Introduction
Esophageal cancer is the fourth most common cancer
type in China and nearly half of all newly diagnosed cases of
esophageal cancer and associated deaths worldwide occur in China
(1,2). The most common histological subtypes
are esophageal squamous cell carcinoma (ESCC) and adenocarcinoma;
however, in China, >90% of esophageal cancer cases are squamous
cell carcinoma (3).
Long non-coding RNAs (lncRNAs) are a class of RNA
molecules that are >200 nucleotides long and do not have any
protein-coding capacity (4,5). In recent years, an increasing number
of studies suggested that lncRNAs have important roles in
regulating biological behaviours, including cell proliferation,
differentiation, migration and invasion (6-8).
They are able to affect biological activities by regulating
mechanisms associated with gene expression, chromatin modification
and cell metabolism (9,10). lncRNAs are also closely associated
with tumour development and influence the prognosis of patients
with tumours (11,12). Abnormal expression of lncRNAs has
been reported in ESCC (13-15).
Studies on ESCC have indicated that lncRNAs may have
a role in suppressing or promoting cancer-associated genes during
the development of esophageal cancer. Previous studies have also
indicated that certain lncRNAs may be potential markers for ESCC
tumour diagnosis, treatment and estimation of prognosis (16,17).
It was also reported that the expression level of lncRNA lymphoid
enhancer binding factor 1 (LEF1)-antisense 1 (AS1) in ESCC tissues
is significantly increased and that LEF1-AS1 is associated with
lymph node metastasis, TNM stage and poor prognosis in patients
with ESCC (16). It has also been
suggested that lnc-ABC A12-3 expression is increased in ESCC and
that this lncRNA is able to promote the migration, invasion and
proliferation of tumour cells by regulating fibronectin 1(17). lncRNA cancer susceptibility 2 is
able to negatively regulate microRNA-181a by inhibiting the Akt
pathway in ESCC cells, thereby increasing the sensitivity of ESCC
cells to cisplatin (18).
c-Myc upregulated lncRNA (MYU), also known as VPS9
domain-containing 1 (VPS9D1)-AS1, is a recently discovered lncRNA
that was produced from the reverse strand of the VPS9D1 gene. To
date, the expression and clinical significance of lncRNA MYU in
ESCC has not been systematically reported in the literature. Thus,
the present study explored the expression of lncRNA MYU in ESCC and
its role in tumour development and progression. The effect of
lncRNA MYU expression on the growth, proliferation and migration of
various gastric cancer cell lines was also investigated.
Materials and methods
Patients and tissue specimens
Tissue specimens were collected from 112 patients
with ESCC who underwent radical resection between January 2015 and
December 2017 at the Affiliated Hospital of Nantong University
(Nantong, China). None of the patients recruited for the study
received any radiotherapy or chemotherapy prior to surgery. The
ages of the patients ranged from 33 to 78 years, with a median age
of 68 years, and there were 71 males and 41 females. The detailed
clinicopathological parameters of the patients are provided in
Table I. Tumor tissue samples were
collected with written informed consent from the patients. The
present study was approved by the Ethics Committee of the
Affiliated Hospital of Nantong University (Nantong, China).
 | Table IlncRNA MYU expression in esophageal
squamous cell carcinoma. |
Table I
lncRNA MYU expression in esophageal
squamous cell carcinoma.
| | lncRNA MYU | |
|---|
| Clinicopathological
features | Number of
patients | High expression | Low expression | χ2
value | P-value |
|---|
| Age (years) | | | | 0.169 | 0.681 |
|
<60 | 34 | 18 | 16 | | |
|
≥60 | 78 | 38 | 40 | | |
| Sex | | | | 1.885 | 0.170 |
|
Male | 71 | 32 | 39 | | |
|
Female | 41 | 24 | 17 | | |
| Histological
grade | | | | 4.390 | 0.036 |
|
High +
moderate | 63 | 26 | 37 | | |
|
Low | 49 | 30 | 19 | | |
| Vascular tumour
thrombus | | | | 0.728 | 0.393 |
|
Present | 30 | 17 | 13 | | |
|
Absent | 82 | 39 | 43 | | |
| T
classification | | | | 5.659 | 0.059 |
|
T1 | 29 | 9 | 20 | | |
|
T2 | 36 | 20 | 16 | | |
|
T3 | 47 | 27 | 20 | | |
| Lymph node
metastasis | | | | 6.196 | 0.013 |
|
Present | 47 | 30 | 17 | | |
|
Absent | 65 | 26 | 39 | | |
| TNM stage | | | | 5.486 | 0.019 |
|
I/II | 70 | 29 | 41 | | |
|
III/IV | 42 | 27 | 15 | | |
A total of four ESCC cell lines (EC9706, KYSE150,
ECA109 and TE-2 cells) and normal human esophageal epithelial cells
(HEECs) were obtained from the Shanghai Institute of Cell Research,
Chinese Academy of Sciences. All cells were cultured in RPMI-1640
(Gibco; Thermo Fisher Scientific, Inc.) medium supplemented with
10% foetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and
maintained under standard conditions (37˚C, 5% CO2).
