Introduction
Hyperlipidemia is a disorder of lipid metabolism
that causes blood lipid levels to increase to abnormally high
levels, including increases in the levels of low-density
lipoprotein-cholesterol (LDL-c) and total cholesterol (TC). In
addition, hyperlipidemia can cause kidney damage (1). Clinical studies have suggested a
causative role of dyslipidemia in the development and progression
of CKD and its complications (2,3).
Increasing evidence has shown that lipid deposition, oxidative
stress, fibrosis and inflammation are major pathophysiological
mechanisms of hyperlipidemia-induced kidney damage, which may
progress to chronic kidney disease (CKD) (4,5).
Exercise has been shown to reduce kidney damage in several studies.
Zeynali et al (6) suggested
that aerobic exercise may reduce cisplatin-induced nephrotoxicity,
and a favorable effect on renal function was induced by increasing
the activation of the antioxidant system (6). In an animal model of high consumption
of cola soft drinks, it has been observed that moderately intense
exercise reduced the kidney damage caused by cola (7). In addition, Braun et al
(8) showed that free-running wheel
exercise may attenuate the long-term effects of high blood pressure
in spontaneously hypertensive rats with kidney damage (8). Apolipoprotein E-deficient
(ApoE-/-) mice, a well-established animal model of
hyperlipidemia, have been extensively used to study the effects of
kidney damage on atherosclerosis and renal injury (9-11).
A hyperlipidemia-induced kidney damage animal model using
ApoE-/- mice was established in the present study by
administering a high-fat diet (HD) and subjecting the mice to
exercise in the form of swimming for 40 min/day, 5 days/week for 12
weeks. Whether exercise reduced renal damage caused by
hyperlipidemia, and the underlying mechanisms, was assessed.
Materials and methods
Animals
For establishment of an animal model of
hyperlipidemia-induced kidney damage, 8-week-old male
ApoE-/- mice (n=28; weight, 24.30±1.04 g) were obtained
from Beijing Vital River Laboratories Animal Technology Co., Ltd.
Mice were provided with ad libitum access to food and water,
and were maintained under constant conditions, including 40-60%
humidity, 24-26˚C and a 12-h light/dark cycle. ApoE-/-
mice were randomly divided into four groups as follows: Mice fed a
normal diet (ND group); mice fed a ND and exercised (ND + E group);
mice fed a HD (HD group); and mice fed a HD and exercised (HD + E
group). Based on a previous study, the number of mice in each group
was seven (12). The HD mouse food
consisted of 1.25% (w/w) cholesterol, 22.5% (w/w) protein, 20.0%
(w/w) cocoa fat and 45.0% carbohydrate (cat. no. MD12017; Jiangsu
Medicience Co., Ltd.; https://www.medicience.com/proshow_13.html). Exercise
training was initiated in an experimental swimming pool
(temperature, 30˚C; water depth, 44 cm; radius, 120 cm) 1 week
prior to administration of the experimental diets. The progressive
exercise program initially involved swimming for 5-10 min and was
gradually extended to 30 min/day. When the experimental diets were
administered, mice were subjected to swimming exercise for 40
min/day, 5 days/week for 12 weeks. After 12 weeks, the mice were
euthanized with a high dose of pentobarbital (100 mg/kg,
intraperitoneally), and lack of respiration and heartbeat was used
as an indicator of mouse death. All animal experiments were
approved by the Ethics Committee of the Zhejiang Rongjun Hospital
(Jiaxing, China).
Biochemical measurements
Blood samples were obtained from the inferior vena
cava and preserved in tubes. The blood samples were immediately
centrifuged at 1,006 x g for 10 min at 4˚C after collection, and
the serum were subsequently stored at -80˚C. Total cholesterol (TC;
cat. no. A111-1-1), triglyceride (TG; cat. no. A110-1-1), LDL-c
(cat. no. A113-1-1), creatinine (CRE; cat. no. C011-2-1),
superoxide dismutase (SOD; cat. no. A011-3-2) and glutathione
peroxidase (GSH-PX; cat. no. A005-1-2) levels were measured using
ELISA kits (Nanjing Jiancheng Bioengineering Institute) according
to the manufacturer's protocols. The concentrations of TC, TG,
LDL-c, CRE, SOD and GSH-PX were calculated based on measurement of
the optical density at the respective wavelengths for each compound
according to the manufacturer's protocol.
Morphological analysis
Kidney tissues were fixed in 10% buffered formalin
solution for 30 min at room temperature and dehydrated in 75%
ethanol overnight, followed by paraffin embedding. Serial sections
(4 µm, n=3/group randomly selected) were stained with hematoxylin
for 15 min and eosin for 5 min at temperature to assess
pathological changes using a BX40 upright light microscope (Olympus
Corporation). Kidney damage scores were determined according to the
extent of kidney injury, as previously described (13,14),
by two blinded researchers. Scoring was primarily based on the
presence or absence of hemorrhaging, tubular cell necrosis, tubular
dilatation and cytoplasmic vacuole formation. The grading system
was scored as follows: 0, 0% damage (normal kidney); 1, 0-5% damage
(minimal damage); 2, 5-25% damage (mild damage); 3, 25-75% damage
(moderate damage); and 4, 75-100% damage (severe damage).
Masson's trichrome and Periodic
acid-Schiff (PAS) staining
Kidney-tissue sections (n=3/group) were
deparaffinized via immersion in xylene (three times, 5 min each)
and rehydrated in a descending alcohol series (100, 90, 80 and 70%
alcohol; 5 min each). Slides were stained using Masson's trichrome
and PAS staining to investigate changes in kidney tissues. In
Masson's trichrome staining, after deparaffinization and
rehydration the procedure was as follows: i) Fixation in Bouin
liquor overnight at 4˚C, followed by washing in running water until
the yellow color disappeared and rinsing in two changes of
distilled water; ii) staining with Mayer's Hematoxylin for 5 min at
room temperature, followed by immersion in 0.5% hydrochloric acid
in 70% alcohol for 5 sec at room temperature, washing in running
tap water for 30 sec and rinsing in two changes of distilled water;
iii) staining with acid ponceau for 5 to 10 min at room
temperature, followed by rinsing in three changes of distilled
water; iv) immersion in 1% phosphomolybdic acid aqueous solution;
and v) staining with aniline blue for 5 min at room temperature,
followed by immersion in 1% glacial acetic acid for 5 min at room
temperature, dehydration of the stained sections and sealing using
resin glue. In PAS staining, after deparaffinization and
rehydration, the sections were placed in 1% periodic acid for 15
min at room temperature followed by washing with water, incubation
with Schiff 's reagent (MilliporeSigma) for 20 min at room
temperature and staining with Gill's Hematoxylin (Thermo Fisher
Scientific, Inc.) for 3 min at room temperature. All sections were
examined using a BX40 upright light microscope (Olympus BX43;
Olympus Corporation). Blue staining indicated collagen accumulation
in Masson's trichrome staining and red staining indicated lipid
deposition in PAS staining.
