Introduction
Hepatitis E virus (HEV) is an important causative
agent of severe hepatitis in humans (1). However, it is a poorly understood
pathogen, and has been indicated to cause high mortality during
pregnancy (2) and result in chronic
hepatitis in immunocompromised individuals, such as transplant
recipients and HIV-infected patients with low CD4 cell counts
(3,4).
HEV belongs to the Orthohepevirus genus of
the Hepeviridae family (5). The
7.2-kb HEV genome comprises three open reading frames (ORFs). ORF1
is a polyprotein required for HEV RNA replication, ORF2 is a capsid
protein and ORF3 is a small phosphoprotein involved in virion
morphogenesis and egress (6). ORF2
is involved in the formation of particle assembly, binding to host
cells and elicitation of neutralizing antibodies (7-9).
Its capsid protein comprises three domains: Domain S (amino acids
118-313) builds the viral shell, domain M (amino acids 314-453)
contributes to a surface plateau at the 3-fold axes of the virus
capsid and domain P (amino acids 454-606) forms a protruding spike
from the shell (10). It also
includes a cell attachment region and neutralizing antigenic region
(11).
In cultured cells and samples from infected
patients, HEV produces three forms of the ORF2 protein:
Intracellular, glycosylated and cleaved ORF2(12). The ORF2 protein sequence comprises
three potential positions for N-linked glycosylation: N137 and N310
in the S domain and N562 in the P domain (13). N562 is highly conserved and located
at the apical center of the protruding spike, which comprises a
cell attachment region and neutralizing antigenic region (14,15).
The location of N562 suggests that it may serve an important role
in viral morphogenesis and immune responses; however, several wild
strains contain an N562D mutation and it was reported that these
strains successfully infected PLC/PRF/5 cells (16). Therefore, the biological functions
of the other mutants at N562, and whether other types of amino acid
mutations will stimulate the body to produce more efficient
neutralizing antibodies, requires further elucidation. The effects
of specific amino acid substitutions at N562 on the dimerization,
antigenicity, immunogenicity and production of neutralizing
antibodies in the P domain remain poorly understood. The present
study performed site-directed mutagenesis on the truncated capsid
protein HEV pORF2 E1 (amino acids 439-617) expressed in Pichia
pastoris (P. pastoris) to investigate the effects
of specific amino acid substitutions at N562 in the dimerization,
antigenicity, immunogenicity and production of neutralizing
antibodies in the P domain.
Materials and methods
Anti-HEV IgG assay
Serum samples were collected from 18 hospital
patients with HEV (10 males and 8 females; age, 36-65 years),
admitted to Jinan Central Hospital Affiliated to Shandong First
Medical University and Shandong Academy of Medical Sciences (Jinan,
China) between January and October 2019. Patients with psychiatric
illness, malignancy, pregnancy, HIV infection, herpes zoster, bowel
disease and celiac disease were excluded. All patients signed
informed consent, and the study was approved by the Bioethics
Committee of Jinan Central Hospital Affiliated to Shandong First
Medical University and Shandong Academy of Medical Sciences. Serum
samples were tested for anti-HEV IgG antibodies according to the
manufacturer's protocol [HEV IgG antibody detection kit
(Chemiluminescence microparticle immunoassay); cat. no. 20190904;
Xiamen InnoDx. Biotech Co., Ltd.) using an automated
chemiluminescent microparticle immunoassay system (Xiamen InnoDx
Biotech Co., Ltd.). The result was considered positive when the
signal-to-cut-off value of the sample was >1.20 (according to
the manufacturer's recommendations). The anti-HEV IgG-positive
human sera were subsequently pooled and stored at -80˚C until
further use.
Homology modeling and in silico
protein simulation
The SWISS-MODEL server accessible via the ExPASy
website (http://swissmodel.expasy.org/) (17,18)
was used for homology modeling. This prediction server can use the
known 3D protein structure with amino acid sequence homologous to
the protein of interest and predict its 3D structure based on this
homology, and automatically score the 3D protein structures. The
best scoring models were used to simulate the pORF2 homodimer using
the SymmDock server, which is an online server for the prediction
of complexes with Cn symmetry by geometry based docking
(http://bioinfo3d.cs.tau.ac.il/SymmDock/), and the
symmetry order used for docking was 2(19).
Site-directed mutagenesis and
construction of recombinant plasmids
Recombinant E1 encompassing residues 439-617 of
genotype 4 HEV ORF2 was cloned into pET-28a (+) plasmid
(Invitrogen; Thermo Fisher Scientific, Inc.), which has been
previously constructed in the laboratory (20), and site-directed mutagenesis was
performed to generate three mutant variants at N562. Amino acids
with molecular weights similar to that of asparagine were selected
(leucine, cysteine and lysine), representing aliphatic,
sulfur-containing and alkaline amino acids, respectively. Regarding
site-directed mutagenesis, the different E1 mutants were generated
using the overlap PCR approach. The primers used for mutagenesis
are listed in Table I, and PCR was
performed using Expand High Fidelity PCR System (Roche) as follows:
Denaturation at 94˚C for 45 sec, annealing at 54˚C for 30 sec and
extension at 72˚C for 60 sec for a total of 35 cycles. The
amplicons were separated by 1% agarose gel electrophoresis and
visualized by ethidium bromide.
 | Table IPrimers used for mutagenesis. |
Table I
Primers used for mutagenesis.
| Construct | Sequence
(5'-3') |
|---|
| E1N562 | F: CCC GAA TTC GTT
ATC CAG GAC TAT GAT AAT C |
| | R: TCT TCT AGA TCA
AGG GTA ATC AAC AGT GTC C |
| E1N562L | F: ATC ACT AGC AGT
AGT TTG ATA ATT GTA TGG G |
| | R: CCA TAC AAT TAT
ttg ACT ACT GCT AGT GAT C |
| E1N562C | F: ATC ACT AGC AGT
AGT ATC ATA ATT GTA TGG G |
| | R: TAC CCA TAC AAT
TAT tgt ACT ACT GCT AGT G |
| E1N562K | F: ATC ACT AGC AGT
AGT TGG ATA ATT GTA TGG G |
| | R: TAC CCA TAC AAT
TAT aag ACT ACT GCT AGT G |
Expression of recombinant proteins in
the P. pastoris strain SMD1168
The E1 gene and three mutants carried EcoRI
and XbaI restriction sites at their 5' and 3' ends,
respectively. All genes were cloned into
EcoRI-XbaI-digested pPICZaA (Invitrogen; Thermo
Fisher Scientific, Inc.). P. pastoris SMD1168 cells
(Invitrogen; Thermo Fisher Scientific, Inc.) were cultured in 3 ml
Yeast Extract-Peptone-Dextrose (YPD) medium (1% yeast extract, 2%
peptone, 2% dextrose) at 30˚C and when the cells were in the log
phase (100 µl) they were mixed with 10 µg plasmid linearized by
SacI and were subsequently transferred to an electroporation
cuvette and pulsed for 6 msec at 1,000 V using BioRad Gene Pulser
(Gene Pulser Xcell; Bio-Rad Laboratories, Inc.). In parallel, a
control P. pastoris clone harboring the empty parent
vector pPICZaA was generated. The transformation mixture (200 µl)
was plated in increasing concentrations of Zeocin™ (Invitrogen;
Thermo Fisher Scientific, Inc.) to select putative multicopy
recombinants at 30˚C for 3 days.
