Introduction
Colorectal cancer (CRC) is the most common
malignancy of the digestive system and it is associated with high
morbidity and mortality rates (1).
According to the cancer statistics from 185 countries, ~800,977 new
CRC cases were reported in 2018 and the mortality rate was as high
as 47.8% (2). Although radical
surgical resection combined with radiotherapy or chemotherapy is
widely used for the treatment of CRC, its efficacy remains low; the
5-year survival rate of patients with CRC without metastasis is
40-90%, which drops to 10-15% in patients exhibiting metastasis
(3). Thus, clinically effective
therapeutic targets for CRC are still limited (4), and the identification of novel
specific molecular targets is required to identify the mechanisms
underlying CRC tumorigenesis and for the development of new
drugs.
Gene expression profiling analysis based on large
datasets serves an increasingly important role in determining
potential molecular markers for different types of cancer (5,6). To
investigate new CRC-related genes, the present study screened
differentially expressed genes (DEGs) in 32 patients with CRC from
RNA-Seq datasets in The Cancer Genome Atlas (TCGA) database and
identified keratin 80 (KRT80) as the most upregulated gene.
KRT80 is located on chromosome 12q13 and encodes a
452-amino-acid protein that is a type II keratin (7,8).
Keratins are intermediate filament cytoskeletal proteins that
maintain the structural integrity of epithelial cells and they have
also been reported to be representative markers for epithelial
cells (9,10). Keratin expression is tissue-specific
and related to advanced tissue or cell differentiation (10). Previous studies have revealed that
keratins were extensively expressed in human cancer and suggested
that they may be used as molecular markers for the diagnosis of
multiple types of tumor, such as basal cell carcinoma, oral
squamous cell carcinoma, bladder cancer, breast cancer,
hepatocellular carcinoma, cervical cancer and gastric
adenocarcinoma (11). Furthermore,
it has been reported that keratins served an important role in the
regulation of cancer cell migration and invasion (12,13).
In fact, the keratin family in humans consists of 28 type I
keratins (KRT9-KRT40) and 26 type II keratins (KRT1-KRT8,
KRT71-KRT86) (14,15). Among them, KRT7, KRT8 and
KRT18-KRT20 were found to participate in the proliferation and
differentiation of colon cells (16-18).
However, to the best of our knowledge, there are few reports
investigating the association between KRT80 expression levels and
human cancer. Especially in CRC, only one previous study has
demonstrated that KRT80 promoted the migration and invasion of CRC
cells (19). Thus, the functional
role of KRT80 in CRC cell proliferation remains relatively
unclear.
In the present study, gene expression profiling
analysis was performed to identify DEGs associated with CRC.
Following the identification of KRT80, the study aimed to
investigate the expression, function and underlying mechanisms of
KRT80 in CRC cell proliferation. The findings supported the role of
KRT80 in CRC development, and provided further experimental
evidence and a theoretical basis for using KRT80 as a therapeutic
target in patients with CRC.
Materials and methods
Data mining and analysis
The mRNA expression profiles of human CRC cases from
RNA-Seq datasets in the TCGA database
(TCGA_COADREAD_exp_HiSeqV2-2015-02-24) were downloaded from the
UCSC Xena Browser (https://xenabrowser.net/datapages/). The DEGs between
32 CRC tissues and adjacent normal tissues were identified using
the limma package (v3.34.9) of R software (20). DEGs with the cutoff criteria of
false discovery rate (FDR) <0.001,
|log2(fold-change)|≥1 and average expression value ≥5
were considered to be statistically significant. The mRNA expression
levels of KRT80 in multiple forms of cancer (including bladder,
brain, breast, cervical, colorectal, esophageal, gastric, head and
neck, kidney, ovarian, liver, lung, pancreatic and prostate cancer
and lymphoma, melanoma, myeloma, leukemia and sarcoma) were further
analyzed using ONCOMINE database (access date: Januay 9th 2019;
www.oncomine.org) (21), with cut-off values of P<0.001 and
a fold-change of 2.0. The proteins co-expressed with KRT80 in
TCGA-CRC cases (false discovery rate values <0.05 and absolute
Spearman's rank correlation coefficient ≥0.4) were identified using
the cBioPortal database (accessed March 2019; http://www.cbioportal.org) (22,23).
The co-expressed proteins were subjected to pathway analysis using
the Reactome database (accessed March 2019; https://reactome.org/PathwayBrowser) (24) and P<0.05 as the cut-off
value.
Cell culture
Human CRC cell lines (HCT116, RKO, LoVo, HT29, SW480
and SW620), the normal colorectal epithelial cell line FHC and
normal colorectal fibroblasts CCD18CO were obtained from the
American Type Culture Collection. HCT 116 and HT-29 cells were
cultured in McCoy's 5A medium (Gibco; Thermo Fisher Scientific,
Inc.); RKO and CCD18CO cells were cultured in Eagle's MEM medium
(Gibco; Thermo Fisher Scientific, Inc.); LoVo cells were cultured
in Ham's F-12K medium (Gibco; Thermo Fisher Scientific, Inc.);
SW480 and SW620 cells were cultured in RPMI-1640 medium (HyClone;
GE Healthcare Life Sciences); and FHC cells were cultured in
DMEM/F12 medium (Gibco; Thermo Fisher Scientific, Inc.). All media
were supplemented with 10% (v/v) FBS (Gibco; Thermo Fisher
Scientific, Inc.), and maintained in a humidified incubator at 37˚C
with 5% CO2.
Clinical tissue samples
The present study was conducted in accordance with
the Declaration of Helsinki and the protocol was approved by the
Ethics Committee of Shantou University Medical College (approval
no. SUMC-2015-42). All patients provided their oral informed
consent prior to participation in the study. The need for
additional written consent was waived by the Ethics Committee of
Shantou University, as the patients had authorized the use of their
samples in additional studies in a written consent form they signed
when donating their tissues for use in an earlier study. None of
the patients had received preoperative treatment or been diagnosed
with other types of primary tumors. In total, 50 pairs of CRC
tissues and matched adjacent normal tissues were collected from
patients (age range, 30-86 years; mean age, 60.7 years; 31 males
and 19 females) at the First Affiliated Hospital of Shantou
University Medical College between October 2015 and January 2017.
The samples were obtained during surgery following the removal of
tissue for routine pathology examination. Postoperatively, the
tumors were histologically classified and staged based on the AJCC
7th edition tumor node metastasis (TNM) system (25), and the clinical characteristics of
these patients with CRC are presented in Table I. All the tissue samples were
immediately flash frozen in liquid nitrogen and subsequently stored
at -80˚C for reverse transcription-quantitative PCR (RT-qPCR)
analysis.
 | Table IClinicopathological characteristics
of the patients with colorectal cancer (N=50). |
Table I
Clinicopathological characteristics
of the patients with colorectal cancer (N=50).
| Variable | Cases n (%) |
|---|
| Age | |
|
≤60
years | 25(50) |
|
>60
years | 25(50) |
| Sex | |
|
Male | 31(62) |
|
Female | 19(38) |
| Degree of
differentiation | |
|
Moderate to
low | 9(18) |
|
High | 38(76) |
|
Unknown | 3(6) |
| Tumor invasion | |
|
T1-2 | 6(12) |
|
T3-4 | 44(88) |
| Distant
metastasis | |
|
No | 40(80) |
|
Yes | 10(20) |
| Lymph node
metastasis | |
|
No | 32(64) |
|
Yes | 18(36) |
RT-qPCR
Total RNA was extracted from CRC tissues and cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Total
RNA was reverse transcribed into cDNA using a PrimeScript RT
reagent kit with gDNA Eraser (Takara Bio, Inc.) according to the
manufacturer's protocol. qPCR was subsequently performed using a
QuantiNova™ SYBR Green PCR mix kit (Roche Diagnostics)
and an ABI Prism 7500 system (Applied Biosystems; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
following primer pairs were used for the qPCR: KRT80 forward,
5'-CCTCCCTAATTGGCAAGGTG-3' and reverse, 5'-AGATGCCCGAGGTCGAAGAT-3';
and β-actin forward, 5'-CTGGAACGGTGAAGGTGACA-3' and reverse,
5'-AAGGGACTTCCTGTAACAATGCA-3'. The following thermocycling
conditions were used for the qPCR: Initial denaturation at 95˚C for
10 min; and 40 cycles of 95˚C for 10 sec and 60˚C for 30 sec.