Immunohistochemistry (IHC)
Paraffin-embedded sections (n=3/group) were
deparaffinized with xylene and rehydrated through a graded series
of ethanol, as described above. Sections were treated with 3%
H2O2 in methanol for 15 min to inactivate
endogenous peroxidases and were then incubated with rabbit
anti-NF-κB (cat. no. 10745-1-AP), anti- nuclear factor
erythroid-2-related factor 2 (NRF2, cat. no. 16396-1-AP),
anti-Smad3 (cat. no. 25494-1-AP) and anti-TGF-β (cat. no.
18978-1-AP) antibody (all at 1:200 dilution; ProteinTech Group,
Inc.) overnight at 4˚C. The tissues were incubated with the
secondary antibody (HRP-labeled goat anti-rabbit IgG) from
N-Histofine Simple stain kit (cat. no. 414341F Nichirei Biosciences
Inc.) for 30 min at room temperature. Subsequently, the signal was
visualized using 3,3'-diaminobenzidine (Metal Enhanced DAB
Substrate Kit; cat. no. DA1015; Beijing Solarbio Science &
Technology Co., Ltd.) for 10 min and the development was stopped by
adding distilled water, according to the manufacturer's
instructions. Finally, the stained sections were dehydrated in an
ascending series of ethanol concentrations (70, 80, 90 and 100%
ethanol; 5 min each) and sealed using resin glue. All sections were
examined using a BX40 upright light microscope (Olympus BX43;
Olympus Corporation). Three sections were randomly selected from
each group. Semi-quantitative assessment of the NF-κB positive
cells was performed using ImageJ software version 1.8.0 (National
Institutes of Health).
Western blotting
Proteins were extracted from kidney tissues
(n=3/group) using RIPA lysis buffer (cat. no. P0013B; Beyotime
Institute of Biotechnology) according to the manufacturer's
protocol. Protein concentrations were determined using a BCA
protein assay kit (cat. no. DQ111-01; Beijing Transgen Biotech Co.,
Ltd.). Protein samples (20 µg per lane) were separated by SDS-PAGE
on 10-15% gels and were transferred to PVDF membranes.
Subsequently, membranes were blocked in Tris-buffered saline with
0.1% Tween-20 (TBS-T) containing 5% skimmed milk and then incubated
at room temperature for 2 h. The membranes were then incubated with
primary antibodies diluted in primary antibody diluent (cat. no.
P0023A; Beyotime Institute of Biotechnology) and lightly shaken
overnight at 4˚C. Primary rabbit anti-Smad3 (1:1,000 dilution; cat.
no. 25494-1-AP), anti-TGF-β (1:1,000 dilution; cat. no.
18978-1-AP), anti-NF-κB (1:1,000 dilution; cat. no. 10745-1-AP) and
anti-β-actin antibodies (1:1,000 dilution; cat. no. 20536-1-AP; all
from ProteinTech Group, Inc.) were used. After washing with TBS-T
three times (15 min each), membranes were incubated with a
secondary antibody (HRP-conjugated goat anti-rabbit IgG; 1:2,000;
cat. no. SA00001-2; ProteinTech Group, Inc.) for 1 h at 37˚C.
Enhanced chemiluminescence reagent (cat. no. 32106; Thermo Fisher
Scientific, Inc.) was used to visualize bands. Signals were imaged
using a Bio-Rad imaging system (Bio-Rad Laboratories, Inc.) with a
Chemi 410 HR camera (Analytik Jena AG) and analyzed using Gel-Pro
Analyzer version 4.0 (Media Cybernetics, Inc.). The analysis was
performed independently three times. Densitometry analysis was
performed using ImageJ software version 1.8.0 (National Institutes
of Health) as described previously (15).
