Introduction
Graves' Ophthalmopathy (GO), which is also known as
thyroid eye disease, is an autoimmune disease that is caused by
thyroid dysfunction and seriously affects the quality of life of
patients suffering from it (1). The
principal clinical manifestations of GO include ocular protrusion,
eyelid contraction, exposure keratitis, restricted eye movement and
oppressive optic neuropathy (2).
The pathogenesis of GO has not yet been fully elucidated, is
complex and has been indicated to involve both genetic and
environmental factors (3). It has
been reported that T cell-induced inflammation, glycosaminoglycan
(GAG) aggregation and fibrosis originating from orbital fibroblasts
are the primary pathological mechanisms of GO (4). Histological evidence has revealed that
GO was consistently accompanied by the infiltration of lymphocytes
in the posterior globules, the accumulation of GAG [primarily
hyaluronic-acid (HA)] and collagen deposition, which resulted in
the development of connective tissue hyperplasia and fibrosis
(5,6). These pathological alterations have
been reported to be associated with orbital fibroblasts (7).
In the pathogenesis of GO, orbital fibroblasts have
been reported to produce large amounts of HA and collagen I
(8). HA is the principal component
of GAG and remains anchored to the cell surface after its synthesis
via binding to either hyaluronan synthase or other surface
receptors, while an amount of HA is also cleaved by hyaluronidase
and is released in the extracellular matrix (9). Owing to the potent hydrophilic nature
of HA, its accumulation has been indicated to accelerate the
expansion of orbital tissues (10).
Collagen I is considered to be a marker of fibrosis (11). Fibrocytes, which express CD34 and
C-X-C chemokine receptor type 4, have been reported to produce
collagen I and infiltrate tissues in response to multiple
chemokines, including C-X-C motif chemokine 12(12), which may result in fibrosis in
orbital tissues.
MicroRNAs (miRNAs/miRs) are non-coding,
single-stranded RNA molecules that are ~22 nucleotides in length,
which have been indicated to regulate gene expression at the
post-transcriptional level (13).
miRNAs have been reported to serve an important role in the
pathogenesis of numerous diseases, including cancer and infectious
diseases (14), and it has been
suggested that they may serve as novel targets for disease
treatment (15). In recent years,
several studies have described the role of miRNAs in inflammatory
autoimmune diseases (16-18).
miR-146a has been reported to serve a role in a variety of
autoimmune diseases, including rheumatoid arthritis, systemic lupus
erythematosus and osteoarthritis (19). Previous studies have indicated that
miR-146a regulated the expression of inflammatory factors in GO
orbital fibroblasts (19,20). The current study aimed to
investigate the effect of miR-146a on the production of HA and
collagen I in GO orbital fibroblasts, and indicated that miR-146a
may be used as a novel target for the treatment of GO.
Materials and methods
Reagents
HA (cat. no. ml026554) and collagen I (cat. no.
ml029092) ELISA kits were purchased from Shanghai Enzyme-linked
Biotechnology Co., Ltd. Antibodies against vimentin (cat. no.
D220268), S100B (cat. no. D161409), myoglobin (cat. no. D222721),
keratin 17 (cat. no. D220232) and desmin (cat. no. D162991) were
purchased from Shanghai Shenggong, Biology Engineering Technology
Service, Ltd. A Histostain-SP kit (cat. no. SP-0023) was purchased
from BIOSS. TRIzol® reagent was supplied by Invitrogen,
Thermo Fisher Scientific, Inc. (cat. no. 15596026), Inc. miR-146a
mimics, miR-146a inhibitor, control mimics, control inhibitor and
Polybrene Infection Reagent were purchased from Shanghai GeneChem
Co., Ltd. Primers for miR-146a and U6 were purchased from Takara
Biotechnology Co., Ltd. The primer sequence for miR-146a was
withheld from the supplier (cat. no. MAQ472). Primers for HA and
collagen I, GAPDH were purchased from Shanghai Shenggong Biology
Engineering Technology Service, Ltd.
Patients
Patients were recruited according to the Bartley
diagnostic criteria and clinical activity scores (CAS) (21), where ≥4 points indicated active GO
lesions and <4 points indicate that GO is inactive. The orbital
connective tissue of patients treated at the First Affiliated
Hospital of Guangxi Medical University from January 2018 to October
2019 was obtained for the current study (Table I). A total of 6 patients with
inactive GO (CAS <4) with adipose tissue after orbital
decompression and 4 patients (non-GO patients) with eyeball removal
or upper lid blepharoplasty were included in the current study.
Patients who had previously used immunosuppressive agents and
exhibited other autoimmune diseases (including asthma, chronic
inflammation and HIV), a recent history of trauma or an active
infection within six months prior to surgery were excluded. Written
informed consent was obtained from all participants prior to
inclusion. The current study was approved by the Ethics Review
Committee of the First Affiliated Hospital of Guangxi Medical
University.
 | Table IClinical information of the patients
included in the current study. |
Table I
Clinical information of the patients
included in the current study.