A simple fed-batch protocol developed for high-level
production of recombinant proteins by P. pastoris was
used, as described previously (21). Briefly, recombinant clones were
grown in 4 ml YPD medium at 30˚C to an optical density=1 at 600 nm
(OD600). Subsequently, 0.5 ml of the cells were inoculated into 50
ml buffered complex glycerol medium [1% yeast extract, 2% peptone,
100 mM potassium phosphate pH 6.0, 1.34% yeast nitrogen base (YNB),
4x10-4% biotin, 1% glycerol] grown at 30˚C until culture
reached OD600=20. The cells were harvested by centrifugation at
1,500 x g for 5 min at room temperature, washed with sterile
double-distilled water, and resuspended in 10 ml buffered
methanol-complex medium (1% yeast extract, 2% peptone, 100 mM
potassium phosphate pH 6.0, 1.34% YNB, 4x10-4% biotin,
0.5% methanol). The expression of recombinant proteins was induced
for 72 h at 30˚C, with 100% methanol being added to a final
concentration of 0.5% every 12 h to maintain the promoter activity.
After induction for 72 h, the cells were harvested by
centrifugation at 13,800 x g for 20 min at room temperature. Both
the cells and supernatant were stored at -80˚C.
SDS-PAGE and western blotting
A total of 20 µl from 10 ml culture supernatant was
mixed with 4 µl 6x SDS-PAGE loading buffer, boiled (100˚C) for 5
min, analyzed on 15% SDS-PAGE and stained with Coomassie blue. For
non-reducing SDS-PAGE, the buffer did not contain β-mercaptoethanol
and the samples were analyzed without boiling. For western blot
analysis, the separated proteins were electrotransferred onto a
nitrocellulose membrane. The membrane was incubated for 1 h at room
temperature with blocking buffer (5% nonfat milk power in 0.05%
Tween-20-TBS]. The monoclonal antibody (mAb) 5G5, which was
produced in our laboratory, and anti-HEV IgG-positive human sera
were used as primary antibodies (1:200) incubated at 4˚C overnight.
HRP-conjugated goat anti-mouse (cat. no. 5220-0287) and anti-human
IgG (cat. no. 5220-0277; both from KPL) were used as secondary
antibodies (1:2,000) for 2 h at room temperature. Color development
was performed with 3,3'-diaminobenzidin tetrahydrochloride
(MilliporeSigma; Merck KGaA) as a substrate for HRP.
Tunicamycin assays
For the detection of protein glycosylation, the
P. pastoris SMD1168 cells were divided into two equal
aliquots during induction periods: One aliquot was supplemented
with tunicamycin, which was dissolved in 1% (v/v) DMSO at a final
concentration of 50 µg/ml according to Ito et al (22), whereas the other aliquot was
incubated without tunicamycin. The culture supernatants were
harvested after 3 days of induction and subjected to SDS-PAGE
analysis, which was the same protocol reported in the previous
paragraph.
Double-antibody sandwich ELISA
Double-antibody sandwich ELISA was performed to
compare the antigenicity of the four recombinant HEV capsid
proteins at the same concentration. Microtiter plates were coated
with mAb 4E9 [100 µl/well, 1:200 dilution in CBS buffer (pH 9.6;
0.05 M; 1.59 g Na2CO3 and 2.93 g
NaHCO3 up to 1,000 ml double-distilled water) at 37˚C
for 2 h. The proteins were serially diluted from 1:100 to 1:51,200
and incubated at 37˚C for 45 min. After incubation with HRP-5G5
(1:5,000) at 37˚C for 45 min, the reaction was developed using 100
µl tetramethylbenzidine (MilliporeSigma; Merck KGaA) at 37˚C for 10
min and stopped using 50 µl 2M H2SO4. Optical
density was measured at 450 nm. The empty vector pPICZaA produced
from SMD1168 was used as a negative control. The mAb 4E9 used in
the present study was a gift from Changchun Institute of Biological
Products Co., Ltd. The HRP-5G5 antibody was previously produced in
our laboratory.
Immunization schedule and anti-HEV
antibody detection
Each protein was emulsified with Freund's complete
adjuvant (MilliporeSigma; Merck KGaA) and was used to immunize a
group of six specific pathogen-free female BALB/c mice at 6 weeks
of age with 20-25 g weight (obtained from the Animal Resource
Centre of Yangzhou University, Yangzhou, China). During the animal
experiment, the mice were maintained at 20-26˚C, 40-70% relative
humidity with a 12/12-h light/dark cycle and had free access to
autoclaved food pellets and autoclaved water, The mice were
subcutaneously inoculated at two locations on the back with 20 µg
of the target protein, quantified using the Bradford method. After
3 and 5 weeks, the mice were administered an intraperitoneal
booster injection with 20 µg of the target protein without the
adjuvant. Blood samples (200 µl) were collected from the ocular
venous plexus of each mouse at 0, 3, 5, 8, 10, 12, 14, 16, 18, 20,
22, 24, 26, 28, 30, 32, 34, 36 and 38 weeks. The ocular venous
plexus was selected to collect blood from the mice, as this method
was indicated to cause only minimal injury to the animals and is
easy to perform. Prior to blood collection, the mice were
anesthetized by an intraperitoneal injection of 400 mg/kg chloral
hydrate. The immune serum samples obtained from each group of mice
were pooled. At the end of the animal experiment, the mice were
euthanized by CO2 inhalation (60%; 3 l/min) in a
10-liter volume euthanasia chamber (23). After the animals gradually lost
consciousness, CO2 concentration was increased to 100%
and maintained for >10 min to ensure the animals' death. At that
time, the animals did not breathe spontaneously for 2-3 min without
blinking reflex. The animals died entirely unconscious. All
procedures involving animals were approved and performed according
to the guidelines for animal experimentation of the Jinan Central
Hospital Affiliated to Shandong First Medical University and
Shandong Academy of Medical Sciences Ethics Service Committee
(Jinan, China). Anti-HEV IgG antibodies in mouse sera were detected
by indirect ELISA, as described previously (24). Briefly, microtiter plates were
coated with 100 µl/well of p166 mix [previously produced in our
laboratory; four p166 recombinant proteins (amino acids 452-617 of
HEV) were generated from W1 (genotype 1; JX857689), Mexico-14
(genotype 2; M74506), US-1 (genotype 3; AF060668) and China-9829
(genotype 4; AY789225) and mixed in equal concentrations] in 1M
urea in PBS at a concentration of 1 µg/ml (24). The plates were incubated at 37˚C for
2 h. A total of 100 µl (1x104 dilution) of immune sera
in 10% ethylene glycol were added to each well, and subsequently
incubated for 45 min at 37˚C, followed by incubation with an
HRP-conjugated goat anti-mouse IgG (cat. no. 5220-0287, 1:5,000;
KPL USA). The reaction was developed using 100 µl
3,3',5,5'-tetramethylbenzidine at 37˚C for 10 min (MilliporeSigma),
and then terminated using 50 µl 2M H2SO4. The
optical density was measured at 450 nm with a reference wavelength
of 630 nm.
