Introduction
Angiogenesis is a process in which new blood vessels
arise from pre-existing capillaries (1). It is not only fundamental to
physiological processes, such as the menstrual cycle and wound
healing (2,3), but also the basis of the pathogenesis
of tumor growth (4). Angiogenesis
promotes tumor progression and metastasis (5). Anti-angiogenic therapy has been
approved as a novel treatment for malignant tumors, such as
medullary thyroid, non-small cell lung, renal cell and metastatic
colorectal cancers (5).
Endothelial cell (EC) migration is the basis of
angiogenesis (6). In the activation
phase of angiogenesis, vascular permeability increases, basement
membranes are degraded and ECs proliferate and migrate into the
extracellular space and form the new capillary buds. These cells
stop proliferating and migrating in the resolution phase, combine
to form the basement membrane and facilitate vessel maturation.
Finally, surrounding the newly formed vessels, mesenchymal cells
are recruited and differentiate into smooth muscle cells and
pericytes (1,7).
Dihydroartemisinin (DHA), a derivative of
artemisinin, has been identified to exert anti-angiogenic and
anti-EC migration effects (8-10);
however, the underlying molecular mechanism remains unclear.
TGF-β1 is the predominant and most ubiquitous member
of the TGF-β superfamily and serves a pivotal role during
angiogenesis (11). TGF-β1
activates two type-I receptors with opposite effects in EC
migration. One of the two type-I receptors is called activin
receptor-like kinase 5 (ALK5). This phosphorylates SMAD2 proteins,
resulting in inhibition of EC migration via the TGF-β1/ALK5/SMAD2
signaling pathway (12). SB-431542,
an ALK5 kinase inhibitor, blocks the inhibition of EC migration
(13). In contrast to ALK5, activin
receptor-like kinase 1 (ALK1), another TGF-β1 type-I receptor,
activates SMAD1/5, resulting in EC migration via the
TGF-β1/ALK1/SMAD1/5 signaling pathway (12,14).
In the present study, it was hypothesized that DHA
could inhibit EC migration via the TGF-β1/ALK5/SMAD2 signaling
pathway. Human umbilical vein ECs (HUVECs) were treated with DHA
and SB431542, followed by migration assays. The effects of DHA
treatment on the TGF-β1/ALK5/SMAD2 signaling pathway were assessed
via western blotting.
Materials and methods
Cell culture
HUVECs were obtained from American Type Culture
Collection and cultured in Dulbecco's modified Eagle's medium
(DMEM) supplemented with 10% fetal bovine serum (FBS; Gibco, Thermo
Fisher Scientific, Inc.) and antibiotics (100 IU/ml penicillin and
100 µg/ml streptomycin). The cells were cultured in humidified air
at 37˚C with 5% CO2. DHA was obtained from Sigma-Aldrich
(Merck KGaA), and SB431542 was purchased from Cell Signaling
Technology, Inc. They were both dissolved at 10 mM in dimethyl
sulfoxide.
Cell proliferation assay
The anti-angiogenesis effect of DHA is
dose-dependent. In our previous study, a concentration of 25 µM or
higher of DHA was demonstrated to reduce the growth of HUVECs
significantly (15). Therefore, 25
µM of DHA was used in the present study. An MTT Cell Proliferation
Assay kit (cat. no. 1009365; Cayman Chemical Company) was used to
evaluate cell proliferation. The cells were cultured at a density
of 5x103 cells/well in a 96-well plate. The next day, 25
µM DHA was added to the culture medium. Similarly, the control
groups were treated with equivalent dimethyl sulfoxide. After 24 h,
MTT solution (10 µl; 5 mg/ml) was added to each well, followed by
incubation for 4 h. The culture medium was discarded, and dimethyl
sulfoxide (150 µl) was added. These steps were all performed at
37˚C. Finally, a 96-well plate reader (Molecular Devices, LLC) was
used to analyze colorimetric intensity at a wavelength of 570 nm.
