Atherosclerosis is an inflammatory disease and
previous studies have demonstrated that the underlying pathology of
atherosclerosis is mainly the damage to vessel walls caused by
lipid metabolism disorders and the inflammatory immune response,
leading to abnormal lipid deposition in the intima and its
underlying smooth muscle, plaque formation and vascular stenosis
(1). Unstable atherosclerotic
plaques are likely to rupture and cause thromboembolism, which may
result in serious clinical events including acute coronary syndrome
and myocardial infarction. Atherosclerosis is a chronic
inflammatory condition in which a variety of cell types and
physical and chemical factors are involved (2). A mounting body of evidence suggests
that these cytokines involved in atherosclerosis exhibit circadian
oscillations (3).
All organisms on Earth, from bacteria to plants and
mammals, have intrinsic body clocks that respond to environmental
changes by controlling the major physiological activities. In
mammals, the central pacemaker of the circadian rhythm exists in
the suprachiasmatic nucleus (SCN) of the hypothalamus and consists
of 24-h oscillations present in most cells of the body (4). These oscillations are of particular
relevance to physiological and biochemical functions, including
sleep/wake cycles, feeding behavior and activity rhythms (4). The core of the molecular clock is the
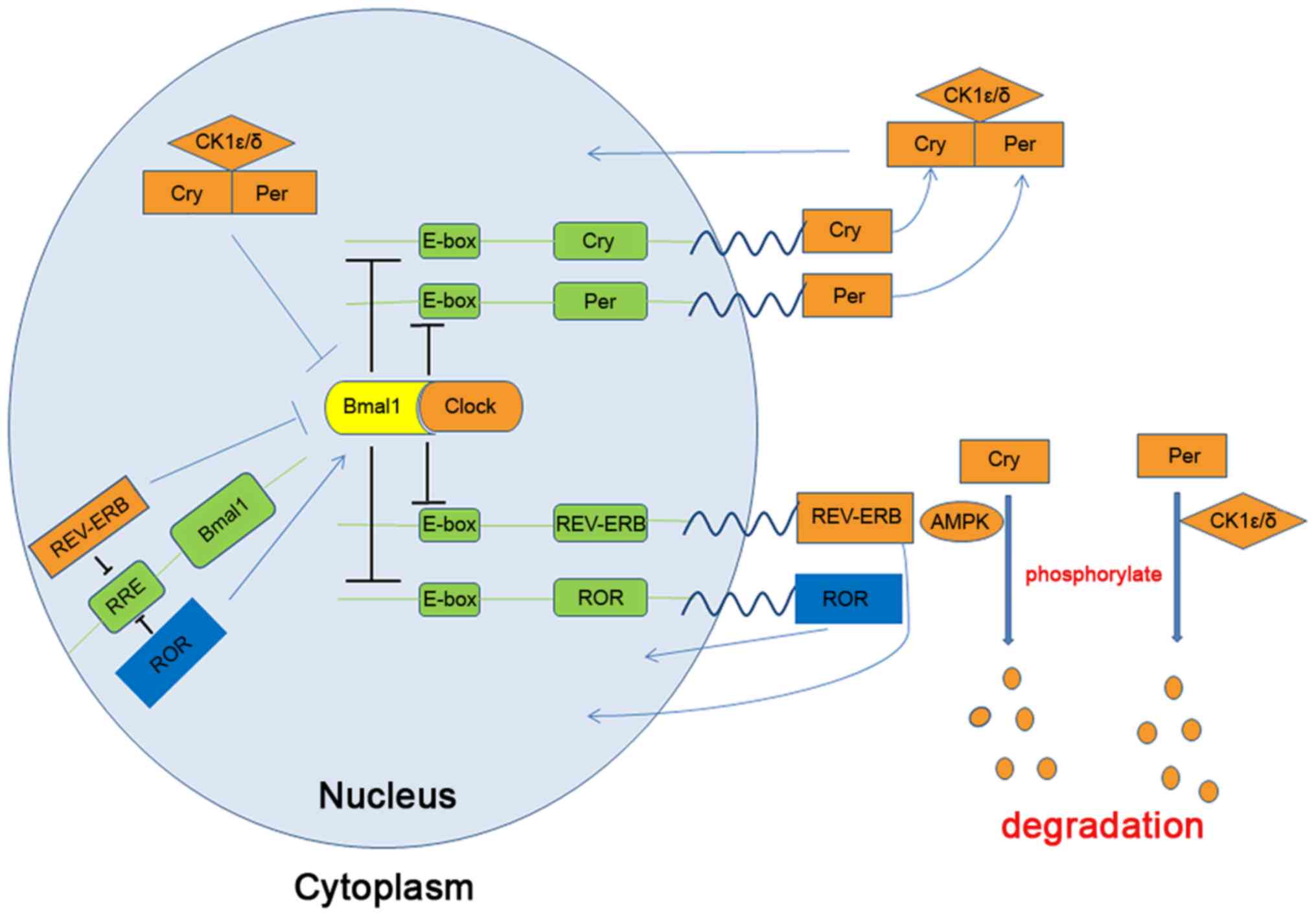
transcription factor heterodimer circadian locomotor output cycle
kaput (Clock)/brain and muscle aryl hydrocarbon receptor nuclear
translocator-like protein 1 (Bmal1), which is driven by two key
negative feedback loops that generate 24-h oscillations of daily
activity (5), as presented in
Fig. 1. The first important
feedback loop consists of Clock, Bmal1, period circadian clock
(Per) and Cryptochromecircadian clock (Cry). In this loop,
Clock/Bmal1 binds to E-box DNA elements to upregulate Per and Cry
levels (3). Per and Cry then
accumulate in the cytoplasm and form a complex with the
serine-threonine kinase casein kinase 1 ε/δ (CK1ε/δ), which
translocates to the nucleus to repress its own transcription and
Clock/Bmal1 activity, thereby forming a negative feedback loop
(5). CK1ε/δ (6,7) and
AMP-activated protein kinase (AMPK) (8) phosphorylate unbound Per and Cry,
respectively, to promote their degradation. Numerous studies have
also demonstrated that other kinase networks, including glycogen
synthase kinase-3, PI3K/Akt and MAPKs, are able to phosphorylate
Per and Cry to promote their degradation (9). The second feedback loop consists of
Clock, Bmal1, nuclear receptor subfamily 1, group D, member 1
(REV-ERB)α and -β, and orphan nuclear receptor (ROR)α, -β or -γ.
