Introduction
Aging is mainly characterized by a progressive
dysfunction of metabolism and various physiological roles, such as
maintaining metabolic rate and delaying cell aging (1,2).
Clinically, abnormalities of the metabolism and physiological
functions can affect the morbidity and mortality of patients. A
previous study reported that injury- or damage-associated risk
factors are able to cause obvious aging of stem cells (3). Reactive oxygen species (ROS) have been
shown to cause injury or damage to cells (4,5).
Recently, radiotherapy has been extensively applied
for the treatment of cancers. Although radiotherapy inhibits the
proliferation of tumor cells, it also suppresses the growth of
hematopoietic stem cells (6).
Therefore, the discovery of drugs that could effectively prevent
the radiation-induced damage of cells in the hematopoietic system
undergoing radiotherapy would be important. Previous studies have
investigated the anti-oxidant capacity of drugs by exploring the
progression of aging and associated oxidative injuries (7,8);
however, studies on the effects of antioxidants on
radiation-induced hematopoietic hypofunction are lacking.
Ginsenoside Rg1 (Rg1) is the most active ingredient
of Panax ginseng, and is characterized by radiation
resistance, anti-aging and anti-oxidation effects (9). A previous study conducted by the
present research team showed that Rg1 could attenuate the aging
processes of hematopoietic stem cells (HSCs) and hematopoietic
progenitor cells (HPCs) (10). Chen
et al (11) also reported
that Rg1 provided resistance against the radiation-induced aging of
mouse HSCs/HPCs. However, the anti-aging mechanism ofRg1 and the
associated regulatory molecules have not yet been discovered.
Sirtuins (SIRTs) area highly conserved family of
proteins, consisting of seven NAD+-dependent
deacetylases, namely SIRT1-7 (12,13).
Among these deacetylases, SIRT1 and SIRT3, as the primary
deacetylases, participate in cell proliferation, apoptosis, aging
and energy metabolism (14,15). A previous study (16) reported that SIRT3 directly regulates
ROS production in mitochondria, and further affects cell aging
processes. Libert and Guarente (17)
reported that SIRT1 serves a critical function in several molecular
processes, including inflammation, senescence/aging and
intracellular transcription. SIRT1 has also been shown to protect
against cellular senescence by deacetylating forkhead box O3
(FOXO3) transcription factors (18).
Another previous study demonstrated that SIRT3 could regulate ROS
levels by changing the expression of superoxide dismutase 2 (SOD2)
(19). A deficiency of SOD2 has also
been shown to be associated with aging (20).
In the present study, Rg1 was used to treat a
γ-irradiation-induced aging mouse model. In addition, the
antagonistic effects of Rg1 on the radiation-induced aging of stem
cell antigen 1 positive (Sca-1+) HSC/HPCs were also
explored. Furthermore, the regulative role of SIRT1/SIRT3 signaling
pathways in the anti-aging effects of Rg1 on Sca-1+
HSCs/HPCs derived from the γ-ray irradiation aging mouse model was
also investigated.
Materials and methods
Mice
A total of 90 clean C57BL/6 mice (weighing 20-25 g,
6-8 weeks old, random selection of 43 males and 47 females) were
purchased from the Experimental Animal Center of Chongqing Medical
University (Chongqing, China). The mice were housed in an
environment with a 12-hlight/dark cycle, 40% humidity at room
temperature with free access to the food and water. All experiments
were approved by the Ethics Committee of the Key Laboratory of Cell
Biology (Kunming, China).
Animal grouping
The mice were divided into three groups, namely the
control group, γ-ray irradiation group and Rg1 group. The mice in
the control group (n=30) were intraperitoneally injected with
normal saline and not exposed to γ-ray irradiation. The mice in the
γ-ray irradiation group (n=30) were intraperitoneally injected with
normal saline for 7 days, followed by exposure to 6.5-Gy γ-ray
total-body irradiation. The γ-rays were delivered by a linear
accelerator at a dose rate of 57.28 Gy/min (11). The mice in the Rg1 group (n=30) were
subjected to the same processes as the γ-ray irradiation group, but
with the replacement of normal saline by the same volume of Rg1 at
dose of 20 mg/kg/day. The time interval between the final injection
and irradiation was 24 h. The specific processes for the treatment
of each group were conducted according to a previously published
study (11). The Rg1 (cat. no.
060427; purity >95%) was purchased from Jilin Hongjiu Biological
Technology Co., Ltd.
Sample preparation
Prior to irradiation treatment, the mice were
anesthetized by the intraperitoneal injection of 55 mg/kg
pentobarbital. Following the 7 day treatment protocol, mice were
sacrificed via the intraperitoneal injection of 150 mg/kg
pentobarbital (Altana AG) on day 8. The Sca-1+ HSC/HPCs
were harvested using an immunomagnetic separation method as
described in a previous study (21).
The harvested Sca-1+ HSC/HPCs were used in several
experiments, including senescence-associated β-galactosidase
(SA-β-Gal) cytochemical staining, flow cytometry (FCM), mixed
hematopoietic progenitor cell colony-forming unit (CFU-Mix),
reverse transcription-quantitative PCR (RT-qPCR) and western blot
assays.
Blood routine analysis
Mice were sacrificed as described above, and blood
was collected from the eyeball. Blood routine tests, including
white blood cell (WBC), red blood cell (RBC) and blood platelet
(PLT) counts were conducted using an XE-2100 hematology analyzer
(Sysmex Corporation).
SA-β-Gal staining
Sca-1+ HSC/HPCs were collected on day 7
from every group. The HSC/HPCs were stained using an Senescence
β-Galactosidase Staining kit (cat. no. 9860; Cell Signaling
Technology, Inc.) according to the manufacturer's instructions. In
brief, the purified cells (1x105) were washed three
times using PBS for 5 min each time, and fixed using 4%
paraformaldehyde (Beyotime Institute of Biotechnology) for 10 min
at room temperature. Then, the Sca-1+ HSC/HPCs were
stained using staining solution at 37˚C for 12 h. The cells were
centrifuged at 1,000 x g at room temperature for 10 min and
cytospin slides were prepared. Subsequently, the slides were
sheet-sealed using 70% glycerol (Sigma-Aldrich; Merck KGaA). In
order to observe and calculate the percentage of blue-stained
positive cells under bright field illumination, ~1x104
cells were separated on a slide, and 400 cells on each slide were
selected randomly and counted.