Antibodies and reagents
TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), the RevertAid First Strand cDNA Synthesis Kit
(Thermo Fisher Scientific, Inc.), a green fluorescent protein
(GFP)-tagged MYU-specific small interfering RNA (siRNA) plasmid
(Shanghai Genechem Co., Ltd.), a Ki-67 antibody (cat. no.
MA5-15690), an E-cadherin antibody (cat. no. MA5-15711) and a
Vimentin antibody (cat. no. MA5-35320; all from Invitrogen; Thermo
Fisher Scientific, Inc.), and Lipofectamine™ 2000 (Santa Cruz
Biotechnology) were used in the present study.
Immunohistochemistry
Tissue specimens were fixed with 4% formaldehyde,
routinely embedded in paraffin, sectioned, dewaxed and hydrated.
Subsequently, endogenous peroxidase activity was blocked. The
samples were then incubated with primary antibodies [Ki-67 (1:500
dilution), E-cadherin (1:200; dilution) and Vimentin (1:200
dilution)] overnight at 4˚C, followed by incubation with the
corresponding secondary antibodies: Goat anti-rabbit (1:2,000; cat.
no. ab207995; Abcam) and goat anti-mouse (1:1,000, cat. no. ab6788;
Abcam) at room temperature for 30 min. The antibodies were
visualized with diaminobenzidine and finally, the samples were
counterstained with haematoxylin.
The immunostaining intensity was evaluated according
to immunoreactivity scores, which were determined based on staining
intensity and the percentage of positive cells. Five high-power
visual fields were observed in each tissue, and 1x103
cells were counted for statistical analysis. The staining intensity
score was multiplied by the score indicating the percentage of
positive cells. The staining intensity of tumour cells was scored
using the following criteria: Negative, 0; weak, 1; moderate, 2;
and strong, 3. The scoring criteria for the percentage of positive
cells were as follows: <5%, 0; 5-25%, 1; 26-50%, 2; 51-75%, 3;
and >75%, 4.
Cell transfection
A lncRNA MYU siRNA and control siRNA were chemically
synthesized by Shanghai GenePharma, Co., Ltd. The sequences of the
lncRNA MYU-specific siRNAs were as follows:
5'-CCCUGCAAGCCAUGGGUAA-3' (siRNA-1) and 5'-CAAGAAGGCUGGUCACAGU-3'
(siRNA-2). The sequence of the lncRNA MYU control siRNA was as
follows: 5'-AAUUCUCCGAACGUGUCACGU-3' (siRNA control). Plasmid
transfection was performed using Lipofectamine™ 2000 in accordance
with the manufacturer's instructions. ESCC TE-2 cells were seeded
into 24-well plates (2x105 cells/well) and transfected
with the lncRNA MYU siRNA or control siRNA at 37˚C for 20 min.
After incubation for 48 h, ESCC TE-2 cells were collected for the
subsequent experiments.
RNA extraction and reverse
transcription-quantitative (RT-qPCR)
Total RNA was extracted from tissues and cultured
cells with TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. RT was conducted
using the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher
Scientific, Inc.). The specific primer sequences for fluorescence
qPCR were as follows: MYU forward, 5'-ATGGGTAACCAGGGGTCAAG-3' and
reverse, 5'-AGTAACAGTGGTAGAGCCGAC-3'; E-cadherin forward,
5'-CGAGAGCTACACGTTCACGG-3' and reverse,
5'-GGGTGTCGAGGGAAAAATAGG-3'; Vimentin forward,
5'-GACGCCATCAACACCGAGTT-3' and reverse,
5'-CTTTGTCGTTGGTTAGCTGGT-3'; GAPDH forward,
5'-AATGGAGCCGTTAGGAAA-3' and reverse, 5'-GCGCCATACGACAATC-3'. The
thermocycling conditions were as follows: Initial step at 94˚C for
40 sec, followed by 30 cycles of 94˚C for 10 sec and 65˚C for 30
sec. An ABI 7500 qPCR instrument (Applied Biosystems; Thermo Fisher
Scientific, Inc.) was used for relative quantitative analysis.
GAPDH was used as the internal reference gene and the relative
expression level of lncRNA MYU was calculated. Each sample was
tested at least three times.
Cell Counting Kit-8 (CCK-8) assay
To analyse the growth of the ESCC cell lines, the
CCK-8 assay (Dojindo Laboratories) was performed according to the
manufacturer's protocol. ESCC cells transfected with lncRNA MYU
siRNA or control siRNA were cultured in 96-well plates overnight.
The cells were incubated for another 2 h and then detected at a
wavelength of 450 nm using a microplate reader (Bio-Rad
Laboratories). GraphPad Prism 8 software (GraphPad Software, Inc.)
was used to plot the cell growth curve. The experiments were
performed at least three times.
Cell cycle analysis
BD cell cycle reagent (BD Biosciences) was used
according to the manufacturer's protocol. ESCC cells were washed
with cold PBS and then fixed in 70% ethanol. After fixation, the
cells were stained with propidium iodide. Finally, the cell
suspensions were detected and analysed with a flow cytometer (BD
Biosciences) and evaluated with ModFit LT 4.1 software (Verity
Software House).