Clock/Bmal1 activate the negative regulators REV-ERBα and -β and
the positive regulators RORα, -β or -γ (10,11). REV-ERBs bind to the retinoic
acid-related orphan receptor response element located in the Bmal1
promoter to inhibit transcription (12). An increasing number of studies have
indicated that the molecular clock has an important role in almost
all metabolic processes in organisms. Furthermore, disruptions to
the circadian rhythm may lead to cardiovascular diseases (13), type 2 diabetes (14) and immune system diseases (15).
The progression of atherosclerosis is characterized
by the accumulation of fatty deposits in the inner layer of
arteries. It is well known that the development of atherosclerosis
is related to lipid metabolism, inflammatory reactions, endothelial
cell dysfunction and immune function, and there is growing evidence
that circadian rhythms have a critical role in the development and
progression of the condition. For instance, it was reported that
low density lipoprotein receptor (Ldlr)-/- and
apolipoprotein E (Apoe)-/- mice with global Clock
knockout fed a standard chow diet developed more lesions at the
aortic arches and aortic root (16), and that upregulated Cry1 expression
(17) or REV-ERBβ agonist delivery
(18) reduced atherogenesis in
Ldlr-/- and Apoe-/- mice.
Glycometabolism is a complex physiological process.
In humans, the daily variation in insulin secretion and insulin
sensitivity over 24 h displays an obvious diurnal rhythm (19). Buxton et al (20) demonstrated that the risk of insulin
resistance/type 2 diabetes may be reduced if shift workers focus on
improving sleep duration and implementing circadian readjustment
strategies (such as sleeping during the biological night and eating
during the biological day). Glucose tolerance is higher in the
morning than in the evening and at night (21). Impaired glucose tolerance of type 2
diabetes (22) appears when the
circadian oscillation of the glycometabolism is disrupted. The
neural and peripheral clocks regulate the enzymes of glycolysis,
fatty acid oxidation and oxidative phosphorylation during the 24-h
day to guarantee that these enzymes function at the appropriate
times during the process of glycometabolism (23-25),
which is associated with the circadian rhythm at the
transcriptional level.
A large number of animal experiments and clinical
studies have reported that glucose tolerance and diabetes are
closely related to circadian rhythm disorders. In a laboratory
test, rodents with SCN lesions exhibited whole or partial clock
gene disruptions and developed glycometabolic disorders, including
impaired glucose tolerance (26),
β-cell failure (27), decreased
insulin sensitivity (28),
hyperglycemia (29) and
hyperinsulinemia (29). Clock-
(29) or Bmal1- (30) mutant mice exhibit impaired glucose
tolerance, reduced insulin secretion and decreased pancreatic islet
proliferation. Furthermore, a study involving mice with Clock gene
mutations reported dampening of the oscillations of hepatic
glycogen and glycogen synthase 2 and expression of the limiting
enzyme of glycogenesis (31).
Pancreas- or β-cell-specific Bmal1-knockout mice had elevated
plasma glucose levels, impaired glucose tolerance and decreased
insulin secretion. In addition, repression of Per2 expression
resulted in reduced plasma glucose levels, enhanced insulin
secretion and impaired gluconeogenesis (32). However, in mice lacking Cry1 and
Cry2, plasma glucose levels were elevated in response to acute
feeding after a 12-h overnight fast (33). REV-ERBα is able to regulate plasma
glucose homeostasis by controlling the expression of
glucose-6-phosphatase and phosphoenolpyruvate carboxylase.
REV-ERBα-mutant mice on a high-fat diet had increased adiposity and
mild hyperglycemia without insulin resistance (34). In general, these studies demonstrate
that the circadian clock system possibly maintains the homeostasis
of glycometabolism by regulating the activities of the key enzymes
of glycometabolism (35).
Levels of serum lipids also display an obvious
circadian rhythm. For instance, plasma levels of lipids exhibit
day-night variations independent of food intake, suggesting that
the circadian clock is an important regulator of lipid metabolism
(36). However, the peak level of
plasma high-density lipoprotein (HDL) appears in the early rest
phase and decreases during the active phase (37). A prospective clinical study
suggested that an unhealthy lifestyle and poor-quality sleep
predict the development of hyperlipidemia and obesity with age
(38). Another study suggested that
reduced sleep duration in children contributes to an increased risk
of being overweight (39). It is
therefore indicated that circadian rhythm disorders are associated
with lipid metabolism in mammals. In a clinical study on a
population with obesity, the expression of REV-ERBα exhibited a
marked positive correlation with the body mass index and waist
circumference, and the expression of RORα and Clock was correlated
with HDL and low-density lipoprotein (LDL) levels, respectively
(40). That study also indicated
that the clock genes Cry2 and REV-ERBα were upregulated in obesity
over a 24-h period (40).
Animal experiments also indicated that clock gene
mutations are closely associated with dyslipidemia. Pan et
al (41) reported that compared
to Apoe-/- mice, Bmal1-/- Apoe-/-
and Bmal1 in liver (L-Bmal1)-/- Apoe-/- mice
had an increased risk of hyperlipidemia and atherosclerosis but
that L-Bmal1-/- Apoe-/- mice with
adenovirus-mediated liver overexpression of Bmal1 had a reduced
risk of hyperlipidemia and atherosclerosis. A recent study
suggested that Bmal1 functions as a positive regulator of vascular
smooth muscle cell (VSMC) proliferation following vascular injury
(42), and liver-specific Bmal1- or
REV-ERBα-knockout mice exhibited increased levels of cholesterol,
triglycerides and free fatty acids (43). Furthermore, physiological studies
have demonstrated that enterocytes expressing the dominant-negative
Clock mutant protein (ClockΔ19/Δ19) protein absorb more
cholesterol from the intestinal lumen and secrete cholesterol and
chylomicrons (16). This evidence
indicates that Clock has a vital role in the regulation of
cholesterol metabolism. Additional supporting evidence for
circadian clock involvement has revealed that other core clock
genes are associated with lipid metabolism. Grimaldi et al
(44) indicated that Per2 was a
natural modulator controlling the proadipogenic activity of
peroxisome proliferating activated receptor (PPAR)γ and a major
regulator of lipid metabolism. In addition, Per1/2-null mice or
Per2-null mice had lower hepatic triglyceride levels than wild-type
mice (37). Furthermore, REV-ERBα
deficiency may cause marked hepatic steatosis (10), and mice lacking REV-ERBα displayed
reduced levels of hepatic triglycerides and cholesterol and
elevated levels of plasma lipids (45).
