Introduction
Glioblastoma multiforme (GBM), a grade IV glioma, is
the most common and aggressive type of primary malignant brain
tumor worldwide, and account for ~60 to 70% of malignant gliomas
(1). Despite the progress made in
the last decade, GBM remains one of the most difficult types of
tumors to treat. The median survival rate is only 12-15 months for
patients with glioblastomas (2).
Thus, investigation of the molecular mechanisms associated with
glioma is essential. In recent years, molecular tumour therapy has
improved the treatment of a number of different cancer types
(3,4). Molecular therapy is expected to attack
malignant cells more specifically with fewer side effects than
conventional chemotherapy.
Actin-like 6A (ACTL6A), also known as BAF53a, Arp4
or INO80K, is an actin-like protein and chromatin-remodeling
factor. Specifically, ACTL6A encodes a subunit of the
switch/sucrose non-fermentable complex that has a mainly
transcriptional role as a regulator of the function of stem and
progenitor cells (5,6). Previous studies have demonstrated that
ACTL6A is involved in the differentiation and proliferation of
neural progenitor cells (7,8). In addition, other studies have shown
that ACTL6A plays an important role in the proliferation of
squamous cell carcinoma cells and is associated with
epithelial-mesenchymal transition in hematoma (9,10).
Although there is evidence to suggest that ACTL6A has an oncogenic
role in numerous types of human cancer, the underlying mechanisms
of ACTL6A in glioma cell migration remain unknown (11,12).
Temozolomide (TMZ) is a first-line chemotherapeutic
drug for GBM. Drug resistance is the predominant obstacle in TMZ
therapy. A large number of gliomas are resistant to TMZ, which is
detrimental to the effectiveness of clinical treatment (2). Therefore, it is essential to
investigate TMZ sensitivity in order to identify how the
therapeutic effects of TMZ may be improved. Previous studies have
demonstrated that several genes are involved in TMZ sensitivity,
including autophagy related 4C cysteine peptidase, N-methylpurine
DNA glycosylase, O-6-methylguanine-DNA methyltransferase and STAT3
(13-15).
However, whether ACTL6A affects the sensitivity of glioma cells to
TMZ is unknown and thus is worthy of investigation.
The aim of the present study was to investigate the
potential role of ACTL6A in glioma. In particular, the effect of
ACTL6A on cell migration and the underlying mechanisms were
evaluated following the knockdown of ACTL6A in two glioma cell
lines, T98G and U251. Furthermore, whether the knockdown of ACTL6A
sensitizes these cells to TMZ was also analyzed.
Materials and methods
Cell culture
Human glioma cell lines T98G and U251 were obtained
from the American Type Culture Collection. The glioma cells were
cultured in Dulbecco's modified Eagle's medium (DMEM; Thermo Fisher
Scientific, Inc.) containing 10% fetal bovine serum (FBS; Thermo
Fisher Scientific, Inc.) at 37˚C in a 5% CO2 incubator.
Cells in the logarithmic growth phase were used for subsequent
experiments.
Cell treatment
The ACTL6A-knockdown and negative control (NC)
lentiviruses containing short hairpin (sh)RNA were synthesized by
and purchased from Shanghai GeneChem, Co., Ltd. The
ACTL6A-knockdown and NC lentiviruses were used to transfect T98G
and U251 cells according to the manufacturer's instructions to
establish shACTL6A and sh control (shCtrl) groups. Cells
transfected with an empty vector were used as controls. The shRNA
target sequence for ACTL6A was ACCTTACGTTTCATAGCTTTA and the shRNA
NC sequence was TTCTCCGAACGTGTCACGT. The concentration of shACTL6A
used was 6x108 TU/ml, and the concentration of shCtrl
used was 8x108 TU/ml cells cultured in moderate medium
remained in a humid environment with a temperature of 37˚C and 5%
CO2, after 72 h, the cells were collected and washed
twice with PBS prior to use in further experiments.
Bioinformatics analysis
The Cancer Cell Line Encyclopedia database (CCLE;
https://portals.broadinstitute.org/ccle) was used to
analyze the transcriptomic expression levels of the ACTL6A gene in
multiple glioma cell lines. Gene Expression Profiling Interactive
Analysis (GEPIA; http://gepia.cancer-pku.cn) is an interactive web
server that enables users to perform expression analyses, including
survival and differential analyses, at the subtype level using data
from The Cancer Genome Atlas and Genotype-Tissue Expression portal.
In the present study, GEPIA was used to examine the differential
expression of the ACTL6A gene between different types of cancer,
including glioma, and healthy tissue. A Kaplan-Meier plot was also
created using GEPIA to calculate the overall survival (OS) rates of
patients with glioma, and analyzed using the log-rank test. The Cox
proportional hazard ratio was also calculated. The thresholds for
the high/low ACTL6A expression level cohorts were adjusted
according to the median value.
Patients
A total of 27 patients with glioma were enrolled in
the present study. The clinicopathological features of the patients
are described in Table I. All
patients had received surgery without chemotherapy or radiotherapy
at the First Affiliated Hospital of Nanchang University (Nanchang,
China) between March 2013 and August 2014. Tumor biopsies were
collected prior to neoadjuvant chemotherapy or wide resection of
the tumor and were fresh-frozen and stored at -80˚C. A total of 5
normal control brain tissues were also obtained from patients with
cerebral trauma. The procedures were approved by the Ethics
Committee of the First Affiliated Hospital of Nanchang University
(approval no. 2010-015). Written informed consent was obtained from
every participant.
 | Table IPatient characteristics and clinical
features for the different types of brain tissue. |
Table I
Patient characteristics and clinical
features for the different types of brain tissue.