Wound-healing assay
ESCC cells transfected with lncRNA MYU siRNA or
control siRNA were seeded in 6-well plates. After the cells reached
confluence in a single layer, the scratch test was performed. A
line-shaped scratch was performed, and the cells were rinsed with
phosphate buffer to remove the detached cells, and then fresh
medium without serum was added. Images of the scraped area with
migrated cells were captured at 0 and 48 h after scratching.
Transwell assay
Serum-deprived ESCC cells were inoculated into the
upper chamber of a Transwell chamber (Corning Inc.) according to
the manufacturer's protocol and culture medium was added to the
lower chamber as a chemoattractant while the serum was not
contained in the upper chamber. A total of 1x105
cells/well were seeded in serum-free medium in the upper chamber
with a membrane with 8-µm width filter pore that was pre-coated
with Matrigel (BD Biosciences). After incubation for 48 h at 37˚C,
the medium was aspirated and the cells that had invaded to the
lower chamber were fixed in 4% formaldehyde for 30 min at room
temperature. Subsequently, the cells were stained with 0.1% crystal
violet for 10 min at room temperature and the cells on the lower
surface of the membrane were counted under a microscope.
Statistical analysis
Each biological experiment was performed three
times. Statistical significance was determined by Student's t-test,
the χ2 test or one-way ANOVA with Tukey's post-hoc test
as appropriate. Survival analysis was performed using the
Kaplan-Meier method and the log-rank test. All statistical analyses
were performed using SPSS 25.0 software (IBM Corp.). P<0.05 was
considered to indicate statistical significance.
Results
lncRNA MYU is increased in ESCC
tissues
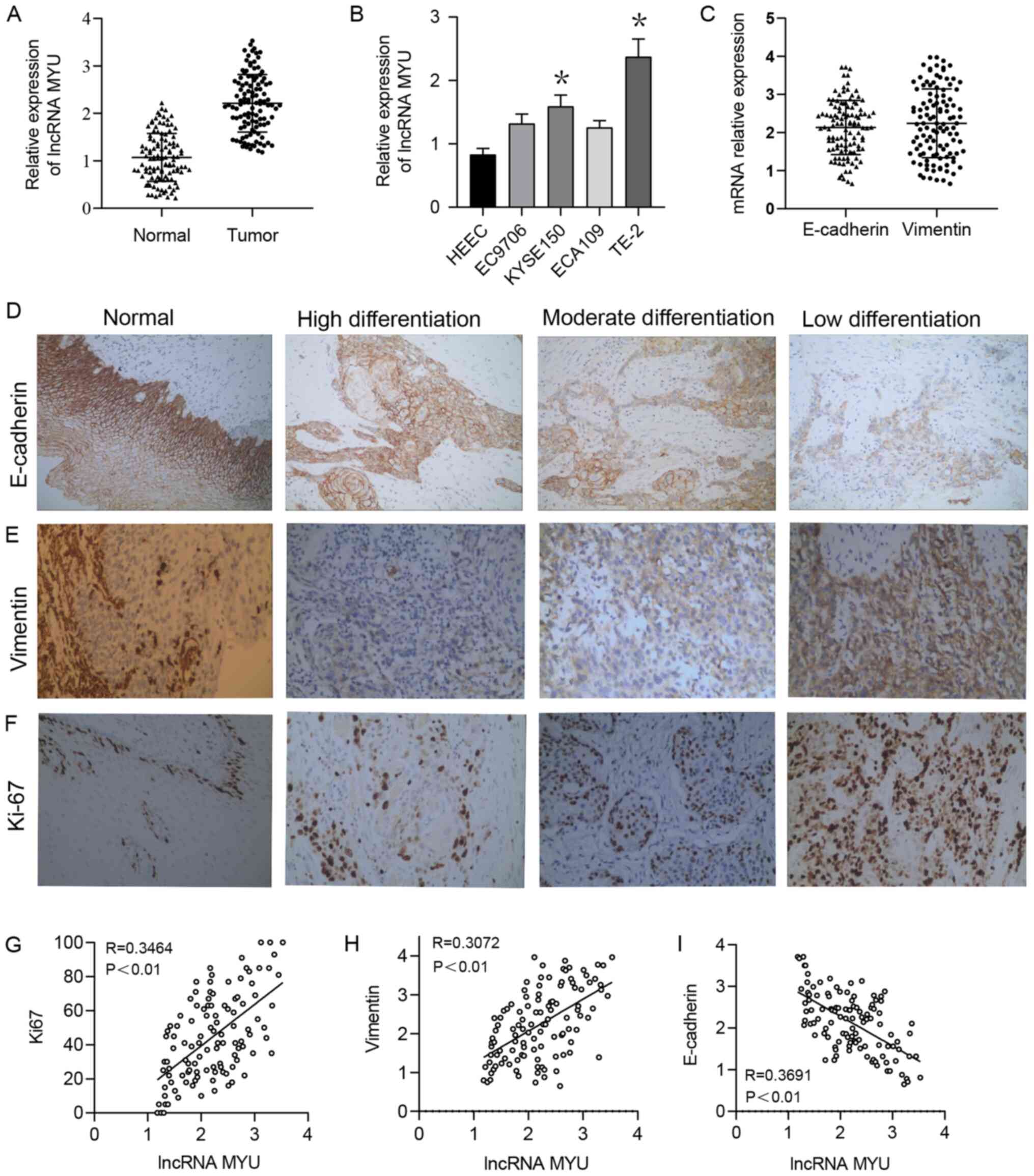
The RT-qPCR results suggested that the relative
expression of lncRNA MYU in ESCC tissues was significantly higher
than that in normal adjacent tissues, with relative expression
scores of 2.2122±0.606 and 1.0712±0.504, respectively (P<0.0001;
Fig. 1A). The median relative
expression of lncRNA MYU in ESCC tumour tissue was 2.195, and based
on this, low MYU expression was defined as an expression score of
<2.195 and high MYU expression was defined as an expression
score ≥2.195.
