Introduction
Sepsis is one of the leading causes of mortality in
the clinic worldwide (1). The
global incidence of sepsis is >18 million cases per year, with
the mortality rate reaching as high as 28-40% (1). Sepsis is defined as a life-threatening
organ dysfunction caused by uncontrolled inflammatory responses,
where the resulting septic liver injury and failure are proposed to
be among the major causes of mortality (1-3).
Septic liver injury has been identified as an independent risk
factor for the prediction of mortality in patients in critical
care, where previous studies have reported that alleviation of
liver injury improved the prognosis and reduced mortality in
patients with sepsis (2-4).
During sepsis, inflammatory cytokines released from
immune cells bind to receptors on hepatocytes and induce the
activation of downstream signaling, resulting in the apoptosis or
necrosis of hepatocytes (4-6).
Previous studies identified inflammasome activation in hepatocytes
as a major cause of hepatocyte cell death and of septic liver
injury and failure (7-10).
Following activation by pro-inflammatory cytokines, including
interleukin (IL)-1β and IL-6, MAPK and NF-κB signaling are
activated in hepatocytes, which promotes the expression of
pro-IL-1β and pro-caspase 1(6).
Pro-IL-1β and pro-caspase are then activated by danger signals,
including extracellular ATP or reactive oxygen species, leading to
the assembly of the nucleotide-binding oligomerization domain,
leucine rich repeat and pyrin domain containing 3 (NLRP3)
inflammasome and IL-1β production (9,11).
IL-1β in turn activates inflammatory signaling in surrounding
hepatocytes, further exacerbating injury and cell death (12). Although it has been speculated that
inflammasome activation suppression may have protective effects on
liver during sepsis, research remains insufficient (11).
Sophocarpine is a pharmaceutical monomer that was
originally derived from herbs of the Sophora species,
including Sophora flavescens and Sophora
alopecuroides, which have been applied in traditional Chinese
medicine (13). Similar extracts,
known as Kudouzi, are typically used in Chinese medicine for the
treatment of fever, inflammation and edema (13). Recent studies suggested that
sophocarpine may alleviate lipopolysaccharide (LPS)-induced liver
injury in mice through suppression of oxidative stress,
inflammation and apoptosis (13-15).
These protective effects have been proposed to be due to inhibition
of the MAPK and NF-κB pathways and reductions in the levels of
inducible nitric oxide synthase and cyclooxgenase-2(13). However, to the best of our
knowledge, the effects of sophocarpine on sepsis-induced liver
injury and inflammasome activation in hepatocytes during sepsis
remains poorly understood.
In the present study, the potential effects of
sophocarpine on septic liver injury were investigated in a
cecal-ligation and puncture (CLP) model, a classical model of
murine sepsis (4). In addition, the
possible effects of sophocarpine on inflammasome activation in the
liver and associated regulatory mechanisms were assessed.
Materials and methods
Animal model and experiments
A total of 86 male C57BL/6J mice (age, 8-10 weeks;
weight, 20-25 g) were purchased from the Qingdao Laboratory Animal
Center. The mice were raised in a specific pathogen-free room at a
temperature of 18-22˚C, humidity of 50-60% and on a 12-h light/dark
cycle. The mice received water and food ad libitum. Animal
experiments in the present study were approved by the Ethics
Committee of Qingdao Municipal Hospital (Qingdao, China). The
authors confirm that the animals received humane care and that all
animal experiments were performed in accordance with the relevant
guidelines and regulations (4). The
CLP mouse model was established as previously reported (16). Mice were administered with
sophocarpine through oral gavage for 3 days. On the day of CLP
procedure, mice were firstly treated with Bafilomycin A or MG132 2
h before the CLP procedure. Then mice were subjected to the CLP
procedure. Mice were anesthetized using 4% sevoflurane (Abbott
Pharmaceutical Co., Ltd.) and positioned on a sanitized surgical
plate. The mouse cecum was isolated and a 0.5 cm section of cecum
was ligated and punctured using a 22G sterile syringe. NC
represents the normal control group. After the CLP procedure, mice
were treated with sophocarpine for 3 more days and then sacrificed
for tissues collection. Sophocarpine treatment (30 or 60 mg/kg) was
adapted from a previous study on oral administration (13). To evaluate the effects of
sophocarpine on septic liver injury, CLP model mice were treated
with sophocarpine at 30 or 60 mg/kg. Mice that underwent the CLP
procedure were administered sophocarpine orally (oral gavage) for 2
days pre-operatively and then for 3 days following the
CLP-procedure. The mice were sacrificed on day 6 and blood and
liver samples were obtained. Bafilomycin A (10 mg/kg) (16) or MG-132 (10 µg/kg; MedChemExpress),
according to previously proposed dosages (17), were injected intraperitoneally 2 h
before the CLP procedure (these inhibitor is administered at the
operation day). After the CLP procedure, mice were administered
sophocarpine for the following 3 days. The liver was harvested on
the fourth day after the CLP procedure. For sample collection mice
were first anesthetized using 4% sevoflurane and then sacrificed by
cervical dislocation, according to a previously reported sacrifice
method (18).
Analysis of liver function
Serum levels of alanine transaminase (ALT) and
aspartate transaminase (AST) were measured to indicate liver
function using biochemical kits (AST kit, cat. no. C010-2-1; ALT
kit, cat. no. C009-2-1) supplied by Nanjing Jiancheng
Bioengineering Institute.
Western blot analysis
Liver tissues or cells were lysed in
immunoprecipitation assay buffer (1 mM EDTA, pH 8.0; 50 mM
Tris-HCl, pH 8.0; 2% sodium dodecyl sulfate; 5 mM dithiothreitol).