Expression levels of KRT80 were quantified using the
2-∆∆Cq method for cell experiments and the
2-∆Cq method for tissue samples (26), and normalized to the loading control
β-actin.
Cell transfection
RNA interference was used to knock down KRT80
expression levels in SW480 and SW620 cells. Briefly, small
interfering RNA (siRNA) against KRT80 (siRNA-KRT80;
5'-CCCTGGATGTCAAGTTGGA-3') and siRNA-negative control (NC:
5'-ACGUGACACGUUCGGAGAATT-3') were obtained from Guangzhou RiboBio
Co., Ltd. A total of 2x105 cells/well were seeded into
six-well plates in RPMI-1640 medium, supplemented with 10% FBS.
Following overnight attachment in a humidified incubator at 37˚C
with 5% CO2, SW480 and SW620 cells were transfected with
100 nM siRNA-KRT80 or siRNA-NC using Lipofectamine RNAiMAX reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37˚C and 5%
CO2. After 48 h, cells were collected and used for RNA
extraction.
Cell Counting Kit-8 (CCK-8) assay
The CCK-8 assay was performed to determine cell
viability, as previously described (27). A total of 4x103 SW480 and
SW620 cells/well were plated into 96-well plates in RPMI-1640
medium, supplemented with 10% FBS. Following overnight attachment
in a humidified incubator at 37˚C with 5% CO2, SW480 and
SW620 cells were transfected with siRNA-KRT80 or siRNA-NC for 24,
48, 72 or 96 h. Subsequently, cell viability was detected using a
CCK-8 assay kit (Dojindo Molecular Technologies, Inc.), according
to the manufacturer's protocol. Optical density values were
measured using a microplate reader (Bio-Rad Laboratories, Inc.) at
450 nm.
Colony formation assay
For the colony formation assay, SW480 and SW620
cells were transfected with siRNA-KRT80 or siRNA-NC for 24 h,
plated at a density of 2x103 cells/well into six-well
plates and incubated in 3 ml RPMI-1640 medium at 37˚C for 10 days.
Subsequently, the cells were fixed with 4% paraformaldehyde for 30
min at 25˚C, air dried and stained with 0.1% crystal violet
(Beyotime Institute of Biotechnology) for 20 min at 25˚C. The
images of the colonies in the full well were captured with camera
and colonies with >50 cells were counted using a light
microscope [magnification, x200; (Nikon Corporation)]. To ensure
all cells were counted only once the well surface area was divided
using a 5x5 grid, the number of colonies in each segment of the
grid were counted.
5-Ethynyl 5'-deoxyuridine (EdU)
incorporation assay
EdU incorporation was performed using the Cell
Light™ Edu Apollo 567 in vitro kit (Guangzhou
RiboBio Co., Ltd.) according to the manufacturer's protocol. A
total of 5x103 SW480 and SW620 cells/well were plated
into 96-well plates in RPMI-1640 medium, containing 10% FBS.
Following overnight attachment in a humidified incubator at 37˚C
with 5% CO2, SW480 and SW620 cells were transfected with
siRNA-KRT80 or siRNA-NC for 48 h. Subsequently, the cells were
incubated with 100 µl 50 µM EdU reagents at 37˚C. Following 2 h of
incubation, the cells were fixed with 50 µl 4% paraformaldehyde for
30 min at 25˚C. Cells were washed three times in 100 µl PBS -0.1%
Triton X-100 prior to incubation with 100 µl 1x Apollo solution
(containing deionized water 93.8 µl, Apollo® reaction
buffer 5 µl, Apollo® catalyst solution 1 µl,
Apollo® fluorescent dye solution 0.3 µl and
Apollo® buffer additive 0.9 mg) for 30 min at room
temperature in the dark. The cells were subsequently stained with
100 µl 1X Hoechst 33342 nuclear dye for 30 min at room temperature.
Stained cells were visualized using a DM IL LED fluorescence
microscope (magnification, x200; Leica Microsystems GmbH), with an
excitation wavelength of 567 nm. Image analysis was performed using
ImageJ v1.8.0 software (National Institutes of Health).
Western blotting
SW480 cells were transfected with siRNA-KRT80 or
siRNA-NC for 48 hand were lysed in radioimmunoprecipitation assay
buffer (Sigma-Aldrich; Merck KGaA), supplemented with protease
inhibitors (Roche Diagnostics). Total protein was quantified using
a bicinchoninic acid assay kit (Pierce; Thermo Fisher Scientific,
Inc.) and 20 µg protein/lane was separated via 12% SDS-PAGE. The
separated proteins were subsequently transferred to PVDF membranes
(EMD Millipore) and blocked with 5% skimmed milk at room
temperature for 1 h. The membranes were incubated with the
following primary antibodies overnight at 4˚C: Anti-KRT80 (1:800;
cat. no. 16835-1-AP; ProteinTech Group, Inc.), anti-protein
phosphatase 1 catalytic subunit α (PPP1CA; 1:2,000; cat. no.
67070-1-Ig; ProteinTech Group, Inc.), anti-cyclin-dependent kinase
inhibitor 1A (p21; 1:1,000; cat. no. 2947; Cell Signaling
Technology, Inc.), anti-cyclin-dependent kinase inhibitor 1B (p27;
1:1,000; cat. no. 3686; Cell Signaling Technology, Inc.) and
anti-β-actin (1:2,500; cat. no. A5441; Sigma-Aldrich; Merck KGaA).
Following the primary antibody incubation, membranes were incubated
with a horseradish peroxidase (HRP)-conjugated anti-mouse IgG (H+L)
secondary antibody (1:5,000; cat. no. SA00001-1; ProteinTech Group,
Inc.) or an HRP-conjugated anti-rabbit IgG (H+L) secondary antibody
(1:5,000; cat. no. SA00001-2; ProteinTech Group, Inc.) at room
temperature for 2 h. Protein bands were visualized using the
SuperSignal™ West Dura Extended Duration Substrate
reagent (Thermo Fisher Scientific, Inc.) and an Amersham Imager 600
(GE Healthcare). Expression levels were quantified using ImageJ
v1.8.0 software (National Institutes of Health) and normalized to
β-actin, the loading control.