Each experiment was repeated three times.
Wound healing assay
Fully confluent HUVECs were starved for 2 h with 2%
FBS (a stress growing condition) at 37˚C. Wounds were created
across the monolayer of cells by scratching the surface with a
sterile pipette tip. Then, 25 µM DHA or 20 µM SB431542 was added to
the DMEM supplemented with 10% FBS, a physiological growing
condition, in DHA treatment group or SB431542 treatment group, as
previously described (16).
Similarly, the control groups were treated with equivalent dimethyl
sulfoxide. Eventually, images were captured after incubation for 0
and 24 h at 37˚C. ImageJ software 1.4.3 (National Institutes of
Health) was used to determine cell migration at the 0 and 24 h time
points. Cells were viewed using x100 magnification with a
phase-contrast microscope (Olympus Corporation), as previously
described (17).
Transwell assay
Briefly, HUVECs (1x105 cells/well) were
seeded in the upper chamber in 24-well plates with 25 µM DHA or 20
µM SB431542 in DMEM supplemented with 2% FBS. Similarly, the
control groups were treated with equivalent dimethyl sulfoxide. The
bottom chambers were filled with 500 µl DMEM supplemented with 10%
FBS overnight at 37˚C. After 24 h of incubation, cells that had
migrated through the membrane were fixed with 79.2% methanol and
stained with 0.1% crystal violet for 30 min at room temperature.
Migration was quantified by manually counting the number of stained
cells at x200 magnification with a phase-contrast microscope
(Olympus Corporation).
Western blot analysis
Western blotting was performed as previously
described (17). GAPDH was used as
the loading control. The following antibodies were used: Rabbit
anti-TGF-β (1:1,000; cat. no. ab66043; Abcam), rabbit anti-ALK1
(1:1,000; cat. no. ab108207; Abcam), rabbit anti-ALK5 (1:1,000;
cat. no. sc-398; cat no. F0315; Santa Cruz Biotechnology, Inc.),
rabbit anti-phospho-SMAD2 (1:5,000; cat. no. ab188334; Abcam),
rabbit anti-SMAD2 (1:5,000; cat. no. ab40855; Abcam), rabbit
anti-GAPDH (1:8,000; cat. no. I0494-I-AP; ProteinTech Group, Inc.)
and goat anti-rabbit IgG (1:10,000; cat. no. ab150077; Abcam).
Reverse transcription-quantitative
(RT-q)PCR
As previously described (18), following the addition of DMSO and
DHA, HUVECs were cultured for 24 h. HUVECs were washed twice using
ice-cold PBS. The RNAiso Plus kit (Takara Bio, Inc.) was used to
extract total RNA. cDNA was generated using a reverse transcriptase
kit (ChamQ Universal SYBR qPCR Master Mix; cat. no. Q711-02/03;
Vazyme Biotech Co., Ltd.). The total RNA (~1,000 ng), 4 x g DNA
wiper Mix (4 µl) and RNase-free ddH2O were incubated for 2 min at
42˚C in A200 Gradient Thermal cycler (Thermo Fisher Scientific,
Inc.). After adding 5xHiScriptIII qRT SuperMix into the
aforementioned mentioned mixture, cDNA was constructed at 37˚C 15
min and 85˚C for 5 sec. cDNA was amplified using a Taq DNA
polymerase kit (Tiangen Biotech Co., Ltd.). The qPCR reaction
consisted of three steps: Pre-denaturation (95˚C for 30 sec),
cyclic reaction (95˚C for 10 sec; 60˚C for 30 sec; repeated x40)
and melting curve (95˚C for 15 sec; 60˚C for 60 sec; and 95˚C for
15 sec). The primer sequences were as follows: ALK5 forward,
5'-GCCGTTTGACTGAAGGCTG-3' and reverse, 5'-GGGCATCCCAAGCCTCATC-3';
and GAPDH forward, 5'-TGATGACATCAAGAAGGTGGTGAAG-3' and reverse,
5'-TCCTTGGAGGCCATGTGGGCCAT-3'.
