Y, Niki E, Okada S, Hirohashi K, et al:
S-allyl cysteine prevents CCl(4)-induced acute liver injury
in rats. Free Radic Res. 41:489–497. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Moriguchi T, Matsuura H, Kodera Y, Itakura
Y, Katsuki H, Saito H and Nishiyama N: Neurotrophic activity of
organosulfur compounds having a thioallyl group on cultured rat
hippocampal neurons. Neurochem Res. 22:1449–1452. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nishiyama N, Moriguchi T, Morihara N and
Saito H: Ameliorative effect of S-allylcysteine, a major
thioallyl constituent in aged garlic extract, on learning deficits
in senescence-accelerated mice. J Nutr. 131 (Suppl 3):1093S–1095S.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baluchnejadmojarad T, Kiasalari Z,
Afshin-Majd S, Ghasemi Z and Roghani M: S-allyl cysteine
ameliorates cognitive deficits in streptozotocin-diabetic rats via
suppression of oxidative stress, inflammation, and
acetylcholinesterase. Eur J Pharmacol. 794:69–76. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Paschen W: Endoplasmic reticulum: A
primary target in various acute disorders and degenerative diseases
of the brain. Cell Calcium. 34:365–383. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Placido AI, Pereira CM, Duarte AI,
Candeias E, Correia SC, Carvalho C, Cardoso S, Oliveira CR and
Moreira PI: Modulation of endoplasmic reticulum stress: An
opportunity to prevent neurodegeneration? CNS Neurol Disord Drug
Targets. 14:518–533. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schröder M and Kaufman RJ: The mammalian
unfolded protein response. Annu Rev Biochem. 74:739–789. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gorman AM, Healy SJ, Jäger R and Samali A:
Stress management at the ER: Regulators of ER stress-induced
apoptosis. Pharmacol Ther. 134:306–316. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nakagawa T and Yuan J: Cross-talk between
two cysteine protease families. Activation of caspase-12 by calpain
in apoptosis. J Cell Biol. 150:887–894. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nakagawa T, Zhu H, Morishima N, Li E, Xu
J, Yankner BA and Yuan J: Caspase-12 mediates
endoplasmic-reticulum-specific apoptosis and cytotoxicity by
amyloid-beta. Nature. 403:98–103. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kosuge Y, Koen Y, Ishige K, Minami K,
Urasawa H, Saito H and Ito Y: S-allyl-L-cysteine selectively
protects cultured rat hippocampal neurons from amyloid
beta-protein- and tunicamycin-induced neuronal death. Neuroscience.
122:885–895. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kosuge Y, Sakikubo T, Ishige K and Ito Y:
Comparative study of endoplasmic reticulum stress-induced neuronal
death in rat cultured hippocampal and cerebellar granule neurons.
Neurochem Int. 49:285–293. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Imai T, Kosuge Y, Ishige K and Ito Y:
Amyloid beta-protein potentiates tunicamycin-induced neuronal death
in organotypic hippocampal slice cultures. Neuroscience.
147:639–651. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kosuge Y, Imai T, Kawaguchi M, Kihara T,
Ishige K and Ito Y: Subregion-specific vulnerability to endoplasmic
reticulum stress-induced neurotoxicity in rat hippocampal neurons.
Neurochem Int. 52:1204–1211. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ito Y, Kosuge Y, Sakikubo T, Horie K,
Ishikawa N, Obokata N, Yokoyama E, Yamashina K, Yamamoto M, Saito
H, et al: Protective effect of S-allyl-L-cysteine, a garlic
compound, on amyloid beta-protein-induced cell death in nerve
growth factor-differentiated PC12 cells. Neurosci Res. 46:119–125.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Imai T, Kosuge Y, Endo-Umeda K, Miyagishi
H, Ishige K, Makishima M and Ito Y: Protective effect of
S-allyl-L-cysteine against endoplasmic reticulum
stress-induced neuronal death is mediated by inhibition of calpain.