Statistical analysis
Statistical analysis was performed using SPSS 23.0
software (IBM Corp.). Statistical differences between groups were
either determined using a two-tailed unpaired Student's t-test, a
paired Student's t-test or one-way ANOVA, followed by Dunnett's
multiple comparison test. All data are presented as the mean ± SD.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Screening of DEGs identifies KRT80 as
the most upregulated gene in CRC
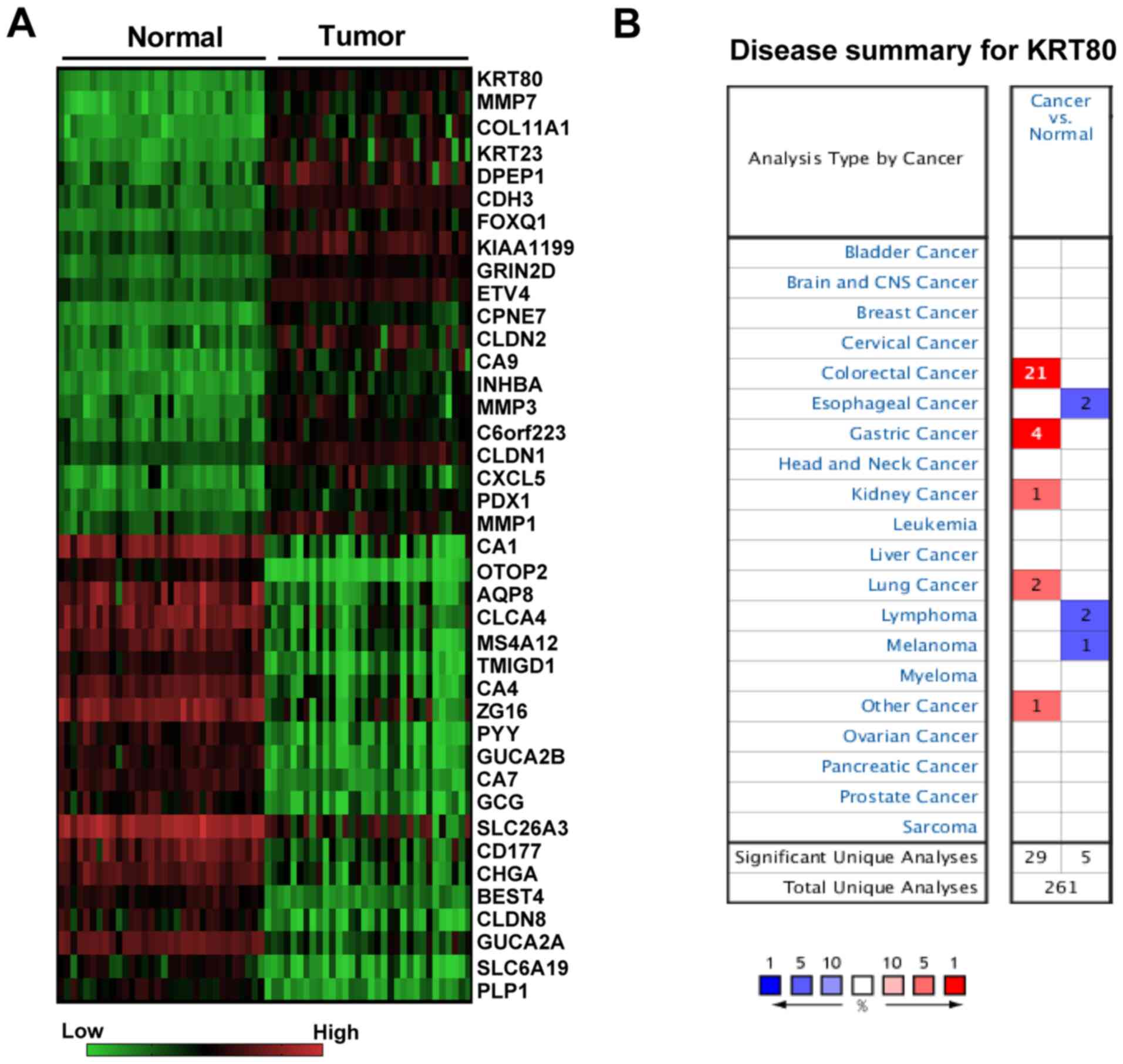
Among the CRC tissue samples from RNA-Seq datasets
in the TCGA database, only 32 CRC specimens had matching adjacent
normal tissues. Therefore, DEG analysis was only performed on these
32 pairs of tissue samples. A total of 2,114 DEGs were screened
(consisting of 844 upregulated and 1,270 downregulated DEGs)
between CRC and normal tissues, and KRT80 was discovered to be the
most upregulated gene (Fig. 1A;
Data S1). The ONCOMINE database
was further used to compare the transcriptional levels of KRT80 in
different types of cancer and the corresponding normal tissue;
compared with the normal tissues, the mRNA expression levels of
KRT80 were revealed to be significantly increased in 29 datasets
(P<0.001), including CRC, gastric, lung and kidney cancer
(Fig. 1B). Among these cancers, the
CRC dataset accounted for 72.4% (21/29) of the cases (Fig. 1B).
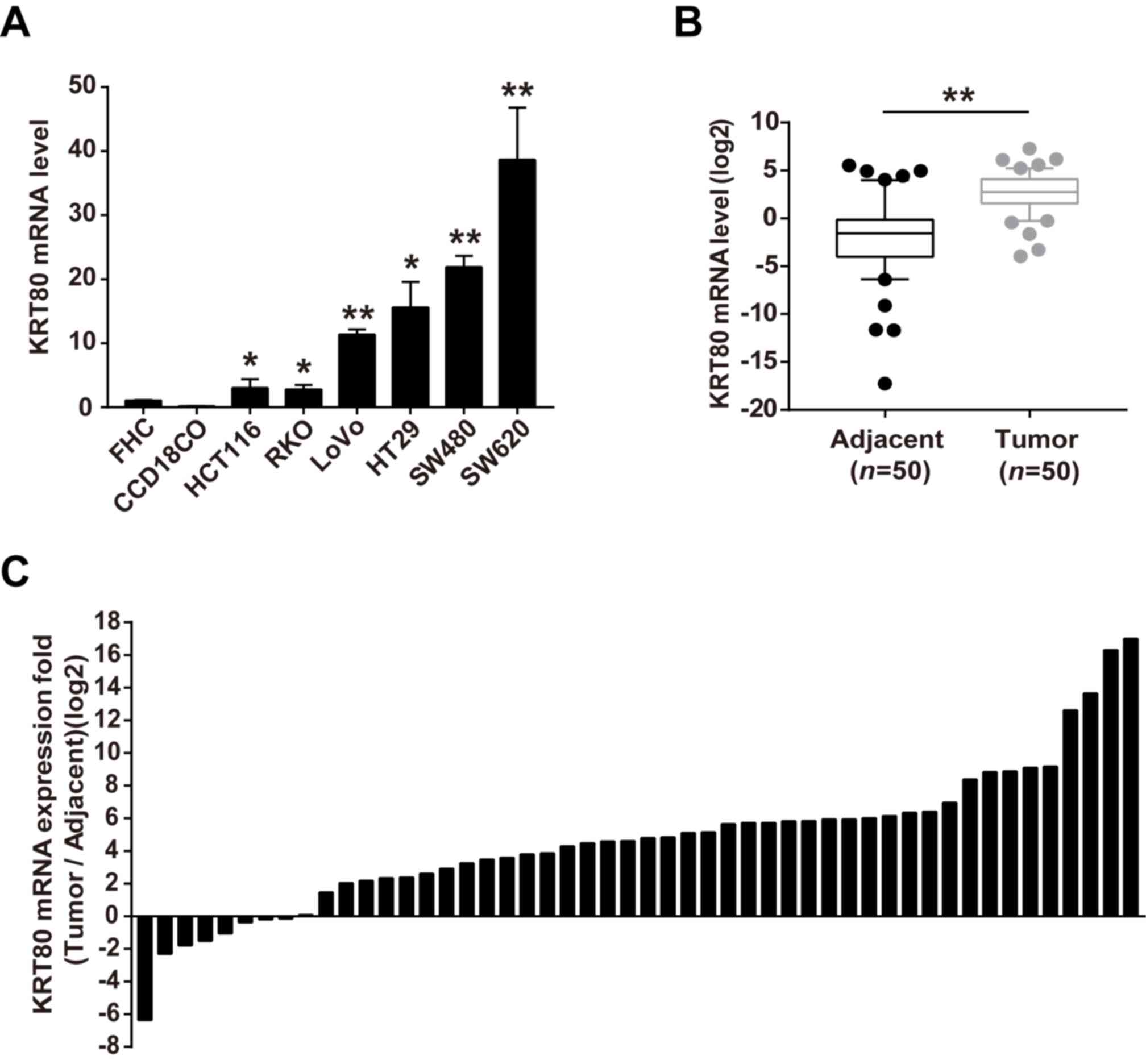
To verify the above findings, KRT80 expression
levels in CRC cell lines and cancerous tissues were further
analyzed using RT-qPCR analysis. The expression levels of KRT80
mRNA were significantly increased in the CRC cell lines HCT116,
RKO, LoVo, HT29, SW480 and SW620, compared with that noted in the
normal colorectal cells FHC and CCD18CO (Fig. 2A). Consistent with the increased
expression of KRT80 in CRC cell lines, increased expression levels
of KRT80 was also observed in CRC tissues compared with the
adjacent normal tissues (Fig. 2B).