In addition, lncRNA MYU expression was examined in 3
human ESCC cell lines (EC9706, KYSE150, ECA109 and TE-2) and HEECs
by RT-qPCR. As lncRNA MYU expression was significantly higher in
TE-2 cells than in the other cells (Fig. 1B), this cell line was selected for
the subsequent experiments.
Expression of lncRNA MYU is associated
with the clinicopathological features of ESCC
Statistical analysis revealed that the relative
expression level of lncRNA MYU in ESCC tissue was significantly
associated with the histological grade (P=0.036), lymph node
metastasis (P=0.013) and TNM stage (P=0.019), but was not
associated with the patients' age or sex, vascular tumour thrombus
or T classification (P>0.05; Table
I).
Expression of lncRNA MYU is correlated
with Ki-67, Vimentin and E-cadherin expression
The expression of Ki-67 and the
epithelial-mesenchymal transition (EMT)-related proteins E-cadherin
and Vimentin in ESCC samples were then examined by
immunohistochemistry (Fig. 1C-F).
The Spearman rank correlation coefficient indicated that lncRNA MYU
expression was positively correlated with the expression of the
proliferation-associated proteins Ki-67 and the EMT-related
proteins Vimentin (Fig. 1G and
H, respectively) and negatively
correlated with the expression of the EMT-related protein
E-cadherin (Fig. 1I) in ESCC tumour
tissues.
Expression of lncRNA MYU is associated
with poor prognosis of patients with ESCC
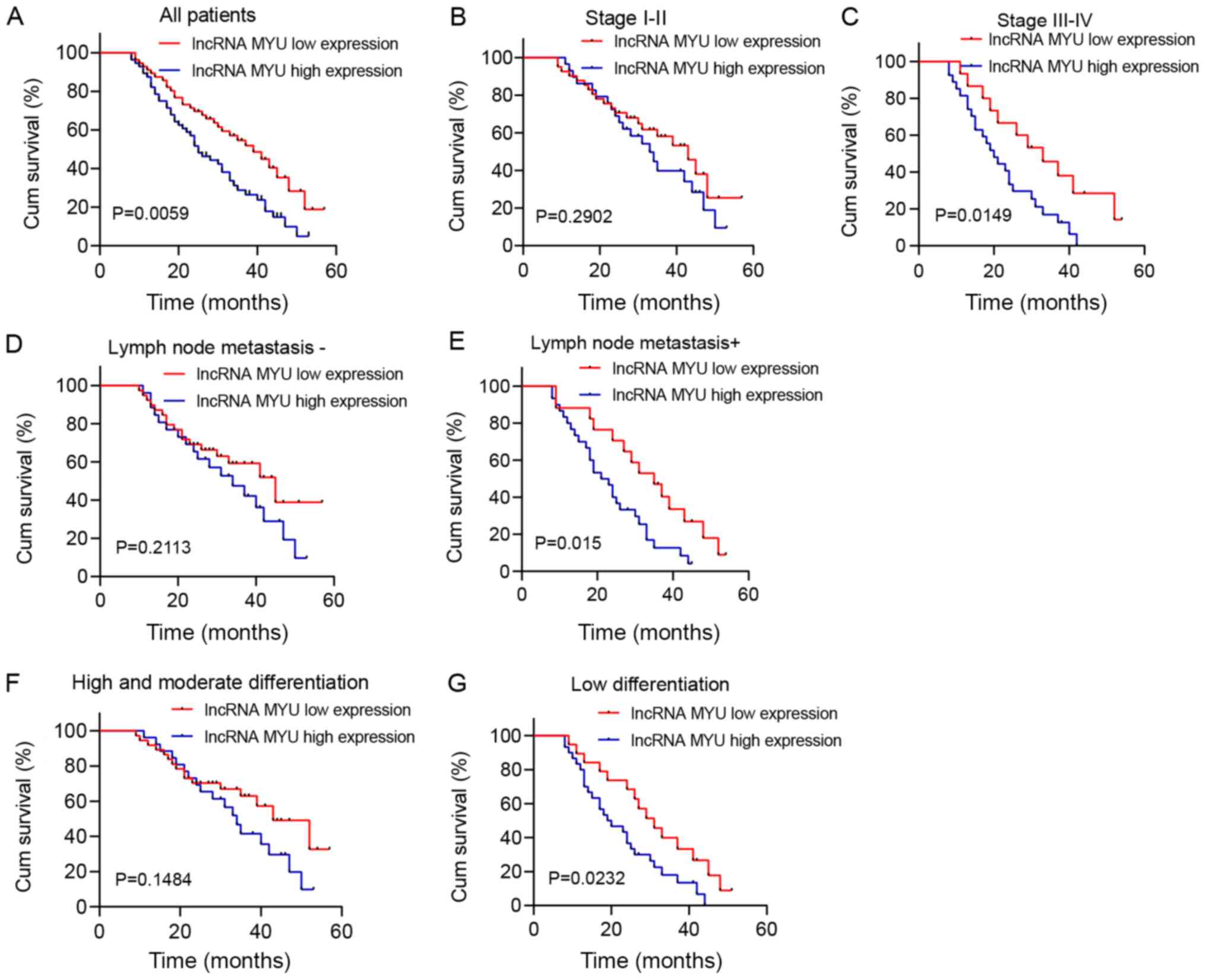
Survival analysis was performed by drawing
Kaplan-Meier plots (Fig. 2). It was
revealed that patients with high lncRNA MYU expression in their
ESCC tissues had shorter overall survival (OS) (median survival of
25 months) than patients with low lncRNA MYU expression (median
survival of 39 months; P=0.0059; Fig.