The protein concentration was then determined using a bicinchoninic
acid protein assay kit (Thermo Fisher Scientific, Inc.). Protein
samples (30 µg) were then subjected to 10% SDS-PAGE, transferred to
PVDF membranes (Invitrogen; Thermo Fisher Scientific, Inc.) and
blocked with 5% non-fat dry milk in phosphate-buffered saline with
Tween-20, pH 7.5 at room temperature. The membranes were then
incubated with the primary antibodies (dilution 1:2,000) for 4 h at
room temperature followed by incubation with horseradish
peroxidase-conjugated secondary antibodies (dilution 1:5,000; cat.
nos. 7074 and 7076; Cell Signaling Technology, Inc.) for 3 h at
room temperature. The primary antibodies used in the present study
were as follows: Anti-mouse NLRP3 (cat. no. ab214185; rabbit;
Abcam), pro-IL-1β (cat. no. 12426; rabbit; Cell Signaling
Technology, Inc.), caspase-1 (cat. no. AG-20B-0042-C100; mouse;
Adipogen Life Sciences, Inc.), gasdermin D (GSDMD; cat. no.
sc393656; mouse; Santa Cruz Biotechnology, Inc.), β-actin (cat. no.
ab8227; rabbit; Abcam), microtubule associated protein 1 light
chain 3 (LC3; cat. no. 2775; rabbit; Cell Signaling Technology,
Inc.) and p62 (cat. no. 5114; rabbit; Cell Signaling Technology,
Inc.). Signal intensity was determined using a Chemiluminescent
Imaging System 5200S (Tanon Science and Technology Co., Ltd.).
Hematoxylin and eosin,
immunohistochemical and immunofluorescence (IF) histochemistry
(IFHC)
Mouse liver tissues were fixed at room temperature
for 24 h with 10% formalin and processed into paraffin blocks and
sectioned into 5-µm slices. Tissues were stained with H&E for
histological analysis according to the standard protocol by
previous proposed method (8) and
subjected to microscopy by Nikon N1 (Japan). All tissues were
examined by three images and subjected to analysis.
For TUNEL staining, section slides were washed with
PBS and then subjected to dewaxing in a descending graded ethanol
series. TUNEL staining of liver sections was conducted using an
In Situ Cell Death Detection kit, according to the
manufacturer's protocol (Roche Diagnostics). The slides were then
subjected to microscopy (Nikon Corporation) for image capture.
For IFHC, the sections were deparaffinized and
hydrated and underwent antigen retrieval using a retrieval solution
(EDTA Antigen Retrieval Solution; Beijing Solarbio Science &
Technology Co., Ltd), according to the manufacturer's protocol
(Beyotime Institute of Biotechnology). Following blocking with
normal goat serum for 1 h at room temperature, the sections were
stained for NLRP3 (dilution 1:500) at room temperature for 3 h
(cat. no. ab214185; Abcam). In total, ≤3 hepatic lobules in three
sections per liver tissue from three mice of each group were
observed and evaluated by IF staining. The fluorescent secondary
antibody used was Alexa® 488-conjugated donkey
anti-rabbit at room temperature for 3 h along with a mounting
medium (NS2832, Molecular Probes; Invitrogen; Thermo Fisher
Scientific, Inc.). Nuclei were counterstained at room temperature
using DAPI for 2 min followed by washing with PBS (Sigma-Aldrich;
Merck KGaA). The slides were then subjected to fluorescence
microscopy using a Nikon N1 microscope (Nikon Corporation) at a
magnification of x200 and processed with a cooled camera and NIS
Viewer software (version 1.2; Nikon Corporation).
IL-1β and IL-6 enzyme-linked
immunosorbent assay (ELISA)
The levels of IL-1β of in liver homogenates and the
levels of IL-6 and IL-1β in serum were analyzed. The liver tissues
were first weighed and incubated in cold PBS containing 2 mM
phenylmethylsulfonyl fluoride, 0.1 mg/ml trypsin inhibitor, 10 nM
EDTA, 0.002% sodium azide and 1.0 mg/ml BSA (Beyotime Institute of
Biotechnology). The tissues were then homogenized and incubated for
2 h at 4˚C. Samples were then centrifuged at 12,000 x g for 10 min
at 4˚C, where the supernatant was subjected to the IL-1β ELISA
(mouse IL-1β/IL-1F2 Quantikine ELISA kit; cat. no. MLB00C; R&D
Systems, Inc.). Serum samples were prepared by centrifuging the
blood samples at 4˚C with 300 x g for 15 min and collected as
previous described (7). The
analysis of the IL-6 (mouse IL-6 Quantikine ELISA kit; cat. no.
M6000B) and IL-1β levels (mouse IL-1β/IL-1F2 Quantikine ELISA kit;
cat. no. MLB00C) was also performed using ELISA, according to
manufacturer's protocols (R&D Systems, Inc.).
Statistical analysis
Data in the present study are presented as the mean
± standard deviation. Student's t-tests were used where a
comparison was made between two groups. One-way ANOVA was used
where a comparison was made between multiple groups. Dunnett's post
hoc test was used to compare all groups to the control and Šidák's
multiple comparisons test was used for comparison between groups.
Statistical analysis was performed using Prism software (version 7;
GraphPad Software, Inc.). P<0.05 was considered to indicate a
statistically significant difference.
Results
Sophocarpine attenuates CLP-induced
liver injury in a dose-dependent manner
Mice that underwent CLP exhibited significantly
elevated serum AST, ALT, IL-6 and IL-1β levels compared with those
in the NC group (Fig. 1A-D).
However, in the groups treated with sophocarpine at both doses
tested (30 and 60 mg/kg), AST, ALT, IL-6 and IL-1β levels were all
found to be significantly reduced compared with those in the CLP
group, with a greater reduction observed when the higher
sophocarpine concentration was applied (Fig. 1A-D). Since hepatocyte cell death is
the primary cause of septic liver dysfunction (2), liver histology was subsequently
analyzed by H&E staining. The structure of the hepatic lobules
was found to be preserved in groups treated with sophocarpine and
damaged in those from the CLP groups (Fig. 1E). TUNEL assay, which was applied to
measure the extent of cell death, also demonstrated that
sophocarpine attenuated hepatocyte death (Fig. 1F), as increased brown staining
indicates increased cell death. These results suggested that
treatment with sophocarpine may attenuate septic liver injury.