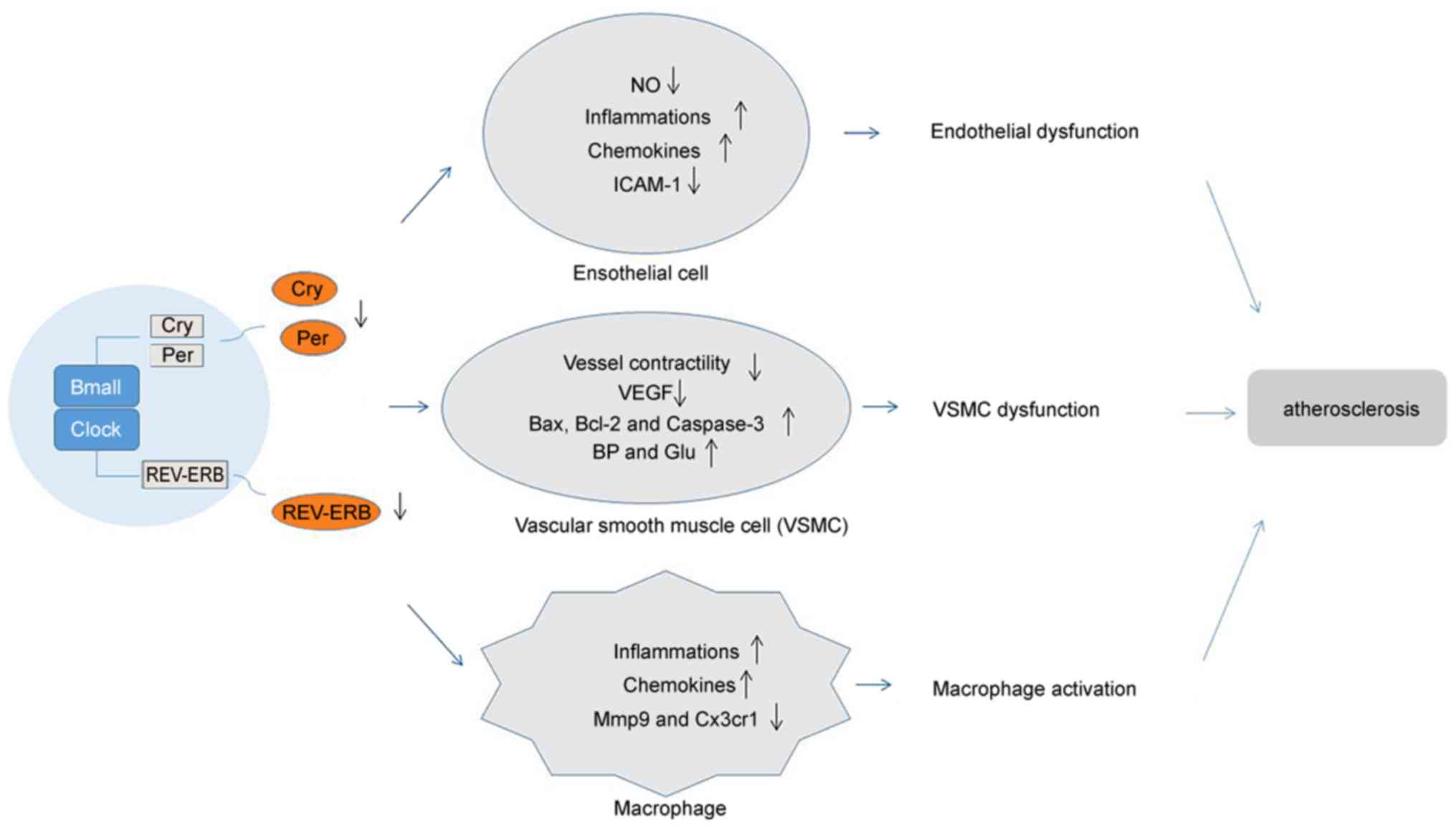
Emerging evidence indicates that circadian rhythm
have a very close connection with vascular cells with regard to
vascular function and health (46).
Circadian rhythms influence the activities of systemic
atherosclerosis mediators, including leukocytes and macrophages,
and locally manipulate cells within the vessel wall. Indeed,
studies have indicated that the functional circadian clock exists
within the vasculature (47). A
growing number of studies (48-55)
have identified that circadian clocks regulate the functions of
endothelial cells, VSMCs and macrophages, suggesting the
possibility of circadian clocks to influence the progression of
atherosclerosis (as presented in Fig.
2). In the present review, a series of clock genes with roles
in vascular cells are presented. Those genes and their functions in
different vascular cell types are listed in Table I.
It is generally known that dysfunction in the
vascular endothelium is a pivotal factor in atherogenesis.
Endothelial cells may be injured and accordingly activated by
numerous stimuli, including oxidized LDLs, hypertension,
hyperglycemia, turbulent blood flow and inflammation. Furthermore,
endothelial cell activation leads to expression of adhesion
molecules, loss of barrier function, migration of leukocytes into
the vascular wall and improvement in inflammatory responses
(46). Tang et al (48) indicated that loss of protective
endothelial Clock expression contributes to the progression of
atherosclerosis and aggravates plaque vulnerability. It appears
that the circadian clock is able to regulate the release of nitric
oxide (NO) and disruption of the clock leads to endothelial
dysfunction (56). Endothelial
dysfunction is associated with decreased bioavailability of
endothelial NO, which is produced by endothelial NO synthase (eNOS)
and the relevant regulatory mechanisms of eNOS activity are closely
interrelated with endothelial dysfunction in atherosclerosis
(57). Human endothelial function
is measured by flow-mediated dilation (FMD) and a recent study
suggested that FMD was decreased in patients with congestive heart
failure and that the circadian variation in endothelial function
was deficient (58).