Furthermore, KRT80 expression was upregulated in 84% (42/50) of the
patients with CRC (Fig. 2C). These
findings are consistent with a previous study (19) and suggested that KRT80 expression
levels may be increased in CRC tissues and cells.
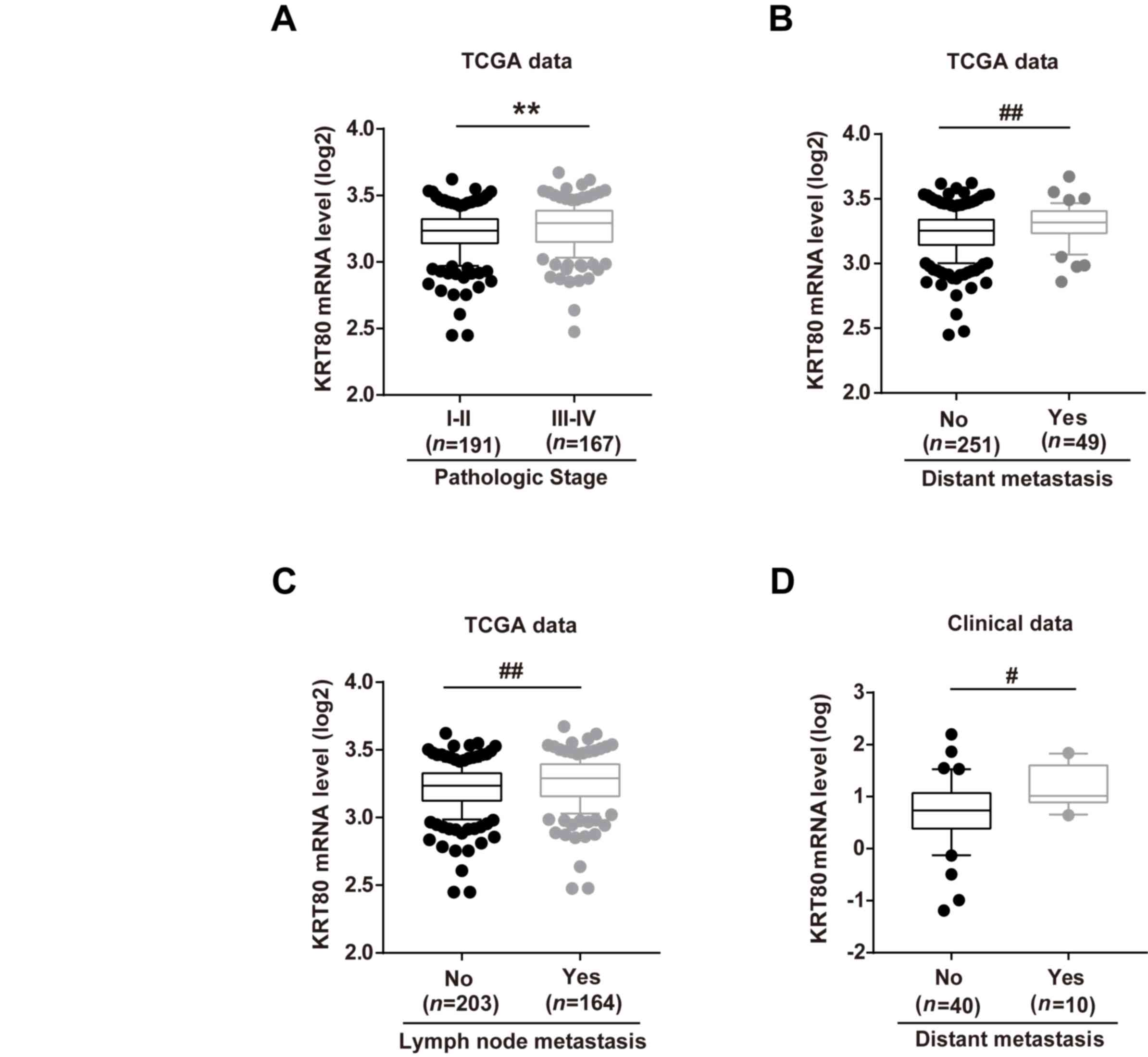
Increased KRT80 expression levels are
associated with the pathological stage and metastasis of patients
with CRC
To investigate the association between KRT80
expression levels and clinicopathological characteristics of
patients with CRC, the clinical data of patients with CRC in the
TCGA database were analyzed. The expression levels of KRT80 mRNA
were significantly increased in patients with a high pathological
stage, and both distant and lymph node metastasis compared with
patients with a low pathological stage and metastasis, respectively
(Fig. 3A-C). Moreover, clinical
tissue sample analysis from 50 patients with CRC validated the
above findings; compared with patients without distant metastasis,
KRT80 expression levels were significantly increased in patients
demonstrating metastasis (Fig.
3D).