Statistical analysis
Data were expressed as mean ± standard error (SE).
n=3, indicated that each experiment was repeated 3 different times.
Densitometric analysis of western blots was performed using Image J
software 1.4.3 (National Institutes of Health). An unpaired
Student's t-test or one-way ANOVA followed by the Tukey's multiple
comparisons test was used for statistical analysis. Statistical
analyses were performed using GraphPad Pro Prism 5.0 (GraphPad
Software, Inc.). P<0.05 was considered to indicate a
statistically significant difference.
Results
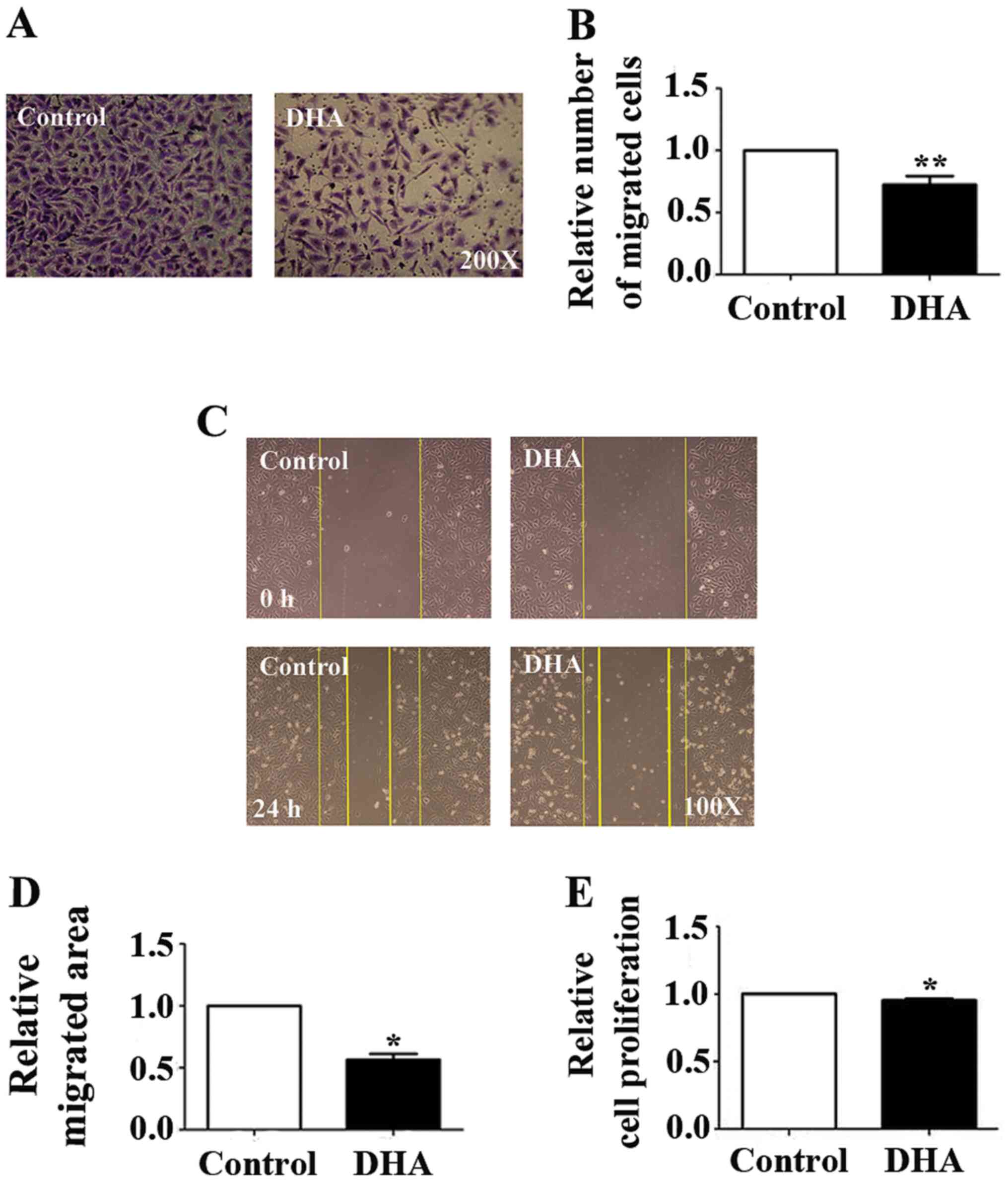
DHA reduces EC migration
To verify the effects of DHA on EC migration,
Transwell and wound healing assays were performed. The numbers of
HUVECs that migrated through the polycarbonate membrane in the
Transwell assay were significantly reduced (33.25%; P<0.01;
Fig. 1A and B). Furthermore, the area of the wound in
the DHA treatment group was significantlyreduced in the wound
healing assay (43.42%; P<0.05; Fig.