| | Tissue |
|---|
| Variable | Glioma (n=27) | Normal (n=5) |
|---|
| Mean age (years) | | |
|
Female | 50.83±8.21 | 44.67±7.37 |
|
Male | 45.13±13.53 | 42.50±3.54 |
| Sex (n) | | |
|
Female | 12.00 | 3.00 |
|
Male | 15.00 | 5.00 |
| Tumor grade | | |
|
Low grades
I–II | 9.00 | 0.00 |
|
High grades
III–IV | 16.00 | 0.00 |
Western blot analysis
Total protein was extracted from the cells using
RIPA buffer (cat. no. R0278; Sigma-Aldrich; Merck KGaA) which
contained 2% phosphatase inhibitor and 1% PMSF. A total of 100 µl
protein buffer was harvested from each well of a 6-well plate. The
protein concentration was quantified by the BCA method and the mass
of protein loaded per lane was ~30 µg. Protein samples were
separated by 10% SDS-PAGE and transferred to PVDF membranes. The
membranes were blocked in 5% skimmed milk with Tris-buffered saline
with 10% Tween-20 (TBST) for 1 h at room temperature, and then
incubated with primary antibodies overnight at 4˚C. The next day,
the membranes were incubated with secondary antibodies for 1 h at
room temperature. After extensive washing using TBST, the blots
were visualized using Amersham Hyperfilm ECL (Cytiva). The
following primary antibodies were used: Anti-ACTL6A (cat. no.
abs133368; absin; dilution 1:3,000), anti-AKT (cat. no. 10176-2-AP;
ProteinTech Group, Inc.; dilution 1:2,000),
anti-Ser473-phosphorylated (p)-AKT (cat. no. 4060T; Cell Signaling
Technology, Inc.; dilution 1:2,000), anti-E-cadherin (cat. no.
3195T; Cell Signaling Technology, Inc,.; dilution 1:6,000),
anti-vimentin (cat. no. 5741T; Cell Signaling Technology, Inc.;
dilution 1:3,000), anti-PI3K (cat. no. abs123958; Cell Signaling
Technology, Inc.; dilution 1:5,000), anti-p-PI3K (cat. no. 17366S;
absin; dilution 1:1,000), and anti-β-actin (abs118937, absin,
dilution 1:5,000). Horseradish peroxidase-conjugated anti-human IgG
was used as the secondary antibody (cat. no. 7074P2; Cell Signaling
Technology, Inc.; dilution 1:10,000). Image Lab V3.0 (Bio-Rad
Laboratories, Inc.) was used for densitometry.
Reverse transcription-quantitative PCR
(RT-qPCR)
The total RNA was extracted from patient tissue
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). A total of 2 µg RNA was converted into cDNA
using a Thermo Scientific RevertAid First Strand cDNA Synthesis kit
(Thermo Fisher Scientific, Inc.). qPCR was then performed using
Maxima SYBR Green qPCR Master Mix (DBI Bioscience) in an ABI PRISM
7500 Fast Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The PCR conditions were as follows: 95˚C for 5
min, and then 40 cycles of amplification for 30 sec at 95˚C, 45 sec
at 60˚C and 45 sec at 72˚C. The primers, purchased from Guangzhou
RiboBio Co. Ltd., were as follows: ACTL6A, forward:
TCAGAGGCACCGTGGAATAC; reverse: GACATAGCCATCGTGGACTG; GAPDH,
forward: TGACTTCAACAGCGACACCCA; reverse: CACCCTGTTGCTGTAGCCAAA. The
gene expression was quantified using the 2-ΔΔCq method
(16).
Wound-healing assay
The transfected cells were seeded in 6-well plates
containing DMEM with 10% FBS. When cultured to 90% confluence, the
cells were starved for 24 h in serum-free medium. A sterile 10-µl
pipette tip was used to create wounds in the cell layer. After
washing away the dislodged cell fragments using PBS, the cells were
cultured in 2% FBS medium. The areas of the wounds were assessed
under an Zeiss AG fluorescence inverted microscope at x5
magnification after 24 h. ImageJ 1.8.0 was then used to quantify
the results (National Institutes of Health).
Single cell tracking assay
The Operetta® High Content Imaging System
(PerkinElmer, Inc.) combines fully automated high-throughput
fluorescence microscopy with multi-parameter quantitative image
analysis, which enables the effects of drugs on cell properties,
such as migration, to be evaluated (10,11).
The transfected cells were seeded at a density of 2,000 cells/well
in a 96-well plate and cultured in an Operetta High Content Imaging
System at 37˚C with 5% CO2 for 20 h. Digital phase
contrast images were acquired every 1 h for 20 h, and the mean
square displacements of cells in the two groups were compared using
Harmony® 3.5 High Content Imaging and Analysis Software
(PerkinElmer, Inc.).
MTT assay
U251 and T98G cells were seeded in 96-well plates at
a density of 5x103 cells/well and cultured in 5%
CO2 at 37˚C. TMZ (Selleck Chemicals) and SC79 were
dissolved in dimethyl sulfoxide (Sigma Aldrich; Merck KGaA). The
cells were cultured for 12 h prior to the knockdown of ACTL6A,
after 48 h transfection, cell were treated with TMZ (0, 100, 500
and 1,000 µM) for 12 h. Similarly, after 48 h of transfection,
cells were treated with 5 µM of SC79 for 12 h. A total of 100 µl
culture medium (DMEM; Thermo Fisher Scientific, Inc.) containing 20
µl MTT was added into each well and the 96-well plates were placed
in an incubator at 37˚C for 4 h. After 4 h, 150 µl/well formanzan
solution (Nanjing KeyGen Biotech Co., Ltd.) was used to dissolve
the blue-purple crystals. The OD value of each well was detected at
the absorbance of 490 nm.
Statistical analysis
Each experiment was repeated three times. Data are
presented as the mean ± SD unless otherwise indicated and were
compared using Student's t-test or one-way analysis of variance
(ANOVA) followed by Bonferroni post hoc tests. The mean square
displacement data were analyzed using two-way ANOVA followed by
Bonferroni post hoc tests. All analyses were performed using
GraphPad Prism (version 7.00; GraphPad Software, Inc.). P<0.05
was considered to indicate a statistically significant
difference.