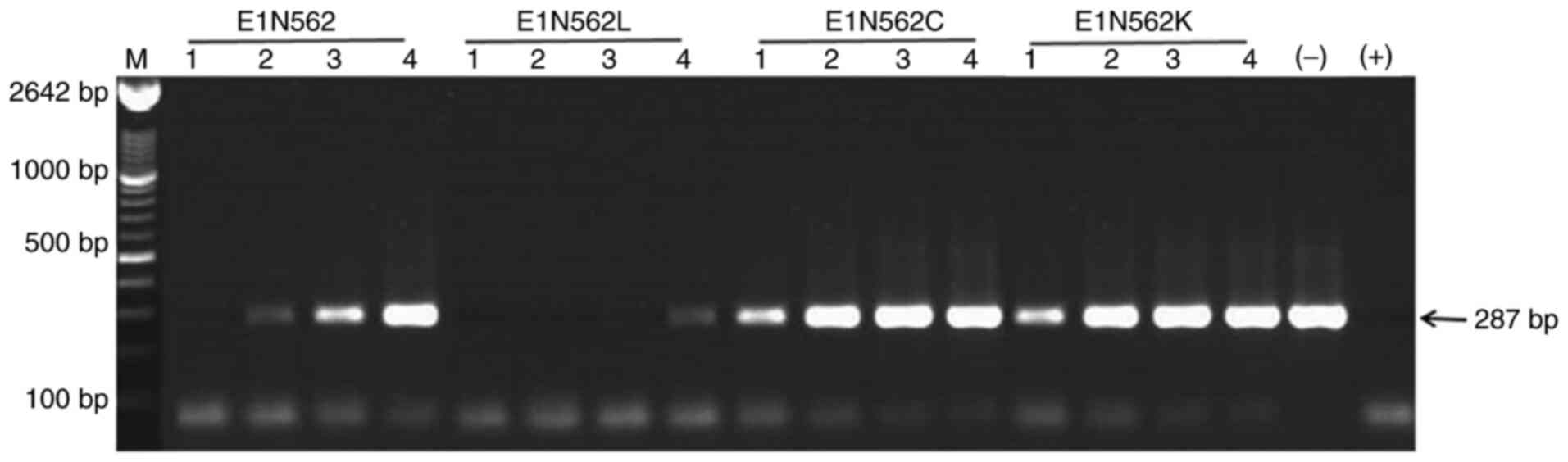
In vitro PCR-based neutralization
assay
The titers of the neutralizing antibodies were
tested by an in vitro PCR-based neutralization assay
(25,26). The inactivated (60˚C for 30 min)
serum samples were serially diluted from 1:10 to 1:80 by two-fold
steps using Hanks' Balanced Salt solution (HBSS). The NJ703 HEV
genotype 4 strain (GenBank accession no. AY789228; isolated from
the stool of an acute HEV-infected patient in our laboratory) was
diluted at 1:500 using HBSS and was mixed with equal volumes (100
µl) of the diluted serum. The mixture was incubated at 37˚C for 1 h
and inoculated onto an A549 cell monolayer (cat. no. CCL-185;
American Type Culture Collection) in DMEM medium supplemented with
10% FBS (Invitrogen; Thermo Fisher Scientific, Inc.). After
incubation for 5 days at 37˚C in 5% CO2, the cells were
washed three times with HBSS, followed by immediate RNA extraction
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). One-step reverse transcription (RT) PCR was
conducted, as described previously (21). OneStep RT-PCR kit (Qiagen GmbH) and
universal HEV PCR primers, forward, 5'-CCGACAGAATTGATTTCGTCGGC-3;
and reverse, 5'-TCGGCGGCGGTGAGAGAGAGCCA-3', were used as follows:
RT, 50˚C for 45 min and 95˚C for 15 min; PCR, denaturation at 94˚C
for 30 sec, annealing at 62˚C for 30 sec and extension at 72˚C for
45 sec for 55 cycles. The amplicons were separated by 1% agarose
gel electrophoresis and visualized using ethidium bromide.
Neutralization was defined by the absence of detectable HEV RNA in
the inoculated cell culture.
Statistical analysis
The double-antibody sandwich ELISA was repeated
twice and the results are presented as the mean ± SD. Differences
in the immunoreactivity with mAb 4E9 between wild-type (WT) E1 and
its three mutant groups were examined by one-way ANOVA followed by
Tukey's post hoc test. SPSS v20.0 (IBM Corp.) was used for
statistical analysis and the results were graphically illustrated
using GraphPad Prism v5.0 (GraphPad Software, Inc.). P<0.05 was
considered to indicate a statistically significant difference.