| | Age (years) | Sex | Smoker | Duration of GO
(years) | CAS | Surgical
treatment |
|---|
| A, Patients with
GO |
| 1 | 61 | Female | N | 0.5 | 0/7 | Decompression |
| 2 | 48 | Female | N | 0.5 | 1/7 | Decompression |
| 3 | 52 | Female | N | 1 | 1/7 | Decompression |
| 4 | 49 | Male | Y | 1 | 3/7 | Decompression |
| 5 | 57 | Male | N | 20 | 1/7 | Decompression |
| 6 | 63 | Male | N | 0.6 | 1/7 | Decompression |
| B, Non-GO control
subjects |
| | Age (years) | Sex | Smoker | Duration of GO
(years) | CAS | Surgical
treatment |
| 1 | 35 | Female | N | n/a | n/a | Eye evisceration |
| 2 | 21 | Male | N | n/a | n/a | Eye evisceration |
| 3 | 50 | Male | Y | n/a | n/a | Eye
evisceration |
| 4 | 33 | Female | N | n/a | n/a | Upper lid
blepharoplasty |
Cell culture and infection
Orbital fat or connective tissues from patients with
GO and non-GO patients were collected under aseptic conditions
during surgery (eyeball removal or upper lid blepharoplasty),
placed immediately into a 50 ml centrifuge tube containing DMEM
high glucose culture fluid (Gibco; Thermo Fisher Scientific, Inc.)
and 1% penicillin/streptomycin on ice. The tissue specimens were
washed three times with PBS under sterile conditions. The blood
vessels, which were visible on the tissue blocks, were removed
using sterile ophthalmic micro scissors. Following three washes
with PBS, the tissue was cut into pieces of ~0.5 mm3 and
transferred to a 15 ml centrifuge tube. Pancreatin digestion
solution containing EDTA (Dalian Meilun Biology Technology Co.,
Ltd.) was added and the centrifuge tube was incubated at 37˚C m for
15 min. Following the addition of DMEM containing 10% FBS (Shanghai
Shuangru Biotechnology Co., Ltd.) to terminate the digestion, the
tube was centrifuged at 25˚C for 15 min at 1,200 x g. The
suspension was subsequently filtered using a 200 Mesh stainless
steel filter (Beijing Solarbio Science & Technology Co., Ltd.)
to remove incompletely digested tissue pieces, and was centrifuged
at 1,200 x g for 15 min at room temperature. The cell pellet was
dissolved in DMEM containing 20% FBS and 1%
penicillin/streptomycin, seeded in cell culture flasks at a density
of 40,000 cells/cm2 and placed in an incubator at 37˚C
with 5% CO2. After 2-3 days, the cell culture medium was
replaced with fresh DMEM containing 10% FBS and 1%
penicillin/streptomycin. Immunohistochemical identification was
performed to determine whether the extracted cells were orbital
fibroblasts. The expression of HA and collagen I in orbital
fibroblasts of patients with GO and non-GO patients was detected
via reverse transcription-quantitative PCR (RT-qPCR). The
fibroblasts were seeded in a 6-well plate at a concentration of
5x104 cells/ml and infected with a lentivirus carrying
miR-146a mimics, miR-146a inhibitor, control mimics or control
inhibitor (each 20 nmol/l) for 12 h. Then, the cell culture medium
was replaced with fresh DMEM containing 10% FBS and 1%
penicillin/streptomycin. After 72 h, a green fluorescence signal
could be observed under a fluorescence microscope, which indicates
successful infection. Uninfected orbital fibroblasts were removed
using complete medium (DMEM containing 10% FBS and 1%
penicillin/streptomycin) containing puromycin (2 µg/ml) for 3-4
days. Then, replace the cell culture medium with fresh DMEM
containing 10% FBS and 1% penicillin/streptomycin and continue
culturing for 2-3 days. At this time, fibroblasts can be used for
subsequent experimentation. The morphology of the orbital
fibroblasts and the infection efficiency were observed using a
light microscope (magnification, x100) and an inverted fluorescence
phase contrast microscope (magnification, x100), respectively. The
expression of miR-146a in the orbital fibroblasts of each group was
detected via RT-qPCR, and the expression of HA and collagen I was
detected via RT-qPCR and ELISA. After 2-3 days, subsequent
experimentation was performed.
Immunohistochemistry (IHC)
IHC was employed to examine the expression of
vimentin, desmin, myoglobin, keratin 17 and S100B. A total of
3-5x104/ml primary orbital fibroblasts were seeded on
slides. After the cells were attached, the supernatant was
discarded, the cells were washed three times with PBS and
subsequently incubated with 4% paraformaldehyde (Solarbio, China)
at 4˚C overnight. The cell slides were washed 3x with PBS and
incubated with 3% H2O2 in deionized water for
10 min at room temperature. Following three washes with PBS, the
slides were blocked with whole goat serum included in the IHC kit
(cat. no. IHC001; Beijing Biosynthesis Biotechnology Co., Ltd.) was
used at 37˚C for 10-20 min. The blocking solution was removed, and
the primary antibodies (vimentin, desmin, myoglobin, keratin 17 and
S100B; each, 1:400) were added and incubated at 4˚C overnight.
After washing three times with PBS, the biotin-labeled goat
anti-rabbit IgG (included in the IHC kit) was added and incubated
at 37˚C for 30-60 min. The cell slides were washed three times with
PBS, and 3,3'-diaminobenzidine reagent was added for color
development. After 4-10 min of hematoxylin counterstaining at room
temperature, the excess dye solution was removed, and the slides
were washed with water for 5 min, dehydrated and sealed with a
neutral gum. A light microscope (magnification, x200) was used to
observe slides.