CFU-mix culture
The CFU-mix assay was performed according to a
previously published study (22)
with a few modifications. Briefly, the cells (1x104
cells/group) were incubated with DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with horse serum (Gibco; Thermo
Fisher Scientific, Inc.), 2-mercaptoethanol (1x10-4
mol/l; Sigma-Aldrich; Merck KGaA), 3% L-glutamine (Sigma-Aldrich;
Merck KGaA), recombinant human erythropoietin (Kyowa Hakko Kirin
China Pharmaceutical Co., Ltd.), recombinant human granulocyte
macrophage colony stimulating factor (Kyowa Hakko Kirin China
Pharmaceutical Co., Ltd.), interleukin-3 (IL-3; Sigma-Aldrich;
Merck KGaA) and 2.7% methylcellulose. The components were mixed
intensively, seeded into 96-well plates (0.2 ml/well) and then
cultured at 37˚C with 5% CO2 for 7 days. The CFU-Mix and
multiple-differentiation capacities were evaluated according to the
CFU-Mix numbers and percentage of Sca-1+ HSC/HPCs.
FCM assay of the cell cycle
Sca-1+ HSC/HPCs were collected from all
three groups and washed using PBS. Sca-1+ HSC/HPCs were
incubated with 70% iced ethanol overnight, and washed with PBS
three times (5 min/time). The Sca-1+ HSC/HPCs were then
incubated with bovine pancreatic ribonuclease (cat. no. R2638;
Sigma-Aldrich; Merck KGaA; 1 mg/ml; 100 µl) at 37˚C for 30 min.
Then, the cells were stained using propidium iodide (PI; 50 µg/ml)
for 30 min at room temperature in the dark. Finally, the stained
cells were analyzed using FCM with a FACSVantage SE instrument (BD
Biosciences). The proportion of cells in each cell cycle phase was
analyzed using Cell Quest software version 3.3 (BD
Biosciences).
RT-qPCR assay
Total cellular RNAs of Sca-1+ HSC/HPCs
were extracted using TRIzol reagent (cat. no. 15596018; Thermo
Fisher Scientific, Inc.). ABeyoRTIII First Strand cDND Synthesis
kit (cat. no. D7178S; Beyotime Institute of Biotechnology) was used
to synthesize cDNAs from the RNA. All processes of the reverse
transcription were conducted according to the manufacturer's
protocol. The synthesized cDNAs were amplified using SYBR-Green I
as the fluorescent dye. RT-qPCR was performed using a Fast
real-time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The following conditions were used for amplification: 94˚C
for 4 min, 94˚C for 20 sec, 60˚C for 30 sec and 72˚C for 30 sec,
for 35 cycles. The primers for SIRT1, SIRT3, FOXO3, SOD2 and
β-actin are listed in Table I. The
relative mRNA expression of the target gene was normalized to that
of β-actin using the comparative threshold cycle
(2-ΔΔCq) method (23).
 | Table IPrimers used for quantitative
PCR. |
Table I
Primers used for quantitative
PCR.
| Primers | Forward
(5'-3') | Reverse
(5'-3') | Gene length
(bp) | Tm (˚C) |
|---|
| SIRT1 |
AAAGTGATGACGATGACAGAACG |
GCCAATCATGAGATGTTGCTG | 104 | 64 |
| SIRT3 |
GCTTCTGCGGCTCTATACACAG |
CACCCTGTCCGCCATCAC | 169 | 66 |
| FOXO3 |
CAGTCACCCATGCAGACTATCC |
GTCGCTGTGGCTGAGTGAGTC | 117 | 68 |
| SOD2 |
GAGGCTATCAAGCGTGACTTTG |
GCAATGGGTCCTGATTAGAGC | 157 | 67 |
| β-actin |
GAGACCTTCAACACCCCAGC |
ATGTCACGCACGATTTCCC | 263 | 61 |
Western blot assay
Proteins were isolated from Sca-1+
HSC/HPCs by lysing cells with lysis buffer (Beyotime Institute of
Biotechnology). The concentration of isolated proteins was
calculated with a BCA protein quantification kit (Beyotime
Institute of Biotechnology). Equal amounts of protein (0.2 µg) were
separated using 15% SDS-PAGE (Sigma-Aldrich; Thermo Fisher
Scientific, Inc.) and electrotransferred onto PVDF membranes
(Pierce; Thermo Fisher Scientific, Inc.). The PVDF membranes were
blocked using 5% defatted milk at 4˚C overnight. Then, the PVDF
membranes were incubated with rabbit anti-mouse SIRT1 polyclonal
antibody (cat. no. sc-15404, 1:3,000; Santa Cruz Biotechnology,
Inc.), rabbit anti-mouse SIRT3 polyclonal antibody (cat. no.
sc-99143, 1:3,000; Santa Cruz Biotechnology, Inc.), rabbit
anti-mouse FOXO3 polyclonal antibody (cat. no. ab47285, 1:3,000;
Abcam), rabbit anti-mouse SOD2 polyclonal antibody (cat. no.
sc-30080; 1:2,000; Santa Cruz Biotechnology, Inc.) and rabbit
anti-mouse GAPDH polyclonal antibody (cat. no. sc-25778; 1:2,000;
Santa Cruz Biotechnology, Inc.) for 2 h at room temperature. The
PVDF membranes were then washed and incubated with horseradish
peroxidase-conjugated goat anti-rabbit IgG (cat. no. sc-2030;
1:1,000; Santa Cruz Biotechnology, Inc.) at 37˚C for 1 h. Finally,
the western blot bands were visualized using an enhanced
chemiluminescence kit (Pierce; Thermo Fisher Scientific, Inc.). The
western blotting density was qualified using the NIH ImageJ
software version 1.46 (National Institutes of Health).
Statistical analysis
Data are presented as the mean ± standard deviation.
Data were analyzed using SPSS software 11.0 (SPSS, Inc.). All data
were obtained from at least three independent tests or experiments.
Tukey's post hoc test was used following ANOVA to compare
continuous data among multiple groups. P<0.05 was considered to
indicate statistical significance.
Results
Rg1 improves peripheral blood levels
in γ-ray irradiated aging mice
In order to investigate Sca-1+
HSC/HPC-induced hematopoietic functions, the WBC, RBC and PLT
levels in the peripheral blood of the mice were examined. The
results indicated that the WBC, RBC and PLT counts in the γ-ray
irradiation group were significantly decreased compared with those
in the control group (P<0.01; Table
II). Notably, Rg1 treatment significantly increased the WBC,
RBC and PLT counts compared with those in the γ-ray irradiation
group (P<0.05; Table II).