Results
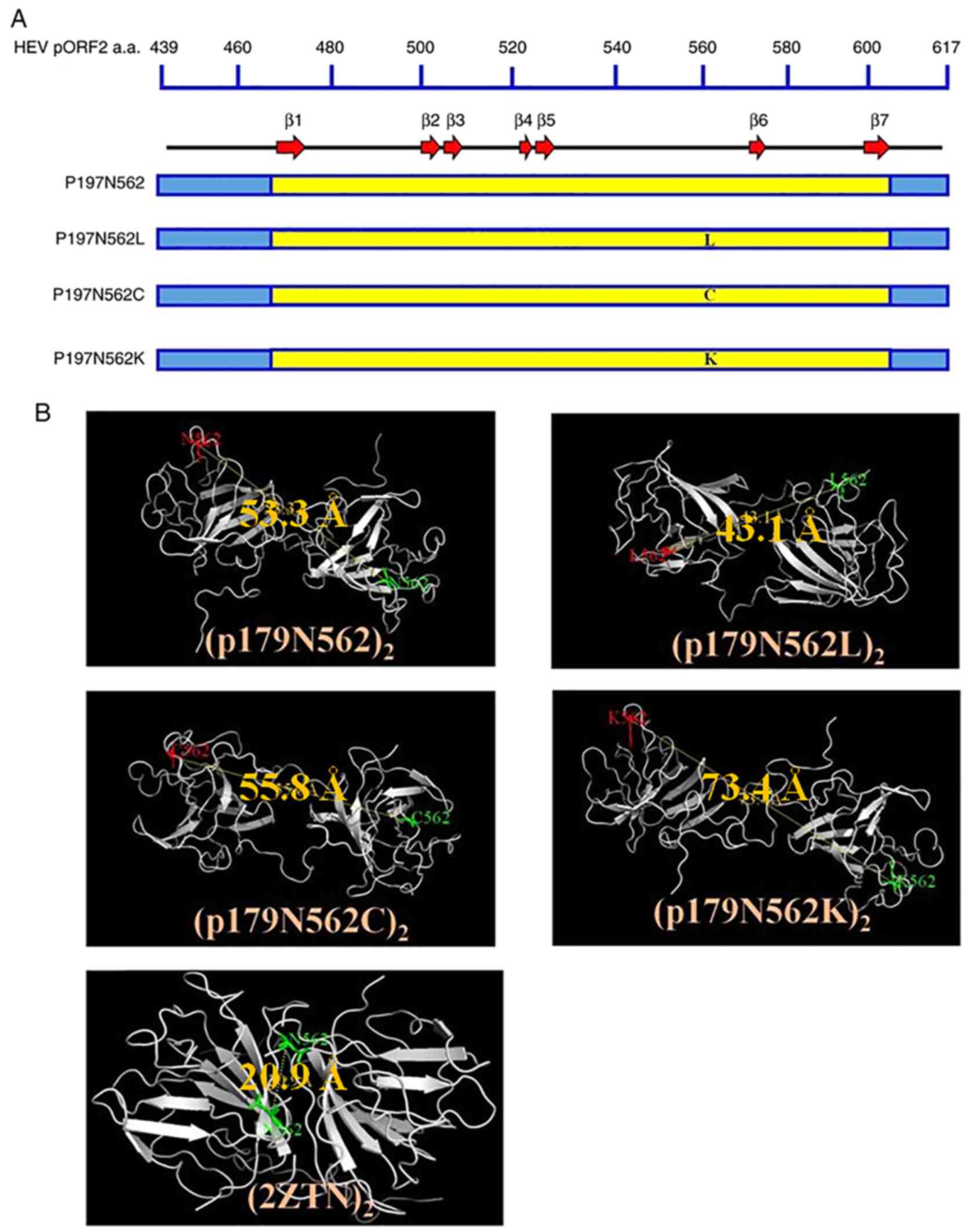
Predicted secondary structure models
and in silico simulations of homodimerization
The WT protein and its mutants exhibited similar
secondary structures, with the WT and the three mutated residues
all located on the coils (Fig. 1A).
The best ranked 3D models were selected among the available E1
protein structures using the SWISS-MODEL server. Subsequently,
dimerization was simulated using the SymmDock server. The best
simulations (the molecular simulations closest to the spatial
structure of the target protein) indicated that N562 was engaged in
the formation of homodimers, which is the case in the proposed
quaternary structure of the HEV-like particle (Protein Data Bank
ID: 2ZTN). To evaluate the engagement of N562 in the four predicted
E1 homodimers, (E1)2, (E1 N562L)2, (E1 N562C)2 and (E1 N562K)2, the
distance between two monomers at N562 was measured and compared
with those in the 2ZTN model (Fig.
1B). The distances between N562 and N562 (the same position in
the second monomer) was 20.9 Å in the 2ZTN model. This distance
allows N562 to engage in hydrogen bonding with the neighboring
amino acids in the opposite monomer and participate in the assembly
of the two monomers. Accordingly, concerning (E1 N562L)2, the
distance between L562 and L562' was 43.1 Å, which was closer to the
distance measured in the 2ZTN model compared with that in the WT,
E1 N562C and E1 N562K, which were predicted to exhibit distances of
53.3, 55.8 and 73.4 Å, respectively. This indicated that the amino
acid at position 562 was better positioned to engage in
homodimerization in the N562L model compared with the WT, N562C and
N562K models.
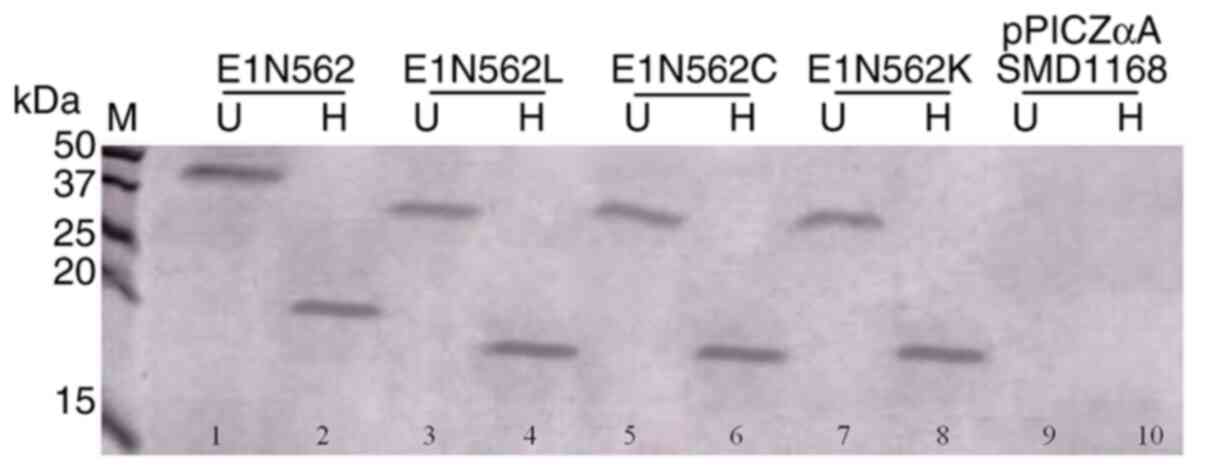
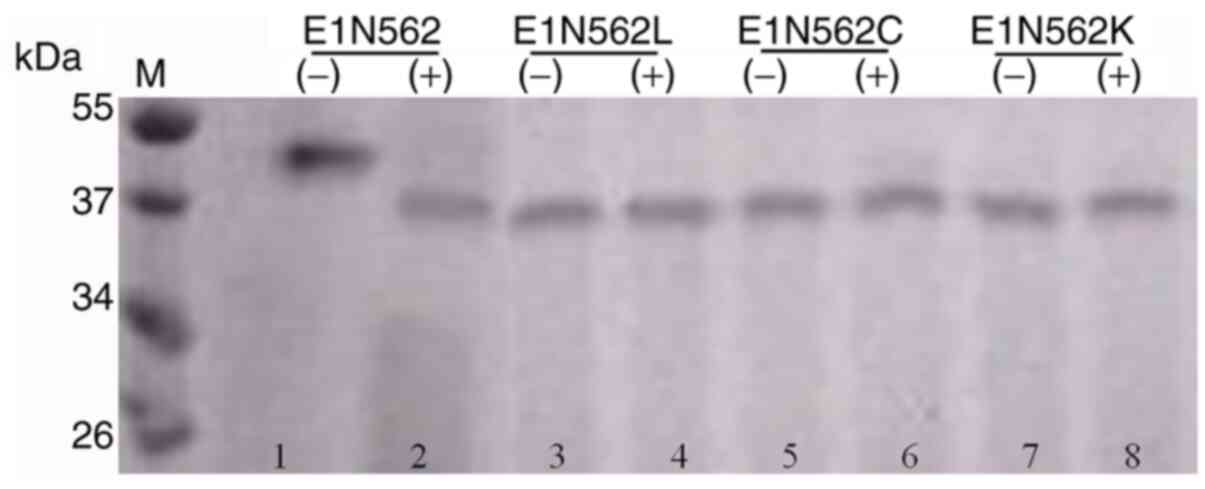
Protein dimerization
Non-reducing SDS-PAGE was performed to verify the
dimerization of E1 N562L, E1 N562C and E1 N562K mutants compared
with that of the WT protein. E1 N562L, E1 N562C and E1 N562K were
assembled into 39-kDa homodimers (Fig.