RT-qPCR
Orbital fibroblast total RNA from the miR-146a
mimics, miR-146a inhibitor, control mimics, control inhibitor
groups was extracted with TRIzol® reagent. The
concentration and quality of RNA were determined using an
ultra-differential photometer (NanoDrop™ 2000; Thermo Fisher
Scientific, Inc.). The RNA samples were reverse transcribed using
PrimeScript RT reagent kit with gDNA Eraser from Takara
Biotechnology Co., Ltd. (37˚C for 60 min; 85˚C for 5 min) and qPCR
was performed using SYBR Premix Ex Taq II from Takara Biotechnology
Co., Ltd. (one cycle of 95˚C for 10 sec, 95˚C for 5 sec and 60˚C
for 20 sec; and 40 cycles of 95˚C for 60 sec, 55˚C for 30 sec and
95˚C for 30 sec). The relative gene expression was normalized using
real-time quantitative PCR and the 2-ΔΔCq method
(22). The expression of mir-146a
was normalized to that of U6. The primer sequences for HA and
collagen I are presented in Table
II, the expression of which was normalized to GAPDH
expression.
| Table IIPrimer sequences used in reverse
transcription-quantitative PCR. |
Table II
Primer sequences used in reverse
transcription-quantitative PCR.
| Gene name | Forward primer | Reverse primer |
|---|
| HA |
5'-CACGTAACGCAATTGGTCTTGTCC-3' |
5'-CCAGTGCTCTGAAGGCTGTGTAC-3' |
| COL1A2 |
5'-CTGGACCTCCAGGTGTAAGC-3' |
5'-TGGCTGAGTCTCAAGTCACG-3' |
| GAPDH |
5'-GACATGCCGCCTGGAGAAAC-3' |
5'-AGCCCAGGATGCCCTTTAGT-3' |
| U6 |
5'-GGAACGATACAGAGAAGATTAGC-3' |
5’-TGGAACGCTTCACGAATTTGCG-3’ |
ELISA
Following infection, the culture supernatant of the
miR-146a mimics, miR-146a inhibitor, control mimics, control
inhibitor group was collected and centrifuged at 4˚C, for 10 min at
800 x g. The expression levels of collagen I and HA in the culture
supernatant of confluent orbital fibroblasts were detected in
triplicates using commercially available Human HA ELISA kits and
Human collagen I ELISA kits according to the manufacturer's
protocol.
Statistical analysis
All experiments were repeated three times and data
were expressed as the mean ± standard deviation. Statistical
analysis was performed using SPSS v22.0 (IBM Corp.). Statistical
differences between two groups were analyzed using the independent
Student's t-test, while multiple group comparisons were made using
one-way ANOVA followed by Fisher's least significant difference
method for pairwise comparisons. P<0.05 was considered to
indicate a statistically significant difference.
Results
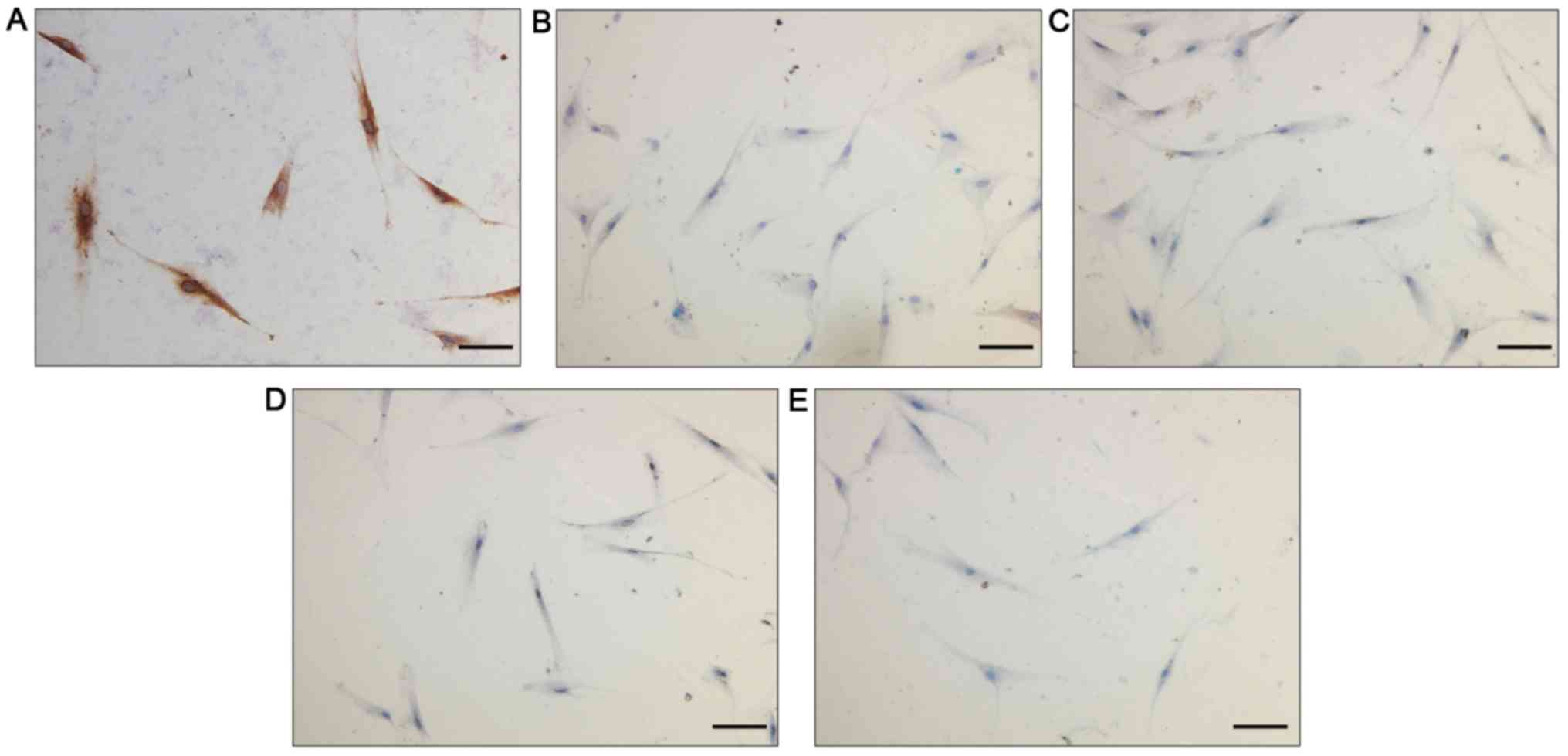
Identification of fibroblasts
As presented in Fig.
