Introduction
Primary nephrotic syndrome (PNS) is the most common
chronic kidney disease in children and is also the main cause of
chronic renal failure in adults in China (1,2).
Previous studies have found that the cause of chronic kidney
diseases in adults originates from PNS during childhood, which can
be alleviated by treatment (3,4). The
etiology and pathogenesis of childhood PNS remain unclear. However,
it is well-documented that proteinuria, hypoproteinemia,
hyperlipidemia and edema are the main pathophysiological
characteristics associated with PNS, with proteinuria being the
core change (2,5). Damage to the glomerular filtration
barrier is the direct cause of proteinuria, where the abnormal
number or function of podocytes are a cause of pathological changes
in glomerular filtration membrane permeability, proteinuria
formation and ultimately glomerular sclerosis (6,7).
Podocyte injuries have been reported to participate in the
formation of urinary fistula and glomerular sclerosis, such that
signs of podocyte injury have been observed in the urine of
patients with primary and secondary glomerular disease (8,9).
Additionally, previous studies have found that a variety of
factors, including anti-capsular antibodies, metabolic factors,
hemodynamics and infection, can result in podocyte damage (10,11).
Interleukin (IL)-17 is a proinflammatory cytokine
secreted by Th17 cells (12) that
promotes the secretion of IL-1β, tumor necrosis factor (TNF)-α and
IL-6 by binding to their corresponding receptors mainly expressed
in the epithelial and endothelial cells of the kidney and spleen
(13-15).
Previous studies have found that the IL-23/IL-17 pathway
contributes significantly to renal tissue injury in experimental
glomerulonephritis (16) and
patients with PNS (17), where the
inhibition of IL-17 expression in patients with renal disease
attenuated crescent formation in nephrotoxic serum nephritis
(18). However, it currently remains
unclear whether IL-17 can influence the development of PNS by
regulating podocyte injury.
In the present study, the relationship between IL-17
and PNS, specifically that between IL-17 and inflammatory podocyte
injury, were investigated. It was found that IL-17 was highly
expressed in renal tissues of patients with PNS and was associated
with indicators of podocyte injury. In addition, IL-17 induced
podocyte apoptosis by activating the Fas/Fas ligand
(FasL)/caspase-8/caspase-3 apoptotic pathway via NF-κB activation
in vitro. In conclusion, data from the present study
provided a potential target for attenuating podocyte injury in
children with PNS by suppressing inflammation.
Materials and methods
Tissues and ethics statement
Renal biopsies of 9 patients with minimal change
nephrotic syndrome (MCNS) (3 female and 6 males; mean age, 8.6±2.3
years), 15 patients with mesangial proliferative glomerulonephritis
(MsPGN) (6 females and 9 males; mean age, 8.2±2.4 years) and 9
patients with focal segmental glomerulosclerosis (FSGS) (3 females
and 6 males; mean age, 8.4±2.8 years) were collected at The First
Hospital of Jilin University (Changchun, China) from January 2015
to December 2017. In addition, normal kidney tissues were collected
from 15 patients (6 females and 9 males, age range, 3-10 years,
mean age, 8.5±3.0 years) at The First Hospital of Jilin University
(Jilin, China) during early renal tumor surgery; normal routine
urine test results and renal function were observed. The inclusion
criteria used were as follows: i) All PNS patients were diagnosed
according to the diagnostic criteria for glomerular nephropathy in
China (19); ii) patients with PNS
were <10 years of age and did not receive any medication prior
to renal puncture; iii) the patients with PNS has complete clinical
data, including age and sex data. The exclusion criteria were as
follows: i) Other organ tissue dysfunction; ii) Infected with HIV,
HCV or tuberculosis; iii) had received surgical treatment within
half a year; iv) exhibited epilepsy, congenital heart disease or
any other genetic diseases. All relevant guardians or patients of
the participants in the present study provided signed informed
consent and the experiments were approved by the Ethics Association
of The First Hospital of Jilin University (Changchun, China). No
animals were used in the present study.
Cell lines and drugs
Podocytes (20,21),
from H-2Kb-tsA58 transgenic mice, were a gift from Professor Peter
Mundel (Mount Sinai School of Medicine, New York University, USA)
and were cultured at 37˚C with 5% CO2 in DMEM (cat. no.
12491-15; Thermo Fisher Scientific, Inc.) supplemented with 10%
fetal bovine serum (cat. no. 10100-147; Thermo Fisher Scientific,
Inc.) and 1% penicillin/streptomycin (cat. no. 1564005; Thermo
Fisher Scientific, Inc.).
A total of 1.5x106 cells/well of
podocytes were seeded into six-well plates and were treated with 0,
50, 100 and 200 ng/ml recombinant mouse IL-17 (rmIL-17; cat. no.
PMC0175; Thermo Fisher Scientific, Inc.) for 24, 48 or 72 h. A
total of 5 µM helenalin (cat. no. 6754-13-8; Cayman Chemical
Company) was added to podocytes with 200 ng/ml rmIL-17 for 72 h.
Cell culture supernatant was collected using centrifugation (2,000
x g; 10 min; 4˚C), and the concentration of IL-1β in the cell
culture supernatant was subsequently measured using the IL-1β mouse
uncoated ELISA kit (cat. no. 88-7013-88; Thermo Fisher Scientific,
Inc.) and TNF-α was measured using the TNF-α mouse ELISA kit (cat.
no. BMS607-3; Thermo Fisher Scientific, Inc.).
Reverse transcription-quantitative PCR
(RT-qPCR)
TRIzol® (cat. no. 15596018; Thermo Fisher
Scientific, Inc.) was used to extract total RNA from kidney tissues
or cells. The extracted RNA was then reverse transcribed into cDNA
by using PrimeScript™ RT Master Mix (cat. no. RR036B;
Takara Biotechnology Co., Ltd.), with the temperature protocol of
37˚C for 60 min and 85˚C for 5 sec. Subsequently, 20 µl
fluorescence quantitative PCR (qPCR) reactions were prepared
according to the manufacturer's protocol of the
SYBR®-Green qPCR Master Mix (cat. no. 638320; Takara
Biotechnology Co., Ltd.) and amplified using ABI 7500 Real-Time PCR
System (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
following thermocycling conditions were used: Initial denaturation
at 95˚C for 30 sec, followed by 40 cycles of 90˚C for 5 sec and
65˚C for 30 sec. Sequences of all primers used for RT-qPCR analysis
are provided in Table I. We used the
2-ΔΔCq method (22) to calculate the relative expression
levels of genes, and GAPDH was used for normalization.