Results
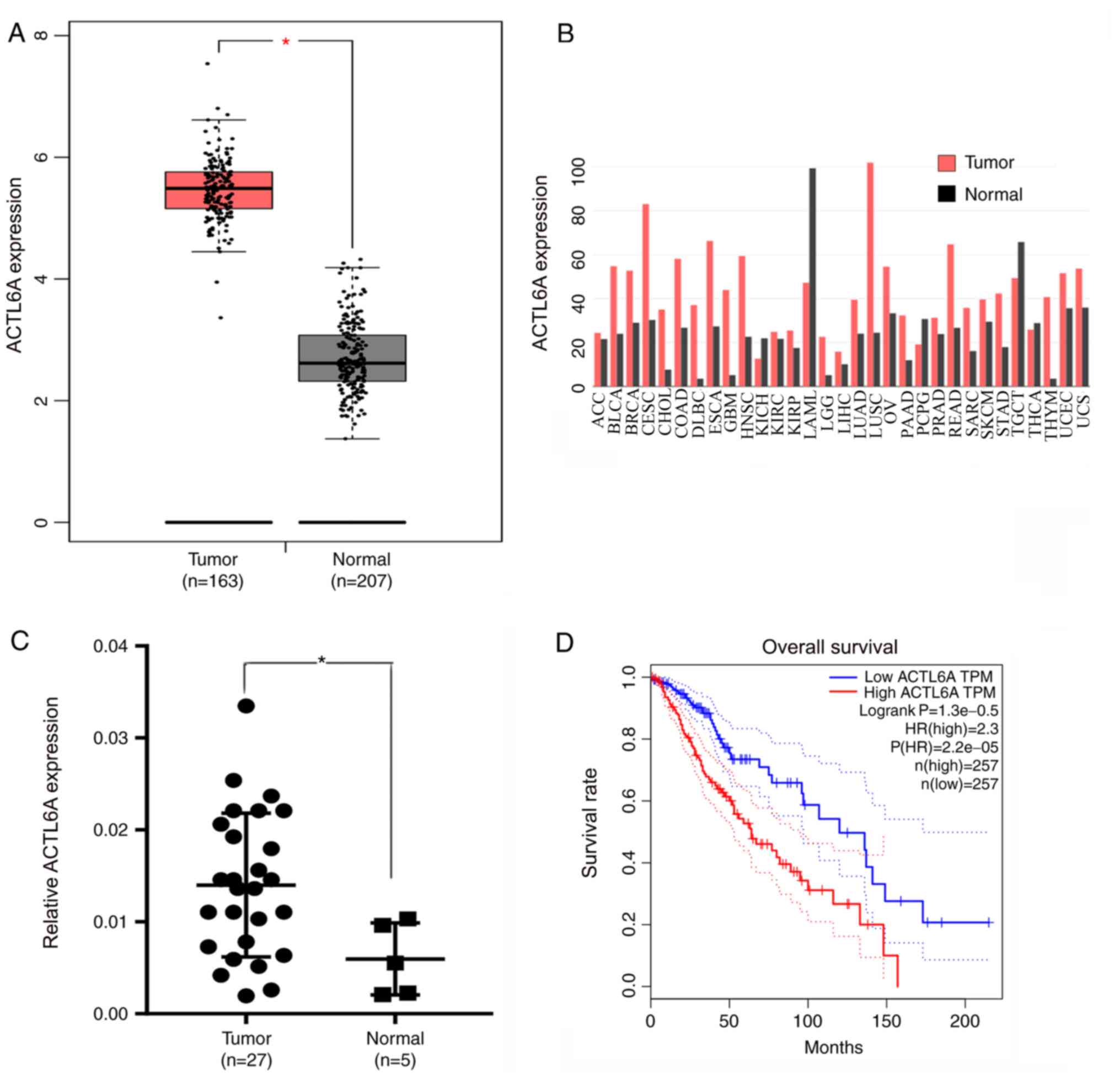
ACTL6A expression is upregulated in
glioma
The differences in ACTL6A expression between glioma
and normal tissue were analyzed using the GEPIA database. The
results revealed a significant difference in the expression levels
of ACTL6A between glioma and normal tissue; the ACTL6A expression
levels in glioma were significantly higher compared with those in
normal tissue (Fig. 1A). The
expression levels of ACTL6A in different types of cancer were also
analyzed using GEPIA. The results reveal that compared with normal
samples, ACTL6A is highly expressed in multiple cancer types,
including in glioma, bladder urothelial carcinoma and breast
carcinoma. However, ACTL6A expression is low in some cancers, such
as in acute myeloid leukemia and testicular germ cell tumors
(Fig. 1B). A total of 27 glioma
samples and 5 normal control brain tissue samples were obtained
from patients at the First Affiliated Hospital of Nanchang
University, and RT-qPCR was used to measure the expression levels
of ACTL6A. The results revealed that ACTL6A was expressed at
significantly higher levels in glioma compared with normal brain
tissue (Fig. 1C). The OS curves for
glioma generated using GEPIA indicated that high expression of
ACTL6A was associated with a poor prognosis, while low expression
of ACTL6A was associated with an improved prognosis (Fig. 1D). These results indicate that
ACTL6A expression is upregulated in glioma tissue, and that ACTL6A
may play a significant role in the development and progression of
glioma.