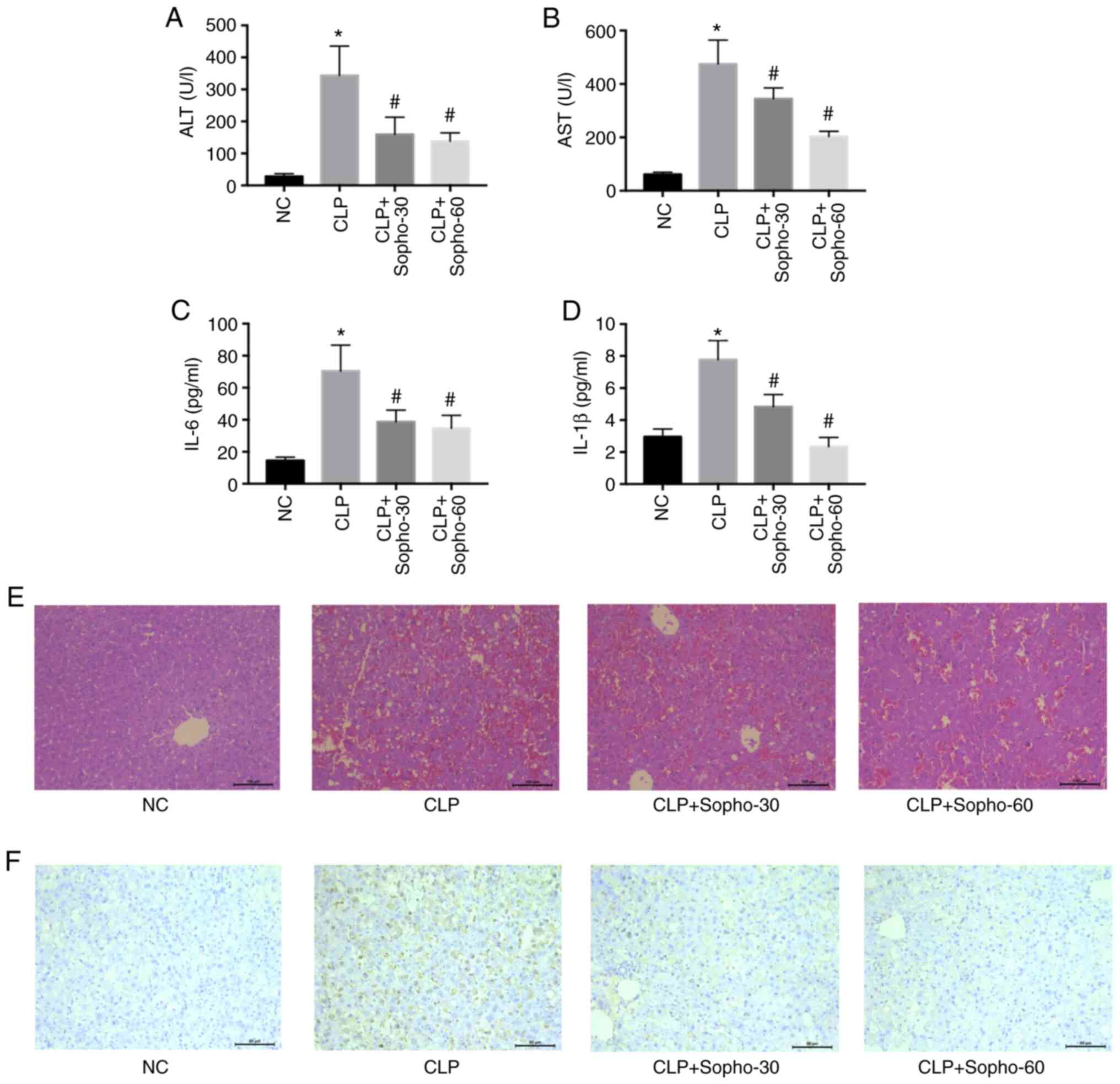 | Figure 1Sophocarpine attenuates septic liver
injury in a dose-dependent manner. Serum levels of (A) ALT, (B)
AST, (C) IL-6 and (D) IL-1β were measured in NC, CLP or mice that
underwent CLP followed by treatment with 30 or 60 mg/kg
sophocarpine. n=5 mice per group. Representative images of (E)
hematoxylin and eosin and (F) TUNEL staining of liver sections.
Scale bar, 50 µm. *P<0.05 vs. NC and
#P<0.05 vs. CLP. ALT; alanine transaminase; AST,
aspartate transaminase; CLP, cecal ligation and puncture; IL,
interleukin; NC, normal control; Sopho-30, sophocarpine treatment
30 mg/kg; Sopho-60, sophocarpine treatment 60 mg/kg. |
Sophocarpine attenuates NLRP3
inflammasome activation following CLP-induced liver injury
Previous studies have suggested that inflammasome
activation, especially that of the NLRP3 inflammasome, serves a key
role in liver injury (9,11,19),
resulting in the release of IL-1β from hepatocytes and subsequent
pyroptosis (6,11). Therefore, following the CLP
procedure and/or sophocarpine treatment, the mice were sacrificed
and the liver was harvested, following which the extent of NLRP3
activation was assessed. NLRP3 inflammasome was found to be highly
activated in the CLP group, as indicated by the elevated NLRP3
expression, activated caspase 1-p20 and elevated GSDMD-p30
(GSDMD-p30) levels compared with those in the NC group (Fig. 2A). IL-1β levels in the liver was
also found to be elevated in the CLP groups compared with those in
the NC group (Fig. 2B). However,
compared with that in the CLP group, activation of caspase 1 and
GSDMD was found to be attenuated in the sophocarpine-treated group,
with reduced levels of liver IL-1β also observed (Fig. 2A and B). To assess NLRP3 inflammasome activation
further, IFHC was used to study NLRP3, an indicator for
inflammasome activation (4) in the
liver. The results indicated that although CLP promoted NLRP3
activation in the liver in comparison to sham, this was reversed by
sophocarpine treatment (Fig. 2C).