1, vimentin exhibited a positive expression in the cytoplasm of
orbital fibroblasts, indicating the mesenchymal origin of these
cells. By contrast, desmin, myoglobin, keratin 17 and S100B were
not expressed in these cells, excluding the possibility that these
cells were smooth muscle cells, striated muscle cells, skin cells,
nerve cells or skin melanocytes, which additionally indicated that
the extracted cell population was not contaminated with other cell
types. These results verified the identity of these cells as
fibroblasts.
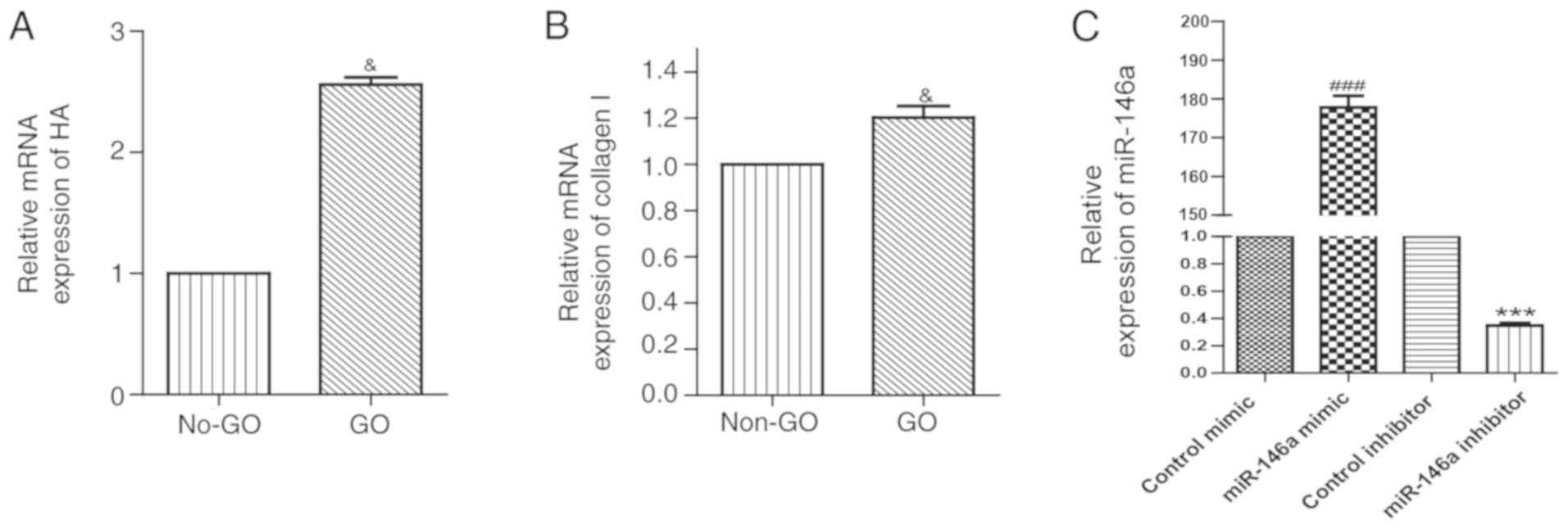
Expression of HA and collagen I in
orbital fibroblasts
The mRNA expression level of HA in orbital
fibroblasts of the GO group was approximately 2.6 folds higher
compared with the non-GO group (Fig.
2A). Similarly, orbital fibroblasts from the GO group exhibited
a higher expression level of collagen Ia2 mRNA compared with the
non-GO group (Fig. 2B).