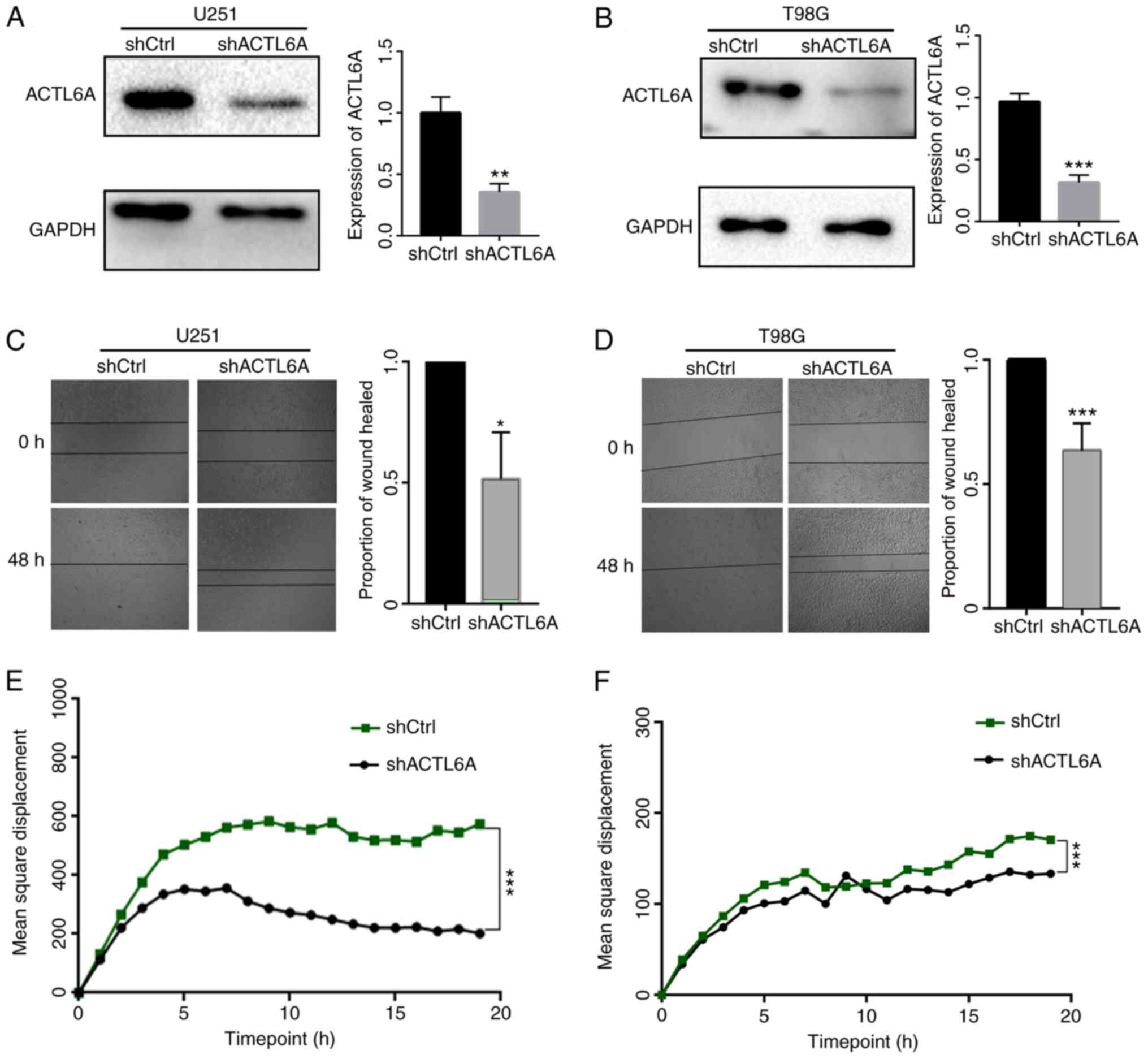
ACTL6A promotes glioma cell migration
in vitro
ACTL6A knockdown U251 and T98G cell lines were
constructed to investigate the functional role of ACTL6A in glioma
cells in vitro. First, the efficiency of ACTL6A knockdown in
U251 and T98G cells was determined by comparison with NC
transfected controls, and the results revealed that transfection
with ACTL6A shRNA significantly reduced the expression of ACTL6A
(Fig. 2A and B). Next, a wound-healing assay was
performed to detect the migration ability of the glioma cells. The
results revealed that the knockdown of ACTL6A significantly reduced
the wound-healing rate compared with that of the shCtrl group
(Fig. 2C and D). These results indicate that the
knockdown of ACTL6A inhibited the migration capacity of the glioma
cells. In addition, the 20-h mean square displacements of the
glioma cells were analyzed. The results confirm that the cells in
which ACTL6A was knocked down migrated more slowly compared with
those in the shCtrl group (Fig. 2E
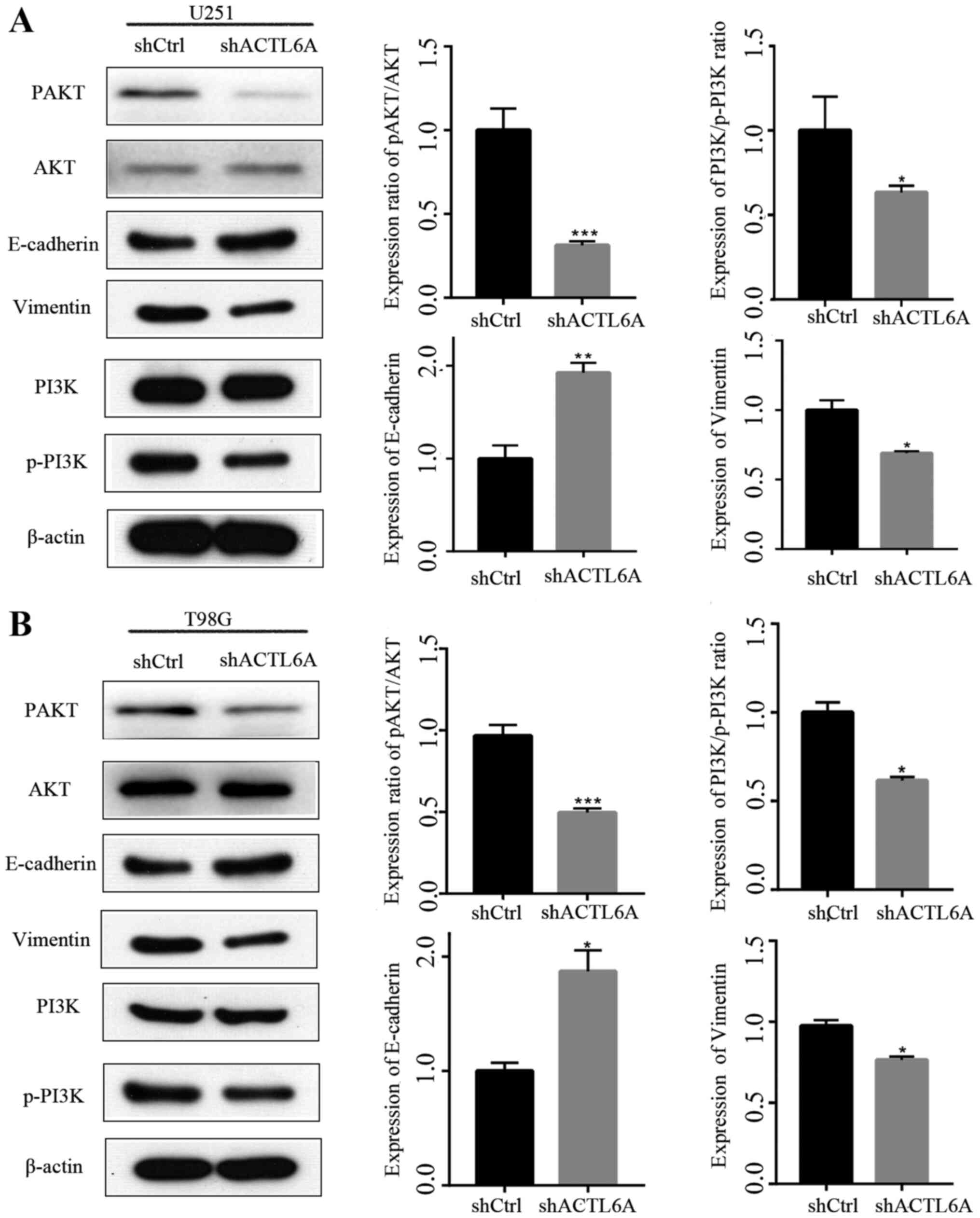
and F). The expression levels of
proteins associated with migration, namely E-cadherin and vimentin,
were also detected and the results revealed that the expression of
E-cadherin was upregulated while that of vimentin was downregulated
in the ACTL6A knockdown groups compared with the respective shCtrl
groups, which further indicate that ACTL6A knockdown inhibited cell
