Introduction
Bone remodeling is a dynamic process orchestrated by
bone-forming osteoblasts and bone-resorbing osteoclasts (1). The bone formation process involves the
activation of different signaling pathways, which modulate multiple
cellular and molecular events during osteoblast proliferation,
differentiation and mineralization stages (1). Genetic studies in both humans and mice
have revealed that the Wnt/β-catenin signaling pathway is an
important mechanism for stimulating osteoblasts function (2).
The Wnt/β-catenin signaling pathway is activated by
the binding of Wnt proteins with the Frizzled (Fzd)/low-density
lipoprotein receptor-related protein (Lrp) (5/6) receptor complex.
The formation of this receptor complex induces the cytoplasmic
accumulation of β-catenin molecules and their nuclear translocation
for the interaction with the T-cell specific transcription factor
(Tcf)/lymphoid-enhancer binding factor (Lef) transcription factors,
as well as the induction of expression of target genes, such as
Lef1, Tcf7, NKD inhibitor of WNT signaling pathway 2 and
anxin 2 (AXIN2) (2,3). In
addition, the induction of AXIN2, a negative regulator of
the signaling pathway, has been proposed to act at the
transcriptional level and may create a negative feedback loop to
silence the Wnt/β-catenin signaling pathway (4).
Under basal conditions, when the Wnt pathway is
inactive, β-catenin is phosphorylated by glycogen synthase kinase 3
(GSK3), which forms part of a complex integrated by AXIN2 and the
protein of the colon APC regulator of WNT signaling pathway (APC)
gene, to be subsequently degraded in the proteasome (5). Therefore, the intracellular levels of
β-catenin are kept relatively low. However, when the Wnt pathway is
activated by the binding of the Wnt ligands to the Fxd/Lrp5/6
receptor complex, the decomposition of the intracellular
AXIN2-APC-GSK3 complex is activated, which results in the
inhibition of the phosphorylation of β-catenin (5). The hypophosphorylated β-catenin
accumulates in the cytoplasm and translocates to the nucleus, where
it regulates gene expression via the activation of various
transcription factors, such as Tcf/Lef1(6). The Wnt/β-catenin signaling pathway is
also regulated by several different extracellular antagonists, such
as the family of secreted Fzd-related proteins 1-4 (SFRPs 1-4), the
four members of the Dickkopf family (DKK 1-4) and the inhibitor Wnt
inhibitory factor 1 (WIF1) (7).
These molecules can act as decoys to compete with the Wnt ligands
to bind with the receptors; for example, some SFRPs are secreted
into the extracellular medium. Furthermore, members within the DKK
protein family, in which type 1 (DKK-1) is particularly important
in bone, can antagonize Wnt signals by binding with the Lrp5/6
co-receptors (4).
The roles of the Wnt signaling-related antagonists
and their effect on bone metabolism and osteoblasts activities have
not yet been fully elucidated. However, previous research suggests
that they are involved in the regulation of osteoblast functions.
For instance, studies in mice have reported that deletion of
SFRP1 or elimination of one allele of DKK1,
stimulates osteoblast proliferation and bone mass formation via the
activation of the Wnt signaling pathway (8,9).
Another study revealed that administration of recombinant SFRP2 or
SFRP4 proteins enhanced the alkaline phosphatase (ALP) activity in
mouse mesenchymal C3H10T1/2 cells, suggesting a role for SFRPs in
osteoblastogenesis (9). Moreover,
the overexpression of WIF1 in murine embryonic mesenchymal
cells, inhibits osteoblast differentiation (10), and knockdown of DKK1 and
DKK2 decreases matrix mineralization in KS483 mesenchymal
stem cells (11). Similarly,
clinical studies have observed an association between genetic
variants in SFRP1 and SFRP4 and bone mass content in
postmenopausal women (12,13). Taken together, these findings
indicate that the Wnt signaling-related extracellular antagonists
influence the osteoblast maturation process and bone formation, and
that they can serve as potential targets to prevent the loss of
bone mass. However, the expression levels of Wnt antagonists during
the human osteoblast proliferation and differentiation stages
remain unknown.
The aim of the present study was to investigate the
dynamics of gene expression of extracellular antagonists, SFRP
1-4, DKK 1-4 and WIF1, during the proliferation
and cell-differentiation stages of osteoblasts maturation. The hFOB
1.19 normal osteoblasts and the MG-63 and Saos-2 osteosarcoma cell
lines were used as a model system. ALP activity and the expression
levels of osterix (OSX) and RUNX family transcription factor
2 (RUNX2), which are markers of the early stage of
osteoblast differentiation, were measured (14). In addition, AXIN2 expression
was investigated to determine the activation status of the
Wnt/β-catenin signaling pathway during the transition of
osteoblasts, from proliferation to differentiation stage.
Materials and methods
Cell culture
The hFOB 1.19 (cat. no. CRL-11372), MG-63 (cat. no.
CRL-1427) and Saos-2 (cat. no. HTB-85) cell lines were purchased
from the American Type Culture Collection. hFOB 1.19 cells were
maintained in DMEM/F-12 culture medium without phenol red
(Sigma-Aldrich; Merck KGaA) and supplemented with 10% FBS (Biowest)
and 0.3 mg/ml G418 (Sigma-Aldrich; Merck KGaA) at 37˚C and 5%
CO2. MG-63 and Saos-2 cells were maintained in EMEM and
McCoy's 5A (both Sigma-Aldrich; Merck KGaA), respectively,
supplemented with 10% FBS (Biowest) and an antibiotic solution
(penicillin and streptomycin, both 100 mg/ml; Gibco; Thermo Fisher
Scientific, Inc.) at 37˚C and 5% CO2.
Cell viability and ALP activity
assays
Cell viability and ALP activity were assessed in the
hFOB 1.19, MG-63 and Saos-2 cell lines at 1, 3, 8, 15 and 21 days
of cell culture. Cells were plated at a density of
70x104 cells/dish in their respective medium and
cultured at 37˚C in a humidified incubator with 5% CO2
for the aforementioned time points. The culture media were removed
and replaced with fresh medium every other day. The media were
replaced with osteogenic media (100 µg/ml ascorbic acid and 5 mM
β-glycerol phosphate) to maintain the osteoblast phenotype from day
8 onwards. Previous studies have reported that the decline in
viability, which occurs after 8 days of cell culture, is essential
to stimulate the osteoblast differentiation-related activities
(15). Subsequently, the medium for
hFOB 1.19 cells was replaced with culture medium supplemented with
0.01 µM menadione, 100 µg/ml ascorbic acid and 5 mM β-glycerol
phosphate (16,17) (all Sigma-Aldrich; Merck KGaA). The
medium for the MG-63 and Saos-2 cell lines was replaced by their
respective medium supplemented with 100 µg/ml ascorbic acid and 5
mM β-glycerol phosphate (all Sigma-Aldrich; Merck KGaA).
In order to test the cell viability, the cells were
harvested at the indicated times using a 0.05% trypsin/EDTA
solution (GIBCO) for 2-3 min at 37˚C to detach the cells from the
adherent substrate, the cell suspension was washed twice and
resuspended in 1 ml of cold phosphate buffer saline (PBS) (4˚C).
One part of the cell suspension was mixed with one part of 0.4%
trypan blue, incubated for 3 min at room temperature and then
visually examined to determine whether cells take up or exclude the
dye, for direct identification and enumeration of live (unstained)
and dead (blue) cells in a given population. and counted on a light
inverted microscope using the 20x objective. The experiments were
performed in triplicate and the cell viability was expressed as the
cell number.