Infection efficiency of miR-146a
mimics and inhibitor in orbital fibroblasts
RT-qPCR indicated that in the overexpression group,
the expression level of miR-146a was ~170-folds higher compared
with the control mimic, while miR-146a expression in the inhibition
group was decreased to ~30% of the control inhibitor expression
level (Fig. 2C) suggesting that
overexpression and inhibition of miR-146a in orbital fibroblasts
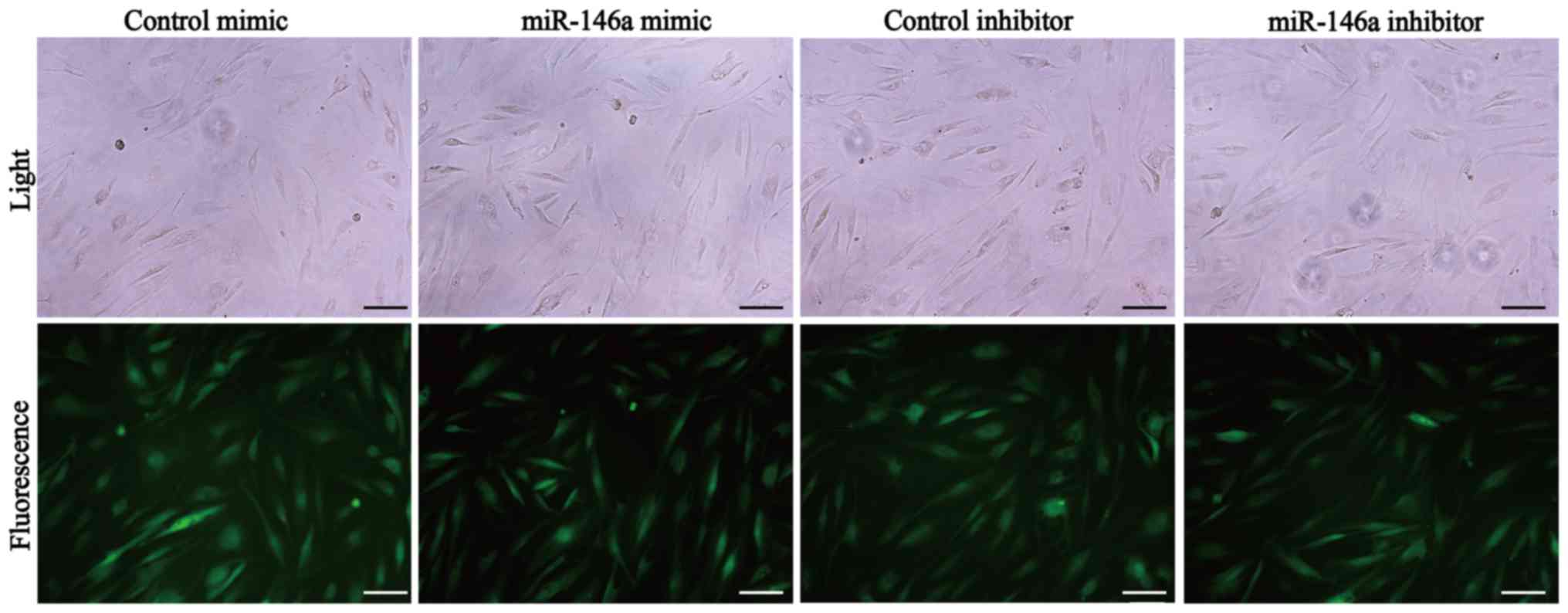
were efficient. The morphology of orbital fibroblasts following
infection was long fusiform or triangular, which was similar to the
morphology of these cells before infection (Fig. 3). On the other hand, a green
fluorescence signal was observed in >80% (an approximation as
observed under fluorescence microscopy) of fibroblasts post
infection, which indicated a high level of infection efficiency
(Fig. 3).
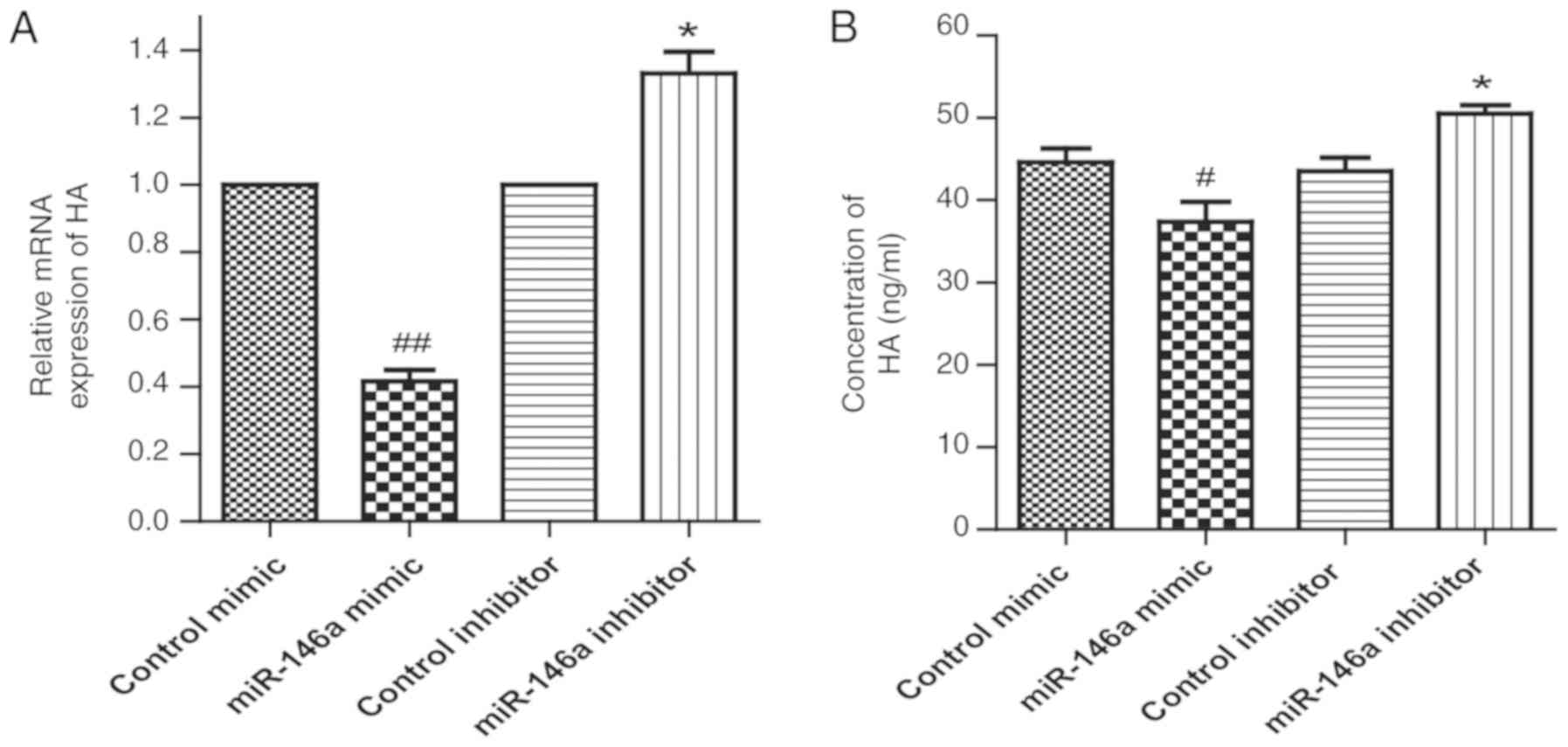
Effect of miR-146a on the expression
of HA in GO orbital fibroblasts
As demonstrated in Fig.