 | Table ISequences of primers used for reverse
transcription-quantitative PCR. |
Table I
Sequences of primers used for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5'-3') |
|---|
| IL-17 | F: |
TCCCACGAAATCCAGGATGC |
| R: |
GGATGTTCAGGTTGACCATCAC |
| Podocalyxin
(human) | F: |
TCCCAGAATGCAACCCAGAC |
| R: |
GGTGAGTCACTGGATACACCAA |
| Podocalyxin
(murine) | F: |
AGTGCCACAACATCAACAGAA |
| R: |
TGTGAATGGTGTAGGGTTGCT |
| WT1 | F: |
AGCACGGTCACTTTCGACG |
| R: |
GTTTGAAGGAATGGTTGGGGAA |
| Nephrin | F: |
ATGGGAGCTAAGGAAGCCACA |
| R: |
CCACACCACAGCTTAACTGTC |
| Synaptopodin | F: |
TCCTCACCTAATGCCACACTC |
| R: |
GCTGGAGGGTTTTGGTTGATA |
| GAPDH (human) | F: |
ACAACTTTGGTATCGTGGAAGG |
| R: |
GCCATCACGCCACAGTTTC |
| GAPDH (murine) | F: |
AGGTCGGTGTGAACGGATTTG |
| R: |
GGGGTCGTTGATGGCAACA |
Western blotting
Tissues and cells were submerged in RIPA lysate
(cat. no. P0013C; Beyotime Institute of Biotechnology), and
detected protein concentration using a BCA kit (cat. no. P0012;
Beyotime Institute of Biotechnology), and a total of 40 µg protein
were separated by 5% SDS-PAGE and then transferred to PVDF
membranes. BSA (5%) which was 5% BSA buffer solution (cat. no.
ST-023; Beyotime Institute of Biotechnology) in PBS as a blocking
solution to block for 1 h at room temperature, and then washed 3
times with PBS buffer. Membranes were subsequently incubated with
primary antibodies against IL-17 (cat. no. sc-374218; 1:500; Santa
Cruz Biotechnology, Inc.), Fas (cat. no. ab82419; 1:1,000; Abcam),
FasL (cat. no. ab15285; 1:1,000; Abcam), active caspase-8 (cat. no.
ab227430; 1:2,000; Abcam), active caspase-3 (cat. no. ab49822;
1:500; Abcam), p65 phosphorylated (p-) S636 (cat. no. ab86299;
1:5,000) or GAPDH (cat. no. ab9484; 1:3,000) overnight at 4˚C,
followed by incubation with Goat Anti-Rabbit IgG H&L (HRP)
(cat. no. ab6721; 1:1,000) or Goat Anti-Mouse IgG H&L (HRP)
(cat. no. ab205719; 1:1,000) secondary antibodies for 1 h at room
temperature, and then ECL solution (cat. no. WBKLS0100, Beijing
Xinjingke Biotechnologies Co., Ltd.) was added for detection.
Protein expression levels were analyzed using ImageJ v1.8.0
(National Institutes of Health), with the calculated gray scale
values of target proteins/GAPDH used to quantify relative
expression.
Immunohistochemistry
IL-17 protein expression was detected by
immunohistochemical staining according to the protocol of the
VECTASTAIN® Elite® ABC Kit (Vector
Laboratories, Inc.; Maravai LifeSciences) in the kidney tissue
sections. The 5 µm paraffin sections were incubated at 60˚C for 2 h
after soaking in 3% hydrogen peroxide solution, dewaxed and
hydrated using xylene and an ethanol gradient, following which
sections were washed with PBS and double distilled water. 5% BSA
which was 5% BSA buffer solution in PBS as a blocking solution to
block for 1 h at room temperature. The sections were subsequently
stained using an anti-IL-17 primary antibody (1:100; cat. no.
sc-374218; Santa Cruz Biotechnologies, Inc.) or PBS (negative
control) overnight at 4˚C, followed by incubation with horseradish
peroxidase-conjugated goat anti-rabbit IgG H&L secondary
antibodies (cat. no. ab6721; 1:500; Abcam) at 37˚C for 2 h.
Finally, DAB staining (cat. no. P0203; Beyotime Institute of
Biotechnology) for 10 min at room temperature and hematoxylin
staining at room temperature (cat. no. C0107; Beyotime Institute of
Biotechnology) for 2 min, and images from 5 fields of view were
obtained per slice. The sections were analyzed using Leica TCS SP5
microscope 217 (magnification, x200; Leica Microsystems, Ltd.) with
LAS AF Lite 4.0 image browser software (Leica Microsystems, Ltd.).
The cells were either stained yellow (1 point), brown (2 points) or
tan (3 points) for positive staining. In addition, ≤25% positive
cells in each field of view was scored as 0 point, 25-75% was
scored as 1 point and ≥75% was scored as 2 points. Final
immunohistochemical score=Score (staining intensity) x Score (% of
IL-17 positive cells) (23).
Urine podocyte counting
Following admission, each patient provided 50 ml
clean mid-stream morning urine 2 days prior to renal puncture.
Urine samples were subsequently centrifuged at 1,000 x g for 10 min
at room temperature to harvest the urine sediments. A total of 50
µl urine sediment which was dissolved in physiological saline
solution was added into an auto-smear centrifuge (TXD3; Changsha
Xiangyi Centrifuge Instrument Co., Ltd.) to produce cell slides by
centrifugation (350 x g, 3 min, room temperature). The slides were
then dried at room temperature and fixed with 99.5% acetone for 4
min at room temperature before being and air-dried.
Slides with cells were incubated in culture plates
and washed with PBS (3x3 min), followed by fixation with 4%
paraformaldehyde for 15 min at room temperature and
permeabilization with 0.5% Triton X-100 dissolved in PBS. Normal 5%
goat serum (cat. no. C0265; Beyotime Institute of Biotechnology)
was then added onto the slides for blocking at room temperature for
30 min before Anti-PODXL antibody [EPR9518] (cat. no. ab150358;
1:1,000; Abcam) was added into slides and incubated in a wet box
overnight at 4˚C. The next day, the slides were rinsed three times
with PBS supplemented with 0.05% Tween-20 (PBS-T), following which
Goat Anti-Rabbit IgG H&L (Alexa Fluor® 488; cat. no.
ab150077; 1:1,000; Abcam) was added and the slides were incubated
in a wet box at 20-37˚C for 1 h. A concentration of 5 µg/ml DAPI
(cat. no. 4083; Cell Signaling Technology, Inc.) was subsequently
added before the slides were incubated in the dark for 5 min at
room temperature to stain the nuclei of the specimen. Following the
removal of excess DAPI by rinsing with PBS-T (4x5 min), the slides
were dried using absorbent paper and an anti-fluorescence quenching
sealant (cat. no. HCY078; Hangzhou Yuxin Biotechnology Co., Ltd.)
was used. A fluorescence microscope was then used to acquire
images. Counting the average of urinary podocytes were counted from
10 high-powered (magnification, x200) fields of view.
Flow cytometric analysis of
apoptosis
Total of 2-3x106 cells/well of podocytes
were collected following treatment for 24, 48 and 72 h with 0, 50,
100 and 200 ng/ml IL-7 and the Annexin V-FITC/propidium iodide kit
(cat. no. V13241; Invitrogen; Thermo Fisher Scientific, Inc.) was
used to detect apoptosis according to manufacturer's protocol.
Beckman CytoFLEX Flow cytometer (Beckman Coulter, Inc.) with Flowjo
10.0 software (Beckman Coulter, Inc.) was used to analyze podocyte
apoptosis.