migration (Fig. 3). These results
suggest that ACTL6A promotes the migration capacity of glioma
cells.
Knockdown of ACTL6A inhibits the
PI3K/AKT signaling pathway
Numerous studies have confirmed that the PI3K/AKT
signaling pathway plays an important role in the regulation of cell
proliferation (17,18). However, whether ACTL6A affects the
PI3K/AKT pathway remains unknown. Western blotting analysis
revealed that the knockdown of ACTL6A significantly decreased
p-AKT/AKT and p-PI3K/PI3K ratios compared with those in the shCtrl
group, while the levels of total AKT and total PI3K were not
changed (Fig. 3). These results
suggest that ACTL6A may regulate biological functions in glioma via
the PI3K/AKT signaling pathway.
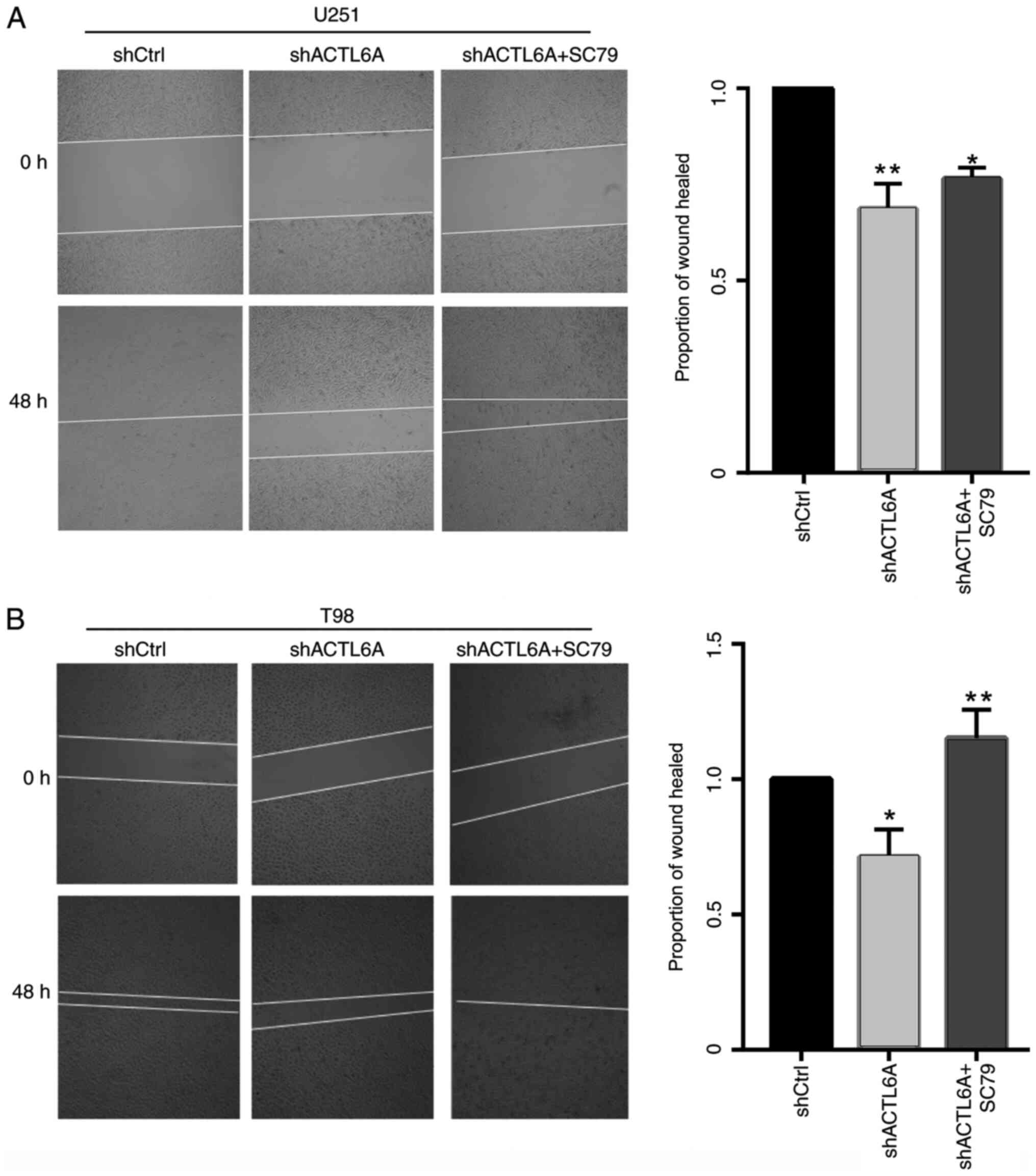
PI3K/AKT agonist reverses the effect
of ACTL6A knockdown
In order to further investigate the role and
mechanism of ACTL6A in cell migration, U251 and T98G cells were
treated with the AKT pathway agonist SC79 or DMSO solvent after
transfection. Following treatment, the wound-healing assay was used
to detect cell migration, and the results showed that the migration
ability of the U251 and T98G cells, which was significantly
decreased following transfection with sh-ACTL6A was recovered by
SC79 (Fig. 4). These results
supports the hypothesis that silencing ACTL6A inhibits cell
migration through deactivation of the AKT pathway.