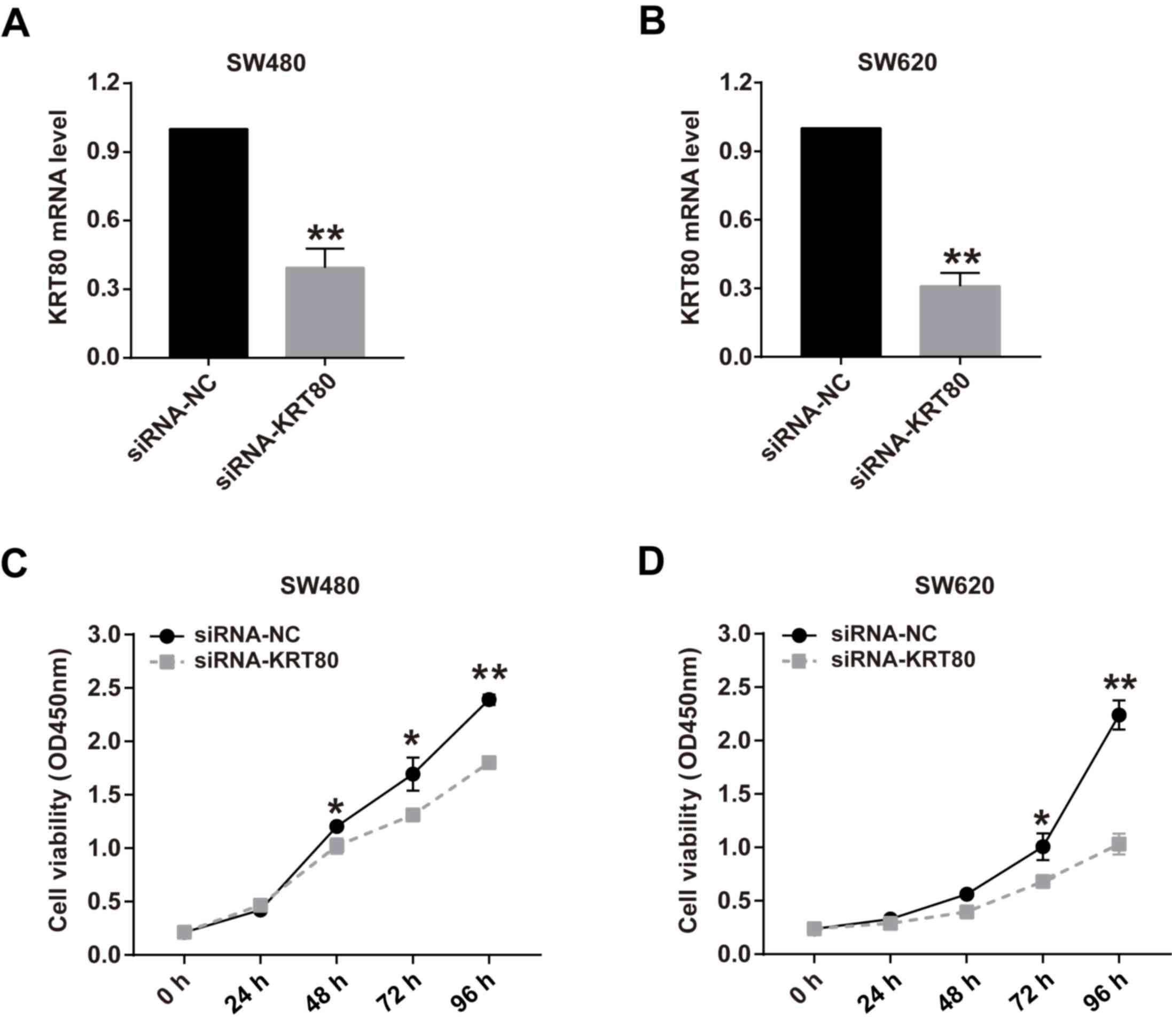
Knockdown of KRT80 expression
suppresses CRC cell proliferation
To determine the effects of KRT80 on the viability
of CRC cells, cell viability was analyzed following KRT80 knockdown
with siRNA. As hypothesized, the genetic knockdown of KRT80 using
siRNA for 48 h significantly decreased KRT80 expression levels to
~40 and 30% of the control levels in SW480 and SW620 cells,
respectively (Fig. 4A and B). The viability of SW480 cells was
significantly decreased following 48 h in siRNA-KTR80-transfected
cells compared with the siRNA-NC-transfected cells, whereas the
cell viability in siRNA-KRT80-transfected cells SW620 cells was
significantly decreased from 72 h compared with
siRNA-NC-transfected cells (Fig. 4C
and D). Moreover, the inhibitory
effect of KRT80 knockdown on cell viability was more pronounced the
longer the cells were cultured (Fig.
4C and D).
Furthermore, the effect of KRT80 on cell
proliferation was determined using colony formation and EdU
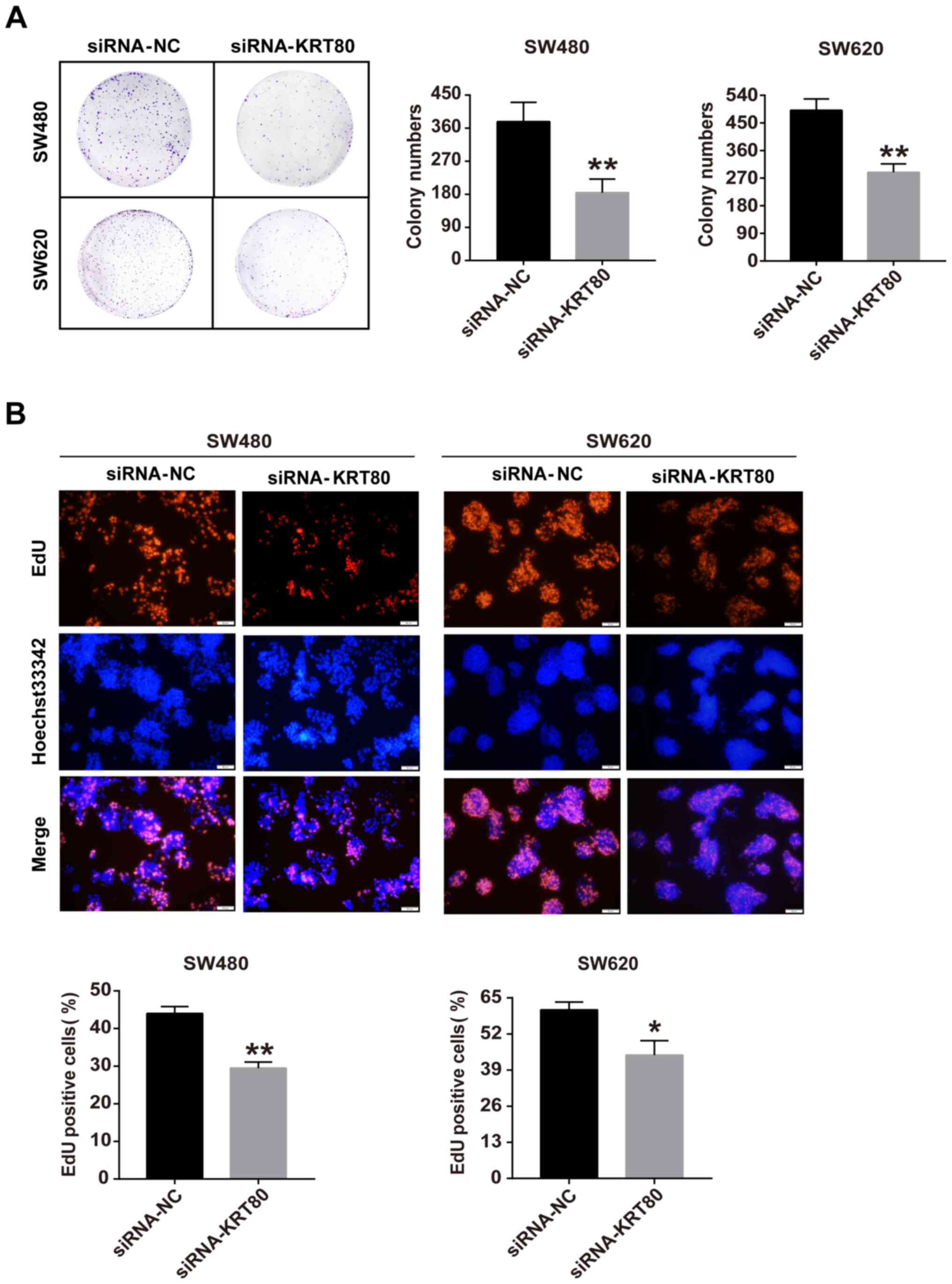
incorporation assays. The colony formation assay demonstrated that
the genetic knockdown of KRT80 significantly decreased the number
of colonies formed by ~50 and ~40% in SW480 and SW620 cells,
respectively, compared with siRNA-NC-transfected cells (Fig. 5A). The EdU assay was conducted to
assess the function of KRT80 with respect to cell proliferation.