Statistical analysis
All data are presented as mean ± standard error of
the mean. SPSS software version 23.0 (IBM Corp.) was used to
analyze the data. Differences between multiple groups were measured
using a one-way ANOVA with a post-hoc Tukey's test. Kidney damage
scores are expressed as median values and were analyzed using a
Kruskal-Wallis test with post hoc Dunn's test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Metabolic characterization
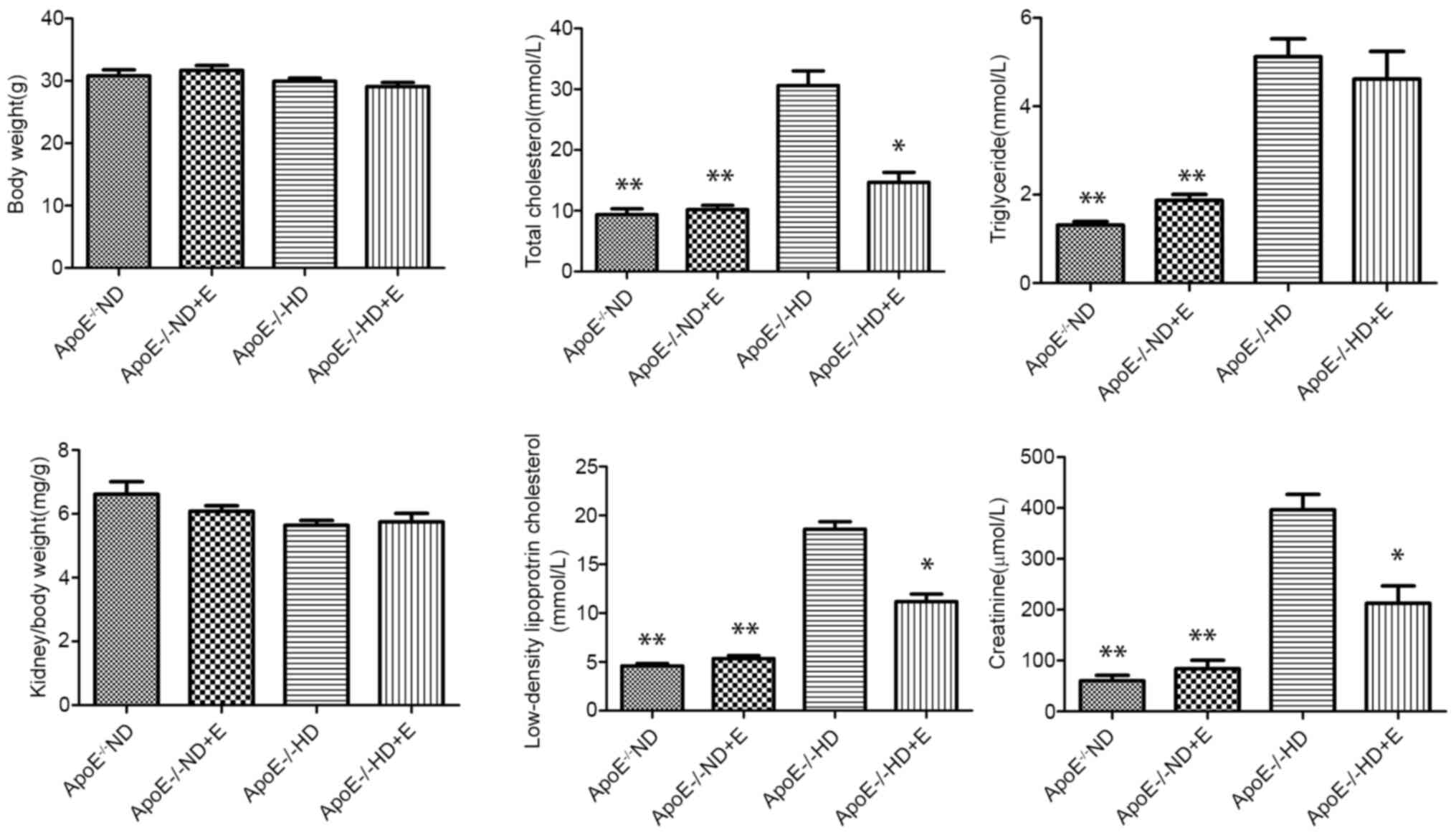
The metabolic characteristics of the animals are
shown in Fig. 1. Body weight did
not differ significantly amongst the four groups. The
ApoE-/- HD and HD + E groups showed significantly
increased LDL-c, TC, TG and CRE levels compared with those in the
ND and ND + E groups. In addition, the levels of TC, LDL-c and CRE
were significantly higher in the HD group compared with those in
the HD + E group, whereas TG levels did not differ significantly.
These results suggested that exercise was effective in reducing TC,
LDL-c and CRE levels in mice fed a HD, but was less effective in
terms of TG status.
Exercise induces histopathological
changes in the kidney tissues of ApoE-/-mice fed a
HD
H&E, Masson's trichrome and PAS staining were
used to evaluate histopathological changes in renal tissues
(Fig. 2). Kidney samples from
ApoE-/- ND mice appeared normal. ApoE-/- HD
mice exhibited obvious renal lipid deposition and pro-inflammatory
cell infiltration compared with in the ND mice. H&E staining
results showed that the kidneys of the HD group mice displayed
hemorrhaging, inflammatory infiltration, detachment and swelling of
tubular epithelial cells, interstitial edema, tubular cell casts,
dilatation and necrosis (Fig. 2A).