However, the WBC, RBC and PLT counts in the Rg1 group were
significantly lower compared with those in the control group
(P<0.01; Table II).
| Table IIEffects of Rg1 treatment on
peripheral blood (mean ± SD; n=6). |
Table II
Effects of Rg1 treatment on
peripheral blood (mean ± SD; n=6).
| Groups | WBC
(x109/l) | RBC
(x1012/l) | PLT
(x1012/l) |
|---|
| Control | 8.67±0.45 | 11.34±1.02 | 17.08±0.73 |
| γ-ray
irradiation |
1.06±0.54a |
3.87±0.92a |
5.25±0.67a |
| Rg1 |
3.24±1.26a,b |
5.09±0.74a,b |
6.12±1.02a,b |
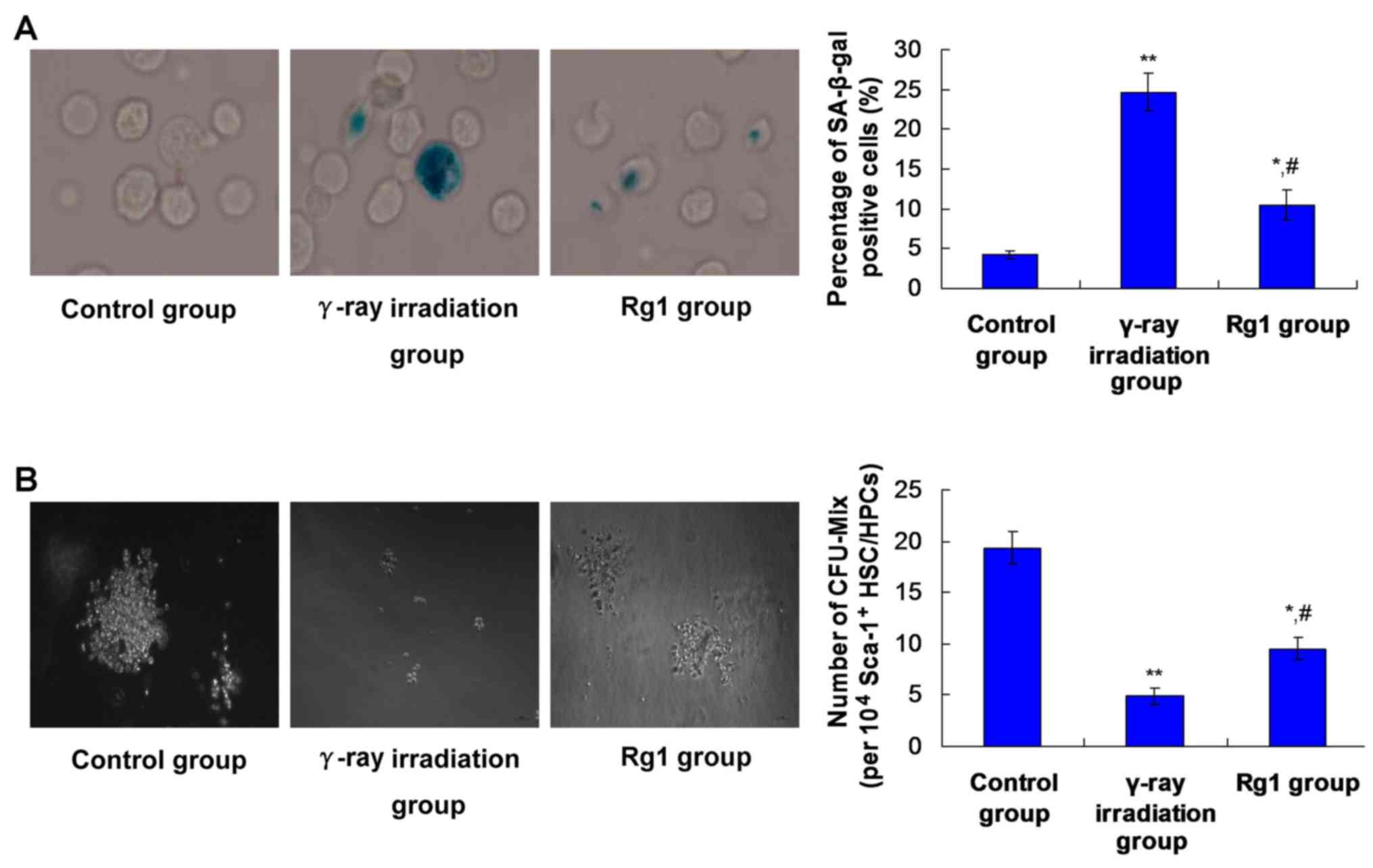
Rg1 affects the senescence of
Sca-1+HSC/HPCs in γ-ray irradiated aging mice
SA-β-Gal is an extensively applied biomarker for
aging cells (24). A decreased
CFU-Mix formation capacity is also an evaluative marker for the
senescence of HSCs (25). Therefore,
an SA-β-Gal staining assay and CFU-Mix formation assay were
performed to evaluate the effects of Rg1 on the senescence of
Sca-1+ HSC/HPCs.
The results showed that γ-ray irradiation induced a
significant increase in the percentage of SA-β-Gal stained
Sca-1+ HSC/HPCs (P<0.01; Fig. 1A) and a significant reduction in the
CFU-Mix counts of Sca-1+ HSC/HPCs, compared with the
control group (P<0.01; Fig. 1B).
In the Rg1 group, the proportion of SA-β-Gal stained HSC/HPCs was
significantly decreased (Fig. 1A;
P<0.05) and the CFU-Mix count was significantly increased
(Fig. 1B; P<0.05), compared with
the respective values in the γ-ray irradiation group. However, the
percentage of SA-β-Gal stained HSC/HPCs and CFU-Mix numbers in the
γ-ray irradiation group were also significantly higher and
significantly lower, respectively, compared with those in the
control group (Fig. 1;
P<0.05).
Rg1 increased the mRNA levels of
SIRT1/SIRT3 signaling molecules in Sca-1+ HSC/HPCs
Cell proliferation-associated SIRT1 and SIRT3
molecules were examined using RT-qPCR. Compared with the control
group, SIRT1 (Fig. 2A) and SIRT3
(Fig. 2B) mRNA levels in the γ-ray
irradiation group were significantly decreased (both P<0.01).
However, Rg1 treatment (Rg1 group) significantly upregulated SIRT1
(Fig. 2A) and SIRT3 (Fig. 2B) mRNA levels compared with those in
the γ-ray irradiation group (P<0.05).