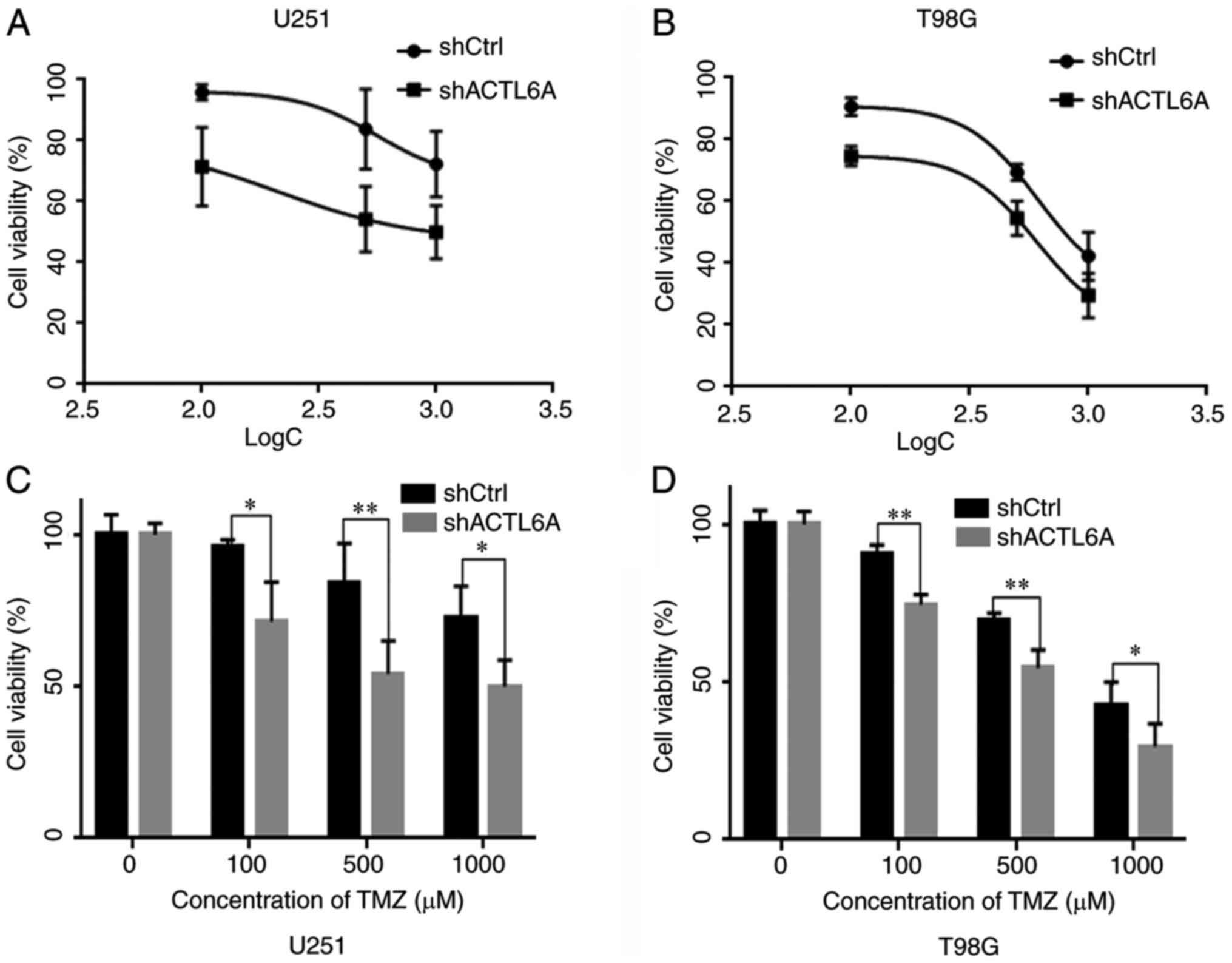
Knockdown of ACTL6A contributes to the
sensitivity of glioma cells to TMZ
In order to determine whether ACTL6A affects the
sensitivity of U251 and T98G cells to TMZ-induced cell death, the
viability of U251 and T98G cells following treatment with TMZ was
tested using an MTT assay. The results revealed that as the
concentration of TMZ increased, cell viability decreased. The cell
viability of the ACTL6A group was significantly lower than that of
the shCtrl group at all concentrations tested, suggesting that the
sensitivity to TMZ was greater in the shACTL6A group compared with
the shCtrl group (Fig. 5).