To determine the ALP activity, at the aforementioned
time points, the cells were lysed with a solution containing 0.1 M
Tris-HCl and 0.1% Tween-20 (pH 7.5) for 2 min on ice. Cell lysates
were freeze-thawed (-70˚C/ice) twice and enzyme activity was
determined using the Lowry method (18) using p-nitrophenyl phosphate
(Sigma-Aldrich; Merck KGaA) as the substrate. Protein concentration
was determined using the Bradford method, using the Bio-Rad Protein
Assay Dye Reagent Concentrate (Bio-Rad Laboratories, Inc.) with
dilution from 0.2 to 0.9 mg/ml of BSA for the standard curve,
following the manufacturer's protocol. All the experiments were
performed three times in triplicate and results are expressed as
enzymatic activity U/mg protein/min.
Gene expression studies
At the end of each incubation time point (1, 3, 8,
15 and 21 days of cell culture), the expression levels of
RUNX2, OSX and AXIN2, as well as those of the
extracellular antagonists were determined using reverse
transcription-quantitative PCR (RT-qPCR). Total RNA was extracted
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to manufacturer's instructions. cDNA
was transcribed from 1 µg total RNA using the TaqMan®
Reverse Transcription Reagents kit (Applied Biosystems; Thermo
Fisher Scientific, Inc.), according to the manufacturer's
instructions under the following conditions: 5 min at 65˚C, 2 min
at 4˚C, 30 min at 37˚C, and 5 min, 95˚C. qPCR was performed using
the following TaqMan gene expression assays (Applied Biosystems;
Thermo Fisher Scientific, Inc.): RUNX2 (assay ID,
Hs00231692_m1), OSX (assay ID, Hs01866874_s1) SFRP1
(assay ID, Hs00610060_m1), SFRP2 (assay ID, Hs00293258_m1),
SFRP3 (assay ID, Hs0017350_m1), SFRP4 (assay ID,
Hs00180066_m1), DKK1 (assay ID, Hs00183740_m1), DKK2
(assay ID, Hs00205294_m1), DKK3 (assay ID, Hs00183740_m1),
DKK4 (assay ID, Hs00205290_m1), WIF1 (assay ID,
Hs00183662_m1) and AXIN2 (assay ID, Hs00610344_m1) under
universal cycling conditions (10 min at 95˚C; 15 sec at 95˚C, 1 min
60˚C, 40 cycles). PCR amplification was performed in triplicate
using a QuantStudio™ 7 Flex Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Gene expression was
normalized using GAPDH, and relative expression was
calculated using the
2-ΔΔCq method (19).
Statistical analysis
Normal distribution was analyzed using the
Shapiro-Wilk test and homogeneity of variance was determined using
the Levene's test. Statistical differences were examined using a
one-way ANOVA followed by Tukey's post hoc test. Correlations were
evaluated using the Spearman correlation coefficient
(rs). P<0.05 was considered to indicate a
statistically significant difference. All analyses were performed
using SPSS v20.0 software (IBM Corp). All assays were performed in
triplicate.
Results
Cell viability and
differentiation
To distinguish between the osteoblast proliferation
and differentiation stages, the viability of the hFOB 1.19, MG-63
and Saos-2 osteoblastic cell lines during 21 days of cell culture
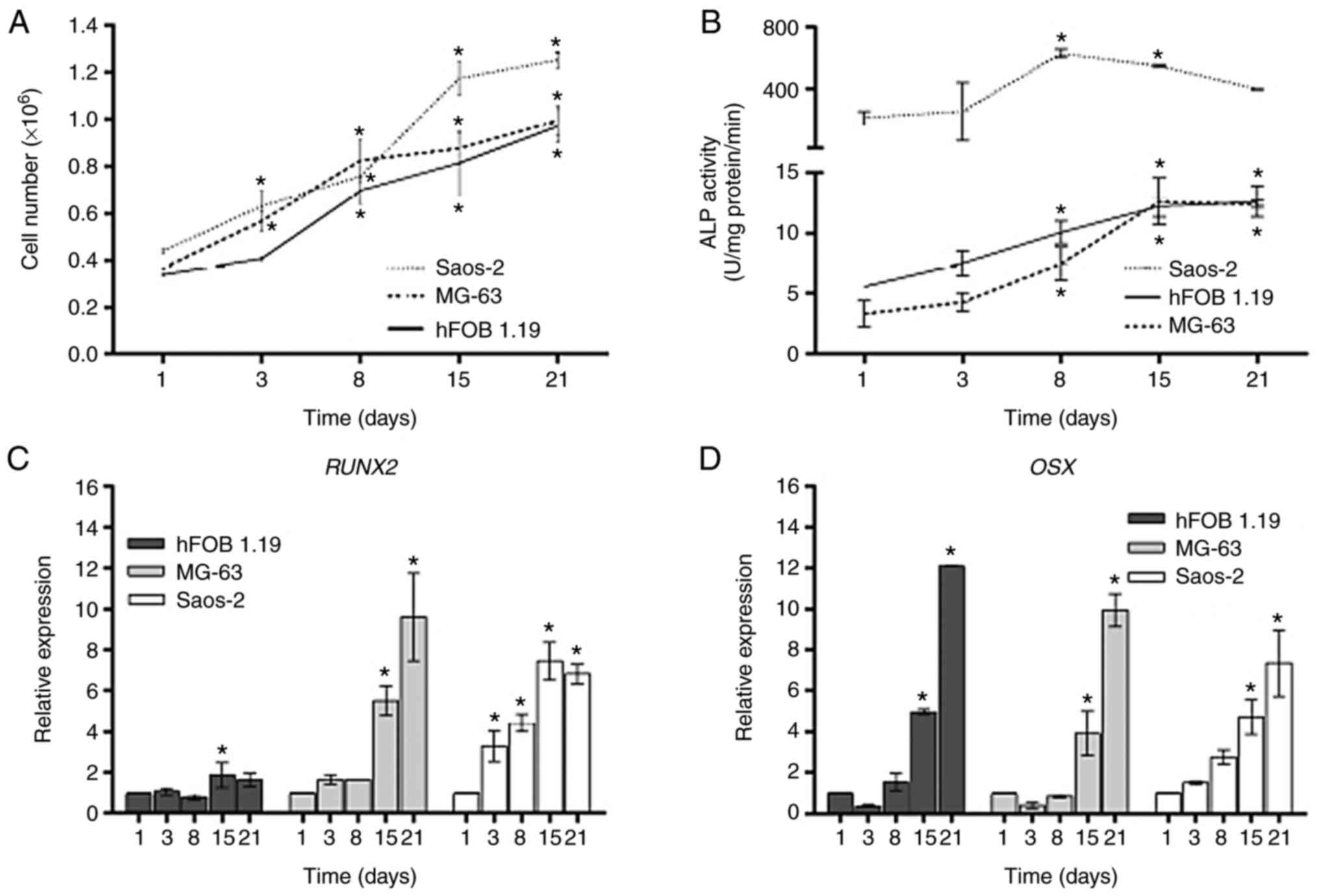
was determined. The three cell lines exhibited distinct viability
rates (Fig. 1A; Tables SI and SII). As expected, the Saos-2 cell line
exhibited the highest viability rate (25.4-fold), followed by MG-63
(17.5-fold), while the hFOB 1.19 cell line had the lowest rate
(13.5-fold). After day 8, the densities of the three osteoblastic
cell lines increased significantly as compared with the respective
day 1, in which the hFOB 1.19 and MG-63 cells reached saturation
densities after 15 days of culture. It was observed that the
cellular proliferation between day 15 and 21 was not
significant.