2; lanes 3, 5 and 7), whereas the WT protein was assembled into
a 44-kDa homodimer (Fig. 2; lane
1). Analysis of the denatured proteins indicated that the E1 N562L,
E1 N562C and E1 N562K mutant proteins were 19-kDa in size (Fig. 2; lanes 4, 6 and 8), whereas WT E1
migrated as a 21-kDa protein on the gel (Fig. 2; lane 2). Proteins from
pPICZaA/SMD1168 were used as negative control.
WT protein is a glycoprotein, but its
mutants are not glycosylated
The observation of a higher molecular weight of the
WT protein suggested that it may be glycosylated, whereas the
mutant proteins were either non-glycosylated or a relatively
smaller fraction was glycosylated. Tunicamycin, which is an
inhibitor of bacterial and eukaryote N-acetylglucosamine
transferases, is known to prevent the glycosylation of newly
synthesized glycoproteins (27).
Therefore, tunicamycin assays were performed to evaluate the
glycosylation status of the WT protein and its mutants. In the
presence of tunicamycin, the glycosylation of the WT protein was
inhibited, leading to a shift from the higher molecular weight to
the lower molecular eight form (Fig.
3; lanes 1 and 2). The WT protein was validated to be a
glycoprotein, as it was susceptible to tunicamycin treatment.
However, the electrophoretic mobility of the three mutants was
unaffected by tunicamycin treatment, suggesting that the mutants
were not glycosylated (Fig. 3;
lanes 3 to 8).
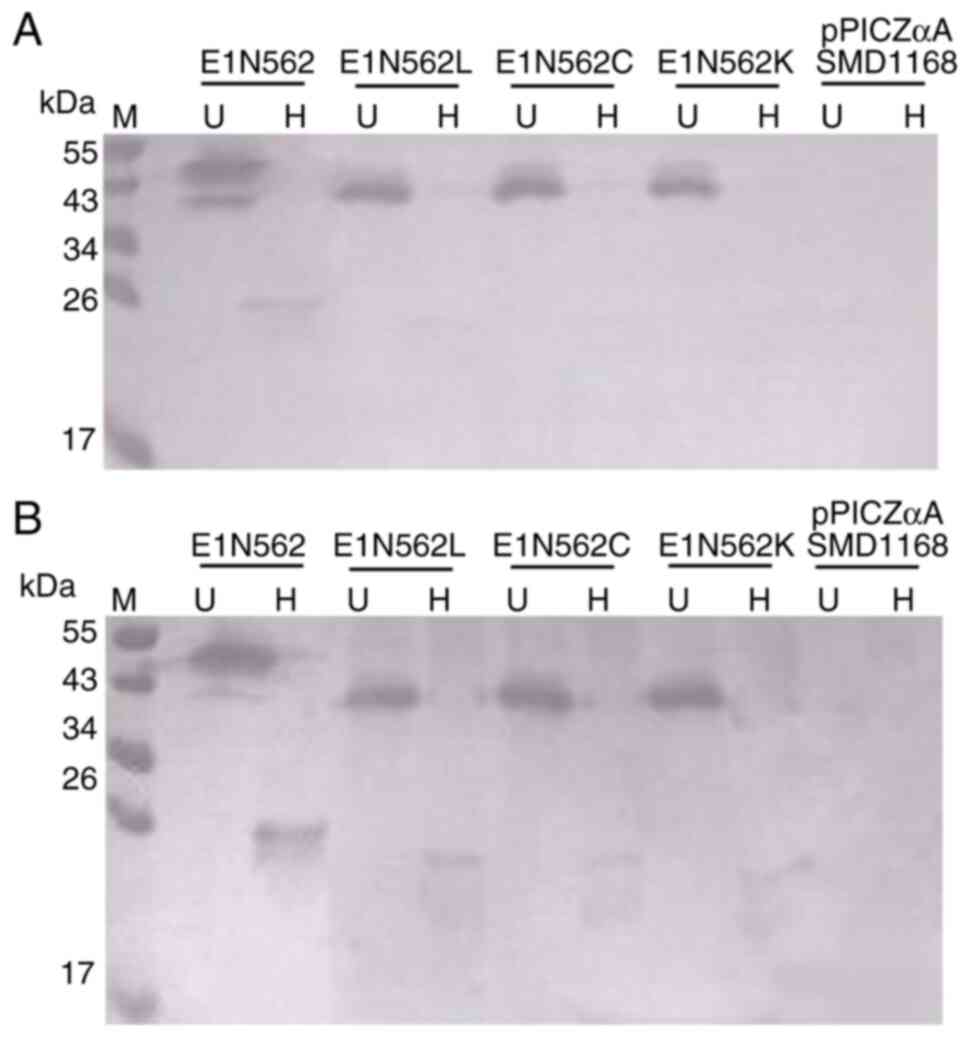
Immunoreactivity of the WT protein and
its three mutants
Western blot analysis demonstrated that the WT
protein and N562L, N562C and N562K immunoreacted with the
HEV-neutralizing mAb 5G5 as homodimers but were not reactive as
monomers (Fig. 4A). Western blot
analysis was also performed using anti-HEV IgG-positive human sera,
and the results were indicated to be similar to those obtained with
mAb 5G5 (Fig. 4B). Proteins from
pPICZaA/SMD1168 were used as negative control.