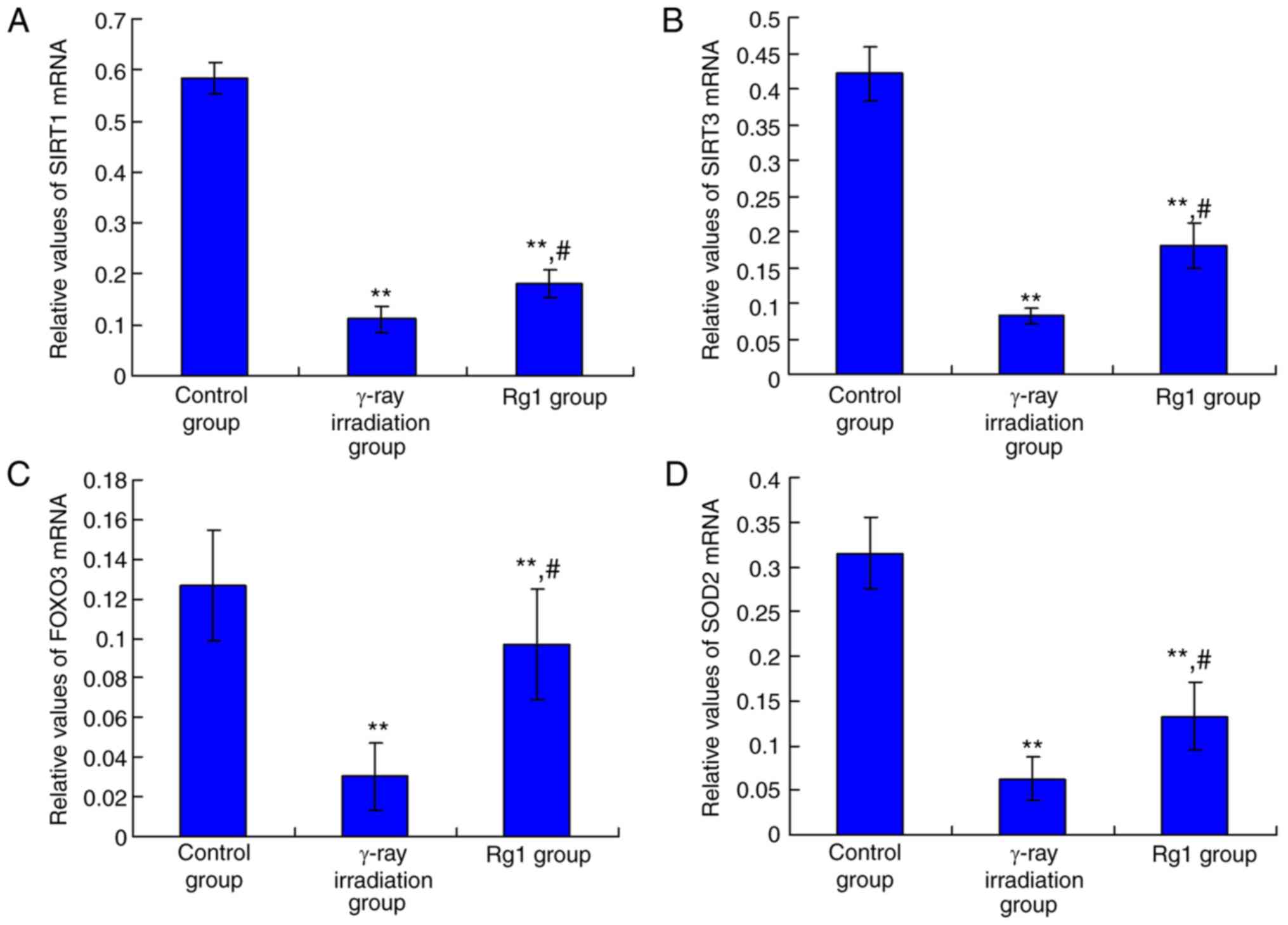 | Figure 2Effects of Rg1 treatment on SIRT1,
SIRT3, FOXO3 and SOD2 mRNA expression in Sca-1+
HSC/HPCs. Relative mRNA expression of (A) SIRT1, (B) SIRT3, (C)
FOXO3 and (D) SOD2. Results are presented as mean ± SD (n=6).
**P<0.01 vs. the control group; #P<0.05
vs. the γ-ray irradiation group. Rg1, ginsenoside Rg1; SIRT,
sirtuin; FOXO3, forkhead box O3; SOD, superoxide dismutase 2;
Sca-1+, stem cell antigen 1 positive; HSC, hematopoietic
stem cell; HPC, hematopoietic progenitor cell. |
The downstream molecule of SIRT1 (FOXO3) and the
downstream molecule of SIRT3 (SOD2) were also examined. The results
indicated that FOXO3 (Fig. 2C) and
SOD2 (Fig. 2D) mRNA levels were
significantly decreased in the γ-ray irradiation group compared
with the control group (both P<0.01). However, Rg1 treatment
significantly increased the mRNA levels of FOXO3 (Fig. 2C) and SOD2 (Fig. 2D) compared with those in the γ-ray
irradiation group (both P<0.05).
Rg1 increased the expression of
SIRT1/SIRT3 signaling molecules in Sca-1+ HSC/HPCs
The SIRT1/SIRT3 signaling pathway-associated
molecules, namely SIRT1, SIRT3, FOXO3 and SOD2, were also examined
using western blotting (Fig. 3A).
The results showed that SIRT1, SIRT3, FOXO3 and SOD2 expression
levels in the γ-ray irradiation group were significantly decreased
compared with their respective levels in the control group
(Fig. 3B; all P<0.01). However,
when compared with the γ-ray irradiation group, the SIRT1, SIRT3,
FOXO3 and SOD2 expression levels in the Rg1 group were
significantly increased (Fig. 3B;
all P<0.05).