To identify the osteoblast differentiation stage,
ALP activity and the expression levels of the transcription factors
RUNX2 and OSX, which are three well-known markers of
osteoblast differentiation (20,21),
were assessed during 21 days of cell culture. The ALP activity at
the different time points is presented in Fig. 1B. It was found that the levels of
ALP activity in the hFOB1.19, MG-63 and Saos-2 cell lines were
correlated with their corresponding cells in terms of viability
(Tables SI and SII). The highest level of ALP activity,
expressed as units of enzyme activity, was found in the Saos-2 cell
line since early days of culture and was maintained until day 21.
Notably, the ALP activity level was similar in the hFOB 1.19 and
MG-63 cell lines during 21 days of culture, in which the enzymatic
activity increased in a time-dependent manner, reaching maximal
levels at day 15 of cell culture compared to the respective day 1.
However, the enzyme activity in Saos-2 cells was much higher
compared with that other two cell lines, which had lower viability
(Fig. 1B).
The expression levels of RUNX2 and OSX
in the hFOB 1.19, MG-63 and Saos-2 cells at days 1, 3, 8, 15 and 21
of cell culture are illustrated in Fig.
1C and D, respectively. The
expression profiles of RUNX2 in MG-63 and Saos-2 cells
demonstrated a strong positive correlation between both cell lines,
and the expression levels increased significantly in a
time-dependent manner during the 21 days of culture, when compared
with the day 1 of culture. By contrast, there was no significant
increase in the expression levels of RUNX2 during the same
days of culture in the hFOB 1.19 cell line (Fig. 1C). With respect to OSX,
similar expression profiles were observed in the three cell lines
(Table SIII).
High expression levels of OSX were observed
at day 15 and reached maximum levels at day 21 in the three
osteoblastic cell lines (Fig. 1D).
These results indicated that the differentiation stage in
osteoblast-like cell lines can be established from day 15 under the
conditions used in the present study. The potential relationship
existing between osteoblast markers is presented in Table SI.
Gene expression levels of Wnt-related
antagonists
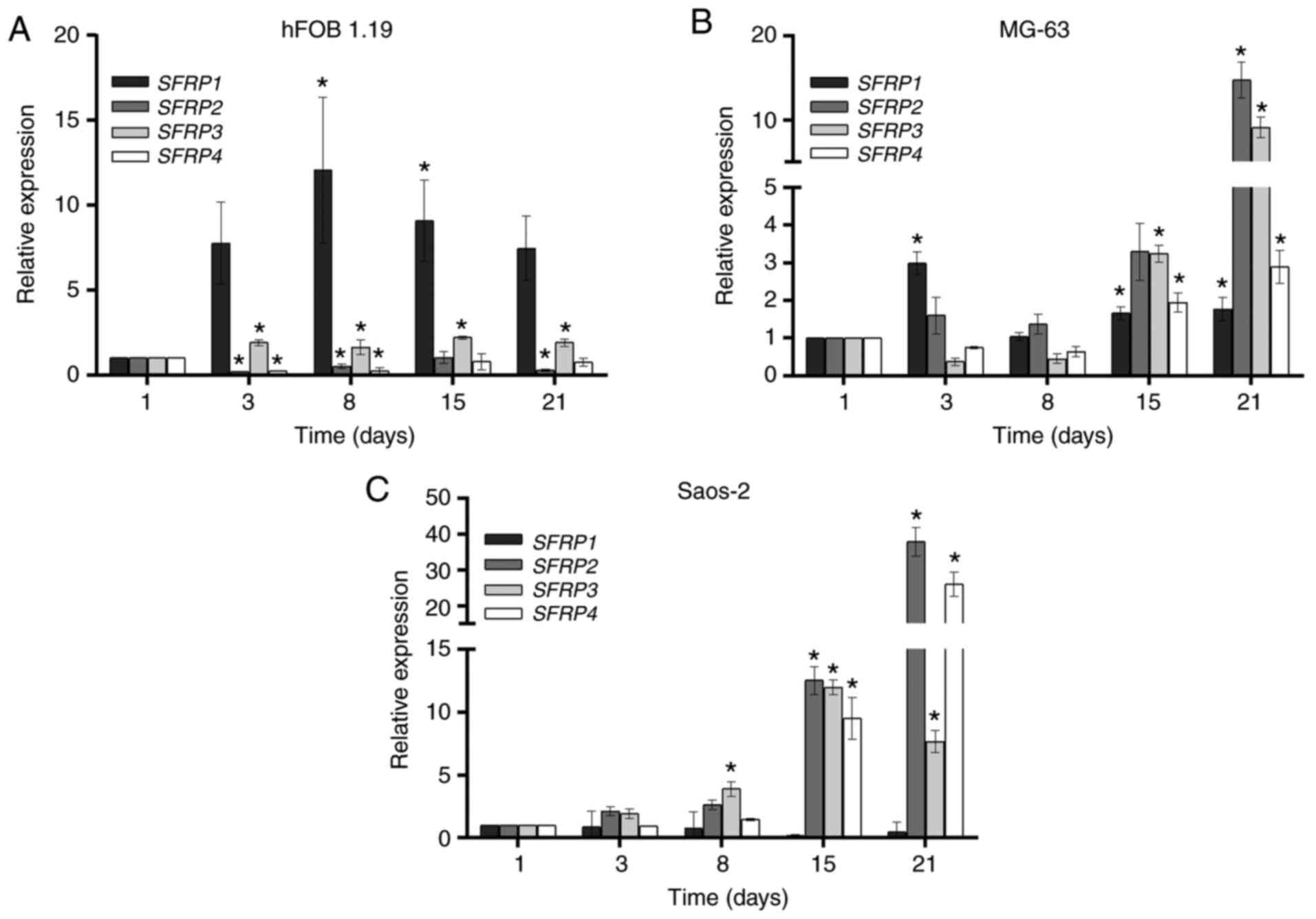
SFRPs genes
The expression levels of SFRP1, SFRP2,
SFRP3 and SFRP4 in the hFOB 1.19, MG-63 and Saos-2
cell lines, during 21 days of cell culture are shown in Fig. 2. The expression levels of
SFRP1 and SFRP3 were increased from day 3 until day
21 in the hFOB 1.19 cell line, compared with those on day 1. By
contrast, the expression levels of SFRP2 and SFRP4
genes decreased from day 3 until day 21, compared with those on day
1 (Fig. 2A).
In the MG-63 cell line, there was an increase in the
expression of SFRP1 from day 3 until day 21, while high
expression levels of SFRP2, SFRP3 and SFRP4
were detected after day 15 of cell culture, reaching maximum levels
at day 21 (Fig. 2B). The Saos-2
cell line had higher expression levels of SFRP2,
SFRP3 and SFRP4 compared with those in the hFOB 1.19
and MG-63 cell lines (Fig. 2C).
These high levels were detected from day 15 and the maximum levels
were detected at day 21. Notably, there were no significant
differences in the expression levels of SFRP1 during the 21
days of culture of Saos-2 cells (Fig.
2C).