Statistical analysis
Data are presented as the mean ± standard deviation
from three independent repetitions per experiment and were analyzed
using SPSS 20.0 (IBM Corp.). Unpaired Student's t-test was used to
compare differences between two groups, whilst comparison between
multiple groups was performed using the One-way ANOVA followed by
Duncan's tests. Correlation between two groups was analyzed by
Pearson's correlation analysis. P<0.05 was considered to
indicate a statistically significant difference.
Results
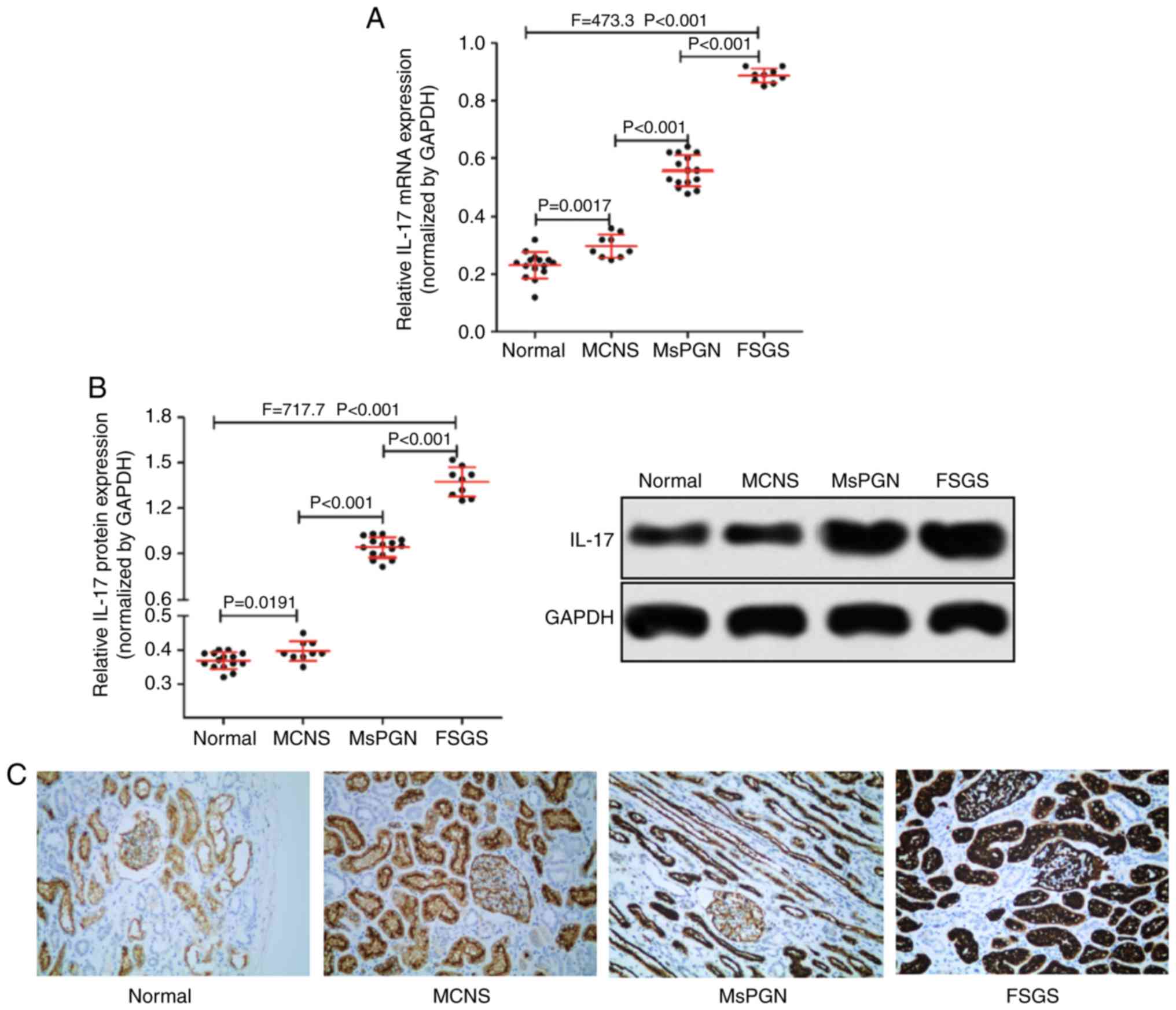
IL-17 is highly expressed in patients
with PNS
RT-qPCR was used to measure the expression of IL-17
mRNA, whereas western blotting and immunohistochemistry were
performed for IL-17 protein expression in patient tissue samples.
The expression of IL-17 in renal tissues of patients with PNS was
found to be significantly higher compared with that of normal
kidney tissues (Fig. 1A and B). In addition, the expression level of
IL-17 also appeared to associate with disease severity in patients
with PNS (Fig. 1C), as IL-17
expression was found to be highest in renal tissues of patients
with FSGS (n=9) and lowest in MCNS (n=9). Taken together, IL-17 was
highly expressed in the renal tissues of patients with PNS and was
associated with the severity of PNS in patients.
IL-17 is associated with podocyte
injury in patients with PNS
Podocalyxin (PCX) is a podocyte-specific marker
protein (24). Podocytes in the
urine are derived from podocytes shed from kidney tissue (25) and PCX levels can be related to
podocyte injury. The expression of PCX mRNA in kidney tissue of
patients with PNS was measured using RT-qPCR and the number of
podocytes (MCNS was 1.67±0.38, MsPGN was 7.06±0.72, FSGS was
10.44±0.69) was counted using immunofluorescence detection of PCX
proteins (Fig. 2A and C). It was found that there was a negative
correlation between IL-17 and PCX mRNA expression in the renal
tissue (Fig. 2B), whilst a positive
correlation was found between IL-17 mRNA expression and the number
of urinary podocytes in patients with PNS (Fig. 2D). The results suggest that the
expression of IL-17 was correlated with podocyte injury in patients
with PNS.