Kidney damage scoring showed that exercise treatment significantly
decreased renal injury in the HD group, consistent with H&E
results (Fig. 2B). Collagen
deposition was determined using Masson's staining. Heavy collagen
deposition was observed in the HD group. Notably, this damage was
suppressed in the ApoE-/- HD + E mice (Fig. 2C).
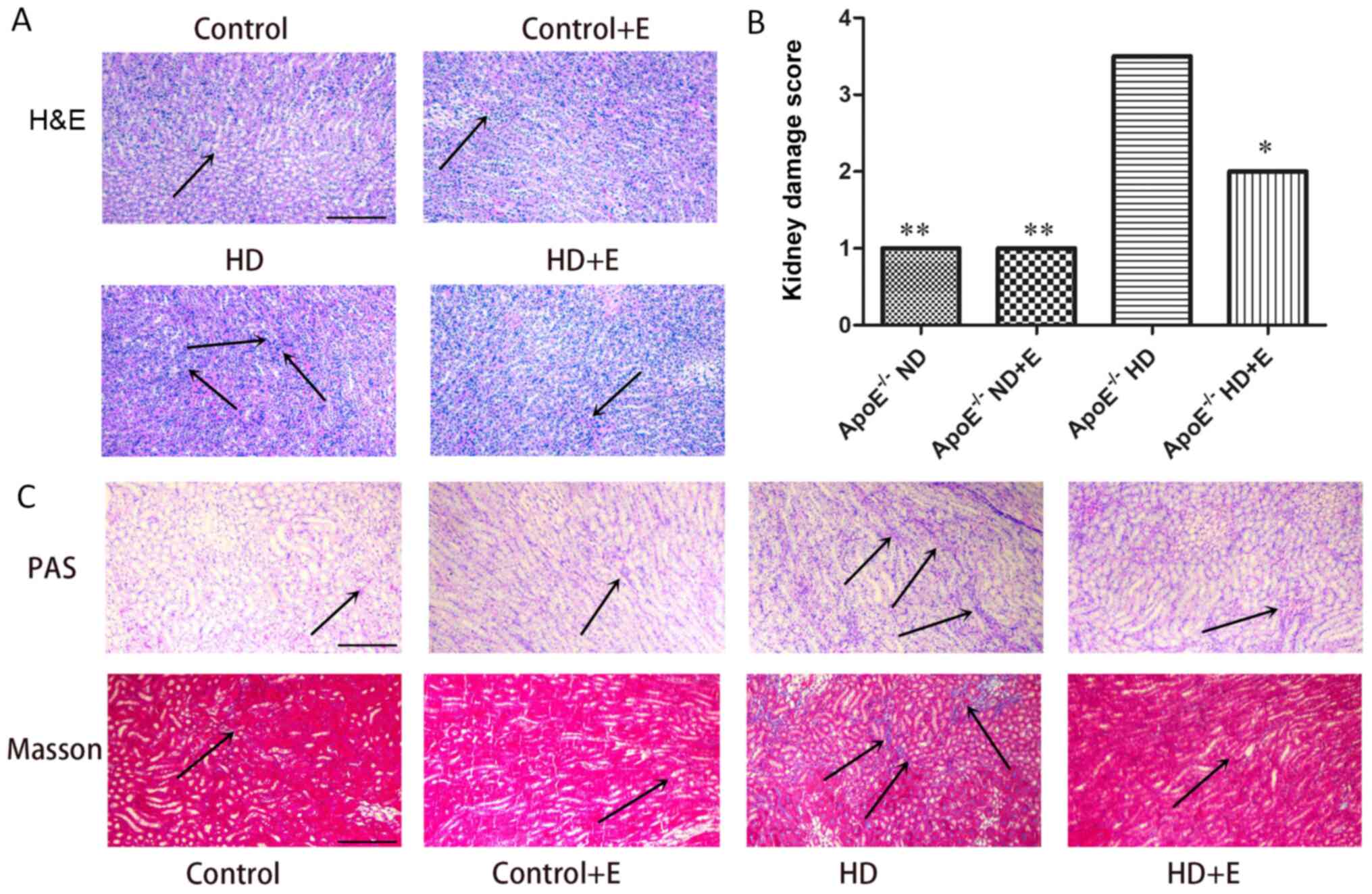 | Figure 2Histological analysis of kidney
samples. (A) Notably reduced inflammatory infiltration was observed
in the kidney tissue of the mice in the HD+E group compared with
the ND group of mice, as determined by H&E staining. Arrows
indicate inflammatory infiltration. (B) Kidney damage scores are
expressed as the median and were analyzed using the Kruskal-Wallis
test and Dunn's post hoc test. Semi-quantitative injury scores
ranged from 0 to 4 [0, normal kidney; 1, minimal damage (0-5%
injury); 2, mild damage (5-25% injury); 3, moderate damage (25-75%
injury); and 4, severe damage (75-100% injury)].
*P<0.05, **P<0.01 vs.
ApoE-/- HD group. (C) Masson and PAS staining in the
kidney tissues with different treatments. Exercise reduced lipid
deposition, collagen deposition and fibrosis in ApoE-/-
HD group mice. Scale bar, 100 µm. n=3/group. Masson staining:
Collagen fibers, mucus and cartilage are blue; muscle fibers,
cellulose and red blood cells are red; and the nucleus is blue and
black. The arrows indicate damage. Data are presented as the mean ±
standard error of the mean. ApoE-/-, apolipoprotein
E-deficient; HD, high-fat diet; ND, normal diet; E, exercise; PAS,
Periodic acid-Schiff. |
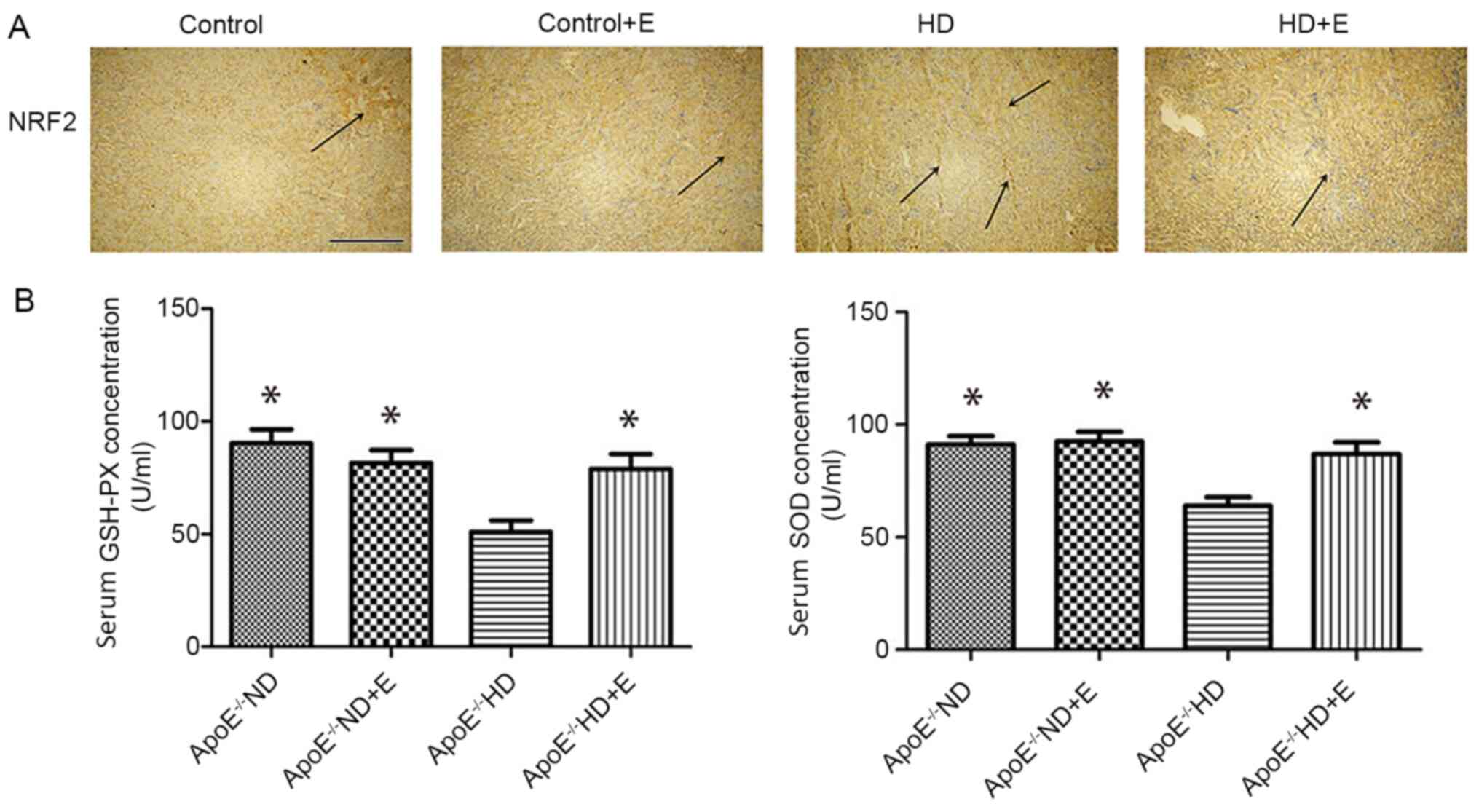
Oxidative stress characteristics
Images of NRF2 expression in the IHC-stained kidney
samples are presented in Fig. 3A.