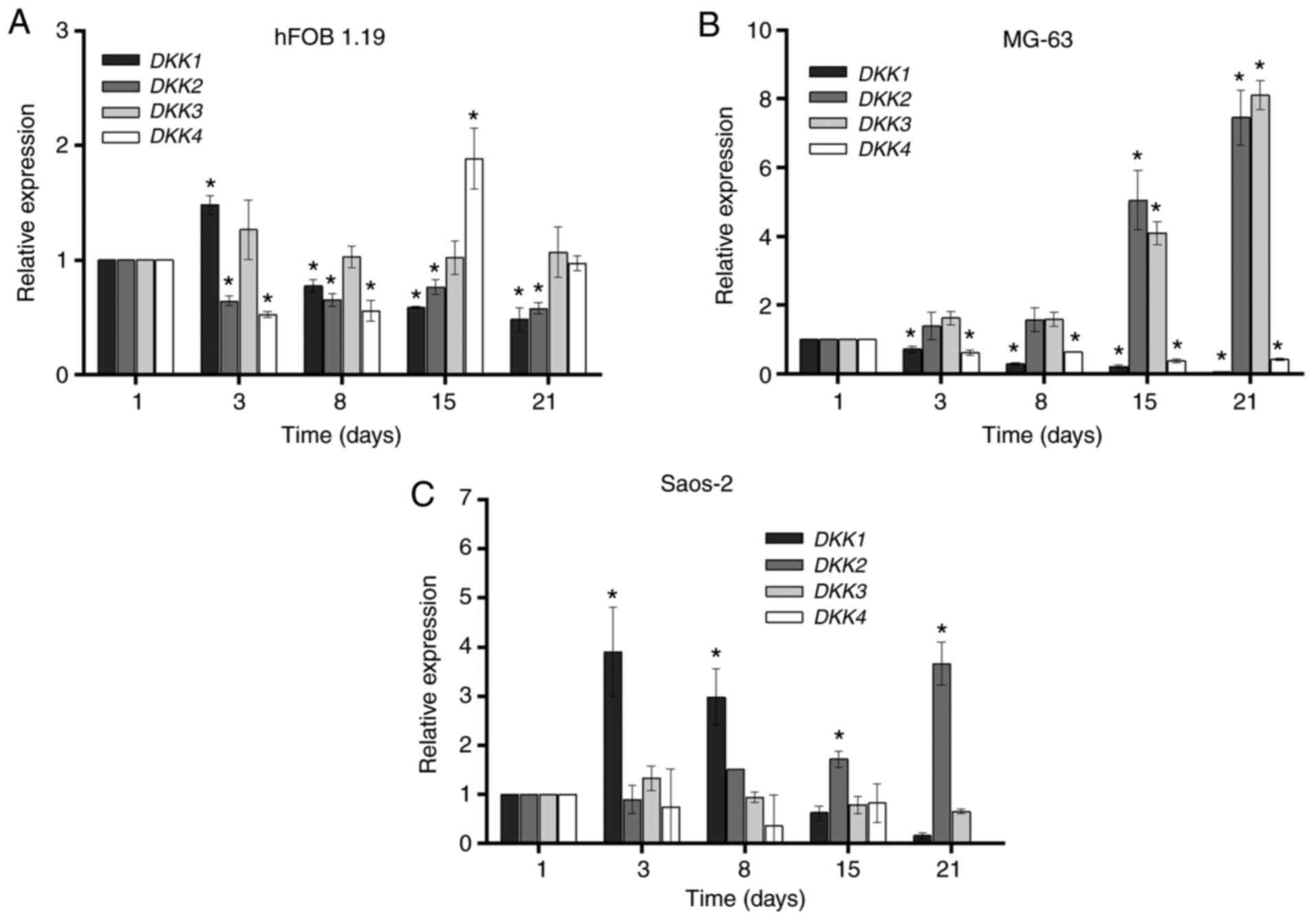
DKK genes
The expression levels of DKK1, DKK2,
DKK3 and DKK4 in the hFOB 1.19, MG-63 and Saos-2 cell
lines at days 1, 3, 8, 15 and 21 of cell culture are presented in
Fig. 3. The hFOB 1.19 cell line
demonstrated an increase in the expression of DKK1 after 3
days of culture, after which the expression decreased to levels
below the basal line to day 21. The expression of DKK4
decreased at day 8 of cell culture compared with that on day 1. At
day 15, there was a statistically significant increase in the
expression of DKK4. With respect to DKK2, there was a
decrease in its expression level during the 21 days of culture
compared with that on day 1. There were no significant changes in
DKK3 expression during the 21 days of cell culture (Fig. 3A).
In the MG-63 cell line, the expression levels of
DKK2 and DKK3 increased in a time-dependent manner,
reaching maximum levels at day 21 of cell culture. On the other
hand, the expression levels of DKK1 and DKK4 were
decreased below their basal levels during the 21 days of cell
culture, as compared with day 1 (Fig.
3B). By contrast, there was high expression of DKK1 on
day 3 in the Saos-2 cell line, which gradually decreased to levels
below its basal level at day 21 of culture. Furthermore, the
expression of DKK2 increased, reaching maximum levels on day
21, and there were no differences in the expression levels of
DKK3 and DKK4 during 21 days of cell culture. The
expression of DKK4 was not observed on day 21 (Fig. 3C).
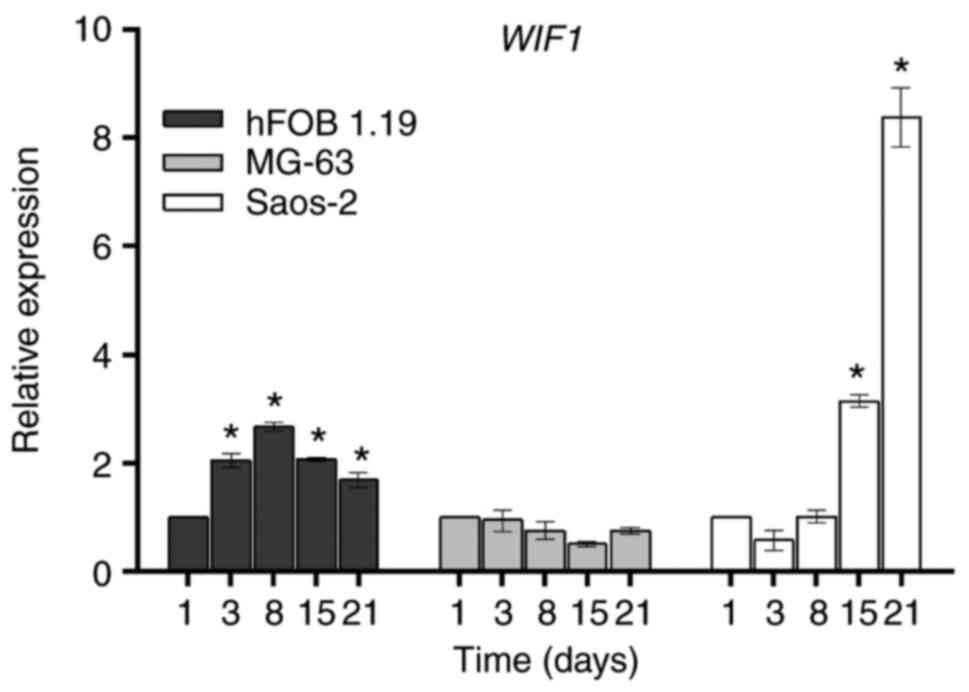
WIF1 gene. WIF1
is an important negative regulator factor of the
Wnt/β-catenin signaling pathway, and is structurally different from
the SFRP and DKK families (22).