There is much evidence to support that
Bmal1-knockout and Clock-mutant mice have endothelial dysfunction.
Genetic ablation of the Bmal1 gene in endothelial cells enhances
the expression of the chemokines C-C motif ligand (Ccl)8, Ccl20 and
chemokine (C-X-C motif) ligand (Cxcl)5, damages endothelial
integrity and barrier function (59) and leads to phenotypic features
similar to those of diabetes (60).
Furthermore, Gao et al (61)
reported that the circadian clock may drive endothelial cells to
express intercellular adhesion molecule-1 and promote the adhesion
of monocytes to endothelial cells. Viswambharan et al
(50) indicated that mutation in
the Per2 gene in mice is associated with aortic endothelial
dysfunction involving decreased production of NO and vasodilatory
prostaglandin and increased release of cyclooxygenase-1-derived
vasoconstrictor. Carvas et al (62) concluded that Per2 gene mutation in
aorta reduces insulin-stimulated NO release from endothelial cells.
In a mouse model of sleep deprivation, Qin and Deng (63) indicated that sleep deprivation
promoted the expression of proinflammatory cytokines and decrease
that of Cry1 in vascular endothelial cells. Furthermore, Savalli
et al (64) studied a
naturalistic animal model of depression, concluding that chronic
mild stress-induced anhedonic behavior is associated with disturbed
diurnal oscillation of Clock, Cry2 and Rev-ERBα expression in the
mouse basolateral amygdala and that Clock gene desynchronization
appeared to be involved in vascular endothelial growth factor
(VEGF) variations. In general, it is well accepted that the
circadian clock is important for maintaining normal endothelial
cell functions.
VSMCs constitute the vascular media. During
atherosclerosis progression, the migration of VSMCs from the middle
layer to the intima is of great significance (65). VSMCs are stimulated by a variety of
cytokines and inflammatory cytokines; the phenotype of SMCs
switches from contractile SMCs to secretory SMCs and VSMCs
simultaneously proliferate and migrate to the intima. Specifically,
VSMCs and the extracellular matrix are the major components of the
neointima and subsequent vascular stenosis (66-68).
Studies have indicated that specific clock gene
knockout of Bmal1 in VSMCs impairs vessel contractility and
decreases the blood pressure; in addition, smooth muscle-specific
Bmal1 has a vital role in normal VSM contraction (69). According to Suyama et al
(70), Bmal1 knockdown resulted in
a decrease in VEGF and VEGF receptor mRNA expression compared with
the control. Furthermore, VSMCs cultured from the carotid arteries
of healthy donors exhibited regular circadian mRNA expression of
Bmal1, Per1, Per2, Per3, Cry1, Cry2 and Rev-ERBα (71). In addition, the circadian rhythm of
the major rhythm genes isolated from human plaque-derived VSMCs is
significantly attenuated compared to that from cells cultured from
the carotid arteries of healthy donors (71). Similarly, hyperglycemia and
hyperlipidemia are associated with circadian gene expression in
VSMCs (72,73). Su et al (72) suggested that expression of Per1/2,
Cry1/2, D site albumin promoter-binding protein (Dbp) and PPARγ in
the aorta and mesenteric arteries in db/db mice was suppressed
compared with that in control mice. Specifically, Migita et
al (55) reported that Rev-ERBα
causes upregulation of NF-кB-responsive genes in VSMCs. These
observations indicate that the circadian clock is of great
significance to VSMC functions.
Macrophages are involved in the formation of
atherosclerosis and have a different role in atherosclerotic lesion
development (46). Macrophage
accumulation within the vascular wall is a hallmark of
atherosclerosis. In atherosclerotic lesions, macrophages respond to
various environmental stimuli, such as modified lipids, cytokines,
and senescent erythrocytes, which can modify their functional
phenotypes. Furthermore, an increase in the inflammatory reaction
decreases plaque stability and results in a thrombotic event. Of
note, as with VSMCs and endothelial cells, circadian rhythms are
able to regulate macrophage function (74).
In the wall of vessels affected by atherosclerosis,
macrophages secrete inflammatory factors, including chemokines and
cytokines. Specifically, the cytokine storms of infected mice
display the greatest reaction at the beginning of infection
(75). In addition, circadian
expression of monocyte chemoattractant protein-1 (MCP-1/JE) in
macrophages is regulated by Bmal1 via activation of NF-кB (76). The molecular mechanisms by which
cytokine production by macrophages is regulated remain to be fully
elucidated. However, evidence indicates that the expression of
Bmal1 is regulated by a circadian rhythm in macrophages (76). Rev-ERBα- or Bmal1-knockout
macrophages displayed increased production of cytokines and
decreased expression of circadian rhythm genes (75). Huo et al (77) concluded that Bmal1 deficiency in
macrophages may exacerbate atherosclerosis by promoting the
recruitment of Ly6Chi monocytes to atherosclerotic lesions.
Similarly, compared to wildtype control mice, Clock-mutant mice
have increased circulating IL-12 and IL-17 levels and altered
NF-кB-induced macrophage activation (16,51).
Furthermore, Rev-ERBα regulate enhancer-derived RNAs, suppressing
the expression of nearby genes, including Mmp9 and CX3C chemokine
ligand receptor 1(Cx3cr1; one of the most expressed genes in
microglia in mice and humans, is implicated in numerous microglial
functions) in macrophages (78),
and Rev-ERBα modulate the inflammatory infiltration of macrophages
by inhibiting the expression of Ccl2(79). It remains elusive whether clock
genes control local macrophage proliferation.
The levels of immune inflammatory immunoreactive
cells and pro-inflammatory cytokines have an obvious daily rhythm
(80) and functionality of the
immune system has been linked to the circadian clock (81). A recent study reported that
circadian disruption by sleep fragmentation accelerates
atherosclerosis development by increasing the number of circulating
monocytes (82). Consequently,
disturbed circadian clock function may contribute to the risk of
atherosclerosis through the pro-inflammatory state. Based on in
vivo and in vitro experiments, the circadian rhythm is
closely linked to inflammatory immunoreactions (52,83).