4A, the mRNA level of HA in the miR-146a overexpression group
was lower in comparison to control mimic, while the HA mRNA level
in the miR-146a inhibitor group was higher compared with the
control inhibitor. ELISA also indicated that overexpression of
miR-146a inhibited HA secretion, while inhibition of miR-146a
increased HA secretion in GO orbital fibroblasts compared with the
respective control group (Fig.
4B).
Effect of miR-146a on the expression
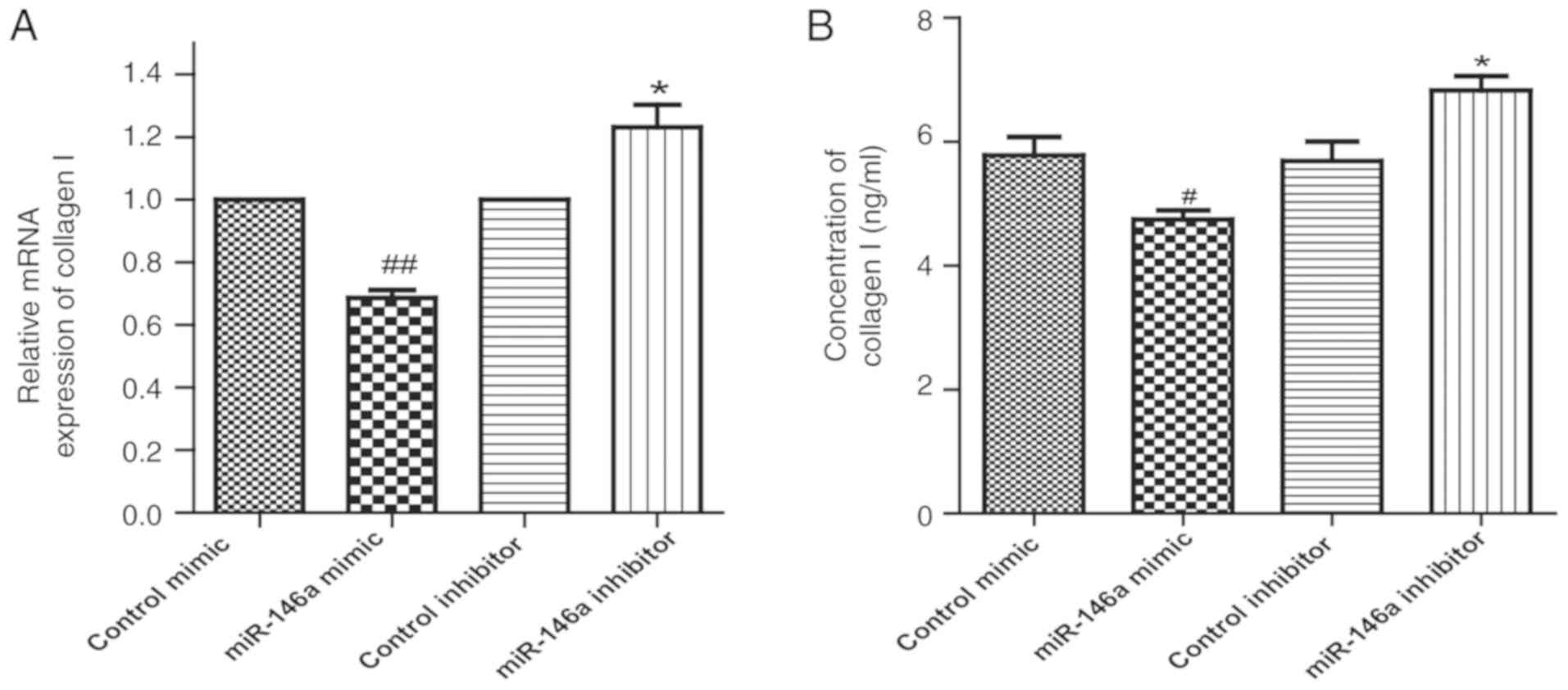
of collagen I in GO orbital fibroblasts
The mRNA level of collagen I in the miR-146a
overexpression group was lower compared with control mimic, while
the level of collagen I mRNA in the miR-146a inhibitor group was
higher compared with control inhibitor (Fig. 5A). Consistently, ELISA results also
indicated that overexpression of miR-146a inhibited, and inhibition
of miR-146a increased the secretion of collagen I in GO orbital
fibroblasts compared with the respective control group (Fig. 5B).