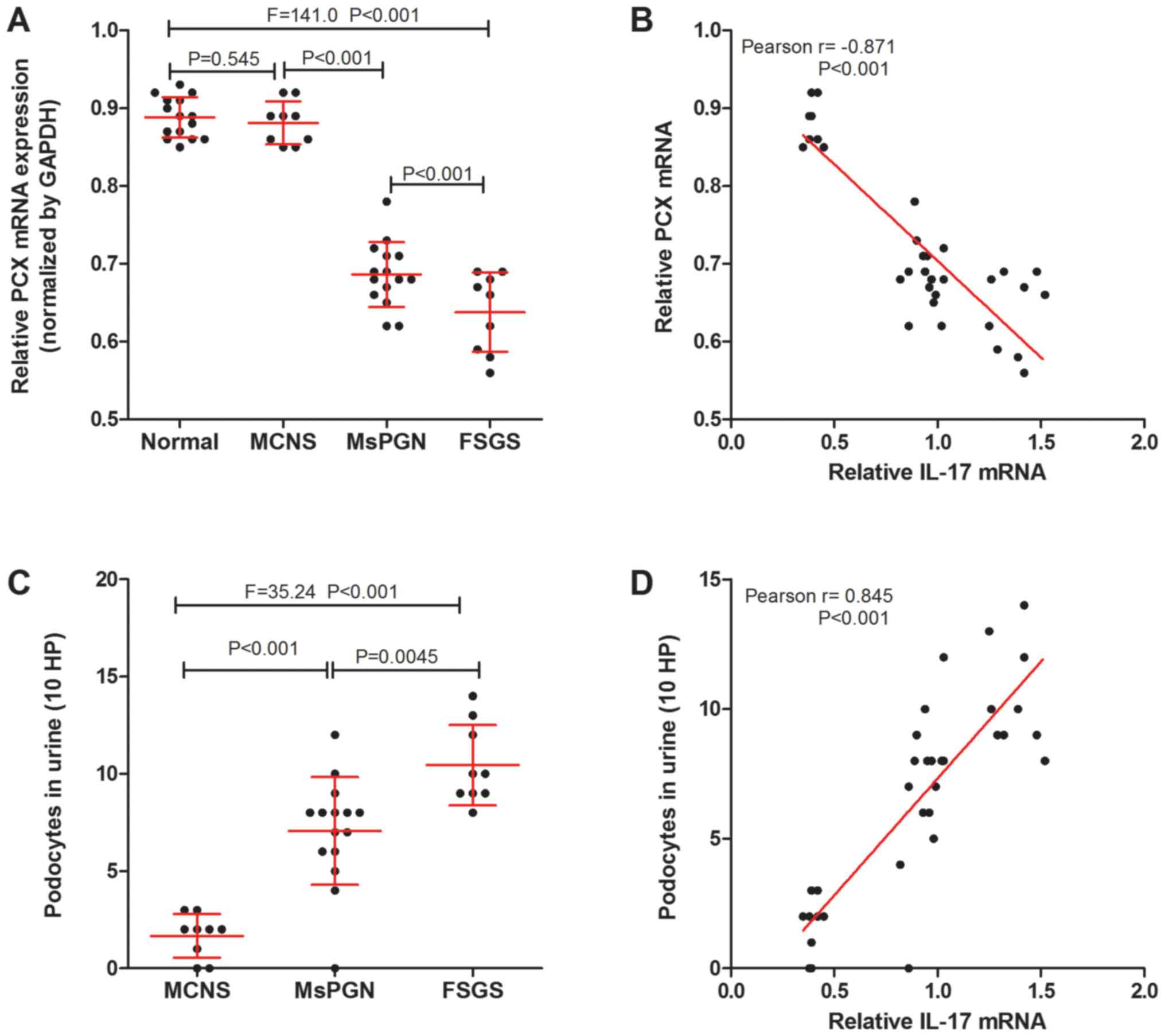 | Figure 2IL-17 is associated with podocyte
injury markers in patients with PNS. (A) Reverse
transcription-quantitative PCR was performed to measure the
expression of PCX mRNA in the kidney tissues of normal patients
without PNS (n=15), and of patients with MCNS (n=9), MsPGN (n=15)
and FSGS (n=9). (B) IL-17 mRNA was negatively correlated with PCX
mRNA expression in the kidney tissues of PNS patients (n=33). (C)
The number of urinary podocytes in patients with MCNS (n=9), MsPGN
(n=15) and FSGS (n=9). (D) A positive correlation was observed
between the levels of IL-17 mRNA in renal tissues and the number of
urinary podocytes in patients with PNS (n=33). IL-17,
interleukin-17; PNS, primary nephrotic syndrome; MCNS, minimal
change nephrotic syndrome; MsPGN, mesangial proliferative
glomerulonephritis; FSGS, focal segmental glomerulosclerosis; PCX,
podocalyxin; HP, high power fields. |
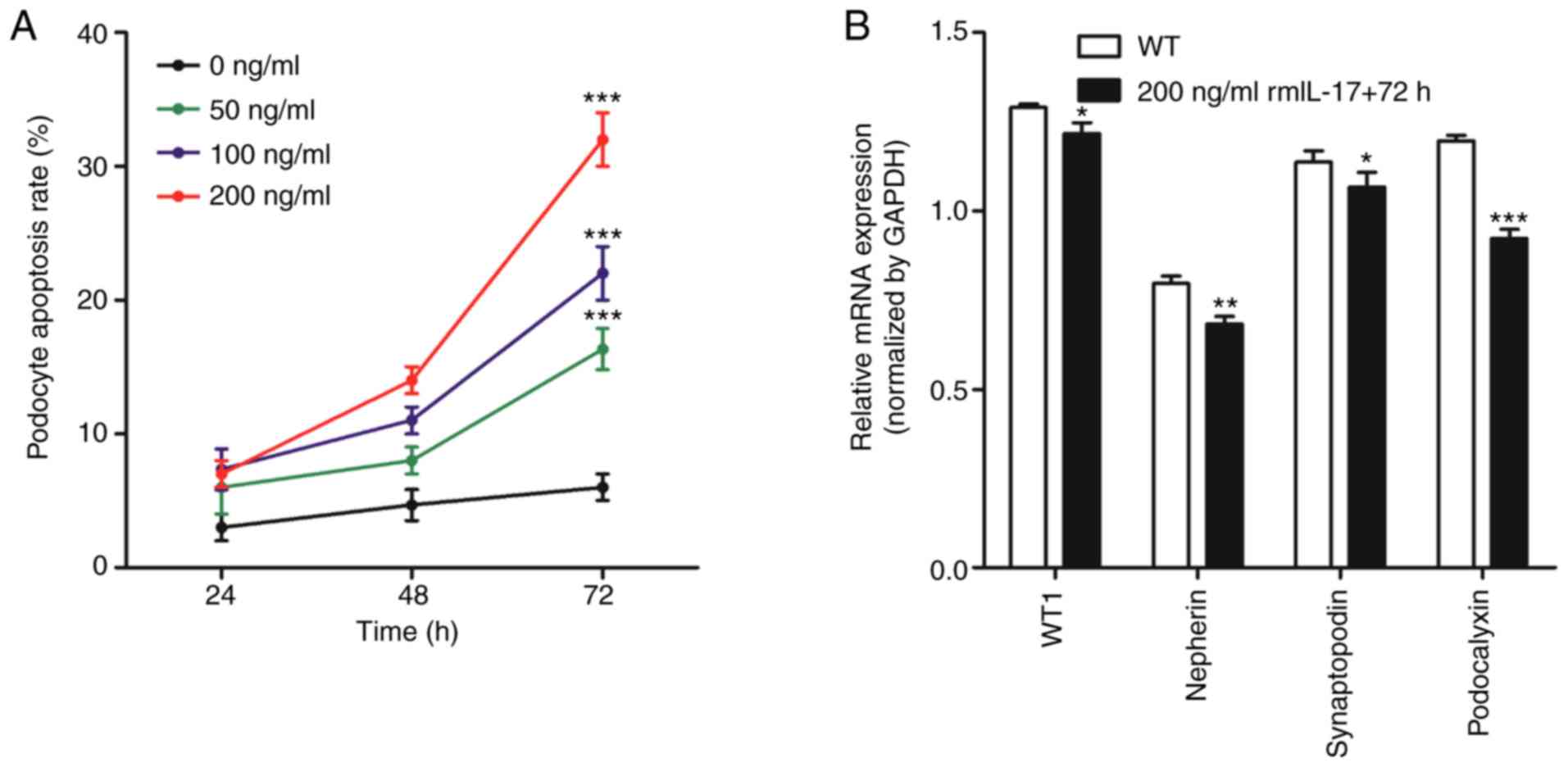
IL-17 induces podocyte apoptosis in
vitro
Flow cytometry was used to measure apoptosis in
podocytes following the addition of ascending concentrations (0,
50, 100 and 200 ng/ml) of rmIL-17 to the culture medium. After 24,
48 and 72 h treatment, podocytes were collected for analysis of
apoptosis and gene expression. For each dose of rmIL-17, the
apoptotic rate of podocytes increased with increasing culture time
(Fig. 3A). Furthermore, at each time
point the apoptotic rate of podocytes increased as the rmIL-17 dose
increased (Fig. 3A). These
observations suggested that IL-17 induced podocyte apoptosis in
vitro in a time- and dose-dependent manner.