Together, these results suggested that sophocarpine attenuated
NLRP3 inflammasome activation following CLP-induced liver
injury.
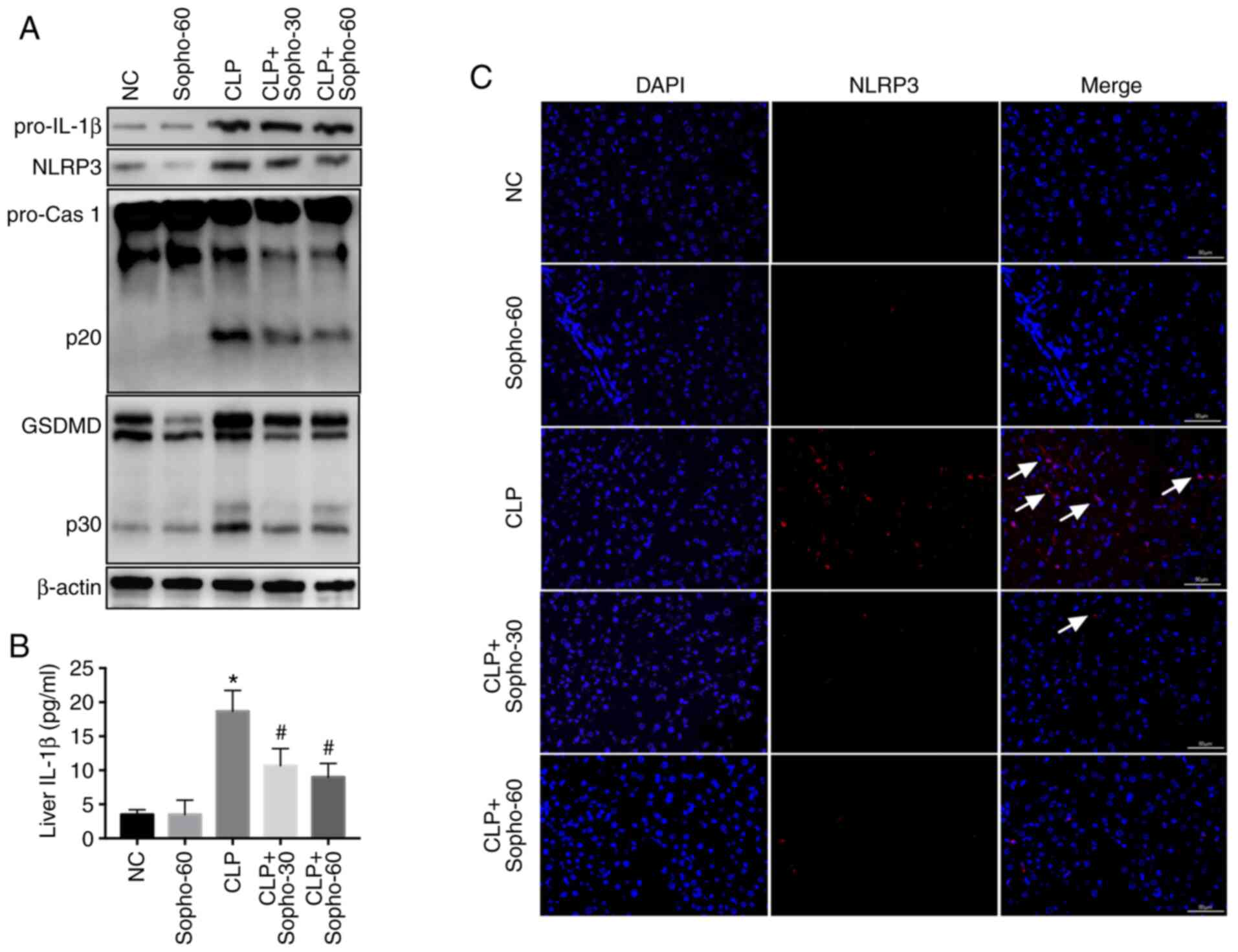 | Figure 2Sophocarpine attenuates NLRP3
inflammasome activation following CLP-induced liver injury. (A)
Representative western blotting images of pro-IL-1β, NLRP3,
pro-caspase 1 and p20, GSDMD and p30 and β-actin. (B) IL-1β
quantification in liver tissues isolated from NC, CLP model and
sophocarpine-treated mice. n=5 mice per group. (C)
Immunofluorescence of NLRP3 staining in liver tissues isolated from
mice in the respective treatment groups. Scale bar, 50 µm.
*P<0.05 vs. NC and #P<0.05 vs. CLP.