Discussion
In the current study, it was identified via IHC
staining that the isolated primary cells were orbital fibroblasts,
and a culture model of orbital fibroblasts was effectively
generated. The results of the present study indicated that the mRNA
levels of HA and collagen I in orbital fibroblasts from the GO
cohort were higher compared with the non-GO cohort. Furthermore,
miR-146a was efficiently overexpressed or inhibited in orbital
fibroblasts. ELISA and RT-qPCR results revealed that the production
of collagen I and HA was significantly reduced following
overexpression of miR-146a, while inhibition of miR-146a
significantly increased the production of collagen I and HA
compared with the control group.
GO is a common eyelid disorder that seriously
affects the appearance and visual function of patients, while
severe GO may also result in blindness (23). GO has been indicated to be initiated
with an autoimmune inflammatory response, HA synthesis and collagen
deposition, resulting in connective tissue hyperplasia and fibrosis
(24). Orbital fibroblasts have
been revealed to be an important pathogenetic factor of GO,
contributing to inflammation, lipogenesis, HA secretion and
fibrosis in GO (24). HA is the
principal component of GAG aggregation and exhibits high
hydrophilicity binding to a large amount of water, thereby
resulting in an abnormal eyelid tissue and extraocular muscle
interstitial edema (25). The
increase in HA production and collagen deposition has been
indicated to increase the volume of orbital tissue in GO, thereby
aggravating the symptoms of eyeball protrusion (26). The results of the current study are
consistent with those of a previous study, which reported that
miR-146a downregulated the expression of collagen I and was
associated with TGF-β-mediated fibrosis (11). In addition, the effect of miR-146a
on HA production was examined in the present study, and it was
revealed that overexpression of miR-146a inhibited HA production,
which suggests that upregulation of miR-146a may reduce GAG
aggregation in patients with GO.
Numerous miRNAs have been indicated to serve
regulatory roles in GO (27).
miR-146a is a member of the miRNA family that regulates gene
expression at the transcriptional level via downregulating its mRNA
targets (13). Previous studies
have indicated that miR-146a participated in several types of
autoimmune diseases, including rheumatoid arthritis (17). In a previous report, miR-146a has
been revealed to reduce the pathogenesis of GO via exerting
anti-inflammatory and anti-fibrotic effects (11), while another study demonstrated that
the upregulation of miR-146a promoted inflammation and disease
progression (20). These studies
have indicated that the effect of miR-146a on GO may be complex, as
miR-146a may exert distinct functions in different cell types or
interact with various target mRNAs. The present study only examined
the role of miR-146a in primary cultures of GO orbital fibroblasts
and additional in-depth investigations are required to verify the
role and the molecular mechanisms of miR-146a function in animal
models of GO.
In conclusion, the present study demonstrated that
miR-146a downregulated the expression of HA and collagen I in GO
orbital fibroblasts, which may reduce GAG aggregation and collagen
deposition, thereby delaying disease progression. Therefore,
miR-146a may represent a novel target for the treatment of GO.
However, therapeutic applications based on miR-146a are not yet
fully developed, and additional studies are required to examine the
effects of miR-146a on other target genes or molecular mechanisms
in GO.
Acknowledgements
Not applicable.
Funding
The present study was funded by National Natural
Science Foundation of China (grant no. 81360152), Guangxi Natural
Science Foundation (grant no. 2018GXNSFAA281234), 2019 Guangxi One
Thousand Young and Middle-Aged College and University Backbone
Teachers Cultivation Program and ‘Medical Excellence Award’ Funded
by the Creative Research Development Grant from the First
Affiliated Hospital of Guangxi Medical University.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
KL proposed the current study and drafted the
manuscript. WL designed and performed the experiments and wrote the
manuscript. CM, HL, LC and SY collected and analyzed experimental
data. All authors read and approved the final manuscript for
publication.
Ethics approval and consent to
participate
The current study was approved [approval no.
2019(KY-E-092)] by the Ethics Review Committee of The First
Affiliated Hospital of Guangxi Medical University and written
informed consent was obtained from all participants prior to
inclusion.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hiromatsu Y, Eguchi H, Tani J, Kasaoka M
and Teshima Y: Graves' ophthalmopathy: Epidemiology and natural
history. Intern Med. 53:353–360. 2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Wang L and Ma JM: Progression of the
pathogenesis of thyroid associated ophthalmopathy. Zhonghua Yan Ke
Za Zhi. 53:474–480. 2017.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
3
|
Gu LQ, Jia HY, Zhao YJ, Liu N, Wang S, Cui
B and Ning G: Association studies of interleukin-8 gene in Graves'
disease and Graves' ophthalmopathy. Endocrine. 36:452–456.
2009.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Park M, Banga JP, Kim GJ, Kim M and Lew H:
Human placenta-derived mesenchymal stem cells ameliorate orbital
adipogenesis in female mice models of Graves' ophthalmopathy. Stem
Cell Res Ther. 10(246)2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Bahn RS and Heufelder AE: Retroocular
fibroblasts: Important effector cells in Graves' ophthalmopathy.