According to the experimental results obtained in
Fig. 3A, a dose of 200 ng/ml rmIL-17
for a treatment of 72 h was chosen for subsequent experiments. The
mRNA expression of podocyte marker molecules, WT1, nephrin,
synaptopodin and podocalyxin were significantly reduced following
IL-17 treatment (Fig. 3B). In
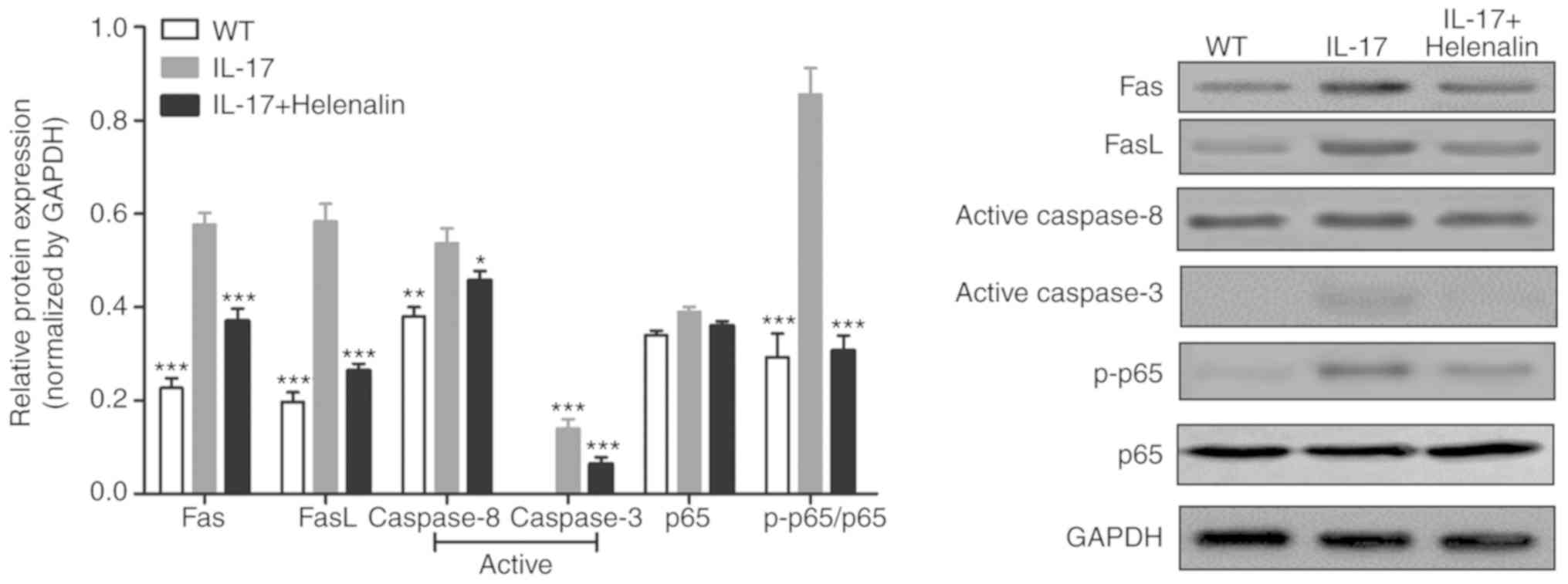
addition, IL-17 treatment increased the expression of Fas, FasL,
active-caspase-8 and active-caspase-3 proteins (Fig. 4). These results suggested that IL-17
induced podocyte apoptosis by activating the
Fas/FasL/caspase-8/caspase-3 signaling pathway in podocytes in
vitro.
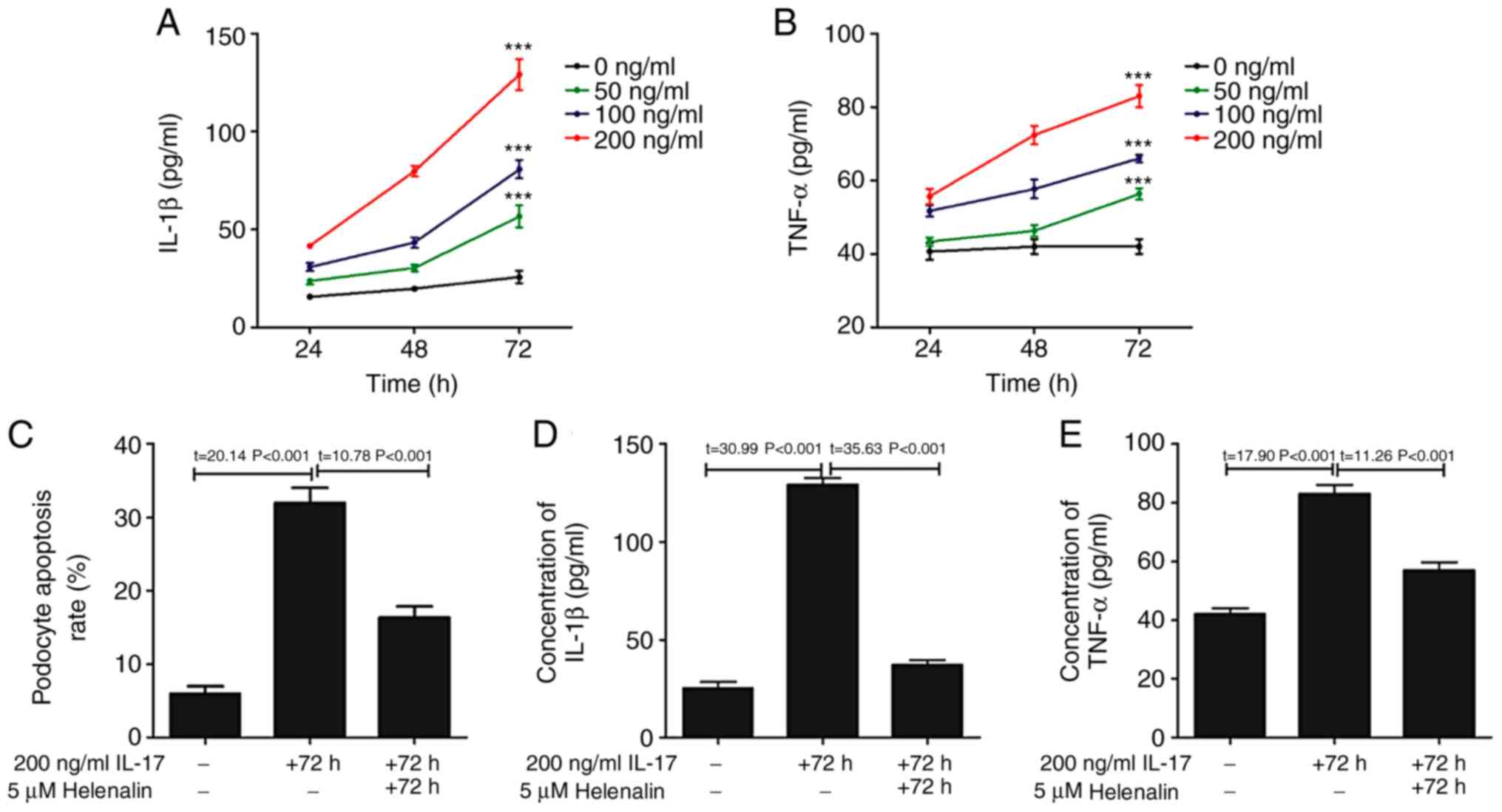
IL-17 activates the NF-κ signaling
pathway in podocytes in vitro
The phosphorylation of p65 subunit of NF-κB was
increased following IL-17 treatment (Fig. 4), suggesting that the NF-κB pathway
was activated (26). Since this
signaling pathway controls the transcription of a plethora of genes
associated with inflammation (27),
the hypothesis that IL-17 induced podocyte apoptosis by
upregulating inflammation was first tested, by measuring the levels
IL-1β and TNF-α content in the podocyte culture supernatant
following IL-17 treatment. IL-17 treatment increased the secretion
of IL-1β and TNF-α in a time- and dose-dependent manner (Fig. 5A and B). To investigate the relationship between
IL-17-induced podocyte apoptosis, IL-1β and TNF-α secretion and the
NF-κB pathway, podocytes were treated with helenalin, a NF-κB
inhibitor, in the presence of IL-17. Helenalin significantly
attenuated IL-17-induced podocyte apoptosis (Fig. 5C), and IL-1β (Fig. 5D) and TNF-α (Fig. 5E) secretion by podocytes.
Additionally, helenalin also significantly inhibited
IL-17-activated NF-κB signaling via p65 phosphorylation and the
Fas/FasL/caspase-8/caspase-3 apoptotic pathway (Fig. 4). These results suggested that
IL-17-induced podocyte apoptosis may be associated with the NF-κB
signaling pathway.
Discussion
Over the past two decades, studies have found that
among children with PNS, the incidence of FSGS has exhibited an
increasing trend, seriously affecting the prognosis of patients
with PNS (28,29). However, development of treatment
strategies for PNS is hindered by the poorly understood mechanism
of glomerular sclerosis pathogenesis. It has been previously
reported that inflammation mediated by CD4+ T cell
dysfunction is associated with the occurrence of glomerular
sclerosis (30,31). Traditionally, CD4+ T cells
are divided into Th1 and Th2 subtypes. Th1 cells are mainly
associated with the secretion of cytokines IL-2 and IFN-γ, which
mediate cellular immune responses, whereas Th2 mainly secrete
cytokines IL-4 and IL-10, which mediate fluid immune responses
(32,33). Although some studies have
demonstrated that the dysfunction of Th1/Th2 and the dominant
activation of Th2 serve an important role in the development of
kidney disease (34,35); however, other studies have opposing
views (36,37). Therefore, the balance of Th1/Th2
activation alone is not sufficient to fully explain the mechanism
of glomerular sclerosis pathogenesis.
In recent years, Th17 cells were discovered as
additional CD4+ T cells, which have a different
mechanism of differentiation, but phenotypes and functions are