1C and D). It was also
demonstrated that relative cell proliferation was significantly
decreased in the DHA-treated groups in the MTT assay (4.70%;
P<0.05; Fig. 1E). These data
suggested that inhibitory effects of DHA on migration were more
significant than those on proliferation.
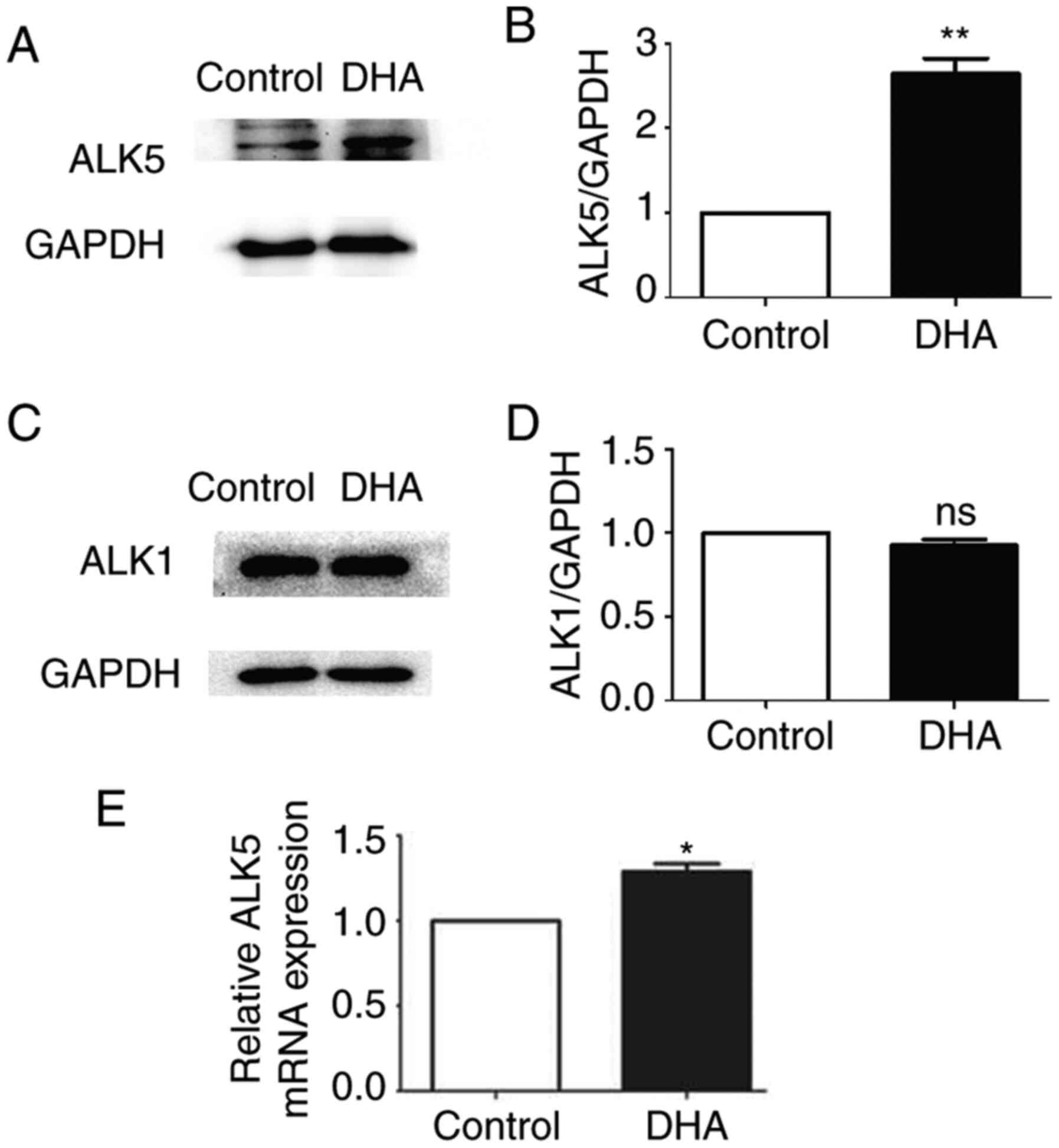
DHA treatment increases the expression
levels of ALK5
In order to examine the activation of two different
TGF-β1 type-I receptors in ECs, the protein expression levels of
ALK1 and ALK5 were analyzed. The present study revealed significant
upregulation of ALK5 protein expression in the DHA treatment group
after 16 h of DHA treatment compared with control group (164.31%;
P<0.01; Fig. 2A and B). However, ALK1 protein expression
remained unchanged in the DHA treatment group (Fig. 2C and D). Additionally, the mRNA expression
levels of ALK5 were examined, and DHA treatment upregulated ALK5
expression at the transcriptional level in the DHA treatment group