WIF1 inhibits the activity of the Wnt signaling pathway by directly
binding to Wnt proteins (22).
Moreover, it is well-known that WIF1 can act as a tumor suppressor
and its downregulation is associated with the development of
various types of cancer (23). The
expression of WIF1 in the hFOB 1.19, MG-63 and Saos-2 cell
lines during the 21 days of culture is presented in Fig. 4. There was an increment on day 3,
which remained constant until day 21 in the hFOB 1.19 cell line,
while in Saos-2 cells, higher levels of WIF1 were observed
from day 15 and reached maximum levels on day 21 when compared with
day 1. By contrast, there were no changes in the expression of
WIF1 in the MG-63 cell line.
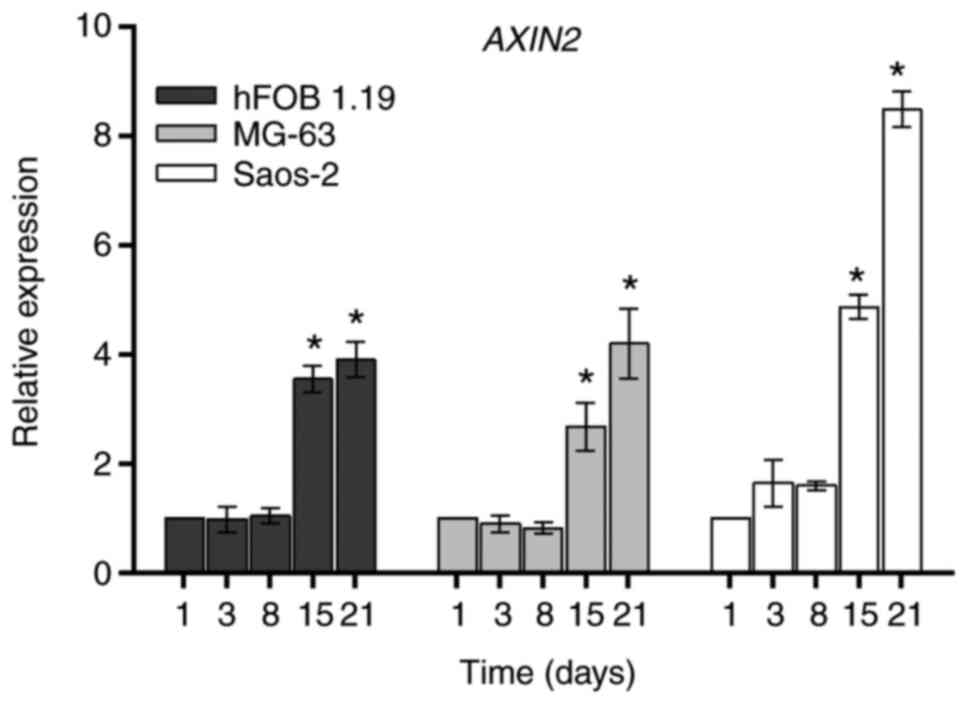
AXIN2 gene
AXIN2 is a negative intracellular regulator of the
Wnt signaling pathway, which forms a complex with APC and GSK3, and
results in the inhibition of the phosphorylation of β-catenin
(5). The expression of AXIN2
in the hFOB 1.19, MG-63 and Saos-2 cell lines during days 1, 3, 8,
15 and 21 of cell culture is illustrated in Fig. 5. The three cell lines had a similar
expression profile of AXIN2 (Table
SIII). The results identified that AXIN2 was constantly
expressed during the proliferative stage and there was a
significant increase on day 15, which reached maximum levels on day
21 of culture in all three osteoblastic cell lines, as compared
with the respective day 1. The relationship between AXIN2
and osteoblastic markers is presented in Tables SI and SII. A positive correlation was observed
between the AXIN2 expression profile and cell markers of
osteoblast proliferation (cell viability) and differentiation
stages in hFOB 1.19, MG-63 and Saos-2 cell lines. On the other
hand, a strong correlation between the mean of AXIN2 gene
expression and differentially expressed extracellular Wnt
antagonists during differentiation stage in the three cell lines
was observed (Tables
SIV-SVI).
Discussion
The results from the present study provide evidence
of the differential expression of certain bone-related Wnt
antagonists, during the proliferation (evaluated as cell viability)
or differentiation stages in human osteoblast-like cells. The roles
of these Wnt antagonists in the control of Wnt signaling have been
only partially described, which is due to the complexity of their
functions and the manner in which they have been investigated
(4).
In the present study, to evaluate the dynamics of
gene expression of the Wnt antagonists in the human osteoblasts,
the transition from proliferation to differentiation stage was
firstly determined in the hFOB 1.19, MG-63 and Saos-2 cells lines.
The differentiation stage was defined by high confluency in cell
culture, increased ALP activity and high expression levels of
RUNX2 and OSX (20,21).
The results demonstrated that arrest of cellular proliferation,
judged by cell confluency at day 15 of cell culture, was associated
with high levels of ALP activity and the expression of RUNX2
and OSX. The present results support the findings observed
in previous studies by Stein et al (24) and Owen et al (25), in which high mRNA and protein
expression levels of ALP, as well as no detectable
expression of RUNX2 and OSX, were identified before
day 12 of cell culture in rodent osteoblasts. Furthermore, in
another study, high levels of ALP activity and osteocalcin, a
differentiation marker, were observed at day 15 of cell culture in
a neonatal rat calvarial osteoblasts model (26). Based on the present results, it was
suggested that day 15 of cell culture was the beginning of the
differentiation stage in the three osteoblast-like cell lines. To
determine the expression levels of Wnt antagonists during the early
differentiation stage, the present study was limited to days 15-21
of cell culture, when ALP reached maximum levels of activity.
Previous studies have reported a significant decrease in enzyme
activity when cell cultures progress into the mineralization stage
(after day 25 of cell culture) (24,25).
However, the association between the Wnt-pathway antagonists and
the mineralization process was beyond the scope of the present
research.
To characterize the expression profiles of the Wnt
antagonists in the three osteoblast-like cell lines during the
proliferation and differentiation stages, the cells were cultured
for 21 days. Distinctive expression patterns were identified during
both the proliferation and differentiation stages for each cell
line. The differential expression patterns of Wnt antagonists
suggested there was a possible balance between the temporal and
spatial expression of Wnt-pathway antagonists during the
progression of proliferative stage towards the differentiation of
the osteoblasts (27,28). The present results demonstrated
there was an overlap between SFPR2, SFRP3,
SFRP4 and DKK2 gene expression levels in MG-63 and
Saos-2 cell lines. The analysis of their expression patterns
identified high levels on days 15 and 21 of cell culture,
suggesting that these antagonists were upregulated during the
differentiation stage in osteosarcoma cell lines. In addition, high
expression of DKK3 in MG-63, but not in Saos-2 cells, was
observed during the differentiation stage. On the other hand, in
the hFOB1.19 cells, high expression of DKK4 was found during
the differentiation stage (day 15). Several studies in murine
osteoblasts cells have reported that the expression levels of
SFRP2, SFRP4 (9,29) and
SFRP3, activated by the β-catenin-independent pathway
(30), can promote osteoblast
differentiation by decreasing cell proliferation and inducing ALP
activity. It has also been revealed that increased expression
levels of DKK2 (31) or
WIF1 (32) in murine
osteoblasts and Saos-2 cells, promote in vitro
mineralization. Moreover, DKK3, in Saos-2 and mesenchymal
cells (33,34) and DKK4 in MC3T3-E1 cells
(35), can increase cell
proliferation and decrease or inhibit osteogenic
differentiation.