derived from Th1/Th2(38). IL-17 is
an important cytokine secreted by Th17 cells, which has been found
to associate closely with the development of coronary heart disease
(39,40), multiple sclerosis (41), inflammation and autoimmune diseases
(15). In kidney diseases, Matsumoto
et al (42) found that the
excretion of IL-17 in urine during the minimally active period was
significantly higher compared with that in the remission period,
which was in turn proportional to the excretion of urinary protein.
In addition, IL-17 was also found to be associated with renal
tissue injury in experimental glomerulonephritis (16), patients with PNS (17) and the severity of IgA nephropathy
(43). In the present study, it was
found that the expression of IL-17 in the renal tissues of patients
with PNS was significantly higher compared with that of normal
kidney tissues, which was also associated with the severity of
disease in patients with PNS. In addition, it was found that IL-17
expression was associated with indicators of podocyte injury.
Podocytes damage is a signature of nephrotic
syndrome (44,45). Podocytes are glomerular visceral
epithelial cells, which belong to a class of terminally
differentiated cells. When podocytes are damaged, the normal
structure of the foot processes is destroyed, resulting in
podocytes detaching from the basement membrane. Since this cannot
be repaired effectively due to the limited proliferative ability of
podocytes (44), the integrity of
the glomerular filtration membrane is compromised, leading to
proteinuria. In the present study, it was found in vitro
that IL-17 treatment induced podocyte apoptosis in addition to
reducing the mRNA expression of podocyte-specific markers,
including WT1, nephrin, synaptopodin and podocalyxin. Indeed,
expression of these markers were directly associated with podocyte
integrity, where studies have found that the loss or mutation of
podocalyxin leads to the development of FSGS and familial nephrotic
syndrome (46,47). Additionally, Fluvastatin treatment,
which is mainly used to treat primary hypercholesterolemia and
primary mixed dyslipidemia, protects podocytes in HIV-associated
nephropathy by increasing the expression of nephrin, WT1 and
synaptopodin (48,49).
To explore the molecular mechanism by which IL-17
induces podocyte apoptosis further, changes in protein expression
in cultured podocytes following IL-17 treatment were examined.
IL-17 treatment increased the expression of Fas, FasL,
active-caspase-8, active-caspase-3 in addition to p65
phosphorylation, all of which were reversed by concomitant
treatment with the NF-κB inhibitor helenalin. A previous study has
shown that inflammatory factors can induce muscle cell apoptosis by
activating the Fas/FasL-caspase-8-caspase-3 pathway (50). NF-κB signaling controls the
transcription of a variety of genes associated with inflammation,
including IL-1β and TNF-α, which in turn can potentiate NF-κB
signaling further (51). Therefore,
it is possible that IL-17 first increased the secretion of IL-1β
and TNF-α by activating the NF-κB pathway in podocytes, where these
inflammatory factors increased podocyte apoptosis of by activating
the Fas/FasL-caspase-8-caspase-3 pathway.