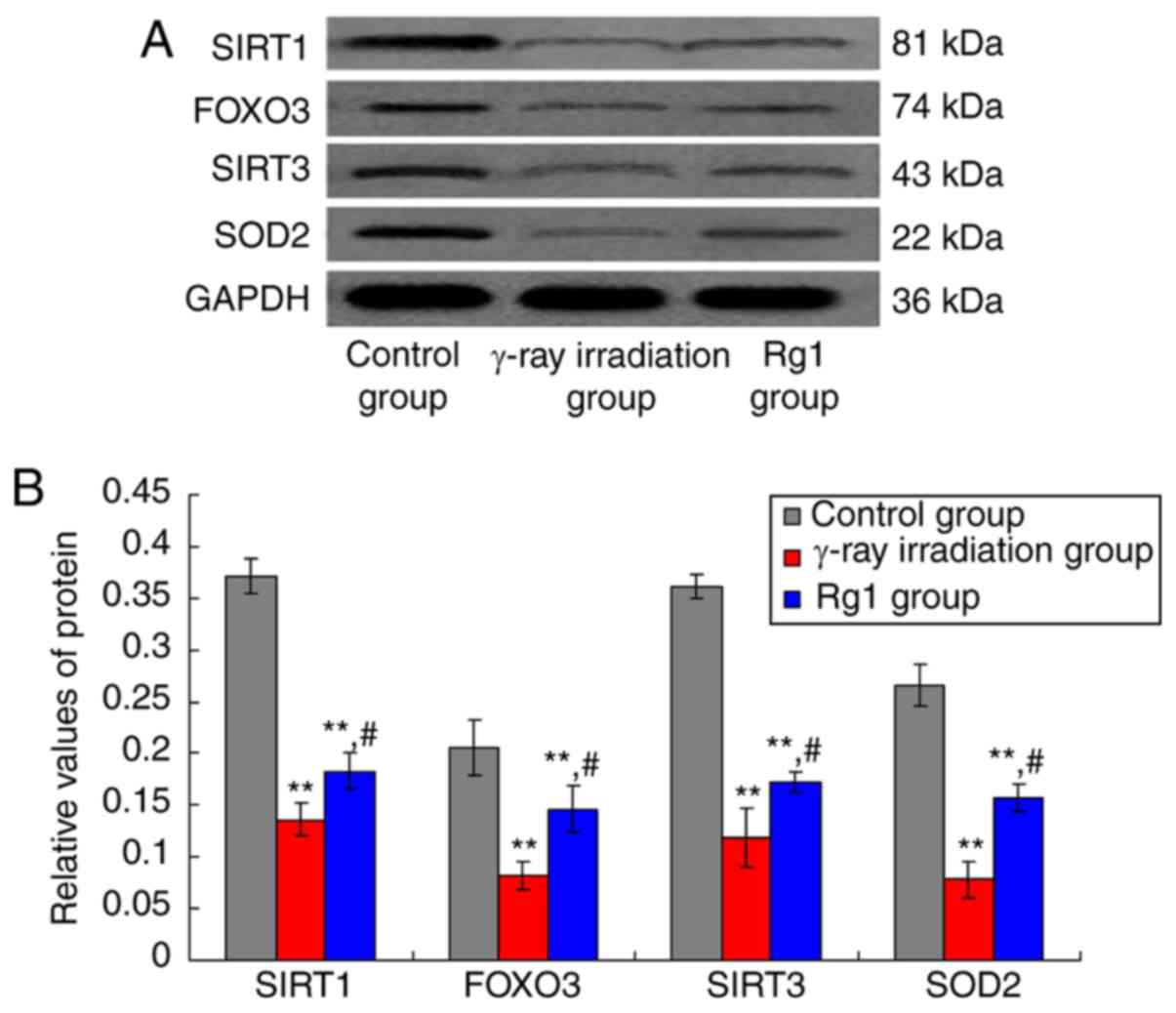 | Figure 3Effects of Rg1 treatment on SIRT1,
SIRT3, FOXO3 and SOD2 protein expression in Sca-1+
HSC/HPCs. (A) Western blot bands for SIRT1, SIRT3, FOXO3 and SOD2.
(B) Quantification of relative SIRT1, SIRT3, FOXO3 and SOD2
expression. Results are presented as mean ± SD (n=6).
**P<0.01 vs. the control group; #P<0.05
vs. the γ-ray irradiation group. Rg1, ginsenoside Rg1; SIRT,
sirtuin; FOXO3, forkhead box O3; SOD, superoxide dismutase 2;
Sca-1+, stem cell antigen 1 positive; HSC, hematopoietic
stem cell; HPC, hematopoietic progenitor cell. |
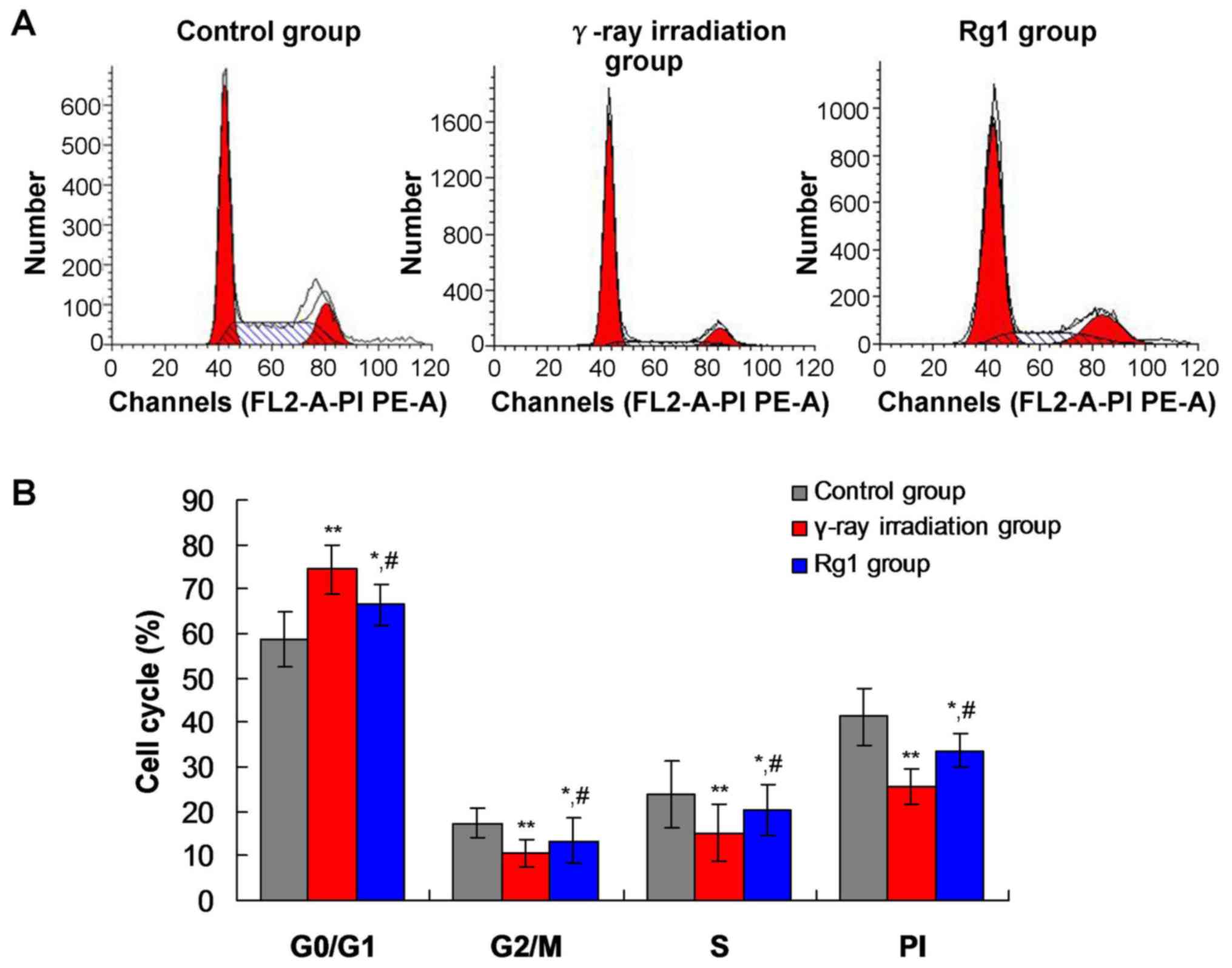
Rg1 impacted the cell cycle
distribution of Sca-1+ HSC/HPCs in the γ-ray irradiation
aging mouse model
Previous studies (26,27)
reported that SIRT1 and SIRT3 participate in cell proliferation and
cell cycle regulation. Furthermore, HSCs undergoing aging are
arrested at the G1 stage (28).