CLP, cecal ligation and puncture; cas, caspase; GSDMD, gasdermin D;
IL, interleukin; NC, normal control; Sopho-30, sophocarpine
treatment 30 mg/kg; Sopho-60, sophocarpine treatment 60 mg/kg;
NLRP3, nucleotide-binding oligomerization domain, leucine rich
repeat and pyrin domain containing 3. White arrows indicate
inflammasome activation induced by NLRP3 staining. |
Autophagy is essential for
sophocarpine-mediated attenuation of NLRP3 inflammasome activation
following CLP-induced liver injury
A previous study has indicated that sophocarpine may
promote autophagy in gastric cancer cells (20). Additionally, the NLRP3 inflammasome
may be degraded by the proteasome or the autophagic process in
macrophage (21). Therefore, the
route by which the inflammasome is degraded during
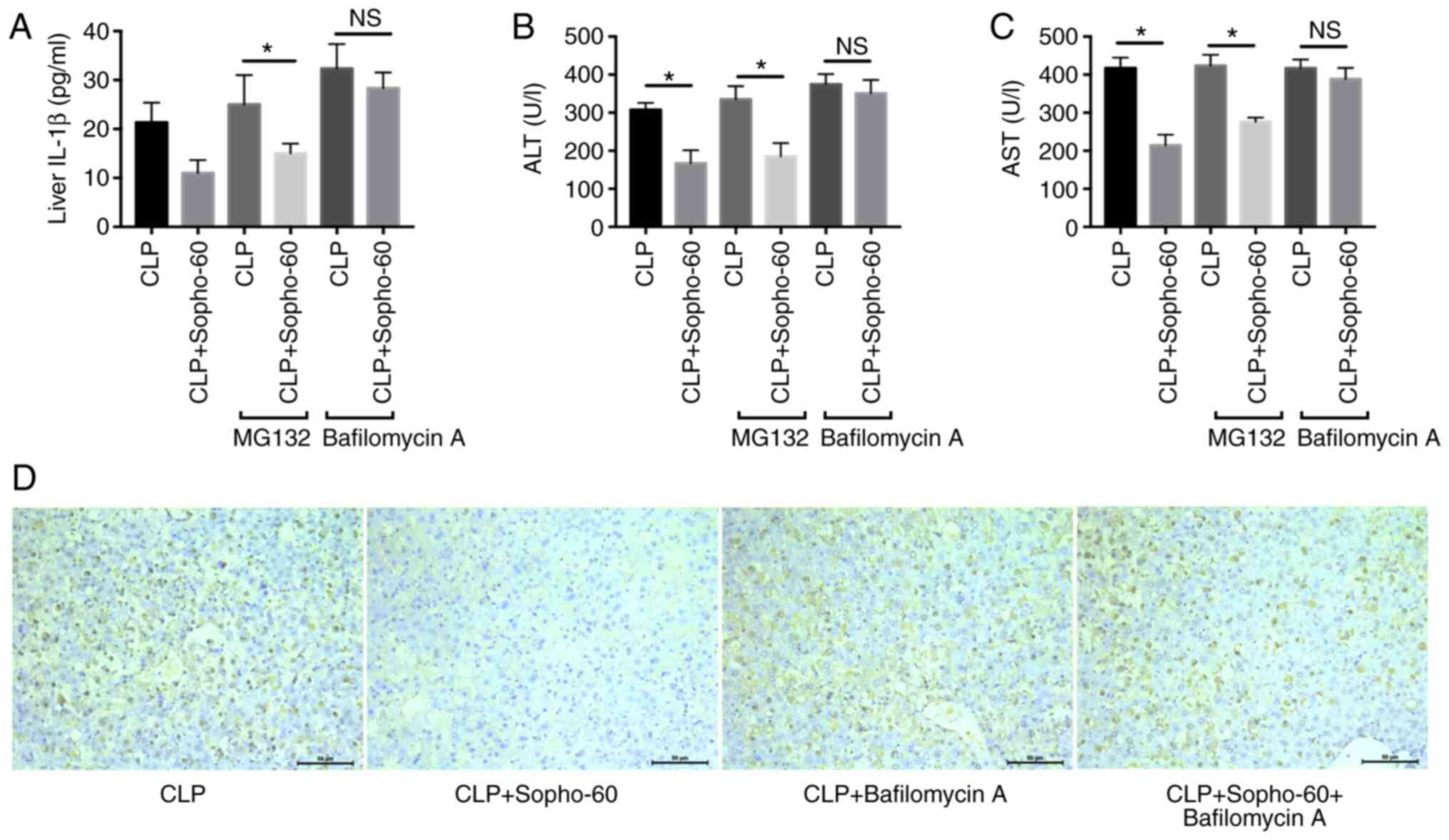
sophocarpine-induced attenuation of liver injury was assessed. As
the results indicated, pre-treatment with MG-132 did not influence
the reduction in AST and ALT as a result of sophocarpine treatment,
whilst bafilomycin A significantly reversed the protective effects
of sophocarpine in CLP-induced liver injury (Fig. 3B and C). In addition, the levels of liver IL-1β
also showed a similar tendency, in that autophagy inhibition by
bafilomycin A increased liver IL-1β levels in the
sophocarpine-treated group compared with those in the CLP group
(Fig. 3A). These results suggested
that autophagy is essential for the protective effect of
sophocarpine in CLP-induced liver injury.
The effect of autophagy on sophocarpine-mediated
NLRP3 inflammasome activation was subsequently investigated by
western blotting. Following the induction of CLP-induced liver
injury, sophocarpine attenuated the activation of NLRP3, caspase
1-p20 and GSDMD-p30 compared with that in the CLP group (Fig. 4A). By contrast following liver
injury induction, In mice treated with bafilomycin A, liver NLRP3
expression and caspase 1-p20 and GSDMD-p30 activation were found to
be expressed at markedly higher levels in the presence of
sophocarpine compared with mice treated with sophocarpine alone
(Fig. 4A). These results indicated
that inhibition of autophagy restored NLRP3 inflammasome activation
following liver injury induction, suggesting that autophagy is
essential for the sophocarpine-mediated attenuation of NLRP3
inflammasome activation during CLP-induced liver injury.
Sophocarpine was further suggested to promote the autophagy
process, through the observed elevation in LC3 levels and reduction
in p62 expression in mouse livers of mice following CLP-induced
liver injury compared with untreated mice following CLP-induced
liver injury (Fig. 4B). These
results suggested that sophocarpine may attenuate NLRP3
inflammasome activation by promoting its degradation by autophagy,
thereby exhibiting protective effects against CLP-induced liver
injury.