2A).
Further subgroup analysis suggested that in patients
with stage III/IV, lymph node metastasis and low degree of
differentiation, the OS of the high lncRNA MYU expression group was
significantly shorter than that of the low lncRNA MYU expression
group (P<0.05; Fig. 2C, E and G).
However, OS was not associated with MYU expression in patients with
stage I/II, lymph node metastasis-negative status and a
high/moderate degree of differentiation (P>0.05; Fig. 2B, D
and F).
lncRNA MYU promotes the proliferation
of TE-2 cells
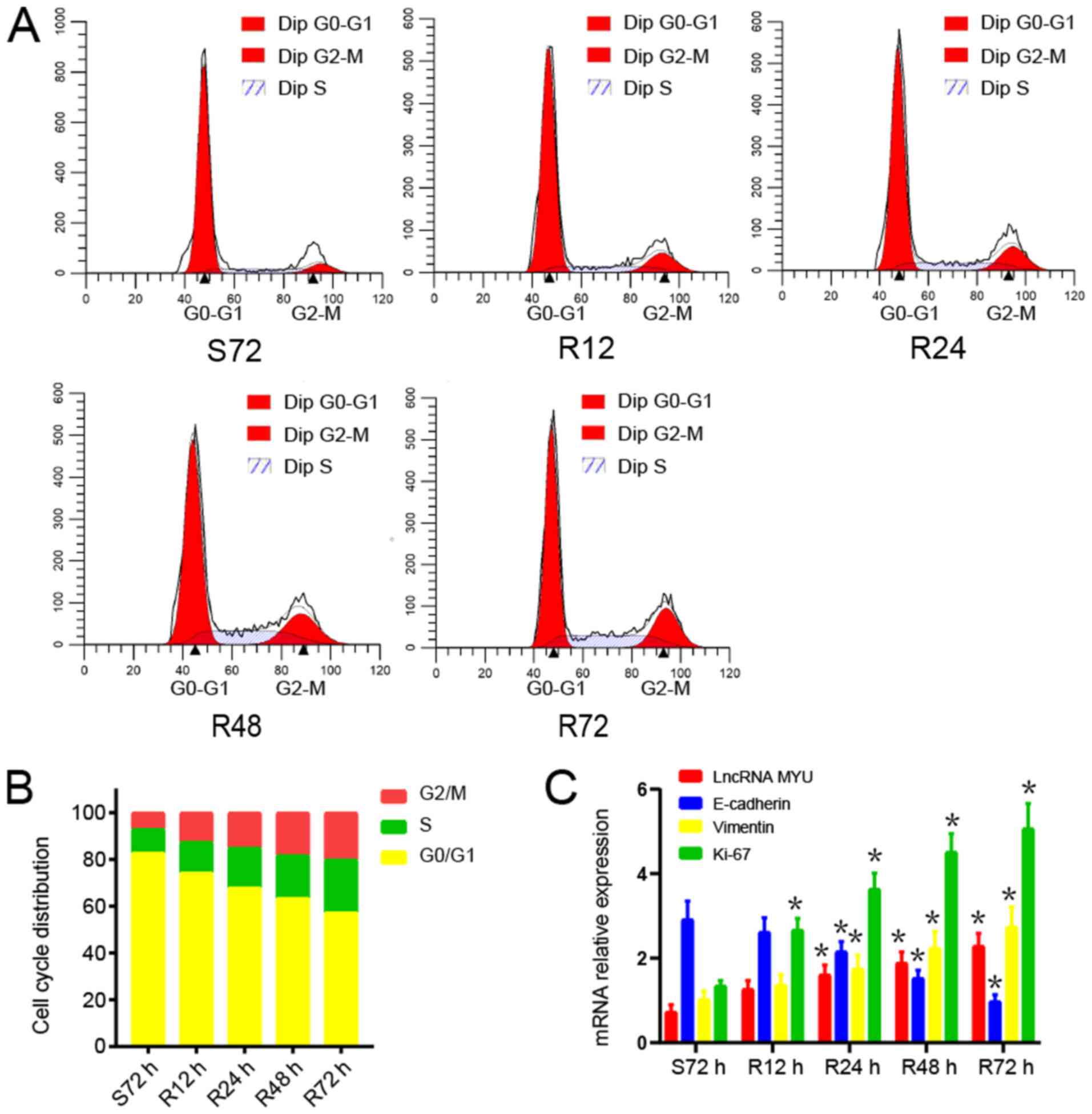
TE-2 cells were used for the serum
starvation-release experiment. After TE-2 cells were subjected to
serum starvation for 72 h, the serum supply was restored.