Therefore, the cell cycle distribution of Sca-1+
HSC/HPCs was analyzed using FCM. The significant G1 phase arrest of
Sca-1+ HSC/HPCs was observed, and the percentages of
cells in the S and M phases were significantly decreased in the
γ-ray irradiation and Rg1 groups, compared with those in the
control group (Fig. 4; all
P<0.05). However, the percentage of Sca-1+ HSC/HPCs
arrested at the G1 phase in the Rg1 group was significantly
decreased compared with that in the γ-ray irradiation group
(Fig. 4; P<0.05).
The proliferative index (PI; S + G2/M) in the γ-ray
irradiation group was significantly decreased compared with that in
the control group (Fig. 4;
P<0.01). However, the PI of Sca-1+ HSC/HPCs in the
Rg1 group was significantly increased compared with that in the
γ-ray irradiation group (Fig. 4;
P<0.05).
Discussion
Scientific studies have demonstrated that the
Chinese herbal drug ginseng is characterized by anti-injury,
anti-aging and anti-oxidant functions, and the ability to enhance
immunity (29,30). Zhu et al (31) found that Rg1 could prolong the life
of mice and delay the aging processes of human lung fibroblasts. In
the aging processes of cells, HSC/HPCs in the hematopoietic system
exhibit progressive and morphological changes (32). Zhou et al (33) reported that Rg1 delays the tert-butyl
hydroperoxide-induced aging of HSCs.
A previous study (34) reported that radiation induces damage
or injury in HSC/HPCs, causes cells to undergo apoptosis and
senescence, and reduces the hematological reconstitution function
of HSC/HPCs. Chen et al (11)
found that Rg1 promotes the proliferation of hematopoietic cells;
however, further evaluation of the mechanism is required. The
present study showed that γ-ray irradiation significantly decreased
the WBC, RBC and PLT counts of mice compared with those in
un-irradiated controls; however, Rg1 was able to attenuate these
reductions in blood cell levels. Therefore, the present findings
confirm that Rg1 prevents irradiation-induced hematopoietic
dysfunction.
It is speculated that the improvement of
hematopoietic function observed in the mice might be triggered by
increased levels of Sca-1+ HSC/HPCs. SA-β-Gal
accumulates in aging cells and reflects the dysfunction of cells
(24). Furthermore, the ability of
mixed hematopoietic progenitor cells to form colonies decreases
following the regression of self-renewal potential in aging
Sca-1+ HSC/HPCs (35).
Therefore, the aging or senescence of Sca-1+ HSC/HPCs
was evaluated using the SA-β-Gal method and CFU-Mix assay in the
present study. The results demonstrated that the percentage of
SA-β-Gal stained HSC/HPCs was significantly increased and CFU-Mix
counts were significantly decreased following γ-ray irradiation
compared with those in the control group. Notably, Rg1 treatment
significantly attenuated the γ-ray irradiation-induced and
aging/senescence-associated changes. These findings are consistent
with a previous study (11), which
reported that Rg1 delays the irradiation-induced senescence of
Sca-1+ HSC/HPCs. Therefore, the present data suggest
that Rg1 effectively counteracts the γ-ray irradiation-induced
persistent senescence/aging of HSC/HPCs.
At present, several signaling pathways have been
demonstrated to participate in cell senescence or aging; these
include SIRT1 and SIRT3 pathways, which may inhibit cellular
senescence by regulating the cell cycle (26,27,36).
SIRT3 upregulates the self-renewal and multi-differentiation
capacity of HSCs, and further delays their aging processes
(37). SIRT3 also enhances the
activity of SOD2 by triggering deacetylation, which plays an
important role in the negative regulation of organism aging
(38). Therefore, SIRT3 and SOD2
expression in Sca-1+ HSC/HPCs were examined in the
present study. The results indicated that irradiation significantly
induced the downregulation of SIRT3 and SOD2 in Sca-1+
HSC/HPCs. This suggests that irradiation might initiate the
progression of aging and oxidative stress. SOD2 is also considered
to be an important anti-oxidant defense enzyme; therefore, it is
speculated that γ-ray irradiation might result in the oxidative
damage of Sca-1+ HSC/HPCs via the reduction of SOD2
activity. However, the results of the present study revealed that
Rg1 significantly inhibited the irradiation-induced downregulation
of SIRT3 and SOD2 expression. This suggests that Rg1 is able to
inhibit the aging of Sca-1+ HSC/HPCs and increaseSOD2
activity, which is critical to prevent Sca-1+ HSC/HPCs
from undergoing irradiation-induced aging or oxidative stress.
It has previously been reported that SIRT1 can delay
cardiovascular and neuron aging (39). Morris et al (40) reported that FOXO3 is an important
gene for human longevity. It is well known that the SIRT1 gene can
interact with FOXO3 and regulate FOXO3 expression in the
mitochondria of cells (41).
Therefore, it is hypothesized that the effects of SIRT1 on aging
might be triggered by FOXO3 expression. Therefore, SIRT1 and FOXO3
expression were examined in the present study, and it was found
that irradiation treatment significantly downregulated SIRT1 and
FOXO3 levels in Sca-1+ HSC/HPCs. However, Rg1 treatment
significantly attenuated the irradiation-induced SIRT1 and FOXO3
downregulation. These results suggest that Rg1 may also suppress
the aging of Sca-1+ HSC/HPCs via the activation of SIRT3
and FOXO3 expression.