(28.97%; P<0.05; Fig. 2E). These
data demonstrated that DHA-induced upregulated ALK5 expression and
may not affect the ALK1/SMAD1/5 signaling pathways.
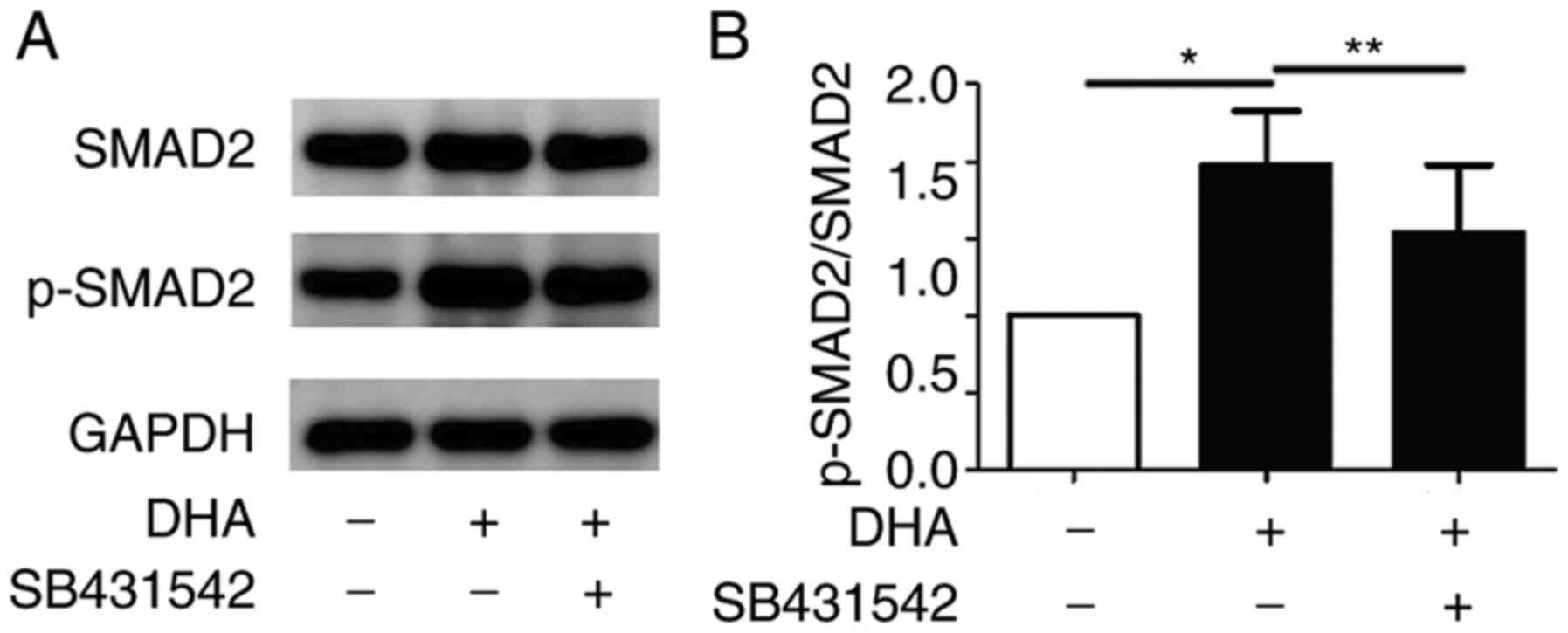
DHA treatment increases SMAD2
phosphorylation
When the TGF-β1/ALK5/SMAD2 signaling pathway is
activated, ALK5 phosphorylates SMAD2. The phosphorylated SMAD2 is
translocated into the nucleus and activates the expression of
target genes (19). Western blot
analysis revealed that the levels of phosphorylated SMAD2 in the
DHA treatment group were increased following DHA treatment (97.49%;
P<0.05; Fig. 3A and B). Additionally, the expression levels of
SMAD2 remained unchanged in the DHA treatment group. In the
presence of SB431542, the increased levels of phosphorylated SMAD2
due to DHA were partially abrogated in the DHA and SB431542
treatment groups (39.17%; P<0.01; Fig. 3A and B).
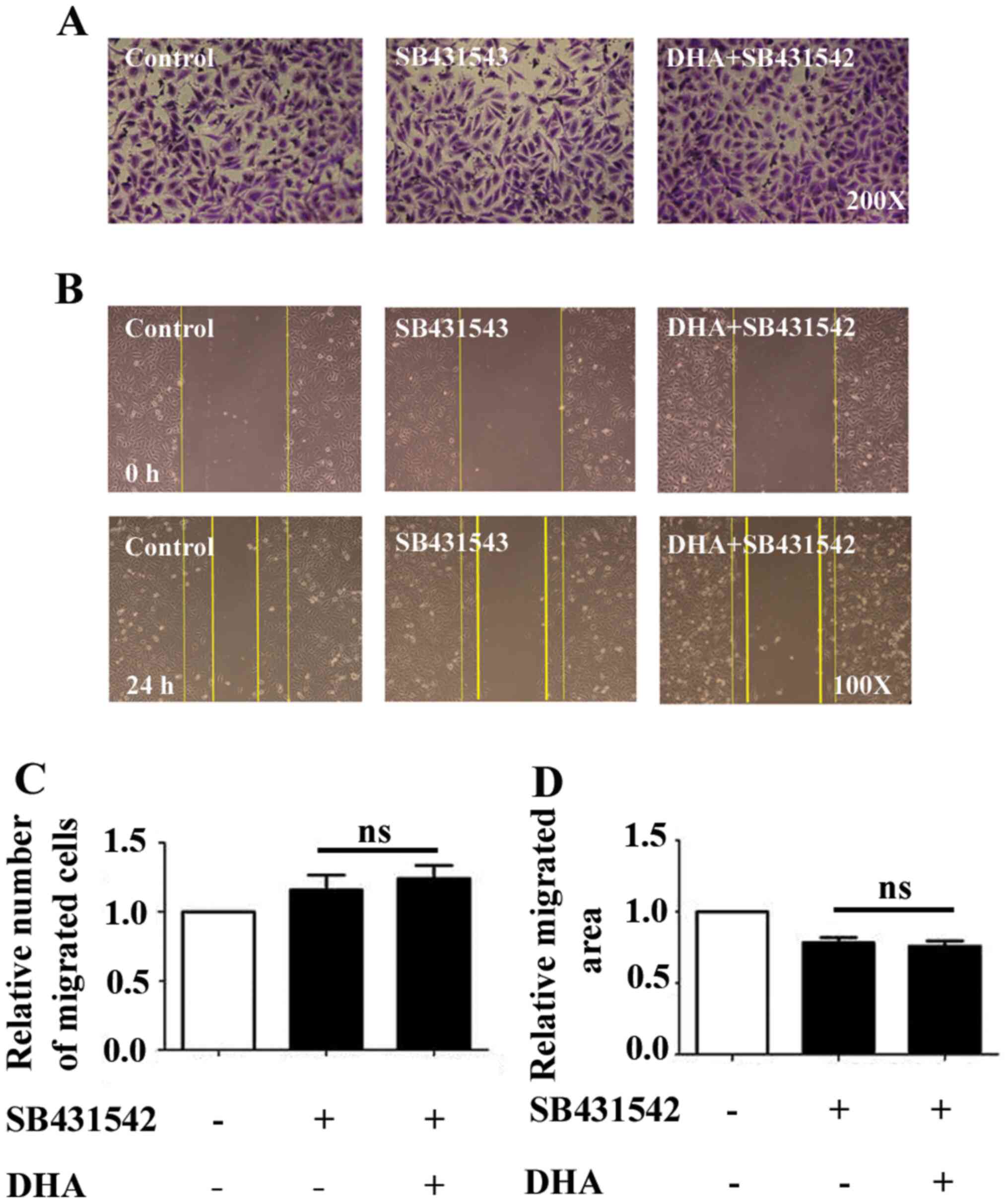