WIF1 is a negative regulator, which acts upstream of
the Wnt signaling pathway, and can inhibit the activation of the
pathway by directly binding with the Wnt signaling proteins
(22). WIF1 has also been found to
act as a tumor suppressor protein (23). The current expression analysis of
WIF1 identified that the MG-63 and Saos-2 cell lines
exhibited low levels during the proliferative stage, and these were
constantly low during the differentiation stage in the MG-63 cells,
but not in Saos-2 cells. In the Saos-2 cell line, WIF1 was
upregulated in a time-dependent manner from days 15 to 21. In
normal hFOB1.19 cells, the expression of WIF1 was constant
in both the proliferation and differentiation stages, but its
levels were higher compared with those in MG-63 cells. These
findings suggested that the activation of the Wnt/β-catenin
signaling pathway may be associated with cell proliferation. A
recent study reported an association between the decreased mRNA and
protein expression levels of WIF1 and the increased levels
of β-catenin and cyclin D1 expression in tumor tissues, compared
with that in healthy tissues (36).
AXIN2 is a known intracellular negative regulator of
the Wnt/β-catenin signaling pathway, which acts by preventing
spontaneous signal transduction in the absence of a Wnt signal
(5). At the same time, AXIN2
expression is repressed by the activation of the Wnt/β-catenin
signaling pathway, which creates a negative feedback loop between
the two (6). Several studies have
revealed that high expression of AXIN2 is associated with
the inhibition of the Wnt/β-catenin signaling pathway (6,37). The
results from the present study demonstrated that high mRNA
expression levels of AXIN2 during differentiation stage was
associated with the overexpression of the Wnt antagonists. The
expression levels of SFRP2, SFRP3, SFRP4 and
DKK2 were upregulated in the Saos-2 and MG-63 cell lines,
DKK3 was upregulated in the MG-63 cell line and WIF1
was upregulated in the Saos-2 cell line, while DKK4 was
upregulated only in the hFOB 1.19 cell line. These results
indicated that extracellular and intracellular antagonists could
modulate the Wnt/β-catenin signaling pathway to decrease cell
proliferation and promote osteoblast differentiation (37). However, additional studies are
required to further assess this hypothesis.
The number of upregulated Wnt antagonists during
cell proliferation is limited (17). The present study identified that
DKK1 was the only gene that had a high expression in the
proliferation stage in the hFOB 1.19 and Saos-2 cell lines.
However, the underlying mechanism of this high expression is
currently unknown. A previous study revealed that overexpression of
DKK1 in the MG-63 and Saos-2 cell lines decreased the lag
time prior to rapid exponential growth during cell proliferation
(38).
The present results suggested that majority of the
Wnt antagonists were downregulated during proliferation and/or
differentiation stages; for instance, SFRP2, SFRP4,
DKK1 and DKK2 in the hFOB 1.19 osteoblasts,
DKK1 and DKK4 in the MG-63 cells and SFRP1 and
DKK1 in the Saos-2 cells. The mechanism underlying the
observed gene repression of the Wnt antagonist was not determined;
however, recent studies have proposed a role for the small
non-coding RNAs in the control of these genes (39,40).
For example, overexpression of microRNA(miR)-29 modulates the
intracellular mRNAs expression levels of DKK1 and
SFRP2, thus promoting the differentiation of human
osteoblasts (17). Furthermore, it
has been shown that the high expression of miR-940 activates the
Wnt/β-catenin signaling pathway by downregulating SFRP1 in
human osteosarcoma tissue (41).
Thus, the expression profile of these small non-coding RNAs in the
progression of proliferation and differentiation in osteoblasts and
osteosarcoma cell lines requires further research.
To the best of our knowledge, the current study
provides for the first time, the distinctive and characteristic
expression patterns of several Wnt antagonists during the
proliferation and differentiation stages of human osteoblast cell
lines (Fig. 6). However, the
association between the gene expression levels and the protein
levels, as well as their functional roles during both stages are
still require further investigation. Moreover, other technologies,
such as next-generation sequencing (seq), including RNA-seq and
small-RNAseq, could be used to analyze additional aspects of RNA
biology to identify the changes in the expression levels of the Wnt
antagonists.
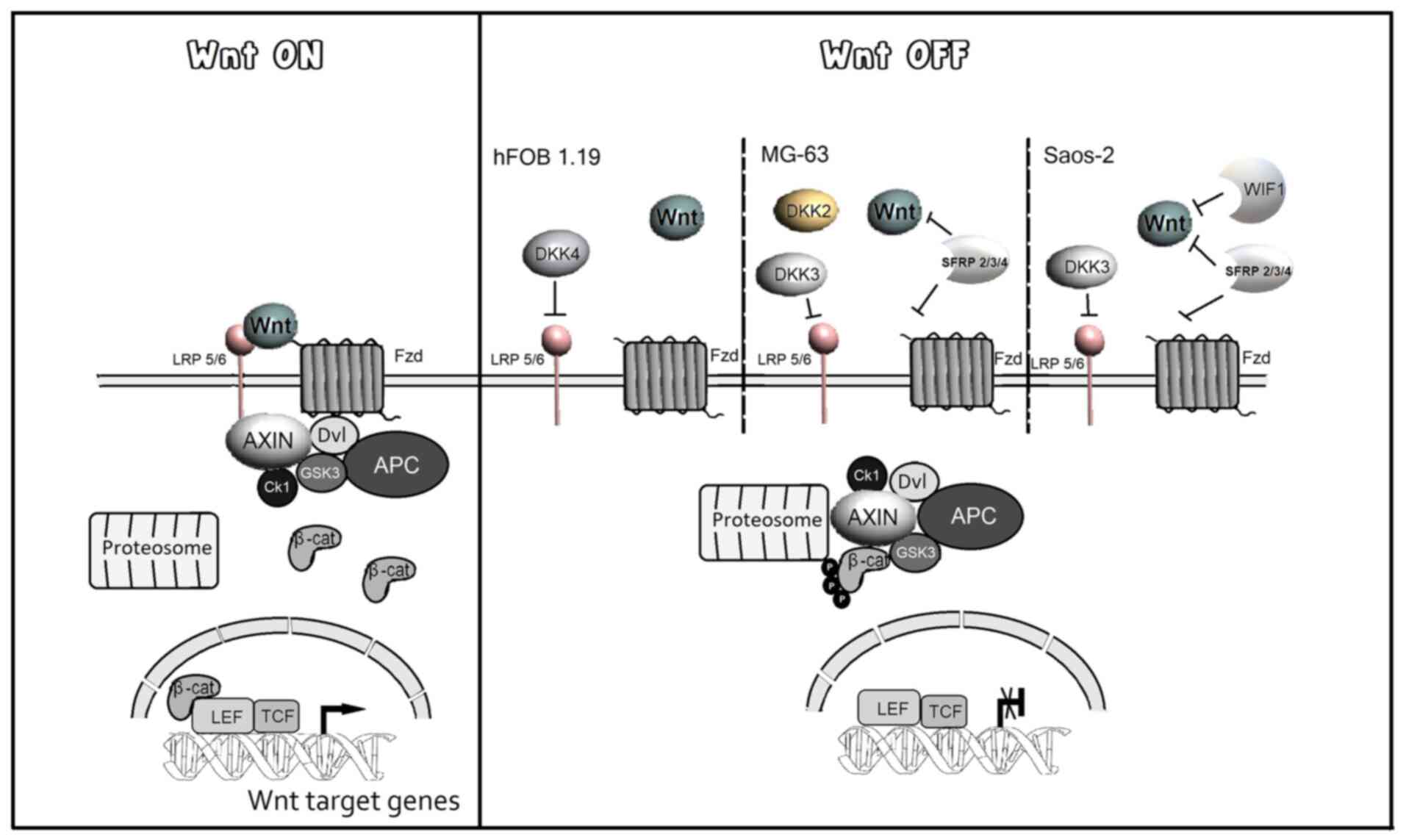 | Figure 6Schematic diagram of the effects of
extracellular antagonists in the Wnt/β-cat signaling pathway during
the proliferation and differentiation in human osteoblast.