In cells lacking Bmal1, lower expression of BMAL1 may affect
mediators of inflammation and oxidative stress (52). Keller et al (74) indicated that Bmal1 mRNA expression
in macrophages was lowest at the active phase (zeitgeber time 12).
Furthermore, in animal models of inflammatory bowel disease
(51), the expression of
inflammatory factors in Clock gene-mutant mice was decreased
compared with that in wild-type mice. In addition, the circadian
rhythm expression of IFN-γ disappeared in Per2-mutant mice
(84). In an arthritis model
(85), simultaneous knockout of
Cry-1 and Cry-2 led to an increase in TNF-α and aggravated the
inflammatory response. In addition, the clock genes Rev-ERBα and
RORα have important roles in inflammatory immunoreactions (86,87).
Numerous studies have suggested that the circadian
rhythm is significant regarding several aspects of atherosclerosis,
including glycometabolism, lipid metabolism, endothelial cell
dysfunction, VSMC phenotype and inflammatory immunoreactions.
However, the mechanism of specific clock genes involved in the
atherosclerotic process remains elusive. Thus, further research on
the relationship between the circadian rhythm and atherosclerosis
is required. There is much to learn about the circadian rhythm,
promoting a healthy lifestyle that includes sufficient, regular
sleep, this may provide novel therapeutic targets and preventative
measures to simultaneously slow the development of atherosclerosis
and reduce cardiovascular mortality.
Not applicable.
The present work was supported by the National
Natural Science Foundation of China (grant nos. 81770456 and
81400794).
Not applicable.
ZZ, JD and XW conceived and designed the article.
ZZ, BY, CL, XZ and TZ collected related articles and analyzed the
relevant literature. ZZ and BY wrote the manuscript and drew the
figures. JD, CL, XZ, TZ and XW revised the manuscript. All authors
read and approved the final version of the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Shah MS and Brownlee M: Molecular and
cellular mechanisms of cardiovascular disorders in diabetes. Circ
Res. 118:1808–1829. 2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Nguyen KD, Fentress SJ, Qiu Y, Yun K, Cox
JS and Chawla A: Circadian gene Bmal1 regulates diurnal
oscillations of Ly6C(hi) inflammatory monocytes. Science.
341:1483–1488. 2013.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Scheiermann C, Kunisaki Y and Frenette PS:
Circadian control of the immune system. Nat Rev Immunol.
13:190–198. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
4
|
Steffens S, Winter C, Schloss MJ, Hidalgo
A, Weber C and Soehnlein O: Circadian control of inflammatory
processes in atherosclerosis and its complications. Arterioscler
Thromb Vasc Biol. 37:1022–1028. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Mohawk JA, Green CB and Takahashi JS:
Central and peripheral circadian clocks in mammals. Annu Rev
Neurosci. 35:445–462. 2012.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Lowrey PL, Shimomura K, Antoch MP,
Yamazaki S, Zemenides PD, Ralph MR, Menaker M and Takahashi JS:
Positional syntenic cloning and functional characterization of the
mammalian circadian mutation tau. Science. 288:483–492.
2000.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Shirogane T, Jin J, Ang XL and Harper JW:
SCFbeta-TRCP controls clock-dependent transcription via casein
kinase 1-dependent degradation of the mammalian period-1 (Per1)
protein. J Biol Chem. 280:26863–26872. 2005.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Lamia KA, Sachdeva UM, DiTacchio L,
Williams EC, Alvarez JG, Egan DF, Vasquez DS, Juguilon H, Panda S,
Shaw RJ, et al: AMPK regulates the circadian clock by cryptochrome
phosphorylation and degradation. Science. 326:437–440.
2009.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Buhr ED and Takahashi JS: Molecular
components of the Mammalian circadian clock. Handb Exp Pharmacol.
217:3–27. 2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Bugge A, Feng D, Everett LJ, Briggs ER,
Mullican SE, Wang F, Jager J and Lazar MA: Rev-erbα and Rev-erbβ
coordinately protect the circadian clock and normal metabolic
function. Genes Dev. 26:657–667. 2012.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Solt LA, Wang Y, Banerjee S, Hughes T,
Kojetin DJ, Lundasen T, Shin Y, Liu J, Cameron MD, Noel R, et al:
Regulation of circadian behaviour and metabolism by synthetic
REV-ERB agonists. Nature. 485:62–68. 2012.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Crumbley C and Burris TP: Direct
regulation of CLOCK expression by REV-ERB. PLoS One.
6(e17290)2011.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Skogstad M, Mamen A, Lunde LK, Ulvestad B,
Matre D, Aass H, Øvstebø R, Nielsen P, Samuelsen KN, Skare Ø and
Sirnes PA: Shift work including night work and long working hours
in industrial plants increases the risk of atherosclerosis. Int J
Environ Res Public Health. 16(521)2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Laermans J and Depoortere I: Chronobesity:
Role of the circadian system in the obesity epidemic. Obes Rev.
17:108–125. 2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
O'Keeffe SM, Beynon AL, Davies JS, Moynagh
PN and Coogan AN: NF-κB signalling is involved in
immune-modulation, but not basal functioning, of the mouse
suprachiasmatic circadian clock. Eur J Neurosci. 45:1111–1123.
2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Pan X, Jiang XC and Hussain MM: Impaired
cholesterol metabolism and enhanced atherosclerosis in clock mutant
mice. Circulation. 128:1758–1769. 2013.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Yang L, Chu Y, Wang L, Wang Y, Zhao X, He
W, Zhang P, Yang X, Liu X, Tian L, et al: Overexpression of CRY1
protects against the development of atherosclerosis via the
TLR/NF-κB pathway. Int Immunopharmacol. 28:525–530. 2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Sitaula S, Billon C, Kamenecka TM, Solt LA
and Burris TP: Suppression of atherosclerosis by synthetic REV-ERB
agonist. Biochem Biophys Res Commun. 460:566–571. 2015.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Kalsbeek A, la Fleur S and Fliers E:
Circadian control of glucose metabolism. Mol Metab. 3:372–383.