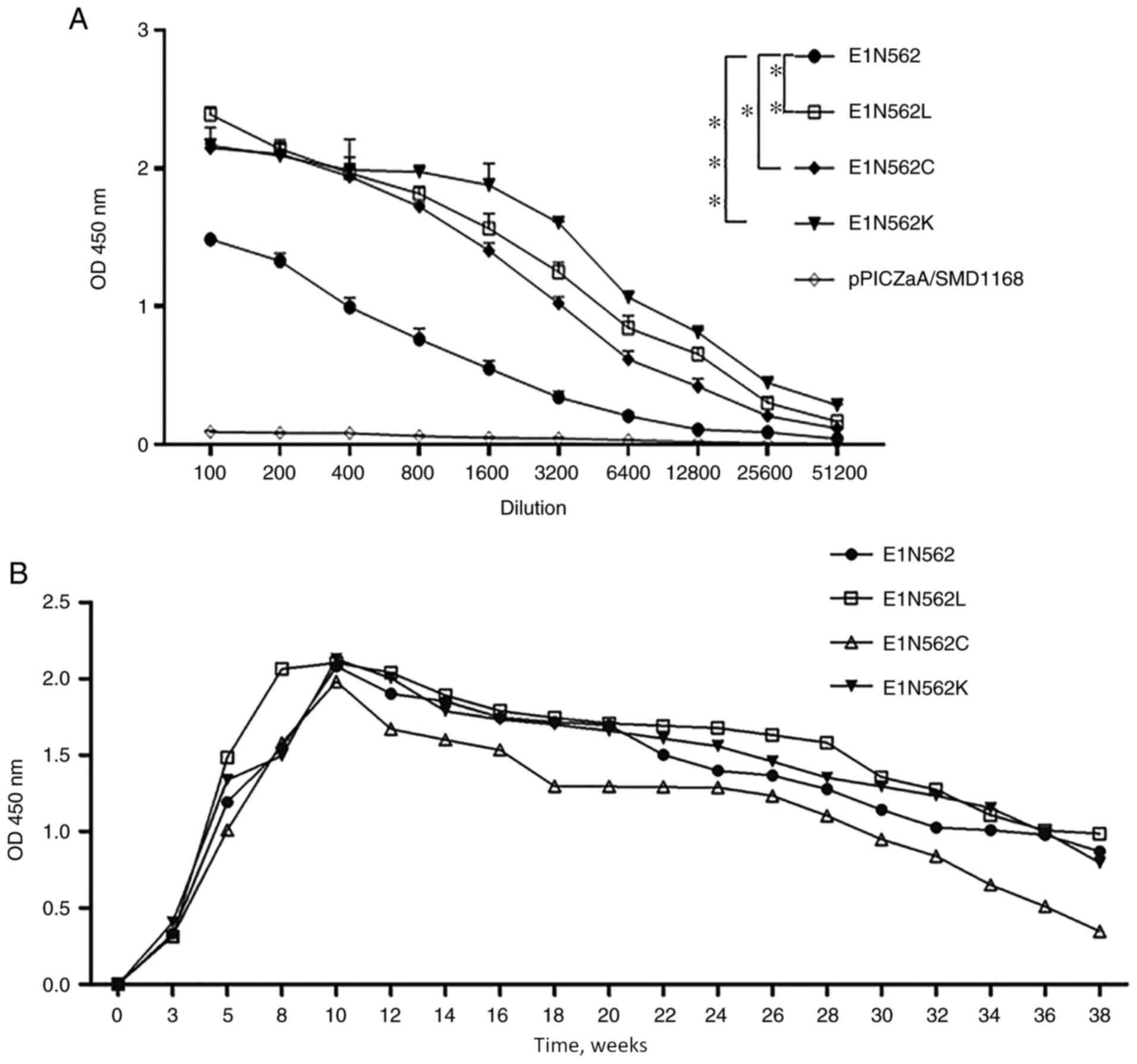
Antigenicity
The antigenicity of the expressed proteins was
assessed using double-antibody sandwich ELISAs. The
immunoreactivity levels were indicated to be decreased as the
dilution increased. The immunoreactivity of WT E1 was significantly
lower compared with that of the three mutant variants (Fig. 5A; P<0.05). Moreover,
pPICZaA/SMD1168 exhibited no reactivity to mAb 4E9. These results
indicated that a sugar chain at N562 may mask the partially
neutralizing epitopes, thereby reducing the reactivity with the
neutralizing mAb.
Immunoreactivity
All mice vaccinated with the recombinant proteins
presented HEV-specific antibodies in sera at 3 weeks post
inoculation (wpi). The E1-, E1 N562L-, E1 N562C- and E1
N562K-vaccinated mice exhibited peak levels of antibody titers at
10 wpi. Overall, the kinetics of antibody responses were highly
similar among the four groups, indicating that the amino acid
substitutions and glycosylation status resulted in no significant
differences in their kinetics. All immunized mice retained the
1x104 dilution of immune sera until 38 wpi (Fig. 5B).
Neutralization activity of mouse
sera
The ability of immune sera to neutralize HEV was
determined by an in vitro PCR-based neutralization assay.
Four pools of the E1, E1 N562L, E1 N562C and E1 N562K serum samples
were prepared from 8 wpi. Pre-inoculation serum specimens were used
as negative controls. The endpoint neutralization titers of the
antibodies against E1 and E1 N562L were 1:20 and 1:80, respectively
(Fig. 6). No neutralization was
observed using E1 N562C, E1 N562K and pre-inoculation sera diluted
1:10.
Discussion
The HEV capsid protein encoded by ORF2 is
responsible for the formation of particle assembly, binding to host
cells and elicitation of neutralizing antibodies (28,29).
Considerable effort has been made for the expression of recombinant
ORF2 protein to create a subunit vaccine (30-32).
In the present study, a secretory truncated version of the ORF2
protein was successfully expressed using P. pastoris
as a host system. Gurramkonda et al (33) previously reported a simple two-phase
strategy. In the first phase, they used a glycerol batch phase to
generate sufficient biomass. In the second phase, they used
methanol induction at a constant methanol concentration to achieve
high-level production of the recombinant protein. In the present
study, this two-phase strategy was successfully employed for the
secretion of E1. The addition of methanol once every 12 h is
recommended to prevent methanol depletion, as performed in the
present study.
The potential glycosylation motif 562NTT
has been indicated to be highly conserved, and only six out of 261
sequences (GenBank accession nos. AB189070, AB425831, AB593690,
AB698071, AB740232 and JQ026407) (34-37)
have been revealed to comprise D562, which is incompatible with the
N-linked glycosylation at this position. N562 is located in the
central region at the top of the P dimer and is exposed on the
surface of HEV (13). Owing to its
location in functionally and structurally important regions of the
HEV capsid protein, this position possibly serves a crucial role in
viral immune responses and morphogenesis (38). In the present study, a set of mutant
proteins containing L, C and K in place of N at position 562 was
constructed and tested for homodimerization, glycosylation,
antigenicity, immunogenicity and neutralization activity.