A previous study (26) reported that SIRT3 overexpression
disrupts mitochondrial proteostasis, induces cell cycle arrest and
inhibits the proliferation of cells. In another study, SIRT1 was
reported to contribute to delay of the epithelial cell cycle in
diabetic corneas (42). Therefore,
the cell cycle status of Sca-1+ HSC/HPCs was
investigated in the present study to determine the proportion of
cells in the G1, S, G2 and M phases. The results indicated that
irradiation triggered cell cycle arrest at the G1 phase and that
cell senescence was induced. However, as discussed above, Rg1
treatment significantly increased SIRT1 and SIRT3 expression in
irradiated Sca-1+ HSC/HPCs. The increased SIRT1 and
SIRT3 expression may have inhibited cells from arresting in the G1
phase and suppressed cell senescence. Irradiation-induced arrest of
the cell cycle at the G1 phase, without entry to the S phase, is
mainly induced by the gradual loss of mitosis reactivity and DNA
synthesis activity in aging cells (28). The cell cycle results in the present
study suggest that Rg1 treatment significantly attenuated the
irradiation-induced G1 arrest by regulating the expression of SIRT1
and SIRT3. This may have triggered the progression of cells from
the G1 phase to the S phase, and thereby delayed the senescence of
Sca-1+ HSC/HPCs.
In conclusion, Rg1 decreased the percentage of
SA-β-Gal stained Sca-1+ HSC/HPCs, and increased the
ability of HSC/HPCs to form colonies. These results suggest that
Rg1 treatment attenuates γ-ray irradiation-induced aging in a mouse
model. Rg1 may exert anti-aging effects via the enhancement of SOD2
activity and reduction of SIRT3, SIRT1 and FOXO3 expression. This
study provides a promising hypothesis for the mechanism by which
Rg1 application delays the aging of Sca-1+ HSC/HPCs.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (grant nos. 81660731 and
81673748).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YLT and YZ designed this study and wrote the
manuscript. YLT, YZ, YPW, YHH and JCD performed the experiments or
tests. YL and CLW analyzed the data. CLW reviewed the literature.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experiments were approved by the Ethics
Committee of the Key Laboratory of Cell Biology (Kunming,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Xu YF, Liang ZJ, Kuang ZJ, Chen JJ, Wu J,
Lu XE, Jiang WW, Fan PL, Tang LY, Li YT, et al: Effect of Suo Quan
Wan on the bladder function of aging rats based on the
β-adrenoceptor. Exp Ther Med. 13:3424–3432. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Li YN, Guo Y, Xi MM, Yang P, Zhou XY, Yin
S, Hai CX, Li JG and Qin XJ: Saponins from Aralia taibaiensis
attenuate D-galactose-induced aging in rats by activating FOXO3a
and Nrf2 pathways. Oxid Med Cell Longev.
2014(320513)2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bustos ML, Huleihel L, Kapetanaki MG,
Lino-Cardenas CL, Mroz L, Ellis BM, McVerry BJ, Richards TJ,
Kaminski N, Cerdenes N, et al: Aging mesenchymal stem cells fail to
protect because of impaired migration and antiinflammatory
response. Am J Respir Crit Care Med. 189:787–798. 2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Montezano AC and Touyz RM: Reactive oxygen
species, vascular Noxs, and hypertension: Focus on translational
and clinical research. Antioxid Redox Signal. 20:164–182.
2014.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Liochev SI: Reactive oxygen species and
the free radical theory of aging. Free Radic Biol Med. 60:1–4.
2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Milano F, Merriam F, Nicoud I, Li J,
Gooley TA, Heimfeld S, Imren S and Delaney C: Notch-expanded murine
hematopoietic stem and progenitor cells mitigate death from lethal
radiation and convey immune tolerance in mismatched recipients.
Stem Cells Transl Med. 6:566–575. 2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Sadowska-Bartosz I and Bartosz G: Effect
of antioxidants supplementation on aging and longevity. Biomed Res
Int. 2014(404680)2014.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Guerra-Araiza C, Alvarez-Mejia AL,
Sanchez-Torres S, Farfan-Garcia E, Mondragon-Lozano R,
Pinto-Almazan R and Salgado-Ceballos H: Effect of natural exogenous
antioxidants on aging and on neurodegenerative diseases. Free Radic
Res. 47:451–462. 2013.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Chu SF and Zhang JT: New achievements in
ginseng research and its future prospects. Chin J Integr Med.
15:403–408. 2009.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Xu FT, Li HM, Yin QS, Cui SE, Liu DL, Nan
H, Han ZA and Xu KM: Effect of ginsenoside Rg1 on proliferation and
neural phenotype differentiation of human adipose-derived stem
cells in vitro. Can J Physiol Pharmacol. 92:467–475.
2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chen C, Mu XY, Zhou Y, Shun K, Geng S, Liu
J, Wang JW, Chen J, Li TY and Wang YP: Ginsenoside Rg1 enhances the
resistance of hematopoietic stem/progenitor cells to
radiation-induced aging in mice. Acta Pharmacol Sin. 35:143–150.
2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Dali-Youcef N, Lagouge M, Froelich S,
Koehl C, Schoonjans K and Auwerx J: Sirtuins: The ‘magnificent
seven’, function, metabolism and longevity. Ann Med. 39:335–345.
2007.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Imai S, Johnson FB, Marciniak RA, McVey M,
Park PU and Guarente L: Sir2: An NAD-dependent histone deacetylase
that connects chromatin silencing, metabolism, and aging. Cold
Spring Harb Symp Quant Biol. 65:297–302. 2000.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Buler M, Aatsinki SM, Izzi V and Hakkola
J: Metformin reduces hepatic expression of SIRT3, the mitochondrial
deacetylase controlling energy metabolism. PLoS One.
7(e49863)2012.PubMed/NCBI View Article : Google Scholar
|
|
15
|
D'Aquila P, Rose G, Panno ML, Passarino G
and Bellizzi D: SIRT3 gene expression: A link between inherited
mitochondrial DNA variants and oxidative stress. Gene. 497:323–329.
2012.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Finley LW, Haas W, Desquiret-Dumas V,
Wallace DC, Procaccio V, Gygi SP and Haigis MC: Succinate
dehydrogenase is a direct target of sirtuin 3 deacetylase activity.