2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Buxton OM, Cain SW, O'Connor SP, Porter
JH, Duffy JF, Wang W, Czeisler CA and Shea SA: Adverse metabolic
consequences in humans of prolonged sleep restriction combined with
circadian disruption. Sci Transl Med. 4(129ra43)2012.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Morris CJ, Yang JN, Garcia JI, Myers S,
Bozzi I, Wang W, Buxton OM, Shea SA and Scheer FA: Endogenous
circadian system and circadian misalignment impact glucose
tolerance via separate mechanisms in humans. Proc Natl Acad Sci
USA. 112:E2225–E2234. 2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Polonsky KS, Given BD, Hirsch LJ, Tillil
H, Shapiro ET, Beebe C, Frank BH, Galloway JA and Van Cauter E:
Abnormal patterns of insulin secretion in non-insulin-dependent
diabetes mellitus. N Engl J Med. 318:1231–1239. 1988.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Panda S, Antoch MP, Miller BH, Su AI,
Schook AB, Straume M, Schultz PG, Kay SA, Takahashi JS and
Hogenesch JB: Coordinated transcription of key pathways in the
mouse by the circadian clock. Cell. 109:307–320. 2002.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Yang X, Downes M, Yu RT, Bookout AL, He W,
Straume M, Mangelsdorf DJ and Evans RM: Nuclear receptor expression
links the circadian clock to metabolism. Cell. 126:801–810.
2006.PubMed/NCBI View Article : Google Scholar
|
|
25
|
McCarthy JJ, Andrews JL, McDearmon EL,
Campbell KS, Barber BK, Miller BH, Walker JR, Hogenesch JB,
Takahashi JS and Esser KA: Identification of the circadian
transcriptome in adult mouse skeletal muscle. Physiol Genomics.
31:86–95. 2007.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Shi SQ, Ansari TS, McGuinness OP,
Wasserman DH and Johnson CH: Circadian disruption leads to insulin
resistance and obesity. Curr Biol. 23:372–381. 2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Lee J, Moulik M, Fang Z, Saha P, Zou F, Xu
Y, Nelson DL, Ma K, Moore DD and Yechoor VK: Bmal1 and β-cell clock
are required for adaptation to circadian disruption, and their loss
of function leads to oxidative stress-induced β-cell failure in
mice. Mol Cell Biol. 33:2327–2338. 2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Coomans CP, van den Berg SA, Lucassen EA,
Houben T, Pronk AC, van der Spek RD, Kalsbeek A, Biermasz NR,
Willems van Dijk K, Romijn JA and Meijer JH: The suprachiasmatic
nucleus controls circadian energy metabolism and hepatic insulin
sensitivity. Diabetes. 62:1102–1108. 2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Turek FW, Joshu C, Kohsaka A, Lin E,
Ivanova G, McDearmon E, Laposky A, Losee-Olson S, Easton A, Jensen
DR, et al: Obesity and metabolic syndrome in circadian Clock mutant
mice. Science. 308:1043–1045. 2005.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Rudic RD, McNamara P, Curtis AM, Boston
RC, Panda S, Hogenesch JB and Fitzgerald GA: BMAL1 and CLOCK, two
essential components of the circadian clock, are involved in
glucose homeostasis. PLoS Biol. 2(e377)2004.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Doi R, Oishi K and Ishida N: CLOCK
regulates circadian rhythms of hepatic glycogen synthesis through
transcriptional activation of Gys2. J Biol Chem. 285:22114–22121.
2010.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zani F, Breasson L, Becattini B, Vukolic
A, Montani JP, Albrecht U, Provenzani A, Ripperger JA and Solinas
G: PER2 promotes glucose storage to liver glycogen during feeding
and acute fasting by inducing Gys2 PTG and G L expression. Mol
Metab. 2:292–305. 2013.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Lamia KA, Papp SJ, Yu RT, Barish GD,
Uhlenhaut NH, Jonker JW, Downes M and Evans RM: Cryptochromes
mediate rhythmic repression of the glucocorticoid receptor. Nature.
480:552–556. 2011.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Delezie J, Dumont S, Dardente H, Oudart H,
Gréchez-Cassiau A, Klosen P, Teboul M, Delaunay F, Pévet P and
Challet E: The nuclear receptor REV-ERBα is required for the daily
balance of carbohydrate and lipid metabolism. FASEB J.
26:3321–3335. 2012.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Tao H, Li X, Qiu JF, Cui WZ, Sima YH and
Xu SQ: Inhibition of expression of the circadian clock gene Period
causes metabolic abnormalities including repression of
glycometabolism in Bombyx mori cells. Sci Rep.
7(46258)2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Chua EC, Shui G, Lee IT, Lau P, Tan LC,
Yeo SC, Lam BD, Bulchand S, Summers SA, Puvanendran K, et al:
Extensive diversity in circadian regulation of plasma lipids and
evidence for different circadian metabolic phenotypes in humans.
Proc Natl Acad Sci USA. 110:14468–14473. 2013.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Adamovich Y, Rousso-Noori L, Zwighaft Z,
Neufeld-Cohen A, Golik M, Kraut-Cohen J, Wang M, Han X and Asher G:
Circadian clocks and feeding time regulate the oscillations and
levels of hepatic triglycerides. Cell Metab. 19:319–330.
2014.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Spiegel K, Tasali E, Leproult R and Van
Cauter E: Effects of poor and short sleep on glucose metabolism and
obesity risk. Nat Rev Endocrinol. 5:253–261. 2009.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Lumeng JC, Somashekar D, Appugliese D,
Kaciroti N, Corwyn RF and Bradley RH: Shorter sleep duration is
associated with increased risk for being overweight at ages 9 to 12
years. Pediatrics. 120:1020–1029. 2007.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Vieira E, Eg R, Figueroa AL, Aranda G,
Momblan D, Carmona F, Gomis R, Vidal J and Hanzu FA: Altered clock
gene expression in obese visceral adipose tissue is associated with
metabolic syndrome. PLoS One. 9(e111678)2014.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Pan X, Bradfield CA and Hussain MM: Global
and hepatocyte-specific ablation of Bmal1 induces hyperlipidaemia
and enhances atherosclerosis. Nat Commun. 7(13011)2016.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Takaguri A, Sasano J, Akihiro O and Satoh
K: The role of circadian clock gene BMAL1 in vascular
proliferation. Eur J Pharmacol. 872(172924)2020.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Jacobi D, Liu S, Burkewitz K, Kory N,
Knudsen NH, Alexander RK, Unluturk U, Li X, Kong X, Hyde AL, et al:
Hepatic Bmal1 regulates rhythmic mitochondrial dynamics and
promotes metabolic Fitness. Cell Metab. 22:709–720. 2015.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Grimaldi B, Bellet MM, Katada S, Astarita
G, Hirayama J, Amin RH, Granneman JG, Piomelli D, Leff T and
Sassone-Corsi P: PER2 controls lipid metabolism by direct
regulation of PPARγ. Cell Metab. 12:509–520. 2010.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Le Martelot G, Claudel T, Gatfield D,
Schaad O, Kornmann B, Lo Sasso G, Moschetta A and Schibler U:
REV-ERBalpha participates in circadian SREBP signaling and bile
acid homeostasis. PLoS Biol. 7(e1000181)2009.PubMed/NCBI View Article : Google Scholar
|
|
46
|
McAlpine CS and Swirski FK: Circadian
influence on metabolism and inflammation in atherosclerosis. Circ
Res. 119:131–141. 2016.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Davidson AJ, London B, Block GD and
Menaker M: Cardiovascular tissues contain independent circadian
clocks. Clin Exp Hypertens. 27:307–311. 2005.PubMed/NCBI
|
|
48
|
Tang H, Zhu M, Zhao G, Fu W, Shi Z, Ding
Y, Tang X and Guo D: Loss of CLOCK under high glucose upregulates
ROCK1-mediated endothelial to mesenchymal transition and aggravates
plaque vulnerability. Atherosclerosis. 275:58–67. 2018.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Takeda N, Maemura K, Horie S, Oishi K,
Imai Y, Harada T, Saito T, Shiga T, Amiya E, Manabe I, et al:
Thrombomodulin is a clock-controlled gene in vascular endothelial
cells. J Biol Chem. 282:32561–32567. 2007.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Viswambharan H, Carvas JM, Antic V,
Marecic A, Jud C, Zaugg CE, Ming XF, Montani JP, Albrecht U and
Yang Z: Mutation of the circadian clock gene Per2 alters vascular
endothelial function. Circulation. 115:2188–2195. 2007.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Bellet MM, Deriu E, Liu JZ, Grimaldi B,
Blaschitz C, Zeller M, Edwards RA, Sahar S, Dandekar S, Baldi P, et
al: Circadian clock regulates the host response to
Salmonella. Proc Natl Acad Sci USA. 110:9897–9902.
2013.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Early JO, Menon D, Wyse CA,
Cervantes-Silva MP, Zaslona Z, Carroll RG, Palsson-McDermott EM,
Angiari S, Ryan DG, Corcoran SE, et al: Circadian clock protein
BMAL1 regulates IL-1β in macrophages via NRF2. Proc Natl Acad Sci
USA. 115:E8460–E8468. 2018.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Pourcet B, Zecchin M, Ferri L, Beauchamp
J, Sitaula S, Billon C, Delhaye S, Vanhoutte J, Mayeuf-Louchart A,
Thorel Q, et al: Nuclear receptor subfamily 1 Group D Member 1
regulates circadian activity of NLRP3 inflammasome to reduce the
severity of fulminant hepatitis in mice. Gastroenterology.
154:1449–1464.e20. 2018.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Chen L, Wu X, Zeb F, Huang Y, An J, Jiang
P, Chen A, Xu C and Feng Q: Acrolein-induced apoptosis of smooth
muscle cells through NEAT1-Bmal1/Clock pathway and a protection
from asparagus extract. Environ Pollut. 258(113735)2020.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Migita H, Morser J and Kawai K:
Rev-Erbalpha upregulates NF-kappaB-responsive genes in vascular
smooth muscle cells. FEBS Lett. 561:69–74. 2004.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Rodrigo GC and Herbert KE: Regulation of
vascular function and blood pressure by circadian variation in
redox signalling. Free Radic Biol Med. 119:115–120. 2018.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Yang Z and Ming XF: Recent advances in
understanding endothelial dysfunction in atherosclerosis. Clin Med
Res. 4:53–65. 2006.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Maruo T, Nakatani S, Kanzaki H, Kakuchi H,
Yamagishi M, Kitakaze M, Ohe T and Miyatake K: Circadian variation
of endothelial function in idiopathic dilated cardiomyopathy. Am J
Cardiol. 97:699–702. 2006.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Gibbs J, Ince L, Matthews L, Mei J, Bell
T, Yang N, Saer B, Begley N, Poolman T, Pariollaud M, et al: An
epithelial circadian clock controls pulmonary inflammation and
glucocorticoid action. Nat Med. 20:919–926. 2014.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Bhatwadekar AD, Beli E, Diao Y, Chen J,
Luo Q, Alex A, Caballero S, Dominguez JM, Salazar TE, Busik JV, et
al: Conditional deletion of Bmal1 accentuates microvascular and
macrovascular injury. Am J Pathol. 187:1426–1435. 2017.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Gao Y, Meng D, Sun N, Zhu Z, Zhao R, Lu C,
Chen S, Hua L and Qian R: Clock upregulates intercellular adhesion
molecule-1 expression and promotes mononuclear cells adhesion to
endothelial cells. Biochem Biophys Res Commun. 443:586–591.
2014.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Carvas JM, Vukolic A, Yepuri G, Xiong Y,
Popp K, Schmutz I, Chappuis S, Albrecht U, Ming XF, Montani JP and
Yang Z: Period2 gene mutant mice show compromised insulin-mediated
endothelial nitric oxide release and altered glucose homeostasis.
Front Physiol. 3(337)2012.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Qin B and Deng Y: Overexpression of
circadian clock protein cryptochrome (CRY) 1 alleviates sleep
deprivation-induced vascular inflammation in a mouse model. Immunol
Lett. 163:76–83. 2015.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Savalli G, Diao W, Schulz S, Todtova K and
Pollak DD: Diurnal oscillation of amygdala clock gene expression
and loss of synchrony in a mouse model of depression. Int J
Neuropsychopharmacol. 18(pyu095)2014.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Lacolley P, Regnault V, Segers P and
Laurent S: Vascular smooth muscle cells and arterial stiffening:
Relevance in development, aging, and disease. Physiol Rev.