Discussion
Glioma-related literature and cell databases were
consulted to assist the selection of cell lines for inclusion in
the present study. The CCLE database revealed that the expression
of ACTL6A was similar in different glioma cell lines. The current
study observed that the long synapse of the U87 cell line renders
it unsuitable for cell scratch experiments, and the proliferation
rate of HS683 cells is rapid, which would adversely affect the cell
migration experiment. Therefore, U251 cells and T98G cell lines
were selected for investigation in the present study. The results
demonstrated that ACTL6A is highly expressed in glioma cells, and
its expression is associated with tumor migration. In vitro
assays indicated that the knockdown of ACTL6A inhibited cell
migration via reduction of the phosphorylation of AKT at Ser473,
and the decrease in cell migration induced by ACTL6A knockdown was
reversed by SC79, an activator of the AKT pathway. Furthermore, MTT
assay results revealed that ACTL6A knockdown enhanced the
sensitivity of the glioma cells to TMZ.
Previous studies have demonstrated that ACTL6A is
involved in the migration of certain types of cancer cells, such as
epidermal squamous cell carcinoma and colorectal carcinoma cells
(19,20). Furthermore, ACTL6A has been shown to
be upregulated in glioma cells and associated with the progression
of glioma (11). The results of
present study are consistent with these previous findings. They
revealed that the knockdown of ACTL6A inhibited the migration
ability of glioma cells, and demonstrated that the ability to
migrate was restored following treatment with an AKT agonist.
AKT is a major cellular kinase involved in cell
proliferation, apoptosis, invasion and migration (21). Numerous studies have shown that AKT
activation increases the migratory or invasive activity of various
types of cells (22-25).
The AKT pathway is regulated by multiple genes, including
insulin-like growth factor 1 receptor and mTOR, and contributes to
wound healing (26-28).
SC79, an AKT pathway agonist, reversed the wound healing by
activating the AKT pathway in glioma (29,30).
The results of the present study suggest that ACTL6A induces the
phosphorylation of AKT at Ser473, and stimulates the migration of
human glioma cells, at least in part through activation of the AKT
signaling pathway.
TMZ is an imidazotetrazine derivative and a prodrug,
the active metabolite of which can also be derived from the
alkylating agent dacarbazine. TMZ is often used as a standard
treatment for glioma. It causes the death of cancer cells via its
ability to methylate or alkylate DNA at the O6 or
N7 sites of guanine residues (31,32).
However, tumor cells can acquire the ability to repair DNA damage,
which may greatly reduce the efficacy of TMZ. Therefore, to improve
the effectiveness of TMZ, it is necessary to further investigate
the mechanism by which TMZ inhibits tumor progression. The present
study revealed that ACTL6A knockdown increased the sensitivity of
glioma cells to TMZ. Future studies are required to assess whether
the enhancement of TMZ sensitivity by ACTL6A knockdown is
associated with DNA damage repair or other mechanisms.
In conclusion, to the best of our knowledge, the
present study revealed for the first time that the knockdown of
ACTL6A reduced the migration of human glioma cells, at least in
part through inactivation of the AKT signaling pathway, and
enhanced the sensitivity of glioma cells to TMZ. The results of the
present study may provide a new therapeutic strategy for the
treatment of GBM.
Acknowledgements
Not applicable.
Funding
The study was funded by the Hunan Provincial Natural
Science Foundation of China (grant no. 2018JJ2600) and the Project
of Scientific Research Plan of Hunan Provincial Health Commission
(grant no. B2017029).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
XP designed the current study and collected data
from recruited cases. XC performed the majority of the experiments.
ZX analyzed data and performed the statistics. XZ and XC analyzed
and wrote the manuscript. DL completed the experiments that were
suggested by reviewers. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All patients provided written informed consent for
their tissues to be used for clinical research. The present study
was approved by the Independent Ethics Committee of the Institute
of Clinical Pharmacology, Central South University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lapointe S, Perry A and Butowski NA:
Primary brain tumours in adults. Lancet. 392:432–446.
2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Posadas EM, Limvorasak S and Figlin RA:
Targeted therapies for renal cell carcinoma. Nat Rev Nephrol.
13:496–511. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Lee YT, Tan YJ and Oon CE: Molecular
targeted therapy: Treating cancer with specificity. Eur J
Pharmacol. 834:188–196. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Marom R, Jain M, Burrage LC, Song IW,
Graham BH, Brown CW, Stevens SJC, Stegmann APA, Gunter AT, Kaplan
JD, et al: Heterozygous variants in ACTL6A, encoding a component of
the BAF complex, are associated with intellectual disability. Hum
Mutat. 38:1365–1371. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Sima X, He J, Peng J, Xu Y, Zhang F and
Deng L: The genetic alteration spectrum of the SWI/SNF complex: The
oncogenic roles of BRD9 and ACTL6A. PLoS One.
14(e0222305)2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Yoo AS, Staahl BT, Chen L and Crabtree GR:
MicroRNA-mediated switching of chromatin-remodelling complexes in
neural development. Nature. 460:642–646. 2009.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Son EY and Crabtree GR: The role of BAF
(mSWI/SNF) complexes in mammalian neural development. Am J Med
Genet C Semin Med Genet. 166C:333–349. 2014.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zeng Z, Yang H and Xiao S: ACTL6A
expression promotes invasion, metastasis and epithelial mesenchymal
transition of colon cancer. BMC Cancer. 18(1020)2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Xiao S, Chang RM, Yang MY, Lei X, Liu X,
Gao WB, Xiao JL and Yang LY: Actin-like 6A predicts poor prognosis
of hepatocellular carcinoma and promotes metastasis and
epithelial-mesenchymal transition. Hepatology. 63:1256–1271.
2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Meng L, Wang X, Liao W, Liu J, Liao Y and
He Q: BAF53a is a potential prognostic biomarker and promotes
invasion and epithelial-mesenchymal transition of glioma cells.
Oncol Rep. 38:3327–3334. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ji J, Xu R, Zhang X, Han M, Xu Y, Wei Y,
Ding K, Wang S, Bin Huang, Chen A, et al: Actin like-6A promotes
glioma progression through stabilization of transcriptional
regulators YAP/TAZ. Cell Death Dis. 9(517)2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Lee ES, Ko KK, Joe YA, Kang SG and Hong
YK: Inhibition of STAT3 reverses drug resistance acquired in
temozolomide-resistant human glioma cells. Oncol Lett. 2:115–121.
2011.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Pan Q, Yang XJ, Wang HM, Dong XT, Wang W,
Li Y and Li JM: Chemoresistance to temozolomide in human glioma
cell line U251 is associated with increased activity of
O6-methylguanine-DNA methyltransferase and can be overcome by
metronomic temozolomide regimen. Cell Biochem Biophys. 62:185–191.
2012.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wen ZP, Zeng WJ, Chen YH, Li H, Wang JY,
Cheng Q, Yu J, Zhou HH, Liu ZZ, Xiao J and Chen XP: Knockdown ATG4C
inhibits gliomas progression and promotes temozolomide
chemosensitivity by suppressing autophagic flux. J Exp Clin Cancer
Res. 38(298)2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Wang S, Zheng Y, He Z, Zhou W, Cheng Y and
Zhang C: SH2B1 promotes NSCLC cell proliferation through
PI3K/Akt/mTOR signaling cascade. Cancer Cell Int.
18(132)2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Xu XY, Zhang J, Qi YH, Kong M, Liu SA and
Hu JJ: Linc-ROR promotes endometrial cell proliferation by
activating the PI3K-Akt pathway. Eur Rev Med Pharmacol Sci.
22:2218–2225. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zhu B, Ueda A, Song X, Horike SI, Yokota T
and Akagi T: Baf53a is involved in survival of mouse ES cells,
which can be compensated by Baf53b. Sci Rep.
7(14059)2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Sun W, Wang W, Lei J, Li H and Wu Y:
Actin-like protein 6A is a novel prognostic indicator promoting
invasion and metastasis in osteosarcoma. Oncol Rep. 37:2405–2417.
2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Chu N, Salguero AL, Liu AZ, Chen Z,
Dempsey DR, Ficarro SB, Alexander WM, Marto JA, Li Y, Amzel LM, et
al: Akt Kinase activation mechanisms revealed using protein
semisynthesis. Cell. 174:897–907.e14. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kwiatkowska A and Symons M: Signaling
determinants of glioma cell invasion. Adv Exp Med Biol.
986:121–141. 2013.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Ao R, Guan L, Wang Y and Wang JN:
Silencing of COL1A2, COL6A3, and THBS2 inhibits gastric cancer cell
proliferation, migration, and invasion while promoting apoptosis
through the PI3k-Akt signaling pathway. J Cell Biochem.
119:4420–4434. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Luo KW, Lung WY, Chun-Xie Luo XL and Huang
WR: EGCG inhibited bladder cancer T24 and 5637 cell proliferation
and migration via PI3K/AKT pathway. Oncotarget. 9:12261–12272.
2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Deng X, Tu Z, Xiong M, Tembo K, Zhou L,
Liu P, Pan S, Xiong J, Yang X, Leng J, et al: Wnt5a and CCL25
promote adult T-cell acute lymphoblastic leukemia cell migration,
invasion and metastasis. Oncotarget. 8:39033–39047. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Ilowski M, Putz C, Weiss TS, Brand S,
Jauch KW, Hengstler JG and Thasler WE: Augmenter of liver
regeneration causes different kinetics of ERK1/2 and Akt/PKB
phosphorylation than EGF and induces hepatocyte proliferation in an
EGF receptor independent and liver specific manner. Biochem Biophys
Res Commun. 394:915–920. 2010.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Guo YY, Wu Y, Jia XW and An W: Augmenter
of liver regeneration potentiates doxorubicin anticancer efficacy
by reducing the expression of ABCB1 and ABCG2 in hepatocellular
carcinoma. Lab Invest. 97:1400–1411. 2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Shang X, Lin K, Yu R, Zhu P, Zhang Y, Wang
L, Xu J and Chen K: Resveratrol protects the myocardium in sepsis
by activating the phosphatidylinositol 3-Kinases
(PI3K)/AKT/mammalian target of rapamycin (mTOR) pathway and
inhibiting the nuclear factor-kappaB (NF-kappaB) signaling pathway.
Med Sci Monit. 25:9290–9298. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Liu W, Jing ZT, Wu SX, He Y, Lin YT, Chen
WN, Lin XJ and Lin X: A Novel AKT Activator, SC79, prevents acute
hepatic failure induced by fas-mediated apoptosis of hepatocytes.
Am J Pathol. 188:1171–1182. 2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Xu Y, Gao YW and Yang Y: SC79 protects
dopaminergic neurons from oxidative stress. Oncotarget.
9:12639–12648. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Hegi ME, Liu L, Herman JG, Stupp R, Wick
W, Weller M, Mehta MP and Gilbert MR: Correlation of
O6-methylguanine methyltransferase (MGMT) promoter methylation with
clinical outcomes in glioblastoma and clinical strategies to
modulate MGMT activity. J Clin Oncol. 26:4189–4199. 2008.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Preusser M, Charles JR, Felsberg J,
Reifenberger G, Hamou MF, Diserens AC, Stupp R, Gorlia T, Marosi C,
Heinzl H, et al: Anti-O6-methylguanine-methyltransferase (MGMT)
immunohistochemistry in glioblastoma multiforme: Observer
variability and lack of association with patient survival impede
its use as clinical biomarker. Brain Pathol. 18:520–532.
2008.PubMed/NCBI View Article : Google Scholar
|