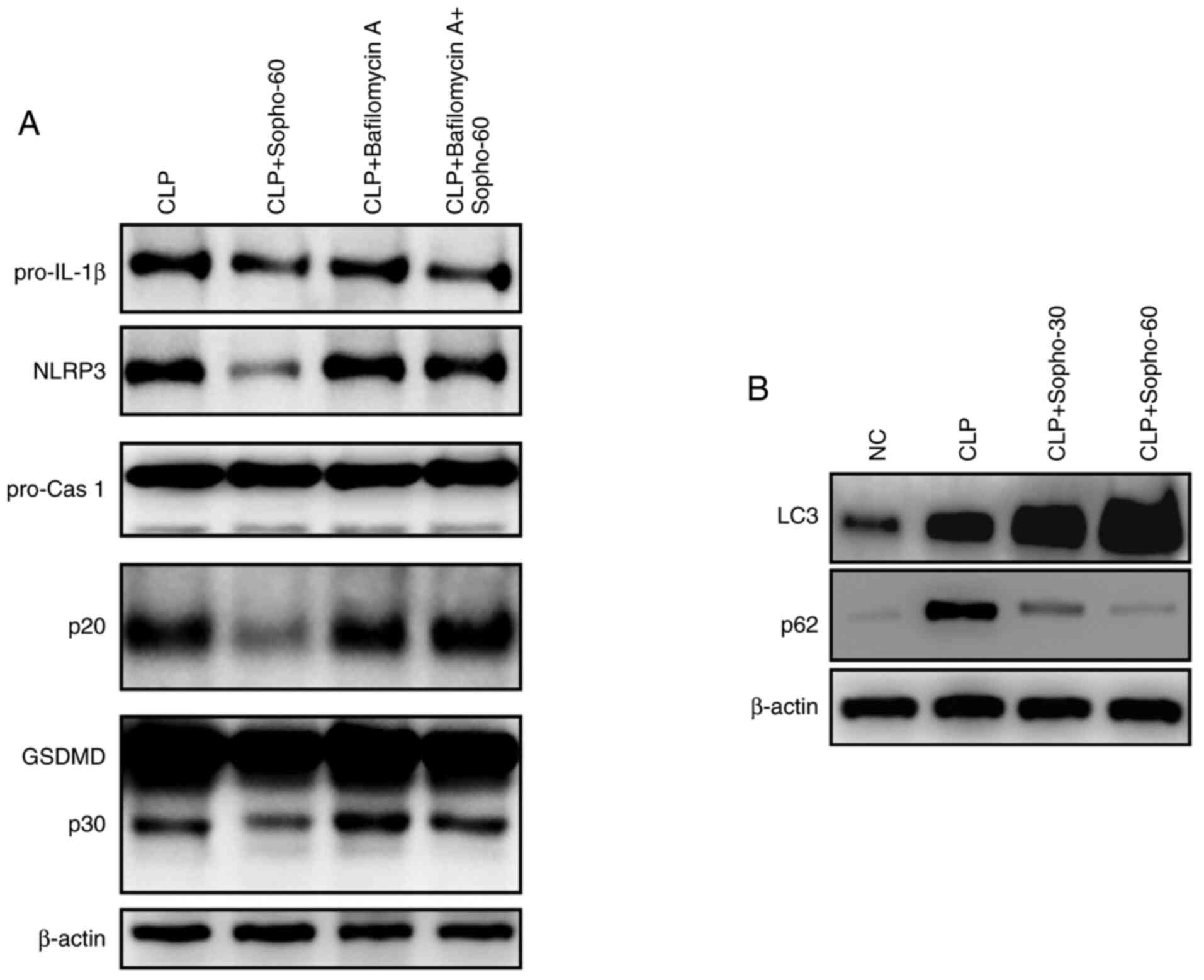 | Figure 4Autophagy is essential for the
sophocarpine-mediated attenuation of NLRP3 inflammasome activation
in CLP-induced liver injury. (A) Representative western blotting
images of pro-IL-1β, NLRP3, pro-caspase 1 and p20, GSDMD and p30
and β-actin. (B) Representative western blotting images of LC3 and
p62 in liver tissues isolated from mice that underwent CLP followed
by Sopho with or without Bafilomycin A treatment. CLP, cecal
ligation and puncture; cas, caspase; GSDMD, gasdermin D; IL,
interleukin; NC, normal control; Sopho-30, sophocarpine treatment
30 mg/kg; Sopho-60, sophocarpine treatment 60 mg/kg; NLRP3,
nucleotide-binding oligomerization domain, leucine rich repeat and
pyrin domain containing 3. |
Discussion
The updated definition of sepsis highlights organ
dysfunction as the primary cause of mortality associated with
sepsis (1,3). Uncontrolled inflammation and pathogen
invasion leads to exacerbated immune responses and activation of
the cytokine storm (1). The
cytokines released, including IL-1β and IL-6, bind to receptors on
hepatocytes and initiate cellular stress responses (2-4).
Although recent research suggest a potential protective effect of
sophocarpine on autoimmune hepatitis (14), LPS-induced liver injury (13), lupus nephritis (10) and arthritis (15), the possible effects of sophocarpine
on septic liver injury remain poorly elucidated. Results from the
present study suggested a protective effect of sophocarpine against
CLP-induced liver injury, as sophocarpine treatment resulted in
reduced AST and ALT levels in the serum and suppression of
hepatocyte cell death compared with those in untreated mice
following CLP. Previous studies have applied sophocarpine as a
pre-treatment for liver injury or LPS-induced inflammation,
typically 5-7 days prior to model establishment (10-11,13-14). In the present
study, sophocarpine was applied 2 days before the CLP procedure and
continuously applied thereafter for 3 days to investigate their
possible therapeutic effects as a perioperative medication for
clinical application.