97:1555–1617. 2017.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Feil S, Hofmann F and Feil R: SM22alpha
modulates vascular smooth muscle cell phenotype during
atherogenesis. Circ Res. 94:863–865. 2004.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Rudolph V and Freeman BA: Cardiovascular
consequences when nitric oxide and lipid signaling converge. Circ
Res. 105:511–522. 2009.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Wang L, Zheng J, Bai X, Liu B, Liu CJ, Xu
Q, Zhu Y, Wang N, Kong W and Wang X: ADAMTS-7 mediates vascular
smooth muscle cell migration and neointima formation in
balloon-injured rat arteries. Circ Res. 104:688–698.
2009.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Xie Z, Su W, Liu S, Zhao G, Esser K,
Schroder EA, Lefta M, Stauss HM, Guo Z and Gong MC: Smooth-muscle
BMAL1 participates in blood pressure circadian rhythm regulation. J
Clin Invest. 125:324–336. 2015.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Suyama K, Silagi ES, Choi H, Sakabe K,
Mochida J, Shapiro IM and Risbud MV: Circadian factors BMAL1 and
RORα control HIF-1α transcriptional activity in nucleus pulposus
cells: Implications in maintenance of intervertebral disc health.
Oncotarget. 7:23056–23071. 2016.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Lin C, Tang X, Zhu Z, Liao X, Zhao R, Fu
W, Chen B, Jiang J, Qian R and Guo D: The rhythmic expression of
clock genes attenuated in human plaque-derived vascular smooth
muscle cells. Lipids Health Dis. 13(14)2014.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Su W, Xie Z, Guo Z, Duncan MJ, Lutshumba J
and Gong MC: Altered clock gene expression and vascular smooth
muscle diurnal contractile variations in type 2 diabetic db/db
mice. Am J Physiol Heart Circ Physiol. 302:H621–H633.
2012.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Chen S, Ding Y, Zhang Z, Wang H and Liu C:
Hyperlipidaemia impairs the circadian clock and physiological
homeostasis of vascular smooth muscle cells via the suppression of
Smarcd1. J Pathol. 233:159–169. 2014.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Keller M, Mazuch J, Abraham U, Eom GD,
Herzog ED, Volk HD, Kramer A and Maier B: A circadian clock in
macrophages controls inflammatory immune responses. Proc Natl Acad
Sci USA. 106:21407–21412. 2009.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Gibbs JE, Blaikley J, Beesley S, Matthews
L, Simpson KD, Boyce SH, Farrow SN, Else KJ, Singh D, Ray DW, et
al: The nuclear receptor REV-ERBα mediates circadian regulation of
innate immunity through selective regulation of inflammatory
cytokines. Proc Natl Acad Sci USA. 109:582–587. 2012.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Hayashi M, Shimba S and Tezuka M:
Characterization of the molecular clock in mouse peritoneal
macrophages. Biol Pharm Bull. 30:621–626. 2007.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Huo M, Huang Y, Qu D, Zhang H, Wong WT,
Chawla A, Huang Y and Tian XY: Myeloid Bmal1 deletion increases
monocyte recruitment and worsens atherosclerosis. FASEB J.
31:1097–1106. 2017.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Lam MT, Cho H, Lesch HP, Gosselin D, Heinz
S, Tanaka-Oishi Y, Benner C, Kaikkonen MU, Kim AS, Kosaka M, et al:
Rev-Erbs repress macrophage gene expression by inhibiting
enhancer-directed transcription. Nature. 498:511–515.
2013.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Sato S, Sakurai T, Ogasawara J, Takahashi
M, Izawa T, Imaizumi K, Taniguchi N, Ohno H and Kizaki T: A
circadian clock gene, Rev-erbα, modulates the inflammatory function
of macrophages through the negative regulation of Ccl2 expression.
J Immunol. 192:407–417. 2014.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Xie M, Tang Q, Nie J, Zhang C, Zhou X, Yu
S, Sun J, Cheng X, Dong N, Hu Y and Chen L: BMAL1-downregulation
aggravates Porphyromonas gingivalis-induced atherosclerosis
by encouraging oxidative stress. Circ Res. 126:e15–e29.
2020.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Lucassen EA, Coomans CP, van Putten M, de
Kreij SR, van Genugten JH, Sutorius RP, de Rooij KE, van der Velde
M, Verhoeve SL, Smit JW, et al: Environmental 24-hr cycles are
essential for health. Curr Biol. 26:1843–1853. 2016.PubMed/NCBI View Article : Google Scholar
|
|
82
|
McAlpine CS, Kiss MG, Rattik S, He S,
Vassalli A, Valet C, Anzai A, Chan CT, Mindur JE, Kahles F, et al:
Sleep modulates haematopoiesis and protects against
atherosclerosis. Nature. 566:383–387. 2019.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Schilperoort M, van den Berg R, Bosmans
LA, van Os BW, Dollé M, Smits N, Guichelaar T, van Baarle D,
Koemans L, Berbée J, et al: Disruption of circadian rhythm by
alternating light-dark cycles aggravates atherosclerosis
development in APOE*3-Leiden.CETP mice. J Pineal Res.
68(e12614)2020.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Arjona A and Sarkar DK: The circadian gene
mPer2 regulates the daily rhythm of IFN-gamma. J Interferon
Cytokine Res. 26:645–649. 2006.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Hashiramoto A, Yamane T, Tsumiyama K,
Yoshida K, Komai K, Yamada H, Yamazaki F, Doi M, Okamura H and
Shiozawa S: Mammalian clock gene Cryptochrome regulates arthritis
via proinflammatory cytokine TNF-alpha. J Immunol. 184:1560–1565.
2010.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Raffatellu M, George MD, Akiyama Y,
Hornsby MJ, Nuccio SP, Paixao TA, Butler BP, Chu H, Santos RL,
Berger T, et al: Lipocalin-2 resistance confers an advantage to
Salmonella enterica serotype Typhimurium for growth and
survival in the inflamed intestine. Cell Host Microbe. 5:476–486.
2009.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Yu X, Rollins D, Ruhn KA, Stubblefield JJ,
Green CB, Kashiwada M, Rothman PB, Takahashi JS and Hooper LV: TH17
cell differentiation is regulated by the circadian clock. Science.
342:727–730. 2013.PubMed/NCBI View Article : Google Scholar
|