The changes in the cell cycle distribution after
serum starvation and restoration were analysed by flow cytometry.
After 72 h of serum starvation, TE-2 cells were arrested in G0/G1
phase. Once serum was restored, TE-2 cells were released from G0/G1
phase and gradually entered the S and G2/M phases (Fig. 3A and B).
The mRNA expression of lncRNA MYU, E-cadherin,
Vimentin and Ki-67 during the serum starvation-release experiment
was then examined. After serum stimulation, the expression of
lncRNA MYU, Vimentin and Ki-67 mRNA in TE-2 cells gradually
increased as the cell proliferation increased, while E-cadherin
mRNA expression declined gradually (Fig. 3C). These results indicated that
lncRNA MYU may promote the proliferation of tumour cells and be
related to tumour metastasis.
Knockdown of lncRNA MYU inhibits the
proliferation, migration and invasion of TE-2 cells
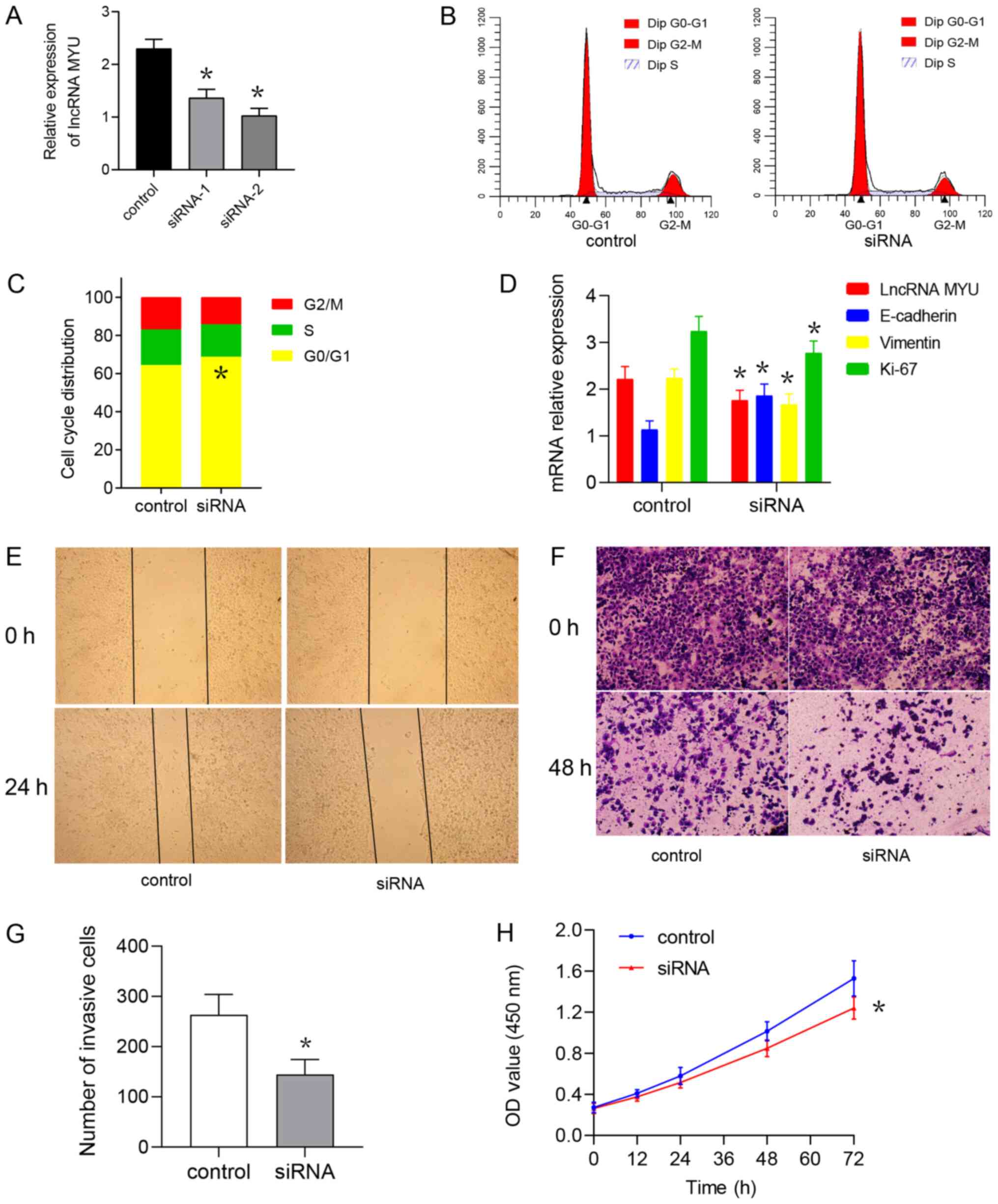
TE-2 cells were transfected with the lncRNA MYU
siRNA plasmid to interfere with the expression of lncRNA MYU.