SB431542 rescues DHA-induced
repression
SB431542 is an effective inhibitor that represses
ALK5/SMAD2 phosphorylation (13).
In the presence of SB431542, DHA did not inhibit EC migration, and
the reduction in EC migration by DHA was rescued in the DHA and
SB431542 treatment groups in the Transwell and wound healing assays
(Fig. 4A and B). The data demonstrated that the blockade
of the TGF-β1/ALK5/SMAD2 signaling pathway mediated the anti-EC
migration effects of DHA.
Discussion
Abnormal angiogenesis serves a crucial role in tumor
survival, growth, invasion and metastasis (5). Anti-angiogenic agents, such as
tyrosine kinase inhibitors, monoclonal antibodies and fusion
peptides, are increasingly being used as treatments for certain
malignant tumors (20,21). However, the targeting of single
angiogenic molecules is insufficient against the countless
stimulators produced by tumors and their microenvironment.
DHA has been identified to exhibit remarkable
activities, such as inhibiting angiogenesis, and inhibiting tube
formation and EC migration (15,17,22).
The present study aimed to examine whether DHA inhibited EC
migration via the TGF-β1/ALK/SMAD signaling pathway. EC migration
is a directional movement caused by chemotactic, haptotactic and
mechanical stimuli, and it is an important part of angiogenesis
(6). The dynamic and multistep
process of EC migration includes cell sensing, extension,
attachment, contraction, rear release and recycling movement. The
whole process is affected by cytoskeletal recombination,
chemokines, extracellular matrix, integrin chemotaxis, mechanical
stimulation and signaling pathways (23). In our previous study, DHA exhibited
dose-dependent anti-angiogenic effects, and treatment with ≥25 µM
DHA markedly reduced the proliferation of ECs (15). Consistent with our previous studies
(15,17), DHA treatment in the present study
not only inhibited proliferation, but also inhibