Introduction
Angiogenesis, the formation of new blood vessels,
plays an important role in bone regeneration and osteoblast
differentiation (1) and provides
essential nutrients and oxygen during bone formation (2). Previous studies have indicated an
association between angiogenesis and osteogenesis during bone
healing and regeneration (2,3). Compared with vasculogenesis,
angiogenesis occurs more frequently and plays a major role during
the process of bone healing (4).
Thus, treatments that improve angiogenesis maybe a beneficial for
bone regeneration (4).
Angiogenesis is a complex process that can be
manipulated by interactions between vascular cells and the
microenvironment (3). Previous
studies have suggested that mesenchymal stem cells (MSCs) may have
a function in cell-based regulation of angiogenesis (5,6). MSCs
can promote tube formation and enhance the proliferation of
endothelial cells via a paracrine pathway (7). Zuo et al (8) found that conditioned medium (CM) from
MSCs could enhance angiogenesis in a post-infarction model. MSCs
can also be obtained from multiple tissues including bone marrow,
adipose tissue, peripheral blood and synovial tissue (9,10).
However, the wide use of MSCs in therapeutic angiogenesis is
limited by the low number of cells that can be isolated from these
tissues and their instability that results from donor age-dependent
differentiation (11). Therefore,
alternative seed cells are required for the pursuit of this type of
therapeutic strategy.
Human amnion-derived mesenchymal stem cells (hAMSCs)
show a powerful ability to promote angiogenesis (12) and exhibit stable viability and low
level anti-inflammatory properties (13). Abundant amounts of amniotic tissuecan
be donated with fewer ethical issues compared with other tissue
types commonly donated for the isolation of seed cells (14). A previous study we carried out showed
that hAMSCs significantly enhanced the angiogenesis of human
umbilical vein endothelial cells (HUVECs) both in vivo and
in vitro (15), however, the
molecular mechanism underlying this process remain largely
unknown.
Circular RNA (circRNA) is a subclass of endogenous
non-coding RNA with a loop structure generated by back splicing
(16). Recent studies have found
that circRNAs are widely expressed in various human cell types and
have multiple biological functions in the process of development
and disease (17,18). The role of circRNAs in angiogenesis
may involve competing endogenous RNAs (ceRNA) (19). Thus, the present study aimed to
investigate whether circRNAs play a role in hAMSC-induced
angiogenesis.
The present study investigated the pro-angiogenic
capacity of CM from hAMSCs on HUVECs in a co-culture system. The
results suggested that circ-ATP binding cassette subfamily B member
10 (ABCB10) may play an important role in the promotion of
angiogenesis induced by hAMSC-CM via the circ-ABCB10/microRNA
(miR)-29b-3p/vascular endothelial growth factor A (VEGFA) axis.
Materials and methods
Ethics statement
Study protocols were approved by The Ethics
Committee of The School of Stomatology, Nanjing Medical University
(approval no. PJ2013-037-001). The methodologies used in the
present study were compliant with the Declaration of Helsinki and
written informed consent was obtained from each participant.
Cell culture
hAMSCs were isolated from donated human placentas
from full term pregnancies (from healthy pregnant females; length
of pregnancy range 38-41 weeks) and identified, following a
previously-described method (15). A
total of three donors from the Obstetrics Department of Nanjing
Maternal and Child Health Hospital Affiliated to Nanjing Medical
University (Nanjing, Jiangsu, China), aged 24-27 (mean age, 25.3),
were enrolled between February 2018 and March 2018 in this study.
The cells were cultured in α-MEM (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.), 100 U/l penicillin and 100 mg/l streptomycin at 37˚C and 5%
CO2. Cells at passages 3-5 were used in all experiments.
HUVECs were purchased from China Infrastructure of Cell Line
Resources and cultured in DMEM (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% FBS, 100 U/l penicillin and 100 mg/l
streptomycin (Gibco; Thermo Fisher Scientific, Inc.) at 37˚C and 5%
CO2.
Collection of CM
When hAMSCs had grown to 70-80% confluence in 15 cm
plates, cells were washed three times with PBS and incubated at
37˚C and 5% CO2 with 15 ml serum-free α-MEM containing
penicillin-streptomycin (100 U/ml penicillin and 100 µg/ml
streptomycin) and 15 ml serum-free DMEM containing
penicillin-streptomycin (100 U/ml penicillin and 100 µg/ml
streptomycin) for 24 h. A 30 ml quantity of medium was collected
after 24 h, centrifuged at 3,000 x g and 4˚C for 3 min followed by
a further centrifugation for 5 min at 1,500 x g and 4˚C. Medium was
then filtered through 0.45-µm filters (Merck KGaA) and stored at
-80˚C for use as CM. For the collection of CM, cells from the three
different donors were collected separately. Subsequently, three
replicate experiments using CM from each donor were conducted
(15), unless otherwise stated.
Serum-free medium (containing 50% v/v DMEM, 50% v/v α-MEM, 100 U/ml
penicillin and 100 µg/ml streptomycin) was used as control
unconditioned medium.
Endothelial cell proliferation
assay
The proliferation of HUVECs was determined using a
Cell Counting Kit 8 (CCK-8) assay (Donjindo Molecular Technologies,
Inc.), according to the manufacturer's instructions. HUVECs were
initially seeded into 96-well plates at a density of
1x103 cells/well and incubated at 37˚C for 12 h. After
synchronization with DMEM containing 2% FBS for 24 hat 37˚Cand
5%CO2, cells were co-cultured with a dilution of 50 or
80% v/v or 100% v/v CM in fresh control medium (containing 50% v/v
DMEM, 50% v/vα-MEM, 100 U/ml penicillin and 100 µg/ml streptomycin)
for 24 h. Cell proliferation was then measured at 450 nm using a
microplate reader (BioTek Instruments, Inc.). All experimental
results are presented as the means of three replicates performed
under the same conditions.
Tube-formation assay
HUVECs were seeded onto 96-well plates coated with
Matrigel reduced factor (50 µl per well at 37˚C for 1 h; Becton,
Dickinson and Company), at a density of 3,000 cells/well and
cultured for 4-6 h at 37˚C and 5% CO2 with hAMSC-CM (80%
v/v CM) or control unconditioned medium, both supplemented with 1%
FBS. Images of tube formation were obtained using an optical light
microscope at 100X (Carl Zeiss, Inc.) and then five fields were
randomly selected from each well for quantitative analysis of the
total tube length with Image J software (version 1.8.0; http://imagej.nih.gov/ij/).
Wound healing assay and transwell
migration
Prior to the wound healing assay, HUVECs cultured in
DMEM containing 10% FBS were plated (1x105 cells per
well in a 6-well plate) at the logarithmic growth phase until a
monolayer was formed. A straight line was scraped with a 200 µl
pipette tip to create a gap across the cell monolayer. The cell
debris was then removed by washing with PBS three times. hAMSC-CM
(80% v/v CM) containing 2% FBS or control unconditioned medium
containing 2% FBS was added and HUVECs were allowed to proliferate
for 16 h (use of 2% FBS is a potential limitation of this study,
but this was necessary to maintain cell viability). Microscopic
images (5 per well at labeled locations) were obtained at 0 and 16
h using an optical light microscope 40X objective lens (Carl Zeiss,
Inc.). For each image, the sizes of the gaps were measured using
Image J software (version 1.8.0; National Institutes of Health).
For each group, five random locations within each gap were
measured. Cell mobility was calculated as [width at 0 h-width at 16
h]/width at 0 h] x100.
For the Transwell migration assay, 8.0-µm pore
culture inserts (24 well; Corning, Inc.) and HUVECs in log phase
were prepared. A total of 1.5x104 cells/well were
suspended in 200 µl control unconditioned medium without FBS and
loaded into the upper chambers of the insert, and 600 µl hAMSC-CM
or control unconditioned medium containing 10% FBS was added into
the lower chambers. After incubation at 37˚C for 12 h, migrated
cells that had attached to the lower surface of the filter were
fixed at 25˚C in 4% paraformaldehyde for 30 min and then stained
with 0.1% crystal violet at 25˚C for 1 h. Cells were counted and
photographed in five random fields under an optical light
microscope using a 100X objective lens (Carl Zeiss, Inc.). The
migration assay was performed independently three times.
RNA interference, miRNA inhibitor and
transfection
Small interfering RNAs (siRNAs) specific for
circ-ABCB10 and miR-29b-3p inhibitor were purchased from Guangzhou
RiboBio Co., Ltd. siRNA (50 nM) or inhibitor (100 nM) was
transfected with Lipofectamine® 2000 transfection
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. After transfection for 48 h,
circ-ABCB10 and VEGFA expression levels were assessed by reverse
transcription-quantitative PCR (RT-qPCR). siRNA and inhibitor
sequences are shown in Table
SI.
Luciferase reporter assay
The potential binding sites of circ-ABCB10 and
miR-29b-3p were determined using TargetScan version 7.1 (http://www.targetscan.org/vert_71) and the
miRanda database (August 2010 release; http://www.microrna.org/microrna/home.do/). Vectors
containing circ-ABCB10 sequences with the predicted miR-29b-3p
binding sites or sequences mutated were purchased from Guangzhou
RiboBio Co., Ltd. and named ‘circ-wild-type (WT)’ and ‘circ-mutant
(Mut)’. Vectors were then co-infected with miR-29b-3p mimic or
mimic negative control (NC) in 293T cells (purchased from China
Infrastructure of Cell Line Resources). A dual-luciferase assay was
then performedwith the Dual-Luciferase Reporter Assay System
(Promega Corporation) according to the manufacturer's instructions
24 h after the co-transfection. The activity of Renilla
luciferase was used as an internal control, and the relative
luciferase activity was calculated and presented as the ratio of
firefly luciferase activity to Renilla luciferase activity.
The predicted sequences are shown in Table SI.
Western blot analysis
CM or control unconditioned medium was removed when
the cells had reached 80% confluence and the cells were washed
three times with pre-chilled 1X PBS, before 300 µl RIPA buffer
(Beyotime Institute of Biotechnology) was added to the cells to
extract cellular protein. After incubation for 30 min on ice, lysed
cells were transferred into a 1.5-ml centrifuge tube and
centrifuged at 4˚C for 15 min at 10,656 x g. Protein quantification
was performed using the bicinchoninic acid method and then loaded
at 40 µg per lane. Protein was separated using 10% SDS-PAGE and
electrotransferred to PVDF membranes (Merck KGaA). The membranes
were incubated with primary antibody against VEGFA (1:200; cat. no.
ab171828; Abcam) at room temperature for 1 h, or GAPDH (1:1,000;
cat. no. ab8245; Abcam), AKT [1:1,000; cat. no. 4691; Cell
Signaling Technology, Inc. (CST)], phosphorylated (p)-AKT (1:1,000;
cat. no. 4060; CST), ERK1/2 (1:1,000; cat. no. 4695; CST) and
p-ERK1/2 (1:1,000; cat. no. 4377; CST) primary antibodies at 4˚C
overnight. Then, membranes were washed with 0.1% Tween-20 in TBS,
and incubated with a secondary antibody (1:10,000; cat. no.
ab150081; Abcam) conjugated to horseradish peroxidase at room
temperature for 1 h. ECL detection was performed using Pierce ECL
reagent (Thermo Fisher Scientific, Inc) and the densitometry
carried out using Image Lab software (version 3.0; Bio-Rad
Laboratories, Inc.).
Enzyme-linked immunosorbent assay
(ELISA)
CM from hAMSCs was collected. HUVECs at 80%
confluence were incubated at 37˚C with CM (80% v/v of hAMSC-CM in
fresh control unconditioned medium) for 24 h, and then the medium
was collected, which was referred to as ‘CM-aft’. To measure the
changes in VEGFA protein expression levels in CM and CM-aft, an
ELISA assay was performed with the human VEGF Quantikine ELISA kit
(cat. no. DVE00; R&D Systems, Inc.) according to the
manufacturer's protocol. Absorbance was detected at 450 nm. Both CM
and CM-aft were transferred into ultra-filtration conical tubes
(EMD Millipore) and concentrated to 100X. Then, both 100X CM and
100X CM-aft were used to extract total RNA.
RT-qPCR
Total RNA was extracted using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). The quantity
and quality of RNA was measured using Nanodrop 2000
spectrophotometry (Thermo Fisher Scientific, Inc.). Total RNA
(1,000 ng) was reverse-transcribed to cDNA at 42˚C for 30 min using
Prime Script RT master mix (Takara Bio, Inc.) and the TaqMan
microRNA RT kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.) was used for the RT of miRNA with specific primers at 42˚C
for 60 min and then 70˚C for 30 min. The level of RNA expression
was detected using SYBR Green Master mix (Takara Bio, Inc.) using
an ABI 7500 system (Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. The thermocycling conditionswere
as follows: 95˚C for 5 min, before 40 cycles of 95˚C for 10 sec and
60˚C for 45 sec. Relative fold changes of RNA expression were
calculated using the 2-ΔΔCq method
(20). GAPDH or U6 were used as the
internal standard controls. RT-qPCR analysis was independently
performed three times. Primer sequences are shown in Table SI.
Statistical analysis
All data were collected and analyzed from three
independent repeats. All image analysis was performed by a blinded
observer. Data are presented as the mean ± SD, unless otherwise
indicated. Data were analyzed using SPSS 20.0 software (IBM Corp.)
to perform one-way ANOVA followed by Tukey's test, or a two-tailed
Student's t-test (unpaired). P<0.05 was considered to indicate a
statistically significant difference.
Results
hAMSC-CM enhances HUVEC proliferation,
migration and tube formation
Dilutions of 50 and 80% v/v hAMSC-CM in fresh
medium, referred to as ‘50% CM’ and ‘80% CM’, were used to
investigate the effect of CM on HUVEC proliferation using a CCK-8
assay. The results suggested that 80% CM promoted the viability of
HUVECs to a higher level compared with 50% CM (Fig. 1A). Therefore, 80% CM was used for all
further experiments. The present study investigated the effects of
hAMSC-CM on the migration of HUVECs during wound healing and in
Transwell assays. The results indicated that significantly
increased migratory ability of HUVECs was induced by hAMSC-CM
(Fig. 1B and C). In addition, tube-formation assays were
performed to investigate the pro-angiogenic role of hAMSC-CM on
HUVECs. It was found that the total tube length formed by HUVECs
was significantly higher when treated with hAMSC-CM compared with
the control (Fig. 1D).
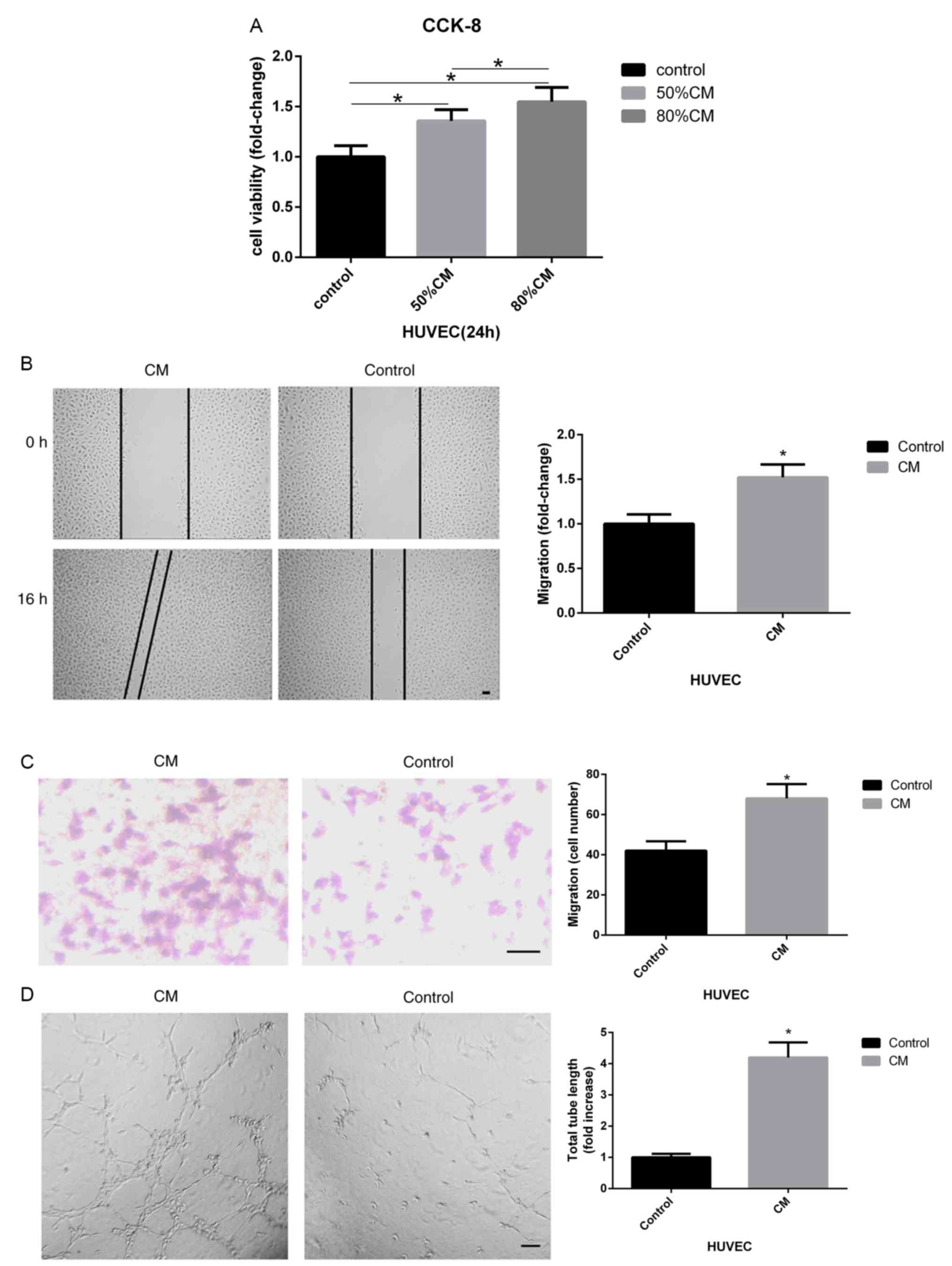 | Figure 1hAMSC-CM increases the proliferation,
migration and tube formation of HUVECs. (A) CCK-8 assay of HUVECs
treated with different dilutions of hAMSC-CM. (B) Influence of
hAMSC-CM on HUVEC migration was measured by wound healing assay.
Scale bar, 100 µm. (C) Effect of hAMSC-CM on HUVEC migration was
measured by Transwell assay. Scale bar, 100 µm. (D) Influence of
hAMSC-CM on HUVEC angiogenesis was measured by tube formation
assay. Scale bar, 200 µm. (E) Changes in mRNA expression levels
that may be involved in the process of angiogenesis in HUVECs were
measured by reverse transcription-quantitative PCR. (F) Protein
expression levels of VEGFA, p-ERK1/2, ERK1/2, p-AKT and AKT in
HUVECs were detected by western blotting. (G) VEGFA concentration
in hAMSC-CM or hAMSC-CM treated with HUVECs for 24 h was assessed
by ELISA. *P<0.05 vs. control unless otherwise
indicated. HUVECs, human umbilical vein endothelial cells; hAMSCs,
human amnion-derived mesenchymal stem cells; CCK-8, Cell Counting
Kit-8;CM, conditioned medium; CM, hAMSC-CM; CM-aft, hAMSC-CM
incubated with HUVECs for 24 h; VEGF, vascular endothelial growth
factor; p-, phosphorylated; MSCs, mesenchymal stem cells; Ang-1,
angiopoietin; HIF-1α, hypoxia-inducible factor 1-α; KDR, kinase
insert domain receptor. |
To investigate the potential molecular mechanism of
hAMSC-CM-induced angiogenesis in HUVECs, the mRNA expression level
of a number of genes that may be involved in angiogenesis was
determined using RT-qPCR. After incubation with hAMSC-CM for 24 h,
HUVECs showed a significantly increased mRNA expression level of
VEGFA compared with that of the control group (Fig. 1E). This change in VEGFA expression
was also identified at the protein level using western blotting. In
addition, western blotting results identified increased levels of
p-ERK1/2 but not p-AKT (Fig. 1F).
MSCs can enhance tube formation in endothelial cells by secreting
proteins such as VEGFA (21).
Therefore, the present study hypothesized that a similar pattern
could occur during tube formation in HUVECs treated with hAMSC-CM.
However, ELISA results suggested that the concentration of VEGFA in
hAMSC-CM was low, while the concentration of VEGFA in hAMSC-CM
significantly increased after treating HUVECs for 24 h (Fig. 1G). It was demonstrated that the
increased levels of VEGFA may have been secreted from HUVECs
treated with hAMSC-CM and not from hAMSC-CM directly.
Collectively, the present results suggested that
hAMSC-CM enhanced HUVEC proliferation, migration and tube
formation, which may result from the induction of the VEGF
signaling pathway.
circ-ABCB10 involvement in the
pro-angiogenic role of hAMSC-CM
Previous studies showed that circRNAs may be
involved in the process of angiogenesis during development or
disease (19,22-26),
including circ_002136, which plays an important role in regulating
angiogenesis in glioma via the miR-138-5p/SOX13 axis (27). The present study investigated several
circRNAs that may potentially regulate the process of angiogenesis.
The present results suggested a number of circRNAs were upregulated
in HUVECs after treatment with hAMSC-CM. Among them, circ-ABCB10
had the highest fold-change, while linear ABCB10 expression level
was unchanged (Fig. 2A). To
investigate the potential function of hAMSC-CM-induced circ-ABCB10
expression in HUVECs, three siRNAs, si-1, si-2 and si-3, were
designed to specifically target the junction site of circ-ABCB10 in
HUVECs. The present study examined the silencing efficiency of
these three siRNAs and found si-2 had the best silencing capacity.
Therefore, si-2 was used in further investigations (Fig. 2B). As expected, both scratch test and
transwell assays in HUVECs treated with hAMSC-CM revealed a reduced
migration in the si-2 group compared with the NC group (Fig. 2C and D). A tube formation assay was performed to
investigate the impact on angiogenesis caused by silencing of
circ-ABCB10 in HUVECs incubated with hAMSC-CM. Similarly, it was
found that the si-2 group showed a significantly decreased total
tube length compared with the NC group (Fig. 2E).
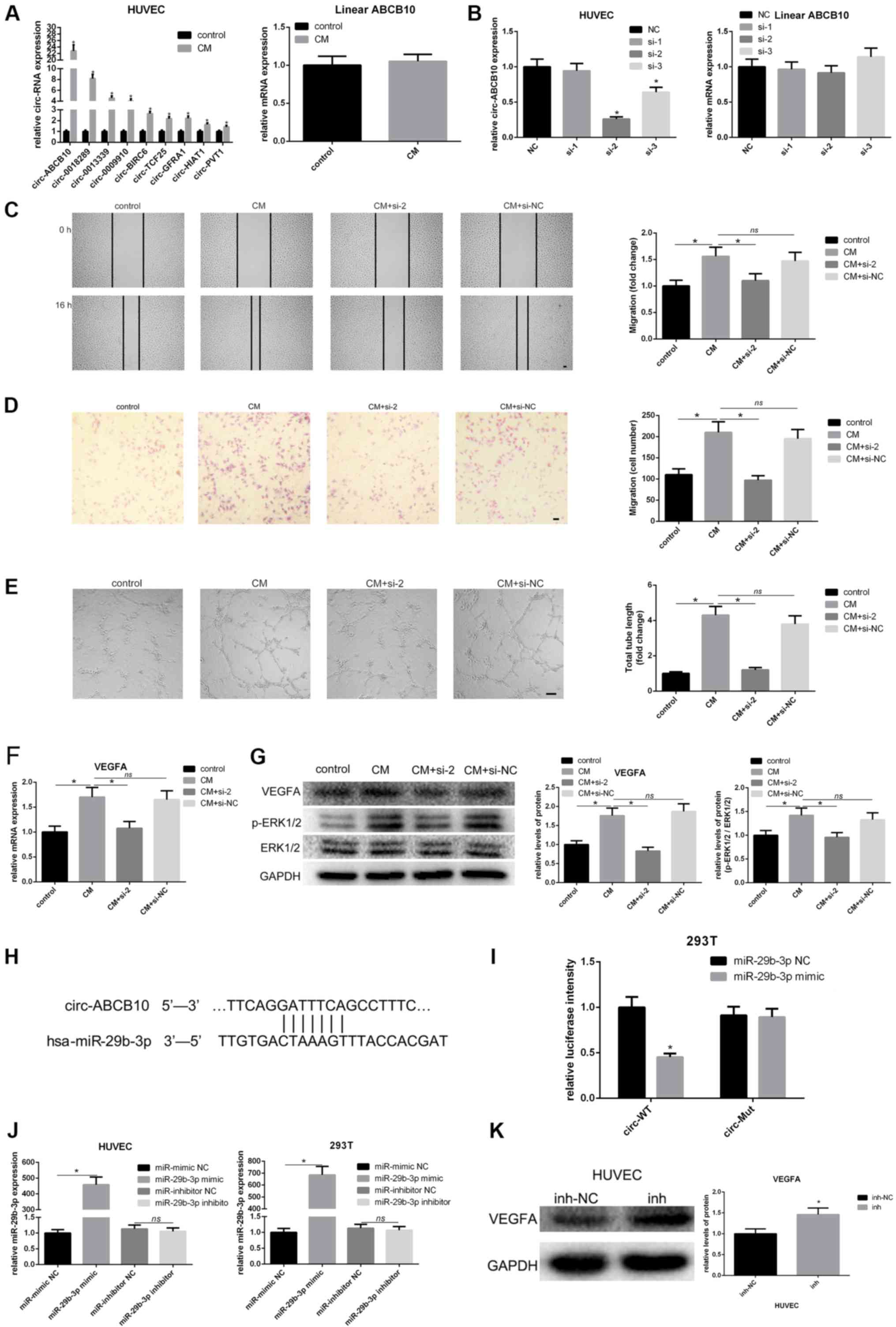 | Figure 2hAMSC-CM-induces loss of function of
circ-ABCB10 in HUVECs. (A) Changes in levels of circular RNAs that
may be involved in angiogenesis in HUVECs were measured by RT-qPCR.
circ-ABCB10 expression showed the highest increase, while linear
ABCB10 remained unchanged. (B) Specific siRNAs targeting
circ-ABCB10 were transfected into HUVECs, and the efficiency and
expression of linear ABCB10 were measured by RT-qPCR. (C) Scratch
test and (D) Transwell assays showed the effect of downregulation
of circ-ABCB10 on HUVEC migration induced by hAMSC-CM. Scale bar,
100 µm. (E) Tube formation assay results showed that the effect of
circ-ABCB10 downregulation on HUVEC angiogenesis was induced by
hAMSC-CM. Scale bar, 200 µm. (F) RT-qPCR results showed changes in
VEGFA mRNA expression levels in HUVECs induced by hAMSC-CM when
circ-ABCB10 was downregulated (G) Western blotting results showed
the corresponding changes in protein expression levels of VEGFA and
p-ERK1/2. (H) Potential miR-29b-3p binding site in the 5' to 3'
sequence of circ-ABCB10. (I) Dual-luciferase assay. HEK293T cells
were co-transfected with hsa-miR-29b-3p or miR-NC and plasmid with
wild-type or mutant circ-ABCB10. (J) The efficiency of transfection
of miR-29b-3p. (K) The expression of VEGFA potentially targeted by
miR-29b-3p in HUVEC transfected with inhibitor-NC or miR-29b-3p
inhibitor was detected by western blot. *P<0.05 vs.
control or NC, unless otherwise indicated. RT-qPCR, reverse
transcription-quantitative PCR; miR, microRNA; HUVECs, human
umbilical vein endothelial cells; hAMSCs, human amnion-derived
mesenchymal stem cells; CM, conditioned medium; VEGF, vascular
endothelial growth factor; p-, phosphorylated; MSCs, mesenchymal
stem cells; WT, wild-type; Mut, mutant; circ, circular RNA; NS, not
significant. |
The present study investigated whether circ-ABCB10
positively regulated VEGFA. RT-qPCR and western blotting results
suggested that silencing of circ-ABCB10 significantly reduced the
expression of VEGFA at both the mRNA and protein level in
hAMSC-CM-treated HUVECs. In addition, activation of p-ERK1/2 showed
the same effect (Fig. 2F and
G). There might be a potential
association between circ-ABCB10 and miR-29b-3p, which matched the
hypothesis of ceRNAs predicted by the TargetScan and miRanda
databases (Fig. 2H). A luciferase
assay was performed to investigate whether circ-ABCB10 could sponge
miR-29b-3p in vitro, and it was demonstrated that luciferase
intensity was significantly decreased when miR-29b-3p mimic was
co-transfected with WT circ-ABCB10 (Fig.
2I). In addition, the efficiency of transfection of miR-29b-3p
mimic was detected (Fig. 2J). In
order to confirm the transfection efficiency of miR-29b-3p mimic
and inhibitor, qPCR was performed. As shown in Fig. 2J, miR-29b-3p mimic significantly
increased miR-29b-3p expression, while miR-29b-3p inhibitor had no
effect. Several studies have shown that the miRNA inhibitor, which
binds to the mature miRNA and blocks its function in RNA-induced
silencing complex (RISC), may not directly degrade the target miRNA
(28-30).
Functional experiments and detection of target genes are still
reliable to evaluate the role of miRNA inhibitor (28-30).
Notably, miR-29b-3p has been reported to target VEGF gene
and inhibit its expression (31).
Furthermore, it has been directly confirmed that miR-29b-3p could
bind the mRNA of VEGFA in RISC by a recent study (32). Thus, the protein expression of VEGFA
was determined by western blotting, following transfection with
miR-29b-3p inhibitor. The results showed the miR-29b-3p inhibitor
significantly increased the VEGFA expression (Fig. 2K). Thus, the
circ-ABCB10/miR-29b-3p/VEGFA axis may be involved in the
pro-angiogenic effects of hAMSC-CM on HUVECs.
circ-ABCB10/miR-29b-3p/VEGFA axis may
be involved in the pro-angiogenic effects of hAMSC-CM on
HUVECs
To investigate whether inhibiting miR-29b-3p can
attenuate the effect caused by silencing circ-ABCB10 in
hAMSC-CM-treated HUVECs, a series of functional experiments were
performed. Wound healing and Transwell assays results suggested
that co-transfection of si-2 and miR-29b-3p inhibitor significantly
reversed the inhibition of migration caused by knockdown of
circ-ABCB10 (Fig. 3A and B). Tube-formation assay results identified
that the suppression of angiogenesis in hAMSC-CM-treated HUVECs,
resulting from knockdown of circ-ABCB10, was rescued by
co-transfection of si-2 and miR-29b-3p inhibitor (Fig. 3C). To understand the association
between circ-ABCB10, miR-29b-3p and VEGFA, the present study
investigated whether reduced expression of VEGFA caused by
knockdown of circ-ABCB10 could be rescued by inhibiting miR-29b-3p
in HUVECs. RT-qPCR and western blotting results suggested that the
reduction of VEGFA caused by knockdown of circ-ABCB10 could be
inhibited by co-transfecting si-2 and miR-29b-3p inhibitor. In
addition, the activation of p-ERK1/2 showed a similar effect
(Fig. 3D and E).
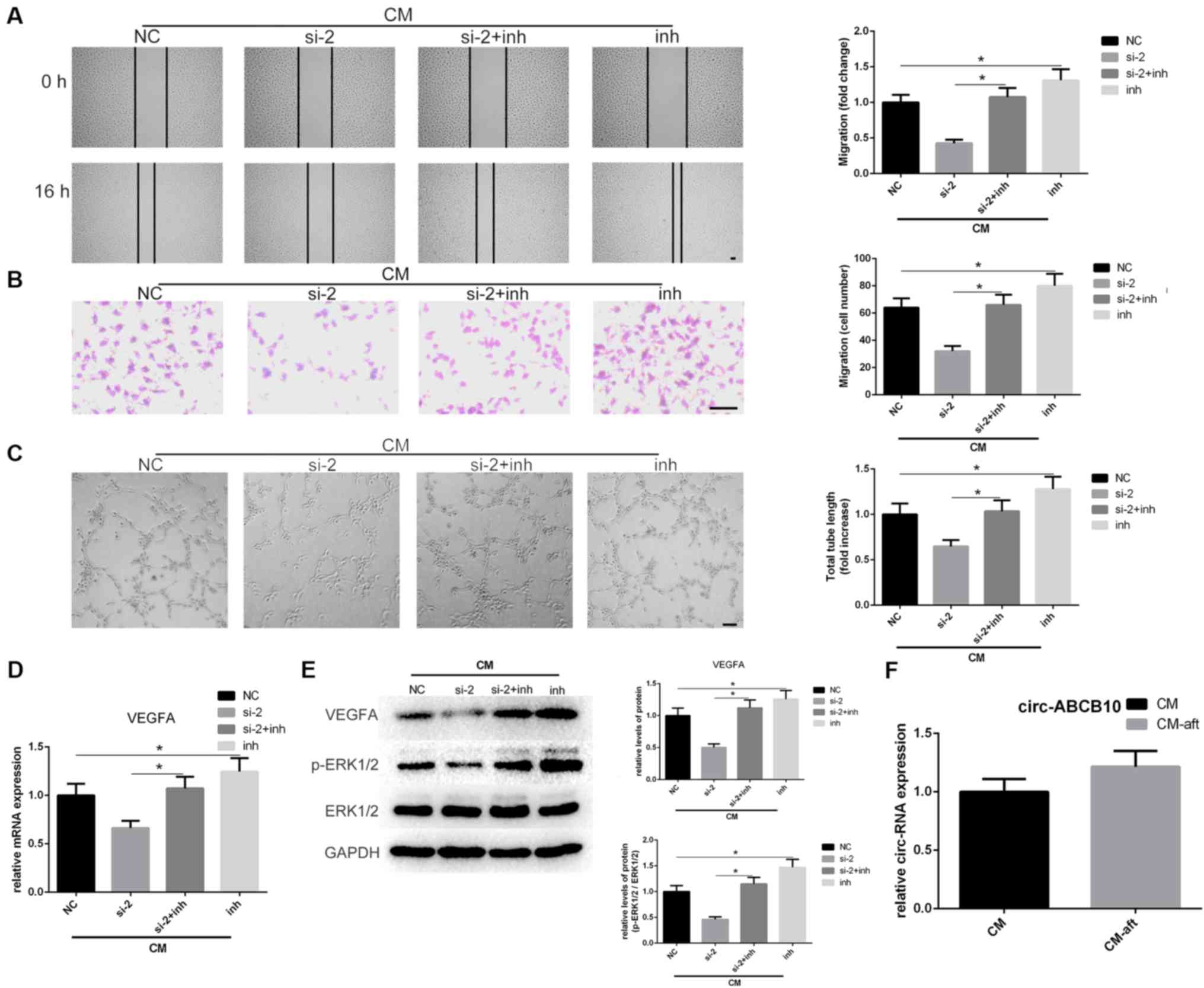 | Figure 3Loss of miR-29b-3p rescues the
effects caused by circ-ABCB10 downregulation in hAMSC-CM-treated
HUVECs. (A) Scratch test and (B) Transwell assays results indicated
that circ-ABCB10 downregulation on hAMSC-CM-induced HUVEC migration
was largely rescued by a miR-29b-3p inhibitor. Scale bar, 100 µm.
(C) Tube-formation assay results showed the effect of circ-ABCB10
downregulation on hAMSC-CM-induced HUVEC angiogenesis was
significantly rescued by the miR-29b-3p inhibitor. Scale bar, 200
µm. (D) VEGFA mRNA expression levels were recovered when HUVECs
were co-transfected with miR-29b-3p inhibitor. (E) VEGFA protein
expression levels and ERK1/2 activation were rescued by miR-29b-3p
inhibitor. (F) circ-ABCB10 expression levels in hAMSC-CM were
unchanged after incubation with HUVECs for 24 h.
*P<0.05. HUVECs, human umbilical vein endothelial
cells; hAMSCs, human amnion-derived mesenchymal stem cells; CM,
conditioned medium; CM, hAMSC-CM; CM-aft, hAMSC-CM incubated with
HUVECs for 24 h; VEGF, vascular endothelial growth factor; p-,
phosphorylated; MSCs, mesenchymal stem cells; siRNA, small
interfering RNA; circ, circular RNA; inh, inhibitor. |
The present study investigated whether increased
circ-ABCB10 in HUVECs originated from hAMSC-CM. The expression
levels of circ-ABCB10 in hAMSC-CM and hAMSC-CM incubated with
HUVECs for 24 h, referred to as hAMSC-CM-aft, were analyzed. It was
demonstrated that unlike the changes of expression of VEGFA,
circ-ABCB10 expression levels between hAMSC-CM and hAMSC-CM-aft
exhibited no statistically significant change (Fig. 3F). Thus, the present results
suggested that hAMSC-CM may partially promote HUVEC migration and
tube formation via the circ-ABCB10/miR-29b-3p/VEGFA axis.
Discussion
Angiogenesis is important in the process of
therapeutic bone regeneration, as bone healing or remodeling
requires oxygen and nutrients from blood vessels to maintain normal
metabolism (2). Therapeutic
angiogenesis induced by stem cells has been considered to be a
potential solution to provide a more beneficial vascular niche
during bone regeneration (33). Kim
et al (12), found that
hAMSCs exhibit angiogenesis, but the details of the molecular
mechanisms involved in the pro-angiogenic process remain unknown.
Previous studies have focused on the significant effect on
angiogenesis in various stem cell types (5-8,34).
MSCs can secrete multiple types of growth factors and cytokines
such as VEGF, insulin-like growth factor 1 and hepatocyte growth
factor in a paracrine manner (7,35). A
previous study revealed promising findings regarding angiogenesis
in a 3D contact co-culturing model of hAMSCs with HUVECs, which may
be involved in hAMSC secretion of matrix metallopeptidases
(15). The present study
hypothesized that hAMSC-CM may promote angiogenesis of HUVECs in a
non-contact co-culture system in a similar paracrine manner. The
present results suggested a pro-angiogenic capacity of CM from
hAMSCs on HUVECs, using a non-contact co-culture model in
vitro. The present study investigated the molecular mechanism
involved in this process. It was found that hAMSC-CM significantly
promoted tube formation and migration of HUVECs with significant
increases in circ-ABCB10 and VEGFA expression levels. In addition,
the present results suggested silencing circ-ABCB10 in HUVECs in
the non-contact co-culture model led to weakened induction of
angiogenesis and reduced expression of VEGFA, following hAMSC-CM
treatment of HUVECs. It was demonstrated that the transfection of
miR-29b-3p inhibitor could largely rescue the weakened angiogenesis
induced by hAMSC-CM treatment of HUVECs and decreased the
expression of VEGFA caused by the knockdown of circ-ABCB10.
Stem cell-based therapy is an important strategy in
the treatment of various diseases, especially angiogenesis-boosting
therapies (5,6). The present results suggested increased
proliferation, migration and tube formation of HUVECs following
hAMSC-CM treatment. VEGFA expression level was increased in HUVECs
cultured with hAMSC-CM. The present study examined whether hAMSC-CM
delivered hAMSC-secreted VEGFA to HUVECs. ELISA assay results
suggested that the protein expression level of VEGFA in hAMSC-CM
was lower compared with hAMSC-CM incubated with HUVECs for 24 h.
This effect may be attributed to a different manner of secretion in
diverse stem cell types (35) or may
be caused by mediation of upstream molecules regulating VEGFA
signaling.
circRNAs are not just byproducts of transcription,
but can also mediate cell proliferation, migration, invasion and
apoptosis in both disease and development (16). circRNAs may have an important role in
angiogenesis by acting as a miRNA sponge (36). In a recent study, circ-SHKBP1 was
found to regulate the angiogenesis of glioma-exposed endothelial
cells via interactions with miR-379 and miR-544a (19). Another previous study indicated that
has_circ_0054633 protects HUVECs from the damage caused by high
glucose by sponging miR-218 and upregulating the expression of
roundabout homolog 1 and heme oxygenase-1(36). Therefore, the present study selected
a number of circRNAs that may be involved in angiogenesis and
detected expression changes in co-cultured HUVECs. Among them,
circ-ABCB10 showed the greatest increase in expression and was
predicted to sponge up miR-29b-3p targeting VEGFA. The present
luciferase reporter assays and loss-of-function and rescue assay
results demonstrated thatcirc-ABCB10 may increase the angiogenesis
of HUVECs via the miR-29b-3p/VEGFA axis. The present results
supported the hypothesis that hAMSC-CM may promote the angiogenesis
of HUVECs by regulating molecules upstream of the VEGFA
pathway.
Several circRNAs involved in biofunctions including
proliferation and angiogenesis were found in cancer exosomes
(37,38). A recent study indicated exosomal
circ-IARS secreted from pancreatic cancer cells could mediate the
permeability of HUVEC monolayers to enhance tumor metastasis
(39). The present study
investigated whether circ-ABCB10, which enhanced the angiogenesis
of HUVECs, originates from exosomes released by hAMSCs. We thereby
measured circ-ABCB10 expression levels in hAMSC-CM and hAMSC-CM
incubated with HUVECs for 24 h. However, the expression levels of
circ-ABCB10 in hAMSC-CM showed no statistically significant
difference in levels in hAMSC-CM incubated with HUVECs for 24 h.
Hence, it is possible that the increased expression of circ-ABCB10
was indirectly induced by hAMSC-CM, possibly by the promotion of
circ-ABCB10 transcription rather than direct delivery by exosomes
in hAMSC-CM. Whilst extensive study on the biofunction of circRNAs
has been undertaken, the molecular mechanism associated with the
transcription of circRNAs remains elusive. Previous studies focused
on transcription factors regulating the transcription of circRNAs
(40,41). Liang et al (40), found that hypoxia-inducible factor
1-α regulates the hypoxia-induced expression of circ-DENND4C in
breast cancer cells. Another study showed that Twist1 promotes the
expression of circular cullin 2 during epithelial-mesenchymal
transition (41). Therefore,
hAMSC-CM may contain secretory proteins that can interact with
transcription factors to mediate the transcription of circ-ABCB10
in HUVECs. However, further studies are required to investigate
this effect.
In summary, the present results suggested hAMSC-CM
enhanced migration and tube formation of HUVECs via the
circ-ABCB10/miR-29b-3p/VEGFA pathway. Thus, the present results
could help to elucidate the molecular mechanism of hAMSC-induced
angiogenesis.
Supplementary Material
Table SI. Sequences of primers,
siRNAs, miR.29b.3p mimic, miR.29b.3p inhibitor, circ.WT and
circ.Mut.
Acknowledgements
Not applicable.
Funding
This work was supported by The National Natural
Science Foundation of China (grant no. 81670966) and A Project
Funded by the Priority Academic Program Development of Jiangsu
Higher Education Institutions (PAPD, 2018-87).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZT, MS and NC contributed to the study design and
conception. ZT, JT, ZY and QZ performed the experiments. ZT and XY
analyzed the data. ZT wrote the manuscript. NC and ZY revised the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Study protocols were approved by The Ethics
Committee of the School of Stomatology, Nanjing Medical University
(approval no. PJ2013-037-001). Written informed consent was
obtained from each participant.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hu N, Jiang D, Huang E, Liu X, Li R, Liang
X, Kim SH, Chen X, Gao JL, Zhang H, et al: BMP9-regulated
angiogenic signalling plays an important role in the osteogenic
differentiation of mesenchymal progenitor cells. J Cell Sci.
126:532–541. 2013.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Kusumbe AP, Ramasamy SK and Adams RH:
Coupling of angiogenesis and osteogenesis by a specific vessel
subtype in bone. Nature. 507:323–328. 2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Ramasamy SK, Kusumbe AP, Wang L and Adams
RH: Endothelial notch activity promotes angiogenesis and
osteogenesis in bone. Nature. 507:376–380. 2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Kanczler JM and Oreffo RO: Osteogenesis
and angiogenesis: The potential for engineering bone. Eur Cell
Mater. 15:100–114. 2008.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Daley GQ and Scadden DT: Prospects for
stem cell-based therapy. Cell. 132:544–548. 2008.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Satija NK, Singh VK, Verma YK, Gupta P,
Sharma S, Afrin F, Sharma M, Sharma P, Tripathi RP and Gurudutta
GU: Mesenchymal stem cell-based therapy: A new paradigm in
regenerative medicine. J Cell Mol Med. 13:4385–4402.
2009.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kinnaird T, Stabile E, Burnett MS, Lee CW,
Barr S, Fuchs S and Epstein SE: Marrow-derived stromal cells
express genes encoding a broad spectrum of arteriogenic cytokines
and promote in vitro and in vivo arteriogenesis through paracrine
mechanisms. Circ Res. 94:678–685. 2004.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zuo S, Jones WK, Li H, He Z, Pasha Z, Yang
Y, Wang Y, Fan GC, Ashraf M and Xu M: Paracrine effect of
wnt11-overexpressing mesenchymal stem cells on ischemic injury.
Stem Cells Dev. 21:598–608. 2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Soncini M, Vertua E, Gibelli L, Zorzi F,
Denegri M, Albertini A, Wengler GS and Parolini O: Isolation and
characterization of mesenchymal cells from human fetal membranes. J
Tissue Eng Regen Med. 1:296–305. 2007.PubMed/NCBI View
Article : Google Scholar
|
|
10
|
Santhagunam A, Dos Santos F, Madeira C,
Salgueiro JB and Cabral JM: Isolation and ex vivo, expansion of
synovial mesenchymal stromal cells for cartilage repair.
Cytotherapy. 16:440–453. 2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Ilancheran S, Moodley Y and Manuelpillai
U: Human fetal membranes: A source of stem cells for tissue
regeneration and repair? Placenta. 30:2–10. 2009.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kim SW, Zhang HZ, Kim CE, An HS, Kim JM
and Kim MH: Amniotic mesenchymal stem cells have robust angiogenic
properties and are effective in treating hindlimb ischaemia.
Cardiovasc Res. 93:525–534. 2012.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Parolini O, Alviano F, Bagnara GP, Bilic
G, Bühring HJ, Evangelista M, Hennerbichler S, Liu B, Magatti M,
Mao N, et al: Concise review: Isolation and characterization of
cells from human term placenta: Outcome of the first international
workshop on placenta derived stem cells. Stem Cells. 26:300–311.
2008.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Leyva-Leyva M, Barrera L, López-Camarillo
C, Arriaga-Pizano L, Orozco-Hoyuela G, Carrillo-Casas EM,
Calderón-Pérez J, López-Díaz A, Hernandez-Aguilar F,
González-Ramírez R, et al: Characterization of mesenchymal stem
cell subpopulations from human amniotic membrane with dissimilar
osteoblastic potential. Stem Cells Dev. 22:1275–1287.
2012.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Jiang F, Ma J, Liang Y, Niu Y, Chen N and
Shen M: Amniotic mesenchymal stem cells can enhance angiogenic
capacity via MMPs in vitro and in vivo. Biomed Res Int.
2015(324014)2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Yao T, Chen Q, Fu L and Guo J: Circular
RNAs: Biogenesis, properties, roles, and their relationships with
liver diseases. Hepatol Res. 47:497–504. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Hou LD and Zhang J: Circular RNAs: An
emerging type of RNA in cancer. Int J Immunopathol Pharmacol.
30:1–6. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Tang W, Ji M, He G, Yang L, Niu Z, Jian M,
Wei Y, Ren L and Xu J: Silencing CDR1as inhibits colorectal cancer
progression through regulating microrna-7. Onco Targets Ther.
10:2045–2056. 2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
He Q, Zhao L, Liu Y, Liu X, Zheng J, Yu H,
Cai H, Ma J, Liu L, Wang P, et al: Circ-SHKBP1 regulates the
angiogenesis of u87 glioma-exposed endothelial cells through
miR-544a/FOXP1 and miR-379/FOXP2 pathways. Mol Ther Nucleic Acids.
10:331–348. 2008.
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Fierro FA, O'Neal AJ, Beegle JR, Chávez
MN, Peavy TR, Isseroff RR and Egaña JT: Hypoxic pre-conditioning
increases the infiltration of endothelial cells into scaffolds for
dermal regeneration pre-seeded with mesenchymal stem cells. Front
Cell Dev Biol. 3(68)2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Liang HF, Zhang XZ, Liu BG, Jia GT and Li
WL: Circular RNA circ-ABCB10 promotes breast cancer proliferation
and progression through sponging miR-1271. Am J Cancer Res.
7:1566–1576. 2017.PubMed/NCBI
|
|
23
|
Gao YL, Zhang MY, Xu B, Han LJ, Lan SF,
Chen J, Dong YJ and Cao LL: Circular RNA expression profiles reveal
that hsa_circ_0018289 is up-regulated in cervical cancer and
promotes the tumorigenesis. Oncotarget. 8:86625–86633.
2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Deng N, Li L, Gao J, Zhou J, Wang Y, Wang
C and Liu Y: Hsa_circ_0009910 promotes carcinogenesis by promoting
the expression of miR-449a target IL6R in osteosarcoma. Biochem
Biophys Res Commun. 495:189–196. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen
B, Luo Y, Lyu D, Li Y, Shi G, et al: Circular RNA profiling reveals
an abundant circHIPK3 that regulates cell growth by sponging
multiple miRNAs. Nat Commun. 7(11215)2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhong Z, Lv M and Chen J: Screening
differential circular RNA expression profiles reveals the
regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in
bladder carcinoma. Sci Rep. 6(30919)2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
He Z, Ruan X, Liu X, Zheng J, Liu Y, Liu
L, Ma J, Shao L, Wang D, Shen S, et al:
FUS/circ_002136/miR-138-5p/SOX13 feedback loop regulates
angiogenesis in glioma. J Exp Clin Cancer Res.
38(65)2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Davis S, Lollo B, Freier S and Esau C:
Improved targeting of miRNA with antisense oligonucleotides.
Nucleic Acids Res. 34:2294–2304. 2006.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Thomson DW, Bracken CP, Szubert JM and
Goodall GJ: On measuring miRNAs after transient transfection of
mimics or antisense inhibitors. PLoS One. 8(e55214)2013.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Hutvágner G, Simard MJ, Mello CC and
Zamore PD: Sequence-specific inhibition of small RNA function. PLoS
Biol. 2(E98)2004.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Tang W, Guo J, Gu R, Lei B, Ding X, Ma J
and Xu G: MicroRNA-29b-3p inhibits cell proliferation and
angiogenesis by targeting VEGFA and PDGFB in retinal microvascular
endothelial cells. Mol Vis. 26:64–75. 2020.PubMed/NCBI
|
|
32
|
Zhao X, Liu Y, Li Z, Zheng S, Wang Z, Li
W, Bi Z, Li L, Jiang Y, Luo Y, et al: Linc00511 acts as a competing
endogenous RNA to regulate VEGFA expression through sponging
hsa-miR-29b-3p in pancreatic ductal adenocarcinoma. J Cell Mol Med.
22:655–667. 2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ren L, Ma D, Liu B, Li J, Chen J, Yang D
and Gao P: Preparation of three-dimensional vascularized MSC cell
sheet constructs for tissue regeneration. Biomed Res Int.
2014(301279)2014.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Edwards SS, Zavala G, Prieto CP, Elliott
M, Martínez S, Egaña JT, Egaña JT, Bono MR and Palma V: Functional
analysis reveals angiogenic potential of human mesenchymal stem
cells from Wharton's jelly in dermal regeneration. Angiogenesis.
17:851–866. 2014.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ratajczak MZ, Kucia M, Jadczyk T, Greco
NJ, Wojakowski W, Tendera M and Ratajczak J: Pivotal role of
paracrine effects in stem cell therapies in regenerative medicine:
Can we translate stem cell-secreted paracrine factors and
microvesicles into better therapeutic strategies? Leukemia.
26:1166–1173. 2012.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Pan L, Lian W, Zhang X, Han S, Cao C, Li X
and Li M: Human circular RNA-0054633 regulates high glucose-induced
vascular endothelial cell dysfunction through the
microrna-218/roundabout 1 and microrna-218/heme oxygenase-1 axes.
Int J Mol Med. 42:597–606. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Ruivo CF, Adem B, Silva M and Melo SA: The
biology of cancer exosomes: Insights and new perspectives. Cancer
Res. 77:6480–6488. 2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Steinbichler TB, Dudás J, Riechelmann H
and Skvortsova II: The role of exosomes in cancer metastasis. Semin
Cancer Biol. 44:170–181. 2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Li J, Li Z, Jiang P, Peng M, Zhang X, Chen
K, Liu H, Bi H, Liu X and Li X: Circular RNA IARS (circ-IARS)
secreted by pancreatic cancer cells and located within exosomes
regulates endothelial monolayer permeability to promote tumour
metastasis. J Exp Clin Cancer Res. 37(177)2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Liang G, Liu Z, Tan L, Su AN, Jiang WG and
Gong C: HIF1α-associated circDENND4C promotes proliferation of
breast cancer cells in hypoxic environment. Anticancer Res.
37:4337–4343. 2017.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Meng J, Chen S, Han JX, Qian B, Wang XR,
Zhong WL, Qin Y, Zhang H, Gao WF, Lei YY, et al: Twist1 regulates
vimentin through Cul2 circular RNA to promote EMT in hepatocellular
carcinoma. Cancer Res. 78:4150–4162. 2018.PubMed/NCBI View Article : Google Scholar
|