RT-qPCR was used to identify the MYU siRNA with the highest
knockdown efficiency and the results suggested that the MYU
siRNA2-transfected group had the lowest lncRNA MYU expression and
the greatest knockdown with good transfection efficiency (Fig. 4A). The flow cytometry results also
indicated that the cell cycle was inhibited and that the proportion
of cells in G1/G0 phase was increased in the MYU siRNA2-transfected
group as compared with that in the MYU siRNA control-transfected
group (Fig. 4B and C). It was also observed that after lncRNA
MYU was knocked down, the mRNA expression of Vimentin and Ki-67 was
decreased in TE-2 cells, while E-cadherin mRNA expression was
elevated (Fig. 4D).
The wound-healing assay suggested that at 48 h after
scratching, the control group exhibited significantly faster
healing than the experimental group (Fig. 4E), indicating that interference with
lncRNA MYU expression is able to inhibit the migration ability of
TE-2 cells. The Transwell invasion test indicated that after the
expression of lncRNA MYU was knocked down, the number of cells
observed under the microscope in the experimental group und was
significantly lower than that observed in the control group
(Fig. 4F and G).
The CCK-8 cell proliferation assay indicated that
the absorbance of the MYU siRNA group at 0, 24, 48, 72 and 96 h was
lower than that of the control group at these time-points,
suggesting that interference with lncRNA MYU expression inhibited
the proliferation of TE-2 cells (Fig.
4H).
The results demonstrated that interference with the
expression of lncRNA MYU may inhibit the proliferation, migration
and invasion ability of TE-2 cells.
Mechanistically, lncRNA MYU may influence migration
and invasion of ESCC cells by regulating the expression of the
EMT-associated proteins E-cadherin and Vimentin.
Discussion
Despite continuous advancements of surgery and other
treatments in recent years, the prognosis of esophageal cancer has
not significantly improved (19).
Studies have indicated that abnormal expression of lncRNAs is
closely associated with the occurrence and development of
esophageal cancer. Certain lncRNAs may become effective markers and
therapeutic targets for the diagnosis and prognostic monitoring of
esophageal cancer.
It has been reported that MYU is highly expressed in
colorectal cancer and is associated with poor patient prognosis
(20). MYU may stabilize the
transcription of cyclin-dependent kinase 6 by combining with
heterogeneous nuclear ribonucleoprotein K, thereby promoting colon
cancer progression (21). A study
on prostate cancer indicated that MYU is significantly upregulated
in tumour tissue (22).
Furthermore, MYU expression was reported to be significantly
increased in squamous cell carcinoma of the lung and to be
associated with the number of lymph node metastases and poor
prognosis (23). The present study
suggested that the expression level of lncRNA MYU in ESCC tumour
tissues was significantly higher than that in normal adjacent
tissues and that the expression of lncRNA MYU was associated with
lymph node metastasis and TNM stage. Furthermore, survival analysis
indicated that the OS of patients with high expression of lncRNA
MYU was significantly shorter than that of patients with low
expression. In addition, it was demonstrated that MYU expression
was correlated with the expression of the proteins Ki-67, Vimentin
and E-cadherin, which were assessed by immunohistochemical
analysis. These results indicated that MYU has an important role in
the progression and metastasis of ESCC.
Interfering with MYU expression is able to inhibit
the growth and migration of cancer cells in prostate cancer and
there is no correlation between MYU expression and VPS9D1
expression (24). MYU may also be
transported outside of cells through exosomes to promote the
proliferation and migration of adjacent cells. The present study
further investigated the effect of MYU on the biological behaviour
of ESCC cell lines. The serum starvation and release assay
suggested that, as cell proliferation increased, the expression of
MYU also increased accordingly, while the mRNA expression of
E-cadherin decreased. Thus, this result suggested that MYU might
play a role in cell cycle regulation (22). However, due to the limitations of
the serum starvation and release assay, the specific role of MYU in
ESCC cell proliferation still needs to be studied further. After
MYU expression was knocked down, the proliferation, migration and
invasion of esophageal cancer cells were inhibited, while the mRNA
expression of E-cadherin was increased. MYU may regulate the
proliferation, migration and invasion of ESCC cells, but the
associated regulatory mechanisms require to be further studied and
discussed.
In conclusion, the results of the present study
suggested that an increase in the expression levels of lncRNA MYU
may be an indicator of poor prognosis in patients with ESCC and may
have value for determining the prognosis of patients with ESCC and
serve as a potential therapeutic target.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Science and
Technology Project of Nantong (grant no. JC2018154).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SG and LQ conceived and designed the study. JM, HS,
YL and SZ were responsible for collecting patient data and
conducted the data analysis. YL, SG and GM performed the
experiments and analyzed the test data. SG, LQ and GM wrote and
revised the manuscript and gave final approval for submission. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Affiliated Hospital of Nantong University
(Nantong, China). The patients provided written informed consent
for the use of their tumour tissues.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Chen W, Zheng R, Zhang S, Zeng H, Xia C,
Zuo T, Yang Z, Zou X and He J: Cancer incidence and mortality in
China, 2013. Cancer Lett. 401:63–71. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Liang H, Fan JH and Qiao YL: Epidemiology,
etiology, and prevention of esophageal squamous cell carcinoma in
China. Cancer Biol Med. 14:33–41. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Sanchez CA, Kawamura Y, Yamamoto Y,
Takeshita F and Ochiya T: Emerging roles of long non-coding RNA in
cancer. Cancer Sci. 109:2093–2100. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Quinn JJ and Chang HY: Unique features of
long non-coding RNA biogenesis and function. Nat Rev Genet.