Compared with siRNA-NC-transfected cells, the number of
EdU-positive cells was significantly reduced in both SW480 and
SW620 cells transfected with the siRNA-KRT80 compared with the
siRNA-NC-transfected cells (Fig.
5B). These data further supported the notion that KRT80 may be
involved in cell proliferation in CRC.
Pathway analysis of KRT80 co-expressed
proteins in CRC samples from TCGA
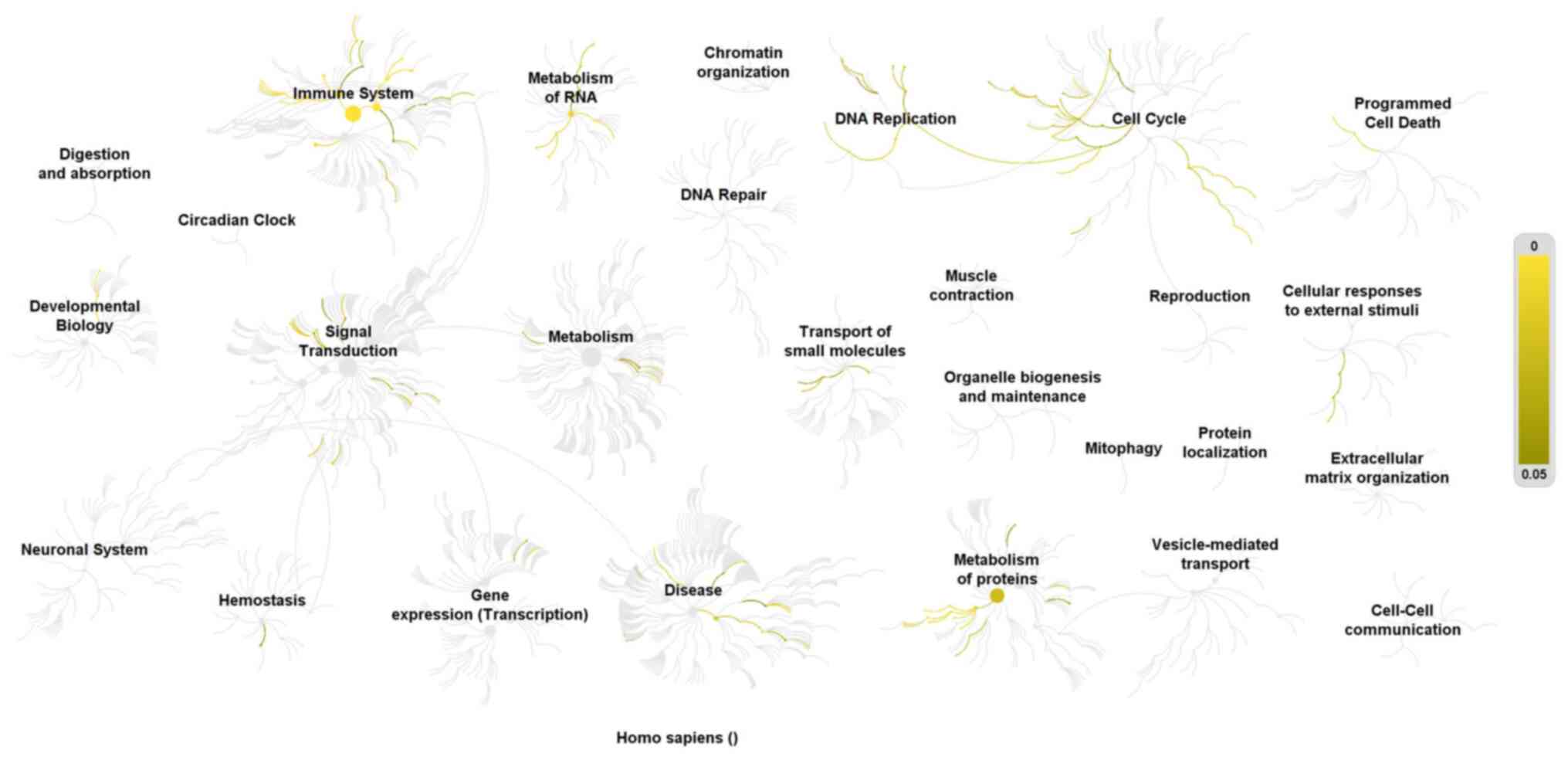
To predict the underlying mechanisms of KRT80 in CRC
cell proliferation, data mining using the cBioPortal for TCGA was
used to identify 354 proteins significantly co-expressed with KRT80
in CRC. In total, 112 co-expressed proteins with absolute
Spearman's r≥0.4 were loaded into the Reactome database for pathway
analysis (Data S2). The proteins
were enriched in the following processes: Cell cycle, DNA
replication, immune system, metabolism of RNA and proteins,
transport of small molecules and signal transduction (Fig. 6). Among them, the pathways
containing the most proteins were the cell cycle and DNA
replication pathways.
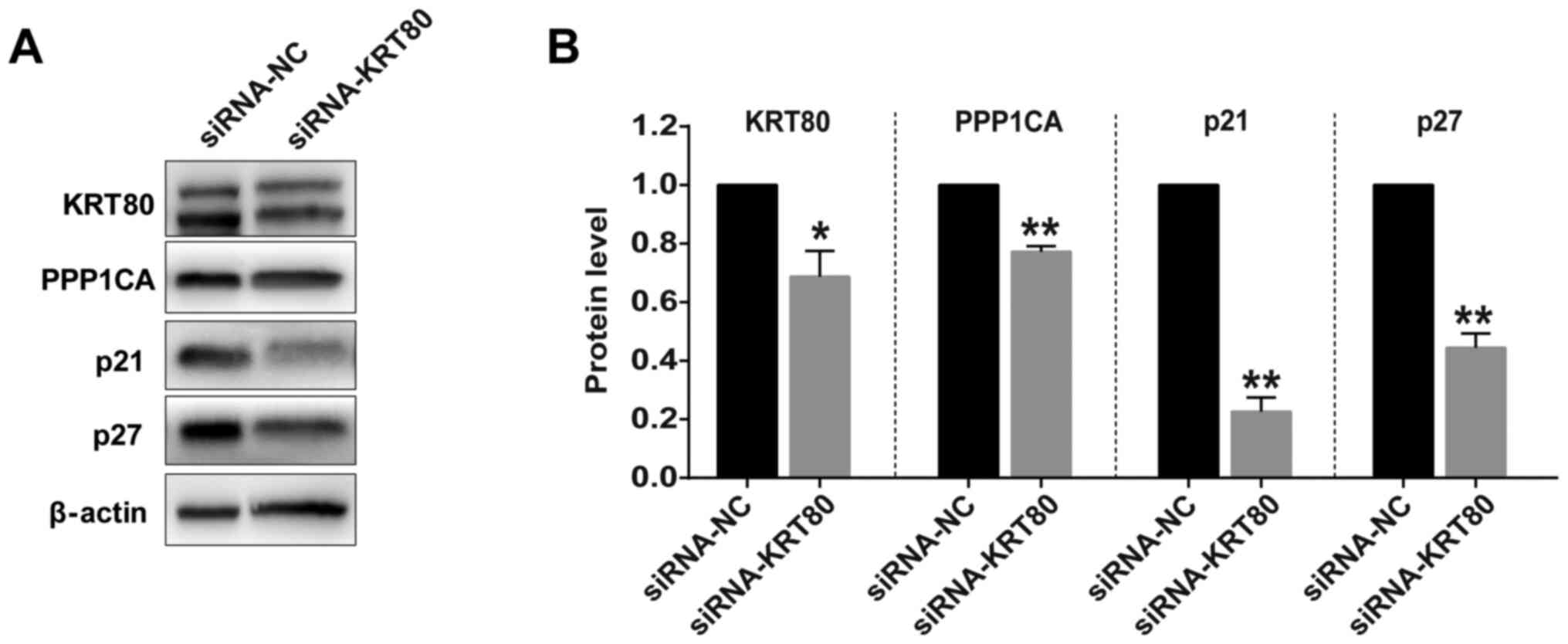
Moreover, to verify the pathway analysis results,
the expression levels of PPP1CA, which is co-expressed with KRT80
(Data S2) and involved in cell cycle and cell division (28), were analyzed in SW480 cells
following the genetic knockdown of KRT80 expression. Western
blotting discovered that the knockdown of KRT80 significantly
decreased the protein expression levels of PPP1CA in SW480 cells
compared with the siRNA-NC-transfected cells (Fig. 7). Additionally, KRT80 knockdown also
significantly reduced the expression levels of cell cycle-related
proteins, p21 and p27, compared with the siRNA-NC transfected cells
(Fig. 7).