Activation of the Wnt signaling induces the expression of genes
that promote cell proliferation. In the transition to
differentiation, the expression of extracellular antagonists
decreases Wnt signaling and promotes the expression of
differentiation markers. During the differentiation, the
upregulation of SFRP2, SFRP3, SFRP4 and
DKK2 is observed in the Saos-2 and MG-63 cell lines.
DKK3 and WIF1 are upregulated in MG-63 and Saos-2
respectively, while DKK4 is uniquely upregulated in the
hFOB1.19 cell line. AXIN2, anxin 2; WIF1, Wnt
inhibitory factor 1; DKK, Dickkopf; SFRP, secreted
Frizzled-related protein; Fzd, Frizzled; LRP, Low-density
lipoprotein receptor-related protein; GSK3, glycogen synthase
kinase 3; CK1, casein kinase 1; β-cat, β-catenin; Tcf, T-cell
specific transcription factor; Lef, lymphoid-enhancer binding
factor; Dvl, Dishevelled. |
In conclusion, the present results provide novel
insights into the expression levels of Wnt antagonists during
proliferation and differentiation stages in human osteoblast-like
cell lines. In addition, the results offer a basis to evaluate
novel potential targets for bone-related Wnt-signaling modulation
and provide an additional area of research into Wnt-signaling in
bone metabolism.
Supplementary Material
Table SI. Positive correlation between
the expression levels of osteoblast markers in hFOB 1.19
cells.
Table SII. Positive correlation
between the expression levels of osteoblast markers in the
osteosarcoma cell lines MG-63 and Saos-2.
Table SIII. Correlation between the
expression levels of osteoblast markers between hFOB 1.19, MG-63
and Saos-2 cell lines.
Table SIV. Correlation between the
mean of AXIN2 gene expression and differentially expressed
extracellular Wnt antagonists in hFOB 1.19 cells.
Table SV. Correlation between the mean
of AXIN2 gene expression and differentially expressed
extracellular Wnt antagonists in MG-63 cells.
Table SVI. Correlation between the
mean of AXIN2 gene expression and differentially expressed
extracellular Wnt antagonists in Saos-2 cells.
Acknowledgements
The authors would like to thank Mr. José Luis
Cruz-Colín (National Institute of Genomic Medicine, INMEGEN) for
his technical assistance with the culturing of cells.
Funding
The present study was partially supported by grants
from Consejo Nacional de Ciencia y Tecnología (grant no.
INFR-2016-01-270405) and the Instituto Nacional de Medicina
Genómica (grant no. 266-17/2016/I).
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
AYPT and RVC conceived and designed the study. JE,
EGRS, RFJO and NP carried out the experiments, acquisition of data,
analysis and interpretation of data. LMTE and MDJCL contributed to
the statistical analysis and interpretation of data. AYPT, JE,
EGRS, MDJCL and RVC drafted, reviewed and edited the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Authors' information
Miss Alma Y. Parra-Torres is a doctoral student
from Programa de Doctorado en Ciencias Biomédicas, Universidad
Nacional Autónoma de México (UNAM) and received fellowship 421295
from CONACYT.
References
|
1
|
Raggatt LJ and Partridge NC: Cellular and
molecular mechanisms of bone remodeling. J Biol Chem.
285:25103–25108. 2010.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Karner CM and Long F: Wnt signaling and
cellular metabolism in osteoblasts. Cell Mol Life Sci.
74:1649–1657. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kobayashi Y, Uehara S, Udagawa N and
Takahashi N: Regulation of bone metabolism by Wnt signals. J
Biochem. 159:387–392. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Boudin E, Fijalkowski I, Piters E and Van
Hul W: The role of extracellular modulators of canonical Wnt
signaling in bone metabolism and diseases. Semin Arthritis Rheum.
43:220–240. 2013.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Nusse R and Clevers H: Wnt/β-catenin
signaling, disease, and emerging therapeutic modalities. Cell.
169:985–999. 2017.
|
|
6
|
Jho E, Zhang T, Domon C, Joo CK, Freund JN
and Costantini F: Wnt/beta-catenin/Tcf signaling induces the
transcription of axin2, a negative regulator of the signaling
pathway. Mol Cell Biol. 22:1172–1183. 2002.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Bodine PVN, Zhao W, Kharode YP, Bex FJ,
Lambert AJ, Goad MB, Gaur T, Stein GS, Lian JB and Komm BS: The Wnt
antagonist secreted frizzled-related protein-1 is a negative
regulator of trabecular bone formation in adult mice. Mol
Endocrinol. 18:1222–1237. 2004.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Morvan F, Boulukos K, Clément-Lacroix P,
Roman SR, Suc-Royer I, Vayssière B, Ammann P, Martin P, Pinho S,
Pognonec P, et al: Deletion of a single allele of the Dkk1 gene
leads to an increase in bone formation and bone mass. J Bone Miner
Res. 21:934–945. 2006.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Cho SW, Her SJ, Sun HJ, Choi OK, Yang JY,
Kim SW, Kim SY and Shin CS: Differential effects of secreted
frizzled-related proteins (sFRPs) on osteoblastic differentiation
of mouse mesenchymal cells and apoptosis of osteoblasts. Biochem
Biophys Res Commun. 367:399–405. 2008.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Cho SW, Yang JY, Sun HJ, Jung JY, Her SJ,
Cho HY, Choi HJ, Kim SW, Kim SY and Shin CS: Wnt inhibitory factor
(WIF)-1 inhibits osteoblastic differentiation in mouse embryonic
mesenchymal cells. Bone. 44:1069–1077. 2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
van der Horst G, van der Werf SM,
Farih-Sips H, van Bezooijen RL, Löwik CWGM and Karperien M:
Downregulation of Wnt signaling by increased expression of
Dickkopf-1 and -2 is a prerequisite for late-stage osteoblast
differentiation of KS483 cells. J Bone Miner Res. 20:1867–1877.
2005.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Sims AM, Shephard N, Carter K, Doan T,
Dowling A, Duncan EL, Eisman J, Jones G, Nicholson G, Prince R, et
al: Genetic analyses in a sample of individuals with high or low
BMD shows association with multiple Wnt pathway genes. J Bone Miner
Res. 23:499–506. 2008.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Fujita M, Urano T, Shiraki M, Momoeda M,
Tsutsumi O, Hosoi T, Clinic K, Orimo H, Ouchi Y and Inoue S:
Association of a single nucleotide polymorphism in the secreted
frizzled-related protein 4 (sFRP4) gene with bone mineral density.