17:47–62. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Chi Y, Wang D, Wang J, Yu W and Yang J:
Long non-coding RNA in the pathogenesis of cancers. Cells.
8(1015)2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Bunch H: Gene regulation of mammalian long
non-coding RNA. Mol Genet Genomics. 293:1–15. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Palmieri G, Paliogiannis P, Sini MC, Manca
A, Palomba G, Doneddu V, Tanda F, Pascale MR and Cossu A: Long
non-coding RNA CASC2 in human cancer. Crit Rev Oncol Hematol.
111:31–38. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Wahlestedt C: Targeting long non-coding
RNA to therapeutically upregulate gene expression. Nat Rev Drug
Discov. 12:433–446. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
11
|
Chen B, Li Y, He Y, Xue C and Xu F: The
emerging roles of long non-coding RNA in gallbladder cancer
tumorigenesis. Cancer Biomark. 22:359–366. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Xie H, Ma B, Gao Q, Zhan H, Liu Y, Chen Z,
Ye S, Li J, Yao L and Huang W: Long non-coding RNA CRNDE in cancer
prognosis: Review and meta-analysis. Clin Chim Acta. 485:262–271.
2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Ma J, Xiao Y, Tian B, Chen S, Zhang B, Wu
J, Wu Z, Li X, Tang J, Yang D, et al: Genome-wide analyses of long
non-coding RNA expression profiles and functional network analysis
in esophageal squamous cell carcinoma. Sci Rep.
9(9162)2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Zhang Y, Li R, Ding X, Zhang K and Qin W:
Upregulation of long non-coding RNA SNHG6 promote esophageal
squamous cell carcinoma cell malignancy and its diagnostic value.
Am J Transl Res. 11:1084–1091. 2019.PubMed/NCBI
|
|
15
|
Razavi M and Ghorbian S: Up-regulation of
long non-coding RNA-PCAT-1 promotes invasion and metastasis in
esophageal squamous cell carcinoma. EXCLI J. 18:422–428.
2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zong MZ, Feng WT, Du N, Yu XJ and Yu WY:
Upregulation of long noncoding RNA LEF1-AS1 predicts a poor
prognosis in patients with esophageal squamous cell carcinoma. Eur
Rev Med Pharmacol Sci. 23:7929–7934. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Ma J, Xiao Y, Tian B, Chen S, Zhang B, Wu
J, Wu Z, Li X, Tang J, Yang D, et al: Long noncoding RNA
lnc-ABCA12-3 promotes cell migration, invasion, and proliferation
by regulating fibronectin 1 in esophageal squamous cell carcinoma.
J Cell Biochem. 121:1374–1387. 2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhu D, Yu Y, Qi Y, Wu K, Liu D, Yang Y,
Zhang C and Zhao S: Long non-coding RNA CASC2 enhances the
antitumor activity of cisplatin through suppressing the Akt pathway
by inhibition of miR-181a in esophageal squamous cell carcinoma
cells. Front Oncol. 9(350)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Hou H, Meng Z, Zhao X, Ding G, Sun M, Wang
W and Wang Y: Survival of esophageal cancer in China: A pooled
analysis on hospital-based studies from 2000 to 2018. Front Oncol.
9(548)2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Yang L, Xu L, Wang Q, Wang M and An G:
Dysregulation of long non-coding RNA profiles in human colorectal
cancer and its association with overall survival. Oncol Lett.
12:4068–4074. 2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Kawasaki Y, Komiya M, Matsumura K, Negishi
L, Suda S, Okuno M, Yokota N, Osada T, Nagashima T, Hiyoshi M, et
al: MYU, a target lncRNA for Wnt/c-Myc signaling, mediates
induction of CDK6 to promote cell cycle progression. Cell Rep.
16:2554–2564. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wang J, Yang X, Li R, Wang L, Gu Y, Zhao
Y, Huang KH, Cheng T, Yuan Y and Gao S: Long non-coding RNA MYU
promotes prostate cancer proliferation by mediating the
miR-184/c-Myc axis. Oncol Rep. 40:2814–2825. 2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Tan J and Yang L: Long noncoding RNA
VPS9D1-AS1 overexpression predicts a poor prognosis in non-small
cell lung cancer. Biomed Pharmacother. 106:1600–1606.
2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wang X, Chen Q, Wang X, Li W, Yu G, Zhu Z
and Zhang W: ZEB1 activated-VPS9D1-AS1 promotes the tumorigenesis
and progression of prostate cancer by sponging miR-4739 to
upregulate MEF2D. Biomed Pharmacother. 122(109557)2020.PubMed/NCBI View Article : Google Scholar
|