Discussion
Keratin, as a molecular marker of epithelial cells
(10), serves an important role in
maintaining the stability and integrity of epithelial cells, and it
is also involved in various intracellular signal transduction
processes, such as cell stress, proliferation and metabolism
(29,30). Numerous studies have reported that
several proteins of the keratin family were closely related to the
development of CRC; for example, keratins were discovered to
regulate colonic epithelial cell differentiation through the Notch
1 signaling pathway (18); keratin
8 deletion-induced colitis caused a predisposition to murine CRC
through the inflammasome and IL-22 pathway (17); and increased expression levels of
KRT7 were observed in CRCs with lymph node metastasis, which was
associated with a poor prognosis (31). However, although KRT80 has been
reported to mediate migration and invasion in CRC through
activating the AKT signaling pathway (19), the role of KRT80 in CRC
proliferation remains unknown.
In the present study, the TCGA and ONCOMINE
databases were used to identify KRT80 as the most
upregulated gene in CRC compared with the normal tissues.
Furthermore, it was confirmed that KRT80 expression levels in
clinical CRC tissues and CRC cell lines were significantly
increased compared with the normal tissues and cells, and increased
KRT80 expression levels were associated with the pathological stage
and metastasis in patients with CRC. These results were consistent
with the study by Li et al (19), which demonstrated that KRT80 was
highly expressed in CRC and promoted the migration and invasion of
CRC cells, suggesting that KRT80 may serve as an oncogene to
promote the proliferation of CRC cells. To confirm this, KRT80
expression was knocked down and it was subsequently found that
reduced KRT80 expression levels decreased the cell viability,
reduced the number of colonies formed and suppressed cell
proliferation. Together with previous reports (19), these findings validated that
KRT80 may be a novel potential oncogene for CRC.
In addition, the results of the present study
revealed that proteins co-expressed with KRT80 were enriched in the
cell cycle and DNA replication processes, and the genetic knockdown
of KRT80 significantly reduced the expression levels of cell
cycle-related proteins, including PPP1CA, p21 and p27. PPP1CA,
which is co-expressed with KRT80 in CRC, is reported to be one of
three catalytic subunits of protein phosphatase 1(28); it has been demonstrated to be
involved in prostate cancer and CRC tumorigenesis via the mitogen
activated protein kinase signaling pathway (32,33).
Although p21 and p27 mediate cell cycle arrest through binding to
and inhibiting cyclin-dependent kinase/cyclin complexes, numerous
studies over the past decade have revealed that p21 and p27 also
serve an important role in carcinogenesis and tumor development
(34-39).
In fact, p21 and p27 have been reported to serve as oncogenic
proteins or tumor suppressors, depending on their localization in
the cytoplasm or the nucleus, respectively (40-43).
In brief, cytoplasmic p21 and p27 have been found to favor
antiapoptotic activities, whereas nuclear p21 and p27 have been
associated with cell cycle arrest (36,39).
Therefore, the findings of the present study provided a valuable
reference for further study to investigate the function and
underlying mechanisms of KRT80 in CRC tumorigenesis.
However, this study has some limitations. Firstly,
only 50 pairs of tissue samples were used in this study, which is a
small sample size. Additionally, some clinicopathological data
related to CRC, such as patient survival, were not included.
Finally, although pathway analysis indicated that proteins
co-expressed with KRT80 were largely enriched in the cell cycle and
DNA replication, the detailed underlying mechanisms have not been
further explored. The above limitations should be resolved in
future studies.
In conclusion, the findings of the present study
suggested that KRT80 may be overexpressed in CRC tissues and
it may function as an oncogene to promote the proliferation of CRC
cells. KRT80 co-expressed proteins were discovered to be largely
enriched in the cell cycle and DNA replication pathways, which are
related to the process of tumorigenesis. Therefore, in-depth
research into KRT80 to further reveal its functions and underlying
mechanisms in CRC may provide novel insights for the early
diagnosis and gene-targeted treatment of CRC.
Supplementary Material
Differentially expressed genes (DEGs)
screened (consisting of 844 upregulated and 1,270 downregulated
DEGs) between CRC and normal tissues.
Supplementary Data
Acknowledgements
Not applicable.
Funding
This study was supported by the Sanming Project of
Medicine in Shenzhen (grant no. SZSM201612071) and the Shenzhen
Science and Technology Project (grant no.
JCYJ20180228163436705).
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in the UCSC Xena Browser (https://xenabrowser.net/datapages/), ONCOMINE
gene expression array datasets (www.oncomine.org), cBioPortal database (http://www.cbioportal.org) and the Reactome database
(https://reactome.org/PathwayBrowser).
Authors' contributions
HY and XX designed the study; JL, XF and JC
performed the experiments and analyzed the data; XX contributed to
obtaining the patient tissue samples; and JL and HY wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All patients provided informed consent for inclusion
before they participated in the study. The study was conducted in
accordance with the Declaration of Helsinki and the study protocol
was approved by the Ethics Committee of Shantou University Medical
College (approval no. SUMC-2015-42).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mercier J and Voutsadakis IA: A systematic
review and meta-analysis of retrospective series of regorafenib for
treatment of metastatic colorectal cancer. Anticancer Res.
37:5925–5934. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Zhang Y, Lin C, Liao G, Liu S, Ding J,
Tang F, Wang Z, Liang X, Li B, Wei Y, et al: MicroRNA-506
suppresses tumor proliferation and metastasis in colon cancer by
directly targeting the oncogene EZH2. Oncotarget. 6:32586–32601.
2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Malapelle U, Mayo de-Las-Casas C, Rocco D,
Garzon M, Pisapia P, Jordana-Ariza N, Russo M, Sgariglia R, De Luca
C, Pepe F, et al: Development of a gene panel for next-generation
sequencing of clinically relevant mutations in cell-free DNA from
cancer patients. Br J Cancer. 116:802–810. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Shahjaman M, Kumar N, Ahmed MS, Begum A,
Islam SMS and Mollah MNH: Robust feature selection approach for
patient classification using gene expression data. Bioinformation.
13:327–332. 2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Strnad P, Paschke S, Jang KH and Ku NO:
Keratins: Markers and modulators of liver disease. Curr Opin
Gastroenterol. 28:209–216. 2012.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Rogers MA, Edler L, Winter H, Langbein L,
Beckmann I and Schweizer J: Characterization of new members of the
human type II keratin gene family and a general evaluation of the
keratin gene domain on chromosome 12q13.13. J Invest Dermatol.
124:536–544. 2005.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Uenishi T, Kubo S, Yamamoto T, Shuto T,
Ogawa M, Tanaka H, Tanaka S, Kaneda K and Hirohashi K: Cytokeratin
19 expression in hepatocellular carcinoma predicts early
postoperative recurrence. Cancer Sci. 94:851–857. 2003.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kurokawa I, Urakawa Y, Senba Y, Kawabata
E, Nishimura K, Omoto Y, Tokime K, Mizutani H and Tsubura A:
Keratin profiles may differ between intraepidermal and intradermal
invasive eccrine porocarcinoma. Oncol Rep. 16:473–477.
2006.PubMed/NCBI
|
|
11
|
Toivola DM, Boor P, Alam C and Strnad P:
Keratins in health and disease. Curr Opin Cell Biol. 32:73–81.
2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Omary MB, Ku NO, Strnad P and Hanada S:
Toward unraveling the complexity of simple epithelial keratins in
human disease. J Clin Invest. 119:1794–1805. 2009.PubMed/NCBI View
Article : Google Scholar
|
|
13
|
Govaere O, Komuta M, Berkers J, Spee B,
Janssen C, de Luca F, Katoonizadeh A, Wouters J, van Kempen LC,
Durnez A, et al: Keratin 19: A key role player in the invasion of
human hepatocellular carcinomas. Gut. 63:674–685. 2014.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Schweizer J, Bowden PE, Coulombe PA,
Langbein L, Lane EB, Magin TM, Maltais L, Omary MB, Parry DA,
Rogers MA and Wright MW: New consensus nomenclature for mammalian
keratins. J Cell Biol. 174:169–174. 2006.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Jacob JT, Coulombe PA, Kwan R and Omary
MB: Types I and II keratin intermediate filaments. Cold Spring Harb
Perspect Biol. 10(a018275)2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Baribault H, Penner J, Iozzo RV and
Wilson-Heiner M: Colorectal hyperplasia and inflammation in keratin
8-deficient FVB/N mice. Genes Dev. 8:2964–2973. 1994.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Misiorek JO, Lähdeniemi IAK, Nyström JH,
Paramonov VM, Gullmets JA, Saarento H, Rivero-Müller A, Husøy T,
Taimen P and Toivola DM: Keratin 8-deletion induced colitis
predisposes to murine colorectal cancer enforced by the
inflammasome and IL-22 pathway. Carcinogenesis. 37:777–786.
2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Lähdeniemi IAK, Misiorek JO, Antila CJM,
Landor SK, Stenvall CA, Fortelius LE, Bergström LK, Sahlgren C and
Toivola DM: Keratins regulate colonic epithelial cell
differentiation through the Notch1 signalling pathway. Cell Death
Differ. 24:984–996. 2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Li C, Liu X, Liu Y, Liu X, Wang R, Liao J,
Wu S, Fan J, Peng Z, Li B and Wang Z: Keratin 80 promotes migration
and invasion of colorectal carcinoma by interacting with PRKDC via
activating the AKT pathway. Cell Death Dis. 9(1009)2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43(e47)2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: A cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBioPortal. Sci Signal. 6(pl1)2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Jassal B, Matthews L, Viteri G, Gong C,
Lorente P, Fabregat A, Sidiropoulos K, Cook J, Gillespie M, Haw R,
et al: The reactome pathway knowledgebase. Nucleic Acids Res. 48
(D1):D498–D503. 2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Edge SB and Compton CC: The American joint
committee on cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474.
2010.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Wang C, Li A, Yang S, Qiao R, Zhu X and
Zhang J: CXCL5 promotes mitomycin C resistance in non-muscle
invasive bladder cancer by activating EMT and NF-κB pathway.
Biochem Biophys Res Commun. 498:862–868. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Figueiredo J, da Cruz E Silva OA and
Fardilha M: Protein phosphatase 1 and its complexes in
carcinogenesis. Curr Cancer Drug Targets. 14:2–29. 2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zhang Q, Shan G, Cao P, He J, Lin Z, Huang
Y and Ao N: Mechanical and biological properties of oxidized horn
keratin. Mater Sci Eng C Mater Biol Appl. 47:123–134.
2015.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Tang J, Zhuo H, Zhang X, Jiang R, Ji J,
Deng L, Qian X, Zhang F and Sun B: A novel biomarker Linc00974
interacting with KRT19 promotes proliferation and metastasis in
hepatocellular carcinoma. Cell Death Dis. 5(e1549)2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Czapiewski P, Bobowicz M, Pęksa R,
Skrzypski M, Gorczyński A, Szczepańska-Michalska K, Korwat A,
Jankowski M, Zegarski W, Szulgo-Paczkowska A, et al: Keratin 7
expression in lymph node metastases but not in the primary tumour
correlates with distant metastases and poor prognosis in colon
carcinoma. Pol J Pathol. 67:228–234. 2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Chen M, Wan L, Zhang J, Zhang J, Mendez L,
Clohessy JG, Berry K, Victor J, Yin Q, Zhu Y, et al: Deregulated
PP1α phosphatase activity towards MAPK activation is antagonized by
a tumor suppressive failsafe mechanism. Nat Commun.
9(159)2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Sun H, Ou B, Zhao S, Liu X, Song L, Liu X,
Wang R and Peng Z: USP11 promotes growth and metastasis of
colorectal cancer via PPP1CA-mediated activation of ERK/MAPK
signaling pathway. EBioMedicine. 48:236–247. 2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Hnit SS, Xie C, Yao M, Holst J, Bensoussan
A, De Souza P, Li Z and Dong Q: p27(Kip1) signaling:
Transcriptional and post-translational regulation. Int J Biochem
Cell Biol. 68:9–14. 2015.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Sharma SS and Pledger WJ: The
non-canonical functions of p27(Kip1) in normal and tumor biology.
Cell Cycle. 15:1189–1201. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Currier AW, Kolb EA, Gorlick RG, Roth ME,
Gopalakrishnan V and Sampson VB: p27/Kip1 functions as a tumor
suppressor and oncoprotein in osteosarcoma. Sci Rep.
9(6161)2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Phillips AH, Ou L, Gay A, Besson A and
Kriwacki RW: Mapping interactions between p27 and RhoA that
stimulate cell migration. J Mol Biol. 430:751–758. 2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Shamloo B and Usluer S: p21 in cancer
research. Cancers (Basel). 11(1178)2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Karimian A, Ahmadi Y and Yousefi B:
Multiple functions of p21 in cell cycle, apoptosis and
transcriptional regulation after DNA damage. DNA Repair (Amst).
42:63–71. 2016.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Huang Y, Wang W, Chen Y, Huang Y, Zhang J,
He S, Tan Y, Qiang F, Li A, Røe OD, et al: The opposite prognostic
significance of nuclear and cytoplasmic p21 expression in
resectable gastric cancer patients. J Gastroenterol. 49:1441–1452.
2014.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Koster R, di Pietro A, Timmer-Bosscha H,
Gibcus JH, van den Berg A, Suurmeijer AJ, Bischoff R, Gietema JA
and de Jong S: Cytoplasmic p21 expression levels determine
cisplatin resistance in human testicular cancer. J Clin Invest.
120:3594–3605. 2010.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Georgakilas AG, Martin OA and Bonner WM:
p21: A two-faced genome guardian. Trends Mol Med. 23:310–319.
2017.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Denicourt C, Saenz CC, Datnow B, Cui XS
and Dowdy SF: Relocalized p27Kip1 tumor suppressor functions as a
cytoplasmic metastatic oncogene in melanoma. Cancer Res.
67:9238–9243. 2007.PubMed/NCBI View Article : Google Scholar
|