In the current study, E1 was indicated to form a
44-kDa homodimer. This molecular weight is slightly higher compared
with the predicted molecular weight of 40 kDa based on the
179-amino acid sequence. The slightly higher molecular weight
resulted from a facultative N-linked glycosylation site (N-X-S/T)
at N562 of the P domain. Tunicamycin is an inhibitor of bacterial
and eukaryotic N-acetylglucosamine transferases, preventing the
formation of N-acetylglucosamine lipid intermediates and the
glycosylation of newly synthesized glycoproteins (27). Tunicamycin has been indicated to
block the formation of N-glycosidic linkages by inhibiting the
transfer of N-acetylglucosamine-1-phosphate to dolichol
monophosphate (27). Tunicamycin
assay revealed that the 44-kDa protein homodimer was glycosylated.
The N562L, N562C and N562K mutations completely abrogated the
glycosylation, thereby supporting the importance of the asparagine
residue for glycosylation at this position, as reported previously
(39).
Li et al (38) reported that the dimerization of the
capsid E2s domain (ORF2; amino acids 455-602) was a prerequisite
for the virus-host interaction and for the binding of certain
neutralizing antibodies to HEV. The findings of the present study
indicated that E1 WT, E1 N562L, E1 N562C and E1 N562K formed a
homodimer, suggesting that replacing N562 with leucine, cysteine or
lysine exhibited no effect on dimerization. It may be possible that
the P. pastoris expression-secretion system used in
the current study was conducive to the formation of homodimers.
E1, E1 N562L, E1 N562C and E1 N562K were revealed to
interact with mAb 5G5 and anti-HEV IgG-positive human sera as
homodimers, whereas their denaturation abrogated this
immunoreactivity.
Furthermore, determination of immune response rates
and antibody neutralization capacities demonstrated that all four
recombinant HEV capsid proteins elicited similar antibody responses
in immunized mice, but not all antibodies were neutralized in the
in vitro PCR-based neutralization assay. The observation
that E1 N562L elicited the strongest neutralizing antibody response
among the tested proteins suggested that specific amino acid
substitutions at position 562 resulted in differential effects on
the immunological properties and neutralization of these proteins.
However, additional experiments are required to support these
results. A comparison experiment between E1 N562L and E1 N562
should be performed by measuring neutralizing antibody levels at
different doses of E1 N562L and E1 N562, such as 5, 10 and 20 µg,
to further support these conclusions.
In conclusion, analyses of the four recombinant HEV
capsid proteins comprising different mutations at position 562
revealed differential effects of mutations at this position on the
neutralizing capacity of the examined antibody. This finding
contributed to a greater understanding of the antigenic
characteristics of the glycosylated and non-glycosylated form of
capsid protein ORF2 and may be helpful in the development of novel
vaccines and diagnostic methods.
Acknowledgements
The authors would like to thank Dr Duanrui Liu
(Shandong University) for providing human clinical samples
collected from patients and Dr Kangle Niu (Shandong University) for
their valuable technical assistance and suggestions.
Funding
The present study was funded by the Academic
Promotion Program of Shandong First Medical University (grant no.
2019QL024), Natural Science Foundation of Shandong Province (grant
no. ZR2017BC002), Jinan Clinical Medicine Science and Technology
Innovation Plan (grant no. 201704117) and Jinan Postdoctoral
Innovation Project Second-Class Funding (grant no. 197249).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
MX performed site-directed mutagenesis and was a
major contributor in writing the manuscript. LS detected and
analyzed the patient with HEV for anti-HEV IgG antibody in the
patient's serum, performed expression of recombinant proteins in
the P. pastoris strain SMD1168. YW performed SDS-PAGE and
western blotting. SG performed the immunization schedule and
anti-HEV antibody detection. WY performed for the purification of
proteins. ML designed the research, performed in vitro
PCR-based neutralization assay and statistical analysis. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All patients signed an informed consent, and the
study was approved by the Bioethics Committee of Jinan Central
Hospital Affiliated to Shandong First Medical University and
Shandong Academy of Medical Sciences (Jinan, China). All procedures
involving animals were approved and performed according to the
guidelines for animal experimentation of the Jinan Central Hospital
Affiliated to Shandong First Medical University and Shandong
Academy of Medical Sciences Ethics Service Committee (Jinan,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yamada H, Takahashi K, Lim O, Svay S,
Chuon C, Hok S, Do SH, Fujimoto M, Akita T, Goto N, et al:
Hepatitis E Virus in Cambodia: Prevalence among the general
population and complete genome sequence of genotype 4. PLoS One.
10(e0136903)2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Pérez-Gracia MT, Suay-García B and
Mateos-Lindemann ML: Hepatitis E and pregnancy: Current state. Rev
Med Virol. 27(e1929)2017.PubMed/NCBI View
Article : Google Scholar
|
|
3
|
Abravanel F, Lhomme S, Fougère M, Saune K,
Alvarez M, Péron JM, Delobel P and Izopet J: HEV infection in
French HIV-infected patients. J Infect. 74:310–313. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Debes JD, Pas SD, Groothuismink ZMA, van
der Ende ME, de Man RA and Boonstra A: A mutation in the
progesterone receptor predisposes to HEV infection in HIV-positive
patients. Liver Int. 38:792–796. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Kasem A, Izopet J, Pavio N, Aggarwal R,
Labrique A, Wedemeyer H and Dalton HR: Epidemiology of hepatitis E
virus infection. Epidemiol Mikrobiol Imunol. 68:176–182.
2019.PubMed/NCBI
|
|
6
|
Tam AW, Smith MM, Guerra ME, Huang CC,
Bradley DW, Fry KE and Reyes GR: Hepatitis E virus (HEV): Molecular
cloning and sequencing of the full-length viral genome. Virology.
185:120–131. 1991.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Li S, Zhang J and Xia N: Lessons from
hepatitis E vaccine design. Curr Opin Virol. 11:130–136.
2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Chandra V, Kalia M, Hajela K and Jameel S:
The ORF3 protein of hepatitis E virus delays degradation of
activated groWTh factor receptors by interacting with CIN85 and
blocking formation of the Cbl-CIN85 complex. J Virol. 84:3857–3867.
2010.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Takahashi M, Yamada K, Hoshino Y,
Takahashi H, Ichiyama K, Tanaka T and Okamoto H: Monoclonal
antibodies raised against the ORF3 protein of hepatitis E virus
(HEV) can capture HEV particles in culture supernatant and serum
but not those in feces. Arch Virol. 153:1703–1773. 2008.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Guu TS, Liu Z, Ye Q, Mata DA, Li K, Yin C,
Zhang J and Tao YJ: Structure of the hepatitis E virus-like
particle suggests mechanisms for virus assembly and receptor
binding. Proc Natl Acad Sci USA. 106:12992–12997. 2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Yamashita T, Mori Y, Miyazaki N, Cheng RH,
Yoshimura M, Unno H, Shima R, Moriishi K, Tsukihara T, Li TC, et
al: Biological and immunological characteristics of hepatitis E
virus-like particles based on the crystal structure. Proc Natl Acad
Sci USA. 106:12986–12991. 2009.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Yin X, Ying D, Lhomme S, Tang Z, Walker
CM, Xia N, Zheng Z and Feng Z: Origin, antigenicity, and function
of a secreted form of ORF2 in hepatitis E virus infection. Proc
Natl Acad Sci USA. 115:4773–4778. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zafrullah M, Ozdener MH, Kumar R, Panda SK
and Jameel S: Mutational analysis of glycosylation, membrane
translocation, and cell surface expression of the hepatitis E virus
ORF2 protein. J Virol. 73:4074–4082. 1999.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Mori Y and Matsuura Y: Structure of
hepatitis E viral particle. Virus Res. 161:59–64. 2011.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Graff J, Zhou YH, Torian U, Nguyen H, St
Claire M, Yu C, Purcell RH and Emerson SU: Mutations within
potential glycosylation sites in the capsid protein of hepatitis E
virus prevent the formation of infectious virus particles. J Virol.