Compared with the ND and ND + E groups, the expression of NRF2 in
the ApoE-/- HD group was visibly increased. Exercise
reduced NRF2 expression in the ApoE-/- HD + E group
compared with that in the ApoE-/- HD group. Serum levels
of GSH and SOD are shown in Fig. 3B
and C. GSH and SOD levels were
significantly higher in the ApoE-/- HD + E mice compared
with those in the ApoE-/- HD mice.
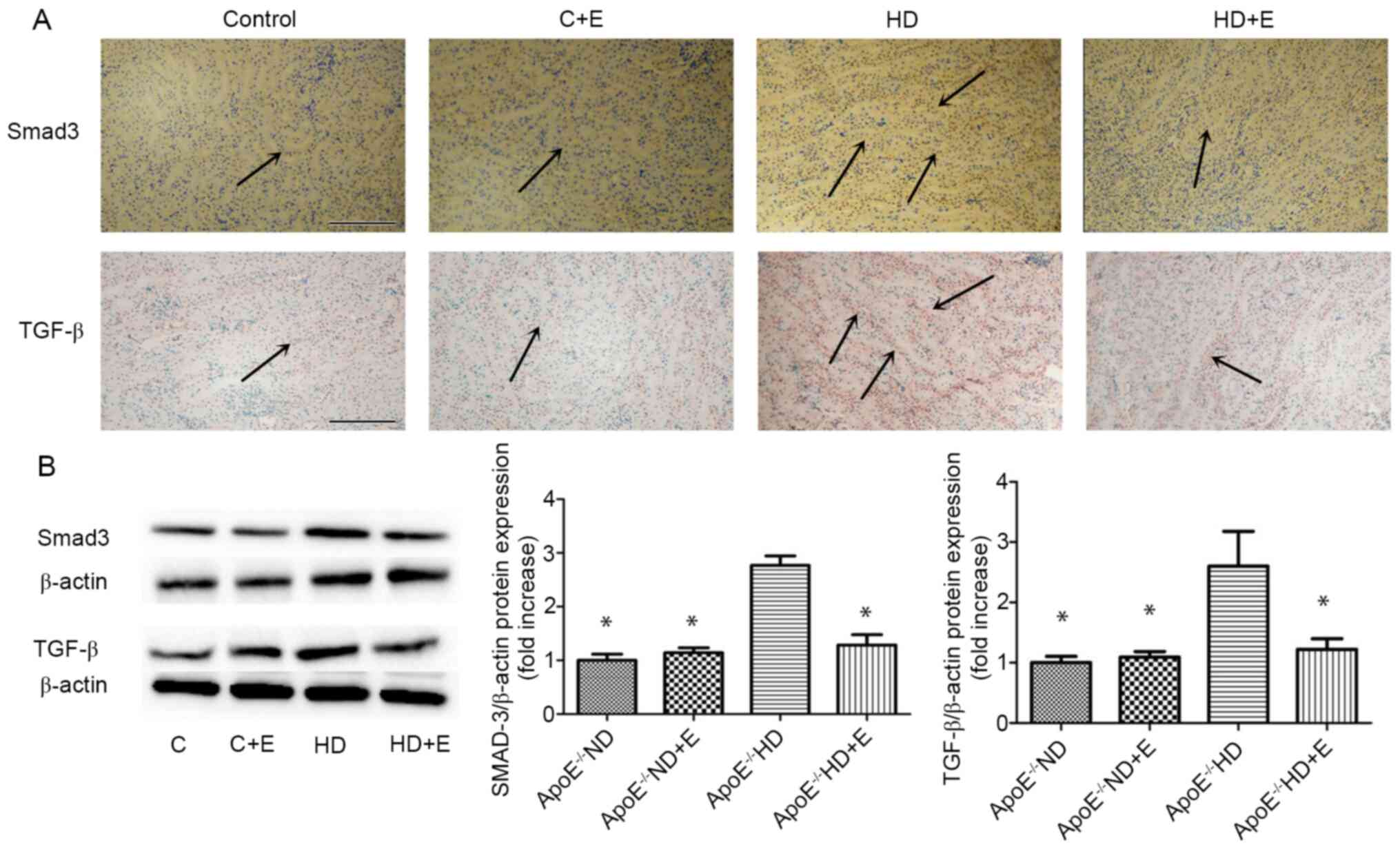
Characteristics of fibrosis
To investigate the mechanism of fibrosis in kidney
damage, TGF-β and Smad3 expression was visualized using IHC
(Fig. 4A) and immunoblotting
(Fig. 4B). IHC staining showed that
the numbers of TGF-β- and Smad3-positive cells in the
ApoE-/- HD mice were markedly increased compared with
the ND group. However, exercise markedly reduced this increase.
Compared with in the ApoE-/- HD mice, HD + E mice
exhibited significantly reduced TGF-β and Smad3 expression levels,
as determined by immunoblotting (Fig.
4B). These results indicated that exercise reduced TGF-β and
Smad3 expression in ApoE-/- HD mice.