Amino Acids. 46:385–393. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yoneda T, Imaizumi K, Oono K, Yui D, Gomi
F, Katayama T and Tohyama M: Activation of caspase-12, an
endoplastic reticulum (ER) resident caspase, through tumor necrosis
factor receptor-associated factor 2-dependent mechanism in response
to the ER stress. J Biol Chem. 276:13935–13940. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rao RV, Hermel E, Castro-Obregon S, del
Rio G, Ellerby LM, Ellerby HM and Bredesen DE: Coupling endoplasmic
reticulum stress to the cell death program. Mechanism of caspase
activation. J Biol Chem. 276:33869–33874. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tan Y, Dourdin N, Wu C, De Veyra T, Elce
JS and Greer PA: Ubiquitous calpains promote caspase-12 and JNK
activation during endoplasmic reticulum stress-induced apoptosis. J
Biol Chem. 281:16016–16024. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Muruganandan S and Cribb AE:
Calpain-induced endoplasmic reticulum stress and cell death
following cytotoxic damage to renal cells. Toxicol Sci. 94:118–128.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Carragher NO: Calpain inhibition: A
therapeutic strategy targeting multiple disease states. Curr Pharm
Des. 12:615–638. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang KK, Nath R, Posner A, Raser KJ,
Buroker-Kilgore M, Hajimohammadreza I, Probert AW Jr, Marcoux FW,
Ye Q, Takano E, et al: An alpha-mercaptoacrylic acid derivative is
a selective nonpeptide cell-permeable calpain inhibitor and is
neuroprotective. Proc Natl Acad Sci USA. 93:6687–6692. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jones MG, Hughes J, Tregova A, Milne J,
Tomsett AB and Collin HA: Biosynthesis of the flavour precursors of
onion and garlic. J Exp Bot. 55:1903–1918. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Imai T, Kosuge Y, Saito H, Uchiyama T,
Wada T, Shimba S, Ishige K, Miyairi S, Makishima M and Ito Y:
Neuroprotective effect of S-allyl-l-cysteine derivatives
against endoplasmic reticulum stress-induced cytotoxicity is
independent of calpain inhibition. J Pharmacol Sci. 130:185–188.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Amano H, Kazamori D and Itoh K:
Pharmacokinetics of S-Allyl-l-cysteine in rats is
characterized by high oral absorption and extensive renal
reabsorption. J Nutr. 146:456S–459S. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ried K, Frank OR and Stocks NP: Aged
garlic extract lowers blood pressure in patients with treated but
uncontrolled hypertension: A randomised controlled trial.
Maturitas. 67:144–150. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Atif F, Yousuf S and Agrawal SK:
S-allyl L-cysteine diminishes cerebral ischemia-induced
mitochondrial dysfunctions in hippocampus. Brain Res. 1265:128–137.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ray B, Chauhan NB and Lahiri DK: Oxidative
insults to neurons and synapse are prevented by aged garlic extract
and S-allyl-L-cysteine treatment in the neuronal culture and
APP-Tg mouse model. J Neurochem. 117:388–402. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chauhan NB: Effect of aged garlic extract
on APP processing and tau phosphorylation in Alzheimer's transgenic
model Tg2576. J Ethnopharmacol. 108:385–394. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tsai SJ, Chiu CP, Yang HT and Yin MC:
S-allyl cysteine, S-ethyl cysteine, and
S-propyl cysteine alleviate β-amyloid, glycative, and
oxidative injury in brain of mice treated by D-galactose. J Agric
Food Chem. 59:6319–6326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen CM, Yin MC, Hsu CC and Liu TC:
Antioxidative and anti-inflammatory effects of four
cysteine-containing agents in striatum of MPTP-treated mice.
Nutrition. 23:589–597. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
García E, Santana-Martínez R, Silva-Islas
CA, Colín-González AL, Galván-Arzate S, Heras Y, Maldonado PD,
Sotelo J and Santamaría A: S-allyl cysteine protects against
MPTP-induced striatal and nigral oxidative neurotoxicity in mice:
Participation of Nrf2. Free Radic Res. 48:159–167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nakka VP, Prakash-Babu P and Vemuganti R:
Crosstalk between endoplasmic reticulum stress, oxidative stress,
and autophagy: potential therapeutic targets for acute CNS
injuries. Mol Neurobiol. 53:532–544. 2016. View Article : Google Scholar : PubMed/NCBI
|