82:1185–1194. 2008.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Shiota T, Li TC, Yoshizaki S, Kato T,
Wakita T and Ishii K: The hepatitis E virus capsid C-terminal
region is essential for the viral life cycle: Implication for viral
genome encapsidation and particle stabilization. J Virol.
87:6031–6036. 2013.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Biasini M, Bienert S, Waterhouse A, Arnold
K, Studer G, Schmidt T, Kiefer F, Gallo Cassarino T, Bertoni M,
Bordoli L and Schwede T: SWISS-MODEL: Modelling protein tertiary
and quaternary structure using evolutionary information. Nucleic
Acids Res. 42:W252–W258. 2014.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Arnold K, Bordoli L, Kopp J and Schwede T:
The SWISS-MODEL workspace: A web-based environment for protein
structure homology modelling. Bioinformatics. 22:195–201.
2006.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zhang C, Vasmatzis G, Cornette JL and
DeLisi C: Determination of atomic desolvation energies from the
structures of crystallized proteins. J Mol Biol. 267:707–726.
1997.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Chen D and Meng J: Expression,
purification and immunogenicity of a novel hepatitis E virus-like
particle. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 22:339–342.
2006.PubMed/NCBI(In Chinese).
|
|
21
|
Xu M, Behloul N, Wen J, Zhang J and Meng
J: Role of asparagine at position 562 in dimerization and
immunogenicity of the hepatitis E virus capsid protein. Infect
Genet Evol. 37:99–107. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Ito K, Qin Y, Guarnieri M, Garcia T, Kwei
K, Mizokami M, Zhang J, Li J, Wands JR and Tong S: Impairment of
hepatitis B virus virion secretion by single-amino-acid
substitutions in the small envelope protein and rescue by a novel
glycosylation site. J Virol. 84:12850–12861. 2010.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Hawkins P, Prescott MJ, Carbone L,
Dennison N, Johnson C, Makowska IJ, Marquardt N, Readman G, Weary
DM and Golledge HD: A Good Death? Report of the second Newcastle
meeting on laboratory animal euthanasia. Animals (Basel).
6(50)2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zhang H, Dai X, Shan X and Meng J:
Characterization of antigenic epitopes of the ORF2 protein from
hepatitis E virus genotype 4. Virus Res. 142:140–143.
2009.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Meng J, Dubreuil P and Pillot J: A new
PCR-based seroneutralization assay in cell culture for diagnosis of
hepatitis E. J Clin Microbiol. 35:1373–1377. 1997.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Emerson SU, Clemente-Casares P, Moiduddin
N, Arankalle VA, Torian U and Purcell RH: Putative neutralization
epitopes and broad cross-genotype neutralization of Hepatitis E
virus confirmed by a quantitative cell-culture assay. J Gen Virol.
87:697–704. 2006.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Fiedler K and Simons K: The role of
N-glycans in the secretory pathway. Cell. 81:309–312.
1995.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zhou YH, Purcell RH and Emerson SU: An
ELISA for putative neutralizing antibodies to hepatitis E virus
detects antibodies to genotypes 1, 2, 3, and 4. Vaccine.
22:2578–2585. 2004.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Meng J, Dai X, Chang JC, Lopareva E,
Pillot J, Fields HA and Khudyakov YE: Identification and
characterization of the neutralization epitope(s) of the hepatitis
E virus. Virology. 288:203–211. 2001.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Purcell RH, Nguyen H, Shapiro M, Engle RE,
Govindarajan S, Blackwelder WC, Wong DC, Prieels JP and Emerson SU:
Pre-clinical immunogenicity and efficacy trial of a recombinant
hepatitis E vaccine. Vaccine. 21:2607–2615. 2003.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Li SW, Zhao Q, Wu T, Chen S, Zhang J and
Xia NS: The development of a recombinant hepatitis E vaccine HEV
239. Hum Vaccin Immunother. 11:908–914. 2015.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zhu FC, Zhang J, Zhang XF, Zhou C, Wang
ZZ, Huang SJ, Wang H, Yang CL, Jiang HM, Cai JP, et al: Efficacy
and safety of a recombinant hepatitis E vaccine in healthy adults:
A large-scale, randomised, double-blind placebo-controlled, phase 3
trial. Lancet. 376:895–902. 2010.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Gurramkonda C, Adnan A, Gäbel T, Lünsdorf
H, Ross A, Nemani SK, Swaminathan S, Khanna N and Rinas U: Simple
high-cell density fed-batch technique for high-level recombinant
protein production with Pichia pastoris: Application to
intracellular production of hepatitis B surface antigen. Microb
Cell Fact. 8(13)2009.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Lorenzo FR, Tanaka T, Takahashi H,
Ichiyama K, Hoshino Y, Yamada K, Inoue J, Takahashi M and Okamoto
H: Mutational events during the primary propagation and consecutive
passages of hepatitis E virus strain JE03-1760F in cell culture.
Virus Res. 137:86–96. 2008.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Okamoto H: Hepatitis E virus cell culture
models. Virus Res. 161:65–77. 2011.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Yamamoto H, Suzuki J, Matsuda A, Ishida T,
Ami Y, Suzaki Y, Adachi I, Wakita T, Takeda N and Li TC: Hepatitis
E virus outbreak in monkey facility, Japan. Emerg Infect Dis.
18:2032–2034. 2012.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Takahashi K, Kitajima N, Abe N and Mishiro
S: Complete or near-complete nucleotide sequences of hepatitis E
virus genome recovered from a wild boar, a deer, and four patients
who ate the deer. Virology. 330:501–505. 2004.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Li S, Tang X, Seetharaman J, Yang C, Gu Y,
Zhang J, Du H, Shih JW, Hew CL, Sivaraman J and Xia N: Dimerization
of hepatitis E virus capsid protein E2s domain is essential for
virus-host interaction. PLoS Pathog. 5(e1000537)2009.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Jameel S, Zafrullah M, Ozdener MH and
Panda SK: Expression in animal cells and characterization of the
hepatitis E virus structural proteins. J Virol. 70:207–216.
1996.PubMed/NCBI View Article : Google Scholar
|