However, it should be emphasized that some
limitations exist for the present study. The podocytes used in the
in vitro study were cell lines and not primary cells.
Secondly, the sample size of the tissues collected from each
disease group were small. In addition, inhibitors for the related
signaling pathways mentioned in the present study were not
used.
In conclusion, results from the present study
suggested that IL-17 was highly expressed in renal tissues of
patients with PNS. The Fas/FasL/caspase-8/caspase-3 apoptotic
pathway was activated in a NF-κB-dependent manner to induce
podocyte apoptosis.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
LZ was responsible for the conception and design of
the study; SZ prepared the manuscript and revised the final draft
of the manuscript; SZ, BS, YZ and LYZ performed the experiments and
analyzed the data.
Ethics approval and consent to
participate
The present study was performed with the approval of
the Ethics Committee of The First Hospital of Jilin University. All
aspects of the study complied with the Declaration of Helsinki. All
relevant guardians or parents of patients provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Trautmann A, Schnaidt S,
Lipska-Ziętkiewicz BS, Bodria M, Ozaltin F, Emma F, Anarat A, Melk
A, Azocar M, Oh J, et al: Long-term outcome of steroid-resistant
nephrotic syndrome in children. J Am Soc Nephrol. 28:3055–3065.
2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Kopp JB: Global glomerulosclerosis in
primary nephrotic syndrome: Including age as a variable to predict
renal outcomes. Kidney Int. 93:1043–1044. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Levey AS, Eckardt KU, Tsukamoto Y, Levin
A, Coresh J, Rossert J, De Zeeuw D, Hostetter TH, Lameire N and
Eknoyan G: Definition and classification of chronic kidney disease:
A position statement from kidney disease: Improving Global Outcomes
(KDIGO). Kidney Int. 67:2089–2100. 2005.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Levey AS, Andreoli SP, Dubose T,
Provenzano R and Collins AJ: Chronic kidney disease: Common,
harmful and treatable-World Kidney Day 2007. Am J Nephrol.
27:108–112. 2007.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Bennett MR: Biomarkers of therapeutic
response in primary nephrotic syndrome: Response. Pediat Nephrol.
28:161–162. 2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Chen W, Jiang Y, Han J, Hu J, He T, Yan T,
Huang N, Zhang Q, Mei H, Liao Y, et al: Atgl deficiency induces
podocyte apoptosis and leads to glomerular filtration barrier
damage. FEBS J. 284:1070–1081. 2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Tian X, Kim JJ, Monkley SM, Gotoh N,
Nandez R, Soda K, Inoue K, Balkin DM, Hassan H, Son SH, et al:
Podocyte-associated talin1 is critical for glomerular filtration
barrier maintenance. J Clin Invest. 124:1098–1113. 2014.PubMed/NCBI View
Article : Google Scholar
|
|
8
|
Petermann A and Floege J: Podocyte damage
resulting in podocyturia: A potential diagnostic marker to assess
glomerular disease activity. Nephron Clin Pract.
106(c61-c66)2007.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Perico L, Conti S, Benigni A and Remuzzi
G: Podocyte-actin dynamics in health and disease. Nat Rev Nephrol.
12(692)2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Nagata M: Podocyte injury and its
consequences. Kidney Int. 89:1221–1230. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Ivanova EA, Arcolino FO, Elmonem MA,
Rastaldi MP, Giardino L, Cornelissen EM, Van Den Heuvel LP and
Levtchenko EN: Cystinosin deficiency causes podocyte damage and
loss associated with increased cell motility. Kidney Int.
89:1037–1048. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Korn T, Bettelli E, Oukka M and Kuchroo
VK: IL-17 and Th17 cells. Annu Rev Immunol. 8:485–517.
2009.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Beringer A, Noack M and Miossec P: IL-17
in chronic inflammation: From discovery to targeting. Trends Mol
Med. 22:230–241. 2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ivanov II, Mckenzie BS, Zhou L, Tadokoro
CE, Lepelley A, Lafaille JJ, Cua DJ and Littman DR: The orphan
nuclear receptor RORgammat directs the differentiation program of
proinflammatory IL-17+ T helper cells. Cell.
126:1121–1133. 2006.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Kuwabara T, Ishikawa F, Kondo M and
Kakiuchi T: The role of IL-17 and related cytokines in inflammatory
autoimmune diseases. Mediators Inflamm.
2017(3908061)2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Paust HJ, Turner JE, Steinmetz OM, Peters
A, Heymann F, Hölscher C, Wolf G, Kurts C, Mittrücker HW, Stahl RA
and Panzer U: The IL-23/Th17 axis contributes to renal injury in
experimental glomerulonephritis. J Am Soc Nephrol.
20(969)2009.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhang L, Yan J, Yang B, Zhang G, Wang M,
Dong S, Liu W, Yang H and Li Q: IL-23 actived γδ T cells affect
Th17 cells and regulatory T cells by secreting IL-21 in children
with primary nephrotic syndrome. Scand J Immunol.
87(36)2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Okada H, Inoue T, Hashimoto K, Suzuki H
and Matsushita S: D1-like receptor antagonist inhibits IL-17
expression and attenuates crescent formation in nephrotoxic serum
nephritis. Am J Nephrol. 30:274–279. 2009.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Jennette JC and Falk RJ: Diagnosis and
management of glomerular diseases. Med Clin North Am. 81:653–677.
1997.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ji Z, Hu Z and Xu Y: APPL1 acts as a
protective factor against podocytes injury in high glucose
environment. Int J Clin Exp Pathol. 8:6764–6771. 2015.PubMed/NCBI
|
|
21
|
Rops AL, van der Vlag J, Jacobs CW,
Dijkman HB, Lensen JF, Wijnhoven TJ, van den Heuvel LP, van
Kuppevelt TH and Berden JH: Isolation and characterization of
conditionally immortalized mouse glomerular endothelial cell lines.
Kidney Int. 66:2193–2201. 2004.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zhang YM, You LF, Chen J and Mao CP:
Expression of kinesin family member 3B is associated with poor
prognosis in epithelial ovarian cancer patients. Int J Clin Exp
Pathol. 10:2834–2842. 2017.
|
|
24
|
Chen WQ, Zhang Y, Jiang H, Li H, Li XY,
Yang X, Feng S, Wang YC, Lin C, Shen XJ, et al: Podocyte-related
proteins in membranous nephropathy progression. Chinese J Med
(Engl). 126:3782–3783. 2013.PubMed/NCBI
|
|
25
|
Yu D, Petermann A, Kunter U, Rong S,
Shankland SJ and Floege J: Urinary podocyte loss is a more specific
marker of ongoing glomerular damage than proteinuria. J Am Soc
Nephrol. 16:1733–1741. 2005.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Buss H, Dörrie A, Schmitz ML, Hoffmann E,
Resch K and Kracht M: Constitutive and interleukin-1-inducible
phosphorylation of p65 NF-{kappa}B at serine 536 is mediated by
multiple protein kinases including I{kappa}B kinase (IKK)-{alpha},
IKK{beta}, IKK{epsilon}, TRAF family member-associated
(TANK)-binding kinase 1 (TBK1), and an unknown kinase and couples
p65 to TATA-binding protein-associated factor II31-mediated
interleukin-8 transcription. J Biol Chem. 279:55633–55643.