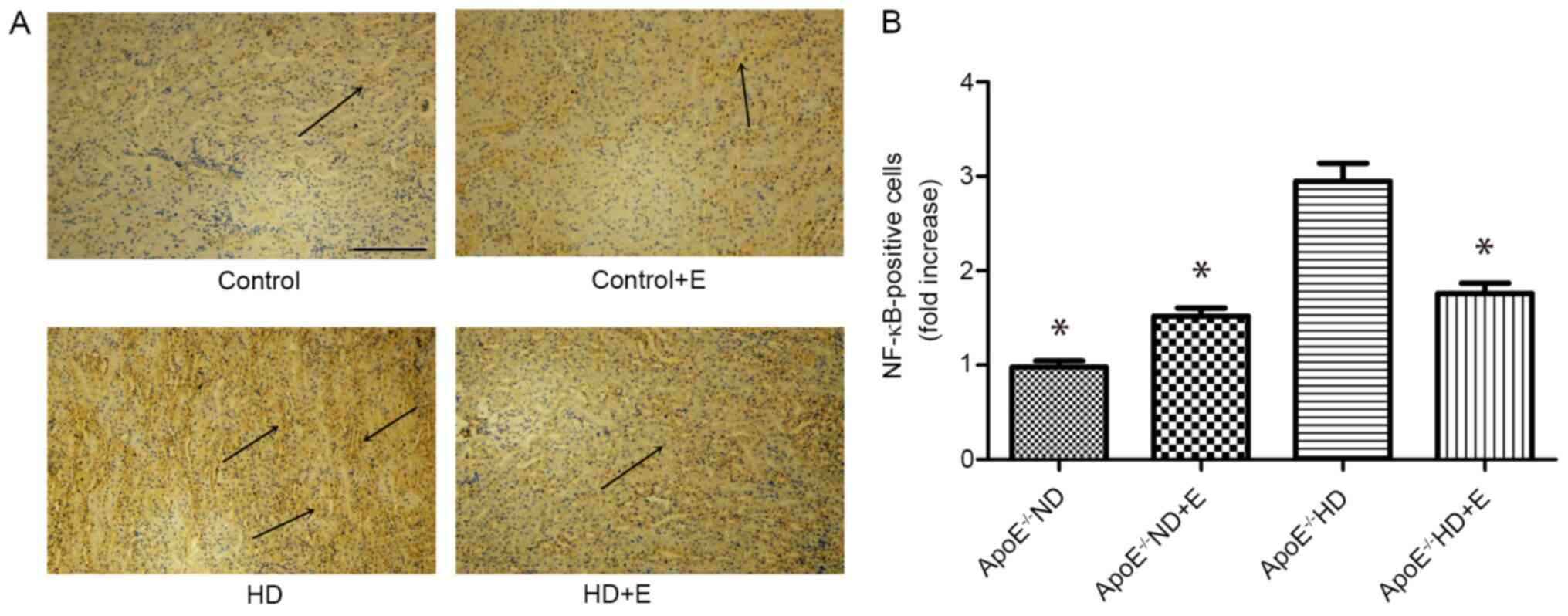
NF-κB signaling pathway
IHC analysis of NF-κB was used to investigate kidney
damage caused by hyperlipidemia (Fig.
5). NF-κB protein expression in kidney tissues was lower in the
ApoE-/- HD + E mice compared with that in the
ApoE-/- HD mice.
Discussion
The present study demonstrated that exercise may
exert a protective effect against kidney damage induced by
hyperlipidemia, including limiting the progression of lipid
deposition, oxidative stress and fibrosis. Compared with in
ApoE-/- mice that consumed a ND, higher LDL-c, TG and TC
levels were observed in ApoE-/- mice that consumed a HD,
in agreement with Faran et al (16). Furthermore, CRE levels were higher
in the HD group compared with those in the ND group. These results
suggested that a hyperlipidemia-induced kidney damage model was
established in the ApoE-/- mice. Notably, LDL-c, TC and
CRE levels were significantly lower in the HD + E mice compared
with those in the HD mice, suggesting that exercise exerted a
protective effect in reducing LDL-c, TC and CRE levels in mice with
kidney damage by progressive lipid deposition, but was less
effective in altering TG levels. Several studies have indicated
that exercise can attenuate kidney damage caused by other factors,
such as cisplatin (6), cola
(7) and hypertension (8).
Hyperlipidemia is a major independent risk factor
for the development of kidney disease (17). In the present study, using H&E,
PAS and Masson staining, it was shown that kidney tissue disorders,
lipid deposition, inflammatory cell infiltration, collagen
accumulation and increased fibrosis resulted in kidney damage in
the HD group. However, kidney damage was significantly reduced in
the HD + E group. These results suggested that exercise may reduce
kidney tissue damage in ApoE-/- mice fed a HD.
Oxidative stress and inflammation are important
characteristics of CKD, and can induce its progression (18,19).
The balance between oxidative stress and antioxidant defenses
maintains stability in living organisms (20). Moreover, hypercholesterolemia has
been reported to cause exacerbation of inflammation and increase
oxidative stress in kidney tissues (21,22).
Napoli and Lerman (23) showed that
increased free radical production was associated with increased
accumulation of cholesterol in serum and tissue (23). Increased lipid peroxidation is
frequently observed concurrent with a reduction in endogenous
antioxidants, such as SOD, catalase and GSH-PX (24). In the present study, SOD and GSH-PX
levels were decreased in the HD group compared with those in the ND
group, suggesting that the HD-induced lipid deposition and
disrupted the oxidative stress/antioxidant defense balance.
However, SOD and GSH levels were increased in the HD + E group
compared with those in the HD group, suggesting that exercise
reduced lipid peroxidation and enhanced antioxidant activity. NRF2
is a key regulator of the cellular response to oxidative stress
(25). Physiologically, NRF2 is
maintained in an inactive state through binding with Keap1 in the
cytoplasm (26). Oxidative stress
can induce nuclear accumulation of NRF2, upregulate downstream
antioxidant gene transcription and promote the expression of
antioxidant enzymes (27,28). In the present study, it was shown
that the protein expression levels of NRF2 were higher in the HD
group compared with those in the ND group, suggesting that
oxidative stress was activated. Additionally, less oxidative stress
was observed in the HD + E group compared with that in the HD
group, as estimated by the lower expression levels of NRF2. Thus,
exercise resulted in a protective effect against kidney damage
caused by hyperlipidemia through attenuation of oxidative stress.
Ishikawa et al (29)
demonstrated that exercise can alleviate diabetic renal injury
through reduced renal oxidative stress and inflammation, in
agreement with the results of the present study.