Geriatrics Gerontol Int. 4:175–180. 2004.
|
|
14
|
Maehata Y, Takamizawa S, Ozawa S, Kato Y,
Sato S, Kubota E and Hata RI: Both direct and collagen-mediated
signals are required for active vitamin D3-elicited differentiation
of human osteoblastic cells: Roles of osterix, an
osteoblast-related transcription factor. Matrix Biol. 25:47–58.
2006.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Coelho MJ and Fernandes MH: Human bone
cell cultures in biocompatibility testing. Part II: Effect of
ascorbic acid, beta-glycerophosphate and dexamethasone on
osteoblastic differentiation. Biomaterials. 21:1095–1102.
2000.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Harris SA, Enger RJ, Riggs LB and
Spelsberg TC: Development and characterization of a conditionally
immortalized human fetal osteoblastic cell line. J Bone Miner Res.
10:178–186. 1995.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Kapinas K, Kessler C, Ricks T, Gronowicz G
and Delany AM: miR-29 modulates wnt signaling in human osteoblasts
through a positive feedback loop. J Biol Chem. 285:25221–25231.
2010.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Waterborg JH and Matthews HR: The lowry
method for protein quantitation. Methods Mol Biol. 1:1–3.
1984.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Hoemann CD, El-Gabalawy H and McKee MD: In
vitro osteogenesis assays: Influence of the primary cell source on
alkaline phosphatase activity and mineralization. Pathol Biol
(Paris). 57:318–323. 2009.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Landim de Barros T, Brito VGB, do Amaral
CCF, Chaves-Neto AH, Campanelli AP and Oliveira SHP: Osteogenic
markers are reduced in bone-marrow mesenchymal cells and femoral
bone of young spontaneously hypertensive rats. Life Sci.
146:174–183. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kerekes K, Bányai L, Trexler M and Patthy
L: Structure, function and disease relevance of Wnt inhibitory
factor 1, a secreted protein controlling the Wnt and hedgehog
pathways. Growth Factors. 37:29–52. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Rubin EM, Guo Y, Tu K, Xie J, Zi X and
Hoang BH: Wnt inhibitory factor 1 decreases tumorigenesis and
metastasis in osteosarcoma. Mol Cancer Ther. 9:731–741.
2010.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Stein GS, Lian JB and Owen TA:
Relationship of cell growth to the regulation of tissue-specific
gene expression during osteoblast differentiation. FASEB J.
4:3111–3123. 1990.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Owen TA, Aronow M, Shalhoub V, Barone LM,
Wilming L, Tassinari MS, Kennedy MB, Pockwinse S, Lian JB and Stein
GS: Progressive development of the rat osteoblast phenotype in
vitro: Reciprocal relationships in expression of genes associated
with osteoblast proliferation and differentiation during formation
of the bone extracellular matrix. J Cell Physiol. 143:420–430.
1990.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Enríquez J, Lemus AE, Chimal-Monroy J,
Arzate H, García GA, Herrero B, Larrea F and Pérez-Palacios G: The
effects of synthetic 19-norprogestins on osteoblastic cell function
are mediated by their non-phenolic reduced metabolites. J
Endocrinol. 193:493–504. 2007.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Vaes BL, Dechering KJ, van Someren EP,
Hendriks JM, van de Ven CJ, Feijen A, Mummery CL, Reinders MJ,
Olijve W, van Zoelen EJ and Steegenga WT: Microarray analysis
reveals expression regulation of Wnt antagonists in differentiating
osteoblasts. Bone. 36:803–811. 2005.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Eijken M, Meijer IMJ, Westbroek I, Koedam
M, Chiba H, Uitterlinden AG, Pols HAP and van Leeuwen JPTM: Wnt
signaling acts and is regulated in a human osteoblast
differentiation dependent manner. J Cell Biochem. 104:568–579.
2008.PubMed/NCBI View Article : Google Scholar
|
|
29
|
von Marschall Z and Fisher LW: Secreted
frizzled-related protein-2 (sFRP2) augments canonical Wnt3a-induced
signaling. Biochem Biophys Res Commun. 400:299–304. 2010.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Chung YS, Baylink DJ, Srivastava AK, Amaar
Y, Tapia B, Kasukawa Y and Mohan S: Effects of secreted
frizzled-related protein 3 on osteoblasts in vitro. J Bone Miner
Res. 19:1395–1402. 2004.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Li X, Liu P, Liu W, Maye P, Zhang J, Zhang
Y, Hurley M, Guo C, Boskey A, Sun L, et al: Dkk2 has a role in
terminal osteoblast differentiation and mineralized matrix
formation. Nat Genet. 37:945–952. 2005.PubMed/NCBI View
Article : Google Scholar
|
|
32
|
Baker EK, Taylor S, Gupte A, Chalk AM,
Bhattacharya S, Green AC, Martin TJ, Strbenac D, Robinson MD,
Purton LE and Walkley CR: Wnt inhibitory factor 1 (WIF1) is a
marker of osteoblastic differentiation stage and is not silenced by
DNA methylation in osteosarcoma. Bone. 73:223–232. 2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Hoang BH, Kubo T, Healey JH, Yang R,
Nathan SS, Kolb EA, Mazza BA, Meyers PA and Gorlick R: Dickkopf 3
inhibits invasion and motility of Saos-2 osteosarcoma cells by
modulating the Wnt-beta-catenin pathway. Cancer Res. 64:2734–2739.
2004.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Aslan H, Ravid-Amir O, Clancy BM,
Rezvankhah S, Pittman D, Pelled G, Turgeman G, Zilberman Y, Gazit
Z, Hoffmann A, et al: Advanced molecular profiling in vivo detects
novel function of dickkopf-3 in the regulation of bone formation. J
Bone Miner Res. 21:1935–1945. 2006.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Hiramitsu S, Terauchi M and Kubota T: The
effects of dickkopf-4 on the proliferation, differentiation, and
apoptosis of osteoblasts. Endocrinology. 154:4618–4626.
2013.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Tang Q, Zhao H, Yang B, Li L, Shi Q, Jiang
C and Liu H: WIF1 gene inhibition and Wnt signal transduction
pathway activation in NSCLC tumorigenesis. Oncol Lett.
13:1183–1188. 2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Yan Y, Tang D, Chen M, Huang J, Xie R,
Jonason JH, Tan X, Hou W, Reynolds D, Hsu W, et al: Axin2 controls
bone remodeling through the beta-catenin-BMP signaling pathway in
adult mice. J Cell Sci. 122:3566–3578. 2009.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Gregory CA, Singh H, Perry AS and Prockop
DJ: The Wnt signaling inhibitor dickkopf-1 is required for reentry
into the cell cycle of human adult stem cells from bone marrow. J
Biol Chem. 278:28067–28078. 2003.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Nakasa T, Yoshizuka M, Andry Usman M,
Elbadry Mahmoud E and Ochi M: MicroRNAs and bone regeneration. Curr
Genomics. 16:441–452. 2015.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Wang J, Liu S, Li J, Zhao S and Yi Z:
Roles for miRNAs in osteogenic differentiation of bone marrow
mesenchymal stem cells. Stem Cell Res Ther. 10(197)2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Lin ZW, Zhang W, Jiang SD, Wei WB and Li
XF: Inhibition of microRNA-940 suppresses the migration and
invasion of human osteosarcoma cells through the secreted
frizzled-related protein 1-mediated Wnt/β-catenin signaling
pathway. J Cell Biochem: Oct 15, 2018 (Epub ahead of print). doi:
10.1002/jcb.27580.
|