Previous studies have demonstrated that hepatocyte
death through mechanisms including apoptosis, necrosis and
autophagy, serves a crucial role in septic liver injury (3-5).
The death of vast numbers of hepatocytes leads to elevated levels
of AST and ALT and subsequent liver failure (7). In addition, pyroptosis, a newly
discovered mechanism of cell death initiated by inflammasome
activation, has been revealed to be an essential mechanism for
liver injury and dysfunction in a number of liver injury models
(6,11). It has been reported that the
activation of inflammasome in hepatocytes participates in acute
hepatic conditions such as drug-induced liver injury and acute
hepatitis (22), in addition to
chronic liver conditions such as liver fibrosis and cancer
(23,24). The majority of studies have
identified the NLRP3 inflammasome as the main type of inflammasome
implicated in liver injury (7,9,11,25).
During cellular stress or inflammation, activated NLRP3, together
with other inflammasome elements such as apoptosis-associated
speck-like protein containing a CARD and pro-caspase 1, assemble to
form the mature NLRP3 inflammasome (11). Pro-caspase 1 becomes activated as a
result, to become cleaved caspase 1-p20 which then induces the
cleavage of IL-1 and GSDMD (26).
Cleaved GSDMD-p30 subsequently forms a membrane pore and releases
IL-1β, leading to the subsequent pyroptosis of hepatocytes
(26). In the present study,
sophocarpine suppressed the activation of the NLRP3 inflammasome,
as indicated by the reduced levels of caspase 1-p20, GSDMD-p30,
IL-1β and the expression of NLRP3 in liver tissues. These results
indicated sophocarpine may alleviate the pyroptosis of hepatocytes
during septic liver injury, suggesting that inflammasome
attenuation may be an important target of sophocarpine.
Regulation of the NLRP3 inflammasome activation
status is mediated by the regulation of its degradation. In
addition to the regulation of complex assembly, based on the ‘two
signal’ theory, during the activation phase of the inflammasome
(21,23,27),
degradation of NLRP3 also serves an essential role in determining
the fate of inflammasome activation (21). It has been reported that NLRP3 is
typically degraded through the autophagy or the proteasome pathway
(17,21). Since sophocarpine was previously
reported to promote autophagy (20), it was hypothesized in the present
study that sophocarpine may regulate NLRP3 inflammasome activation
by degradation via the autophagy pathway. The present study
suggested that the inhibitor of autophagy, bafilomycin A,
significantly reversed the protective effects of sophocarpine in
CLP-induced liver injury, whilst the proteasome inhibitor, MG-132,
exerted little effects. Following treatment with bafilomycin A, the
inhibition of NLRP3 inflammasome activation by sophocarpine was
also found to be reversed, suggesting that autophagy serve as the
major mechanism in inflammasome inhibition by sophocarpine.
There are some limitations associated with the
present study. Firstly, the protocol for the application of
sophocarpine require a more comprehensive investigation. The
present study adopted a short-term pre-treatment strategy combined
with post-treatment, which uncovered the protective effects of
sophocarpine. Future studies may adopt a single post-treatment
approach in a specific disease model to evaluate its effects, which
may provide superior indications for future clinical applications.
Although sophocarpine was found to promote NLRP3 inflammasome
degradation through autophagy, the possible effects of sophocarpine
during the activation phase should be addressed. Studies into the
regulation of inflammasome activation have focused on the
two-signal mechanism and have unveiled several important regulatory
pathways, where reactive oxygen species have been discovered to act
as an important signaling molecule (21). A previous study also suggested that
sophocarpine could alleviate oxidative stress in hepatocytes
(13). Therefore, it is reasonable
to speculate that sophocarpine may also regulate the activation of
NLRP3 inflammasome.
Collectively, the results of the present study
indicated that sophocarpine could attenuate septic liver injury and
NLRP3 inflammasome activation in hepatocytes. Sophocarpine-induced
NLRP3 degradation by autophagy may be beneficial in the alleviation
of septic liver injury.
Acknowledgements
Not applicable.
Funding
The current study was supported by the Research
Program of Qingdao Municipal Hospital (grant no. QD20190223).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
NH performed the majority of the experiments and
prepared the manuscript. XD, WL and HYa assisted with the CLP
procedures and contributed to the western blotting analysis. HYu,
JL, HL and XS assisted with animal preparation and contributed to
manuscript preparation. DA designed the study and reviewed the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Animal experiments in the present study were
approved by the Ethics Committee of Qingdao Municipal Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Singer M, Deutschman CS, Seymour CW, Hari
MS, Annane D, Bauer M, Bellomo R, Bernard GR, Chiche JD,
Coopersmith CM, et al: The Third International Consensus
Definitions for Sepsis and Septic Shock (Sepsis-3). Jama.
315:801–810. 2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Strnad P, Tacke F, Koch A and Trautwein C:
Liver-guardian, modifier and target of sepsis. Nat Rev
Gastroenterol Hepatol. 14:55–66. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Yan J and Li S and Li S: The role of the
liver in sepsis. Int Rev Immunol. 33:498–510. 2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Savio LEB, de Andrade Mello P, Figliuolo
VR, de Avelar Almeida TF, Santana PT, Oliveira SDS, Silva CLM,
Feldbrügge L, Csizmadia E, Minshall RD, et al: CD39 limits P2X7
receptor inflammatory signaling and attenuates sepsis-induced liver
injury. J Hepatol. 67:716–726. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Alhusaini A, Faddaa L, Ali HM, Hassan I,
El Orabi NF and Bassiouni Y: Amelioration of the protein expression
of Cox2, NFκB, and STAT-3 by some antioxidants in the liver of
sodium fluoride-intoxicated rats. Dose-Response.
16(1559325818800153)2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Geng Y, Ma Q, Liu YN, Peng N, Yuan FF, Li
XG, Li M, Wu YS, Li BL, Song WB, et al: Heatstroke induces liver
injury via IL-1β and HMGB1-induced pyroptosis. J Hepatol.