Fibrosis is a characteristic of CKD and has been
recognized as an independent predictor of the progression of kidney
disease (30). In diabetic
glomerular injury, oxidative stress has been shown to induce mRNA
expression of TGF-β (31). TGF-β
acts through a canonical signaling pathway that involves
phosphorylation and activation of Smad3 by the TGF-β receptor, and
then induces renal fibrosis (32).
Renal fibrosis can thus be accelerated by increasing the levels of
TGF-β and Smad3 (31,32). In the present study, collagen
deposition was determined using Masson's staining. Heavy collagen
deposition was prominently observed in the HD group, whereas
exercise reduced collagen deposition in the HD + E group. In
addition, the expression of TGF-β and Smad3 was examined using IHC
and immunoblotting. Compared with in the ApoE-/- HD
group, TGF-β and Smad3 were significantly suppressed in mice in the
ApoE-/- HD + E group. This result indicated that
exercise training reduced renal fibrosis caused by
hyperlipidemia.
Several studies have shown that the NF-κB signaling
pathway is associated with fibrosis (33,34).
NF-κB is constitutively expressed in various types of tissues
during inflammation and fibrosis (35-38).
Inhibiting the activation of activated hepatic stellate cells to
alleviate NF-κB signaling has been shown to contribute to the
treatment of hepatic fibrosis (39). Furthermore, Zhang et al
(40) showed that all-trans
retinoic acid suppressed epidural fibrosis by regulating the NF-κB
signaling pathway (40). To
investigate the inflammatory responses in kidney damage caused by
hyperlipidemia, IHC analysis of NF-κB was performed. NF-κB protein
expression in kidney tissues was lower in the ApoE-/- HD
+ E mice compared with in the ApoE-/-HD mice, thus
suggesting that exercise may regulate the NF-κB pathway, and
improve inflammation and oxidative stress status in
hyperlipidemia-induced kidney damage.
In conclusion, the results of the present study
showed that exercise exhibited a protective effect against kidney
damage caused by hyperlipidemia. Thus, exercise may be an
additional means of clinical management of CKD.
Acknowledgements
Not applicable.
Funding
This study was supported by funding from Jiaxing Key
Discipline Construction Fund (grant no. 2019-22).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YS and HL designed the present study. CQ and QY
performed the experiments. HZ, XY and LG analyzed and interpreted
the results of experiments. CQ prepared figures. QY drafted the
manuscript. LG revised the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Ethics
Committee of Zhejiang Rongjun Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ding M, Si D, Zhang W, Feng Z, He M and
Yang P: Red yeast rice repairs kidney damage and reduces
inflammatory transcription factors in rat models of hyperlipidemia.
Exp Ther Med. 8:1737–1744. 2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Peev V, Nayer A and Contreras G:
Dyslipidemia, malnutrition, inflammation, cardiovascular disease
and mortality in chronic kidney disease. Curr Opin Lipidol.
25:54–60. 2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Ruan XZ, Varghese Z and Moorhead JF: An
update on the lipid nephrotoxicity hypothesis. Nat Rev Nephrol.
5:713–721. 2009.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Pei Z, Zhu L, Liu Y, Li N, Yang G and Liu
H: Thymoquinone reduces kidney damage in apolipoprotein E-deficient
mice fed a high-cholesterol diet. RSC Advances. 83:53002–53009.
2017.
|
|
5
|
Pei Z, Okura T, Nagao T, Enomoto D, Kukida
M, Tanino A, Miyoshi K, Kurata M and Higaki J: Osteopontin
deficiency reduces kidney damage from hypercholesterolemia in
Apolipoprotein E-deficient mice. Sci Rep. 6(28882)2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zeynali F, Nematbakhsh M, Mojtahedi H,
Poorshahnazari A, Talebi A, Pezeshki Z, Mazaheri S and Moslemi F:
Protective role of aerobic exercise against cisplatin-induced
nephrotoxicity in rats. Asian J Sports Med.
6(e24901)2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Cao G, González J, Müller A, Ottaviano G,
Ambrosio G, Toblli JE and Milei J: Beneficial effect of moderate
exercise in kidney of rat after chronic consumption of cola drinks.
PLoS One. 11(e0152461)2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Braun K, Atmanspacher F, Schreckenberg R,
Grgic I and Schlüter KD: Effect of free running wheel exercise on
renal expression of parathyroid hormone receptor type 1 in
spontaneously hypertensive rats. Physiol Rep.
6(e13842)2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Piedrahita JA, Zhang SH, Hagaman JR,
Oliver PM and Maeda N: Generation of mice carrying a mutant
apolipoprotein E gene inactivated by gene targeting in embryonic
stem cells. Proc Natl Acad Sci USA. 89:4471–4475. 1992.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Sastre C, Rubio-Navarro A, Buendía I,
Gómez-Guerrero C, Blanco J, Mas S, Egido J, Blanco-Colio LM, Ortiz
A and Moreno JA: Hyperlipidemia-associated renal damage decreases
Klotho expression in kidneys from ApoE knockout mice. PLoS One.
8(e83713)2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Muñoz-García B, Moreno JA, López-Franco O,
Sanz AB, Martín-Ventura JL, Blanco J, Jakubowski A, Burkly LC,
Ortiz A, Egido J, et al: Tumor necrosis factor-like weak inducer of
apoptosis (TWEAK) enhances vascular and renal damage induced by
hyperlipidemic diet in ApoE-knockout mice. Arterioscler Thromb Vasc
Biol. 29:2061–2068. 2009.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Xu J, Zhu L, Liu H, Li M, Liu Y, Yang F
and Pei Z: Thymoquinone reduces cardiac damage caused by
hypercholesterolemia in apolipoprotein E-deficient mice. Lipids
Health Dis. 17(173)2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Hong X, Zhao X, Wang G, Zhang Z, Pei H and
Liu Z: Luteolin treatment protects against renal ischemia
reperfusion injury in rats. Mediators Inflamm.
2017(9783893)2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Shingu C, Koga H, Hagiwara S, Matsumoto S,
Goto K, Yokoi I and Noguchi T: Hydrogen-rich saline solution
attenuates renal ischemia-reperfusion injury. J Anesth. 24:569–574.