2004.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Monaco C, Andreakos E, Kiriakidis S, Mauri
C, Bicknell C, Foxwell B, Cheshire N, Paleolog E and Feldmann M:
Canonical pathway of nuclear factor κB activation selectively
regulates proinflammatory and prothrombotic responses in human
atherosclerosis. Proc Natl Acad Sci USA. 101:5634–5639.
2004.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Yin XL, Zou MS, Zhang Y, Wang J, Liu TL,
Tang JH, Qiu LR, Chen Y, Yuan HQ and Zhou JH: Twenty-three-year
review of disease patterns from renal biopsies: An experience from
a pediatric renal center. J Nephrol. 26:699–707. 2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Yuen LK, Lai WM, Lau SC, Tong PC, Tse KC
and Chiu MC: Ten-year review of disease pattern from percutaneous
renal biopsy: An experience from a paediatric tertiary renal centre
in Hong Kong. Hong Kong Med J. 14:348–355. 2008.PubMed/NCBI
|
|
30
|
Umemoto S, Okamoto T, Yoshimura K,
Sakumura T, Murata T, Fukai T, Yano M and Matsuzaki M: Toll-like
receptor 4 function is essential for increasing oxidative stress,
and promoting the inflammation and glomerular sclerosis by
angiotensin II. J Am Coll Cardiol. 67:2243. 2016. View Article : Google Scholar
|
|
31
|
Reggiani F and Ponticelli C: Focal
segmental glomerular sclerosis: Do not overlook the role of immune
response. J Nephrol. 29:525–534. 2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Cousins DJ, Lee TH and Staynov DZ:
Cytokine coexpression during human Th1/Th2 cell differentiation:
Direct evidence for coordinated expression of Th2 cytokines. J
Immunol. 169:2498–2506. 2002.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Egwuagu CE, Yu CR, Zhang M, Mahdi RM, Kim
SJ and Gery I: Suppressors of cytokine signaling proteins are
differentially expressed in Th1 and Th2 cells: Implications for Th
cell lineage commitment and maintenance. J Immunol. 168:3181–3187.
2002.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Essa S, Pacsa A, Raghupathy R, Said T,
Nampoory MRN, Johny KV and Alnakib W: Low levels of Th1-type
cytokines and increased levels of Th2-type cytokines in kidney
transplant recipients with active cytomegalovirus infection.
Transplant Proc. 41:1643–1647. 2009.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Hwang YJ, Yun MO, Jeong KT and Park JH:
Uremic toxin indoxyl 3-sulfate regulates the differentiation of Th2
but not of Th1 cells to lessen allergic asthma. Toxicol Lett.
225:130–138. 2014.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Lama G, Luongo I, Tirino G, Borriello A,
Carangio C and Salsano ME: T-lymphocyte populations and cytokines
in childhood nephrotic syndrome. Am J Kidney Dis. 39:958–965.
2002.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Kaneko K, Tuchiya K, Fujinaga S, Kawamura
R, Ohtomo Y, Shimizu T and Yamashiro Y: Th1/Th2 balance in
childhood idiopathic nephrotic syndrome. Clin Nephrol. 58:393–397.
2002.PubMed/NCBI View
Article : Google Scholar
|
|
38
|
Weaver CT, Harrington LE, Mangan PR,
Gavrieli M and Murphy KM: Th17: An effector CD4 T cell lineage with
regulatory T cell ties. Immunity. 24:677–688. 2006.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Gong F, Liu Z, Liu J, Zhou P, Liu Y and Lu
X: The paradoxical role of IL-17 in atherosclerosis. Cell Immunol.
297:33–39. 2015.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Potekhina AV, Pylaeva E, Provatorov S,
Ruleva N, Masenko V, Noeva E, Krasnikova T and Arefieva T:
Treg/Th17 balance in stable CAD patients with different stages of
coronary atherosclerosis. Atherosclerosis. 238:17–21.
2015.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Babaloo Z, Aliparasti MR, Babaiea F,
Almasi S, Baradaran B and Farhoudi M: The role of Th17 cells in
patients with relapsing-remitting multiple sclerosis:
Interleukin-17A and interleukin-17F serum levels. Immunol Lett.
164:76–80. 2015.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Matsumoto K and Kanmatsuse K: Increased
urinary excretion of interleukin-17 in nephrotic patients. Nephron.
91:243–249. 2002.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Matsumoto K and Kanmatsuse K:
Interleukin-17 stimulates the release of pro-inflammatory cytokines
by blood monocytes in patients with IgA nephropathy. Scand J Urol
Nephrol. 37:164–171. 2003.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Niranjan T, Bielesz B, Gruenwald A, Ponda
MP, Kopp JB, Thomas DB and Susztak K: The Notch pathway in
podocytes plays a role in the development of glomerular disease.
Nat Med. 14(290)2008.PubMed/NCBI View
Article : Google Scholar
|
|
45
|
Reiser J, von Gersdorff G, Loos M, Oh J,
Asanuma K, Giardino L, Rastaldi MP, Calvaresi N, Watanabe H,
Schwarz K, et al: Induction of B7-1 in podocytes is associated with
nephrotic syndrome. J Clin Invest. 113:1390–1397. 2004.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Kavoura E, Gakiopoulou H, Paraskevakou H,
Marinaki S, Agrogiannis G, Stofas A, Boletis I, Patsouris E and
Lazaris AC: Immunohistochemical evaluation of podocalyxin
expression in glomerulopathies associated with nephrotic syndrome.
Hum Pathol. 42:227–235. 2011.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Mollet G, Ratelade J, Boyer O, Muda AO,
Morisset L, Lavin TA, Kitzis D, Dallman MJ, Bugeon L, Hubner N, et
al: Podocin inactivation in mature kidneys causes focal segmental
glomerulosclerosis and nephrotic syndrome. J Am Soc Nephrol.
20:2181–2189. 2009.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Sakurai N Kuroiwa T, Ikeuchi H, Hiramatsu
N, Takeuchi S, Tomioka M, Shigehara T, Maeshima A, Kaneko Y and
Hiromura K: Fluvastatin prevents podocyte injury in a murine model
of HIV-associated nephropathy. Nephrol Dial Transplant.
24:2378–2383. 2009.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Wei P, Grimm PR, Settles DC, Balwanz CR,
Padanilam BJ and Sansom SC: Simvastatin reverses podocyte injury
but not mesangial expansion in early stage type 2 diabetes
mellitus. Ren Fail. 31:503–513. 2009.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Kondo M, Murakawa Y, Harashima N,
Kobayashi S, Yamaguchi S and Harada M: Roles of proinflammatory
cytokines and the Fas/Fas ligand interaction in the pathogenesis of
inflammatory myopathies. Immunology. 128 (Suppl
1)(e589-e599)2009.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Sun SC: The non-canonical NF-κB pathway in
immunity and inflammation. Nat Rev Immunol. 17:545–558.
2017.PubMed/NCBI View Article : Google Scholar
|