PLoS One. 6(e23295)2011.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Libert S and Guarente L: Metabolic and
neuropsychiatric effects of calorie restriction and sirtuins. Annu
Rev Physiol. 75:669–684. 2013.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yao H, Chung S, Hwang JW, Rajendrasozhan
S, Sundar IK, Dean DA, McBurney MW, Guarente L, Gu W, Ronty M, et
al: SIRT1 protects against emphysema via FOXO3-mediated reduction
of premature senescence in mice. J Clin Invest. 122:2032–2045.
2012.PubMed/NCBI View
Article : Google Scholar
|
|
19
|
Chen Y, Zhang J, Lin Y, Lei Q, Guan KL,
Zhao S and Xiong Y: Tumour suppressor SIRT3 deacetylates and
activates manganese superoxide dismutase to scavenge ROS. EMBO Rep.
12:534–541. 2011.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Miao L and St Clair DK: Regulation of
superoxide dismutase genes: Implications in disease. Free Radic
Biol Med. 47:344–356. 2009.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Liwski CJ, Padley DJ, Gustafson MP,
Winters JL, Gastineau DA and Jacob EK: Discordant CD34+
cell results in peripheral blood and hematopoietic progenitor
cell-apheresis product: Implications for clinical decisions and
impact on patient treatment. Transfusion. 54:541–544.
2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhou Y, Yang B, Yao X and Wang Y:
Establishment of an aging model of Sca-1+ hematopoietic
stem cell and studies on its relative biological mechanisms. In
Vitro Cell Dev Biol Anim. 47:149–156. 2011.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Schmittgen TD, Zakrajsek BA, Mills AG,
Gorn V, Singer MJ and Reed MW: Quantitative reverse
transcription-polymerase chain reaction to study mRNA decay:
Comparison of endpoint and real-time methods. Anal Biochem.
285:194–204. 2000.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Dimri GP, Lee X, Basile G, Acosta M, Scott
G, Roskelley C, Medrano EE, Linskens M, Rubelj I and Pereira-Smith
O: A biomarker that identifies senescent human cells in culture and
in aging skin in vivo. Proc Natl Acad Sci USA. 92:9363–9367.
1995.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Tang YL, Zhou Y, Wang YP, Wang JW and Ding
JC: SIRT6/NF-κB signaling axis in ginsenoside Rg1-delayed
hematopoietic stem/progenitor cell senescence. Int J Clin Exp
Pathol. 8:5591–5596. 2015.PubMed/NCBI
|
|
26
|
Giralt A and Villarroya F: SIRT3, a
pivotal actor in mitochondrial functions: Metabolism, cell death
and aging. Biochem J. 444:1–10. 2012.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Atkins KM, Thomas LL, Barroso-Gonzalez J,
Thomas L, Auclair S, Yin J, Kang H, Chung JH, Dikeakos JD and
Thomas G: The multifunctional sorting protein PACS-2 regulates
SIRT1-mediated deacetylation of p53 to modulate p21-dependent
cell-cycle arrest. Cell Rep. 8:1545–1557. 2014.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Stein GH, Beeson M and Gordon L: Failure
to phosphorylate the retinoblastoma gene product in senescent human
fibroblasts. Science. 249:666–669. 1990.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Chen X, Zhang J, Fang Y, Zhao C and Zhu Y:
Ginsenoside Rg1 delays tert-butyl hydroperoxide-induced premature
senescence in human WI-38 diploid fibroblast cells. J Gerontol A
Biol Sci Med Sci. 63:253–264. 2008.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Asadullina NR, Usacheva AM and Gudkov SV:
Protection of mice against X-ray injuries by the post-irradiation
administration of inosine-5'-monophosphate. J Radiat Res.
53:211–216. 2012.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhu J, Mu X, Zeng J, Xu C, Liu J, Zhang M,
Li C, Chen J, Li T and Wang Y: Ginsenoside Rg1 prevents cognitive
impairment and hippocampus senescence in a rat model of
D-galactose-induced aging. PLoS One. 9(e101291)2014.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Hu W, Jing P, Wang L, Zhang Y, Yong J and
Wang Y: The positive effects of Ginsenoside Rg1 upon the
hematopoietic microenvironment in a D-Galactose-induced aged rat
model. BMC Complement Altern Med. 15(119)2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Zhou Y, Yang B, Jiang R, Yao X and Wang
YP: Mechanism of ginsenoside Rg1 in the delayed senescence of
hematopoietic stem cell. Zhonghua Yi Xue Za Zhi. 90:3421–3425.
2010.(In Chinese). PubMed/NCBI
|
|
34
|
Wang Y, Liu L, Pazhanisamy SK, Li H, Meng
A and Zhou D: Total body irradiation causes residual bone marrow
injury by induction of persistent oxidative stress in murine
hematopoietic stem cells. Free Radic Biol Med. 48:348–356.
2010.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Kamminga LM, van Os R, Ausema A, Noach EJ,
Weersing E, Dontje B, Vellenga E and de Haan G: Impaired
hematopoietic stem cell functioning after serial transplantation
and during normal aging. Stem Cells. 23:82–92. 2005.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Kwon Y, Kim J, Lee CY and Kim H:
Expression of SIRT1 and SIRT3 varies according to age in mice. Anat
Cell Biol. 48:54–61. 2015.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Brown K, Xie S, Qiu X, Mohrin M, Shin J,
Liu Y, Zhang D, Scadden DT and Chen D: SIRT3 reverses
aging-associated degeneration. Cell Rep. 3:319–327. 2013.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Lu J, Cheng K, Zhang B, Xu H, Cao Y, Guo
F, Feng X and Xia Q: Novel mechanisms for superoxide-scavenging
activity of human manganese superoxide dismutase determined by the
K68 key acetylation site. Free Radic Biol Med. 85:114–126.
2015.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Chang HC and Guarente L: SIRT1 and other
sirtuins in metabolism. Trends Endocrinol Metab. 25:138–145.
2014.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Morris BJ, Willcox DC, Donlon TA and
Willcox BJ: FOXO3: A major gene for human longevity-a mini-review.
Gerontology. 61:515–525. 2015.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Das S, Mitrovsky G, Vasanthi HR and Das
DK: Antiaging properties of a grape-derived antioxidant are
regulated by mitochondrial balance of fusion and fission leading to
mitophagy triggered by a signaling network of
Sirt1-Sirt3-Foxo3-PINK1-PARKIN. Oxid Med Cell Longev.
2014(345105)2014.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Gao J, Wang Y, Zhao X, Chen P and Xie L:
MicroRNA-204-5p-mediated regulation of SIRT1 contributes to the
delay of epithelial cell cycle traversal in diabetic corneas.
Invest Ophthalmol Vis Sci. 56:1493–1504. 2015.PubMed/NCBI View Article : Google Scholar
|