2010.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Rasband WS: ImageJ, U.S. National
Institutes of Health, Bethesda, MD, 1997-2018.
|
|
16
|
Faran SA, Asghar S, Khalid SH, Khan IU,
Asif M, Khalid I, Gohar UF and Hussain T: Hepatoprotective and
renoprotective properties of lovastatin-loaded ginger and garlic
oil nanoemulsomes: Insights into serum biological parameters.
Medicina (Kaunas). 55(579)2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Schaeffner ES, Kurth T, Curhan GC, Glynn
RJ, Rexrode KM, Baigent C, Buring JE and Gaziano JM: Cholesterol
and the risk of renal dysfunction in apparently healthy men. J Am
Soc Nephrol. 14:2084–2091. 2003.PubMed/NCBI
|
|
18
|
Himmelfarb J and Hakim RM: Oxidative
stress in uremia. Curr Opin Nephrol Hypertens. 12:593–598.
2003.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Vaziri ND: Roles of oxidative stress and
antioxidant therapy in chronic kidney disease and hypertension.
Curr Opin Nephrol Hypertens. 13:93–99. 2004.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Blokhina O, Virolainen E and Fagerstedt
KV: Antioxidants, oxidative damage and oxygen deprivation stress: A
review. Ann Bot. 91:179–194. 2003.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Deepa PR and Varalakshmi P: Favourable
modulation of the inflammatory changes in hypercholesterolemic
atherogenesis by a low-molecular-weight heparin derivative. Int J
Cardiol. 106:338–347. 2006.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Scheuer H, Gwinner W, Hohbach J, Gröne EF,
Brandes RP, Malle E, Olbricht CJ, Walli AK and Gröne HJ: Oxidant
stress in hyperlipidemia-induced renal damage. Am J Physiol Renal
Physiol. 278:F63–F74. 2000.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Napoli C and Lerman LO: Involvement of
oxidation-sensitive mechanisms in the cardiovascular effects of
hypercholesterolemia. Mayo Clin Proc. 76:619–631. 2001.PubMed/NCBI View
Article : Google Scholar
|
|
24
|
Kaplowitz N: Mechanisms of liver cell
injury. J Hepatol. 32 (Suppl 1):39–47. 2000.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Loboda A, Damulewicz M, Pyza E, Jozkowicz
A and Dulak J: Role of Nrf2/HO-1 system in development, oxidative
stress response and diseases: An evolutionarily conserved
mechanism. Cell Mol Life Sci. 73:3221–3247. 2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Periyasamy P and Shinohara T: Age-related
cataracts: Role of unfolded protein response, Ca2+
mobilization, epigenetic DNA modifications, and loss of Nrf2/Keap1
dependent cytoprotection. Prog Retin Eye Res. 60:1–19.
2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Calkins MJ, Johnson DA, Townsend JA,
Vargas MR, Dowell JA, Williamson TP, Kraft AD, Lee JM, Li J and
Johnson JA: The Nrf2/ARE pathway as a potential therapeutic target
in neurodegenerative disease. Antioxid Redox Signal. 11:497–508.
2009.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Petri S, Körner S and Kiaei M: Nrf2/ARE
signaling pathway: Key mediator in oxidative stress and potential
therapeutic target in ALS. Neurol Res Int.
2012(878030)2012.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ishikawa Y, Gohda T, Tanimoto M, Omote K,
Furukawa M, Yamaguchi S, Murakoshi M, Hagiwara S, Horikoshi S,
Funabiki K, et al: Effect of exercise on kidney function, oxidative
stress, and inflammation in type 2 diabetic KK-Ay mice. Exp
Diabetes Res. 2012(702948)2012.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Campanholle G, Ligresti G, Gharib SA and
Duffield JS: Cellular mechanisms of tissue fibrosis. 3. Novel
mechanisms of kidney fibrosis. Am J Physiol Cell Physiol.
304:C591–C603. 2013.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Jeong SI, Kim KJ, Choo YK, Keum KS, Choi
BK and Jung KY: Phytolacca americana inhibits the high
glucose-induced mesangial proliferation via suppressing
extracellular matrix accumulation and TGF-beta production.
Phytomedicine. 11:175–181. 2004.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Meng XM, Nikolic-Paterson DJ and Lan HY:
TGF-β: The master regulator of fibrosis. Nat Rev Nephrol.
12:325–338. 2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Domino M, Jasinski T, Kautz E,
Juszczuk-Kubiak E, Ferreira-Dias G, Zabielski R, Sady M and
Gajewski Z: Expression of genes involved in the NF-κB-dependent
pathway of the fibrosis in the mare endometrium. Theriogenology.
147:18–24. 2020.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Luedde T and Schwabe RF: NF-κB in the
liver--linking injury, fibrosis and hepatocellular carcinoma. Nat
Rev Gastroenterol Hepatol. 8:108–118. 2011.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Umezawa K: Possible role of peritoneal
NF-κB in peripheral inflammation and cancer: Lessons from the
inhibitor DHMEQ. Biomed Pharmacother. 65:252–259. 2011.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Lind DS, Hochwald SN, Malaty J, Rekkas S,
Hebig P, Mishra G, Moldawer LL, Copeland EM III and Mackay S:
Nuclear factor-kappa B is upregulated in colorectal cancer.
Surgery. 130:363–369. 2001.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Sosińska P, Baum E, Maćkowiak B,
Staniszewski R, Jasinski T, Umezawa K and Bręborowicz A: Inhibition
of NF-kappaB with Dehydroxymethylepoxyquinomicin modifies the
function of human peritoneal mesothelial cells. Am J Transl Res.
8:5756–5765. 2016.PubMed/NCBI
|
|
38
|
Sun L, Zhang S, Chang Q and Tan J:
Establishment and comparison of different intrauterine adhesion
modelling procedures in rats. Reprod Fertil Dev.
31(1360)2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Mann DA and Marra F: Fibrogenic signalling
in hepatic stellate cells. J Hepatol. 52:949–950. 2010.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Zhang C, Kong X, Ning G, Liang Z, Qu T,
Chen F, Cao D, Wang T, Sharma HS and Feng S: All-trans retinoic
acid prevents epidural fibrosis through NF-κB signaling pathway in
post-laminectomy rats. Neuropharmacology. 79:275–281.
2014.PubMed/NCBI View Article : Google Scholar
|