Thyroid. 2:89–94. 1992.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Weetman AP, Cohen S, Gatter KC, Fells P
and Shine B: Immunohistochemical analysis of the retrobulbar
tissues in Graves' ophthalmopathy. Clin Exp Immunol. 75:222–227.
1989.PubMed/NCBI
|
|
7
|
Khong JJ, McNab AA, Ebeling PR, Craig JE
and Selva D: Pathogenesis of thyroid eye disease: Review and update
on molecular mechanisms. Br J Ophthalmol. 100:142–150.
2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kumar S, Coenen M, Iyer S and Bahn RS:
Forkhead transcription factor FOXO1 is regulated by both a
stimulatory thyrotropin receptor antibody and insulin-like growth
factor-1 in orbital fibroblasts from patients with Graves'
ophthalmopathy. Thyroid. 25:1145–1150. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Underhill CB: The interaction of
hyaluronate with the cell surface: The hyaluronate receptor and the
core protein. Ciba Found Symp. 143:87–99; discussion 100-106,
281-285. 1989.PubMed/NCBI View Article : Google Scholar
|
|
10
|
van Steensel L, Paridaens D, Schrijver B,
Dingjan GM, van Daele PL, van Hagen PM, van den Bosch WA, Drexhage
HA, Hooijkaas H and Dik WA: Imatinib mesylate and AMN107 inhibit
PDGF-signaling in orbital fibroblasts: A potential treatment for
Graves' ophthalmopathy. Invest Ophthalmol Vis Sci. 50:3091–3098.
2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Jang SY, Park SJ, Chae MK, Lee JH, Lee EJ
and Yoon JS: Role of microRNA-146a in regulation of fibrosis in
orbital fibroblasts from patients with Graves' orbitopathy. Br J
Ophthalmol. 102:407–414. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wang Y and Smith TJ: Current concepts in
the molecular pathogenesis of thyroid-associated ophthalmopathy.
Invest Ophthalmol Vis Sci. 55:1735–1748. 2014.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Li K, Du Y, Jiang BL and He JF: Increased
microRNA-155 and decreased microRNA-146a may promote ocular
inflammation and proliferation in Graves' ophthalmopathy. Med Sci
Monit. 20:639–643. 2014.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Bhaskaran M and Mohan M: MicroRNAs:
History, biogenesis, and their evolving role in animal development
and disease. Vet Pathol. 51:759–774. 2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Bernardo BC, Ooi JYY, Lin RCY and McMullen
JR: miRNA therapeutics: A new class of drugs with potential
therapeutic applications in the heart. Future Med Chem.
7:1771–1792. 2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wu F, Zikusoka M, Trindade A, Dassopoulos
T, Harris ML, Bayless TM, Brant SR, Chakravarti S and Kwon JH:
MicroRNAs are differentially expressed in ulcerative colitis and
alter expression of macrophage inflammatory peptide-2α.
Gastroenterology. 135:1624–1635.e24. 2008.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Nakasa T, Miyaki S, Okubo A, Hashimoto M,
Nishida K, Ochi M and Asahara H: Expression of microRNA-146 in
rheumatoid arthritis synovial tissue. Arthritis Rheum.
58:1284–1292. 2008.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Tang Y, Luo X, Cui H, Ni X, Yuan M, Guo Y,
Huang X, Zhou H, de Vries N, Tak PP, et al: MicroRNA-146A
contributes to abnormal activation of the type I interferon pathway
in human lupus by targeting the key signaling proteins. Arthritis
Rheum. 60:1065–1075. 2009.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Jang SY, Chae MK, Lee JH, Lee EJ and Yoon
JS: Role of miR-146a in the regulation of inflammation in an in
vitro model of Graves' orbitopathy. Invest Ophthalmol Vis Sci.
57:4027–4034. 2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Wang N, Chen FE and Long ZW: Mechanism of
microrna-146a/notch2 signaling regulating il-6 in Graves
ophthalmopathy. Cell Physiol Biochem. 41:1285–1297. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Li H, Ma C, Liu W, He J and Li K:
Gypenosides protect orbital fibroblasts in Graves ophthalmopathy
via anti-inflammation and anti-fibrosis effects. Invest Ophthalmol
Vis Sci. 61(64)2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Blandford Z, Zhang D, Chundury RV and
Perry JD: Dysthyroid optic neuropathy: Update on pathogenesis,
diagnosis, and management. Expert Rev Ophthalmol. 12:111–121.
2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Yang IH, Rose GE, Ezra DG and Bailly M:
Macrophages promote a profibrotic phenotype in orbital fibroblasts
through increased hyaluronic acid production and cell
contractility. Sci Rep. 9(9622)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Iyer S and Bahn R: Immunopathogenesis of
Graves' ophthalmopathy: The role of the TSH receptor. Best Pract
Res Clin Endocrinol Metab. 26:281–289. 2012.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Longo CM and Higgins PJ: Molecular
biomarkers of Graves' ophthalmopathy. Exp Mol Pathol. 106:1–6.
2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Jang SY, Chae MK, Lee JH, Lee EJ and Yoon
JS: MicroRNA-27 inhibits adipogenic differentiation in orbital
fibroblasts from patients with Graves' orbitopathy. PLoS One.
14(e0221077)2019.PubMed/NCBI View Article : Google Scholar
|