63:622–633. 2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Lebeaupin C, Proics E, de Bieville CH,
Rousseau D, Bonnafous S, Patouraux S, Adam G, Lavallard VJ, Rovere
C, Le Thuc O, et al: ER stress induces NLRP3 inflammasome
activation and hepatocyte death. Cell Death Dis. 6:e1879.
2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Li W, Li Y, Siraj S, Jin H, Fan Y, Yang X,
Huang X, Wang X, Wang J, Liu L, et al: FUN14 domain-containing
1-mediated mitophagy suppresses hepatocarcinogenesis by inhibition
of inflammasome activation in mice. Hepatology. 69:604–621.
2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Han CY, Rho HS, Kim A, Kim TH, Jang K, Jun
DW, Kim JW, Kim B and Kim SG: FXR inhibits endoplasmic reticulum
stress-induced NLRP3 inflammasome in hepatocytes and ameliorates
liver injury. Cell Rep. 24:2985–2999. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Li X, Wang M, Hong H, Luo C, Liu Z and
Yang R: Sophocarpine attenuates murine lupus nephritis via
inhibiting NLRP3 inflammasome and NF- κB activation. Immunol Res.
66:521–527. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zhang X, Luan J, Chen W, Fan J, Nan Y,
Wang Y, Liang Y, Meng G and Ju D: Mesoporous silica nanoparticles
induced hepatotoxicity via NLRP3 inflammasome activation and
caspase-1-dependent pyroptosis. Nanoscale. 10:9141–9152.
2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Yang CS, Kim JJ, Kim TS, Lee PY, Kim SY,
Lee HM, Shin DM, Nguyen LT, Lee MS, Jin HS, et al: Small
heterodimer partner interacts with NLRP3 and negatively regulates
activation of the NLRP3 inflammasome. Nature Commun.
6(6115)2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Jiang Z, Meng Y, Bo L, Wang C, Bian J and
Deng X: Sophocarpine attenuates LPS-induced liver injury and
improves survival of mice through suppressing oxidative stress,
inflammation, and apoptosis. Mediators Inflamm.
2018(5871431)2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Sang XX, Wang RL, Zhang CE, Liu SJ, Shen
HH, Guo YM, Zhang YM, Niu M, Wang JB, Bai ZF and Xiao XH:
Sophocarpine protects mice from ConA-induced hepatitis via
inhibition of the IFN-Gamma/STAT1 pathway. Front Pharmacol.
8(140)2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Zhu L and Zhu L: Sophocarpine suppress
inflammatory response in human fibroblast-like synoviocytes and in
mice with collagen-induced arthritis. Eur Cytokine Netw.
28:120–126. 2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Jiang Z, Bo L, Meng Y, Wang C, Chen T,
Wang C, Yu X and Deng X: Overexpression of homeodomain-interacting
protein kinase 2 (HIPK2) attenuates sepsis-mediated liver injury by
restoring autophagy. Cell Death Dis. 9(847)2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Harhouri K, Navarro C, Depetris D, Mattei
MG, Nissan X, Cau P, De Sandre-Giovannoli A and Lévy N:
MG132-induced progerin clearance is mediated by autophagy
activation and splicing regulation. EMBO Mol Med. 9:1294–1313.
2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Roustan A, Perrin J, Berthelot-Ricou A,
Lopez E, Botta A and Courbiere B: Evaluating methods of mouse
euthanasia on the oocyte quality: Cervical dislocation versus
isoflurane inhalation. Lab Anim. 46:167–169. 2012.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Gehrke N, Hövelmeyer N, Waisman A, Straub
BK, Weinmann-Menke J, Wörns MA, Galle PR and Schattenberg JM:
Hepatocyte-specific deletion of IL1-RI attenuates liver injury by
blocking IL-1 driven autoinflammation. J Hepatol. 68:986–995.
2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Huang Y, Chen X, Guo G, Guo W, Ma Q and
Yuan J: Sophocarpine inhibits the growth of gastric cancer cells
via autophagy and apoptosis. Front Biosci (Landmark Ed).
24:616–627. 2019.PubMed/NCBI
|
|
21
|
Xu M, Jiang Z, Wang C, Li N, Bo L, Zha Y,
Bian J, Zhang Y and Deng X: Acetate attenuates inflammasome
activation through GPR43-mediated Ca2+-dependent NLRP3
ubiquitination. Exp Mol Med. 51(1)2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Woolbright BL and Jaeschke H: Role of the
inflammasome in acetaminophen-induced liver injury and acute liver
failure. J Hepatol. 66:836–848. 2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Luan J and Ju D: Inflammasome: A
double-edged sword in liver diseases. Front Immunol.
9(2201)2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Alegre F, Pelegrin P and Feldstein AE:
Inflammasomes in liver fibrosis. Semin Liver Dis. 37:119–127.
2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Zheng T, Yang X, Li W, Chen L, Wu Dan,
Bian F, Xing S and Jin S: Salidroside attenuates high-fat
diet-induced nonalcoholic fatty liver disease via AMPK-dependent
TXNIP/NLRP3 pathway. Oxid Med Cell Longev.
2018(8597897)2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Shi J, Zhao Y, Wang K, Shi X, Wang Y,
Huang H, Zhuang Y, Cai Tao, Wang F and Shao F: Cleavage of GSDMD by
inflammatory caspases determines pyroptotic cell death. Nature.
526:660–665. 2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Kayagaki N, Stowe IB, Lee BL, O'Rourke K,
Anderson K, Warming S, Cuellar T, Haley B, Roose-Girma M, Phung QT,
et al: Caspase-11 cleaves gasdermin D for non-canonical
inflammasome signalling. Nature. 526:666–671. 2015.PubMed/NCBI View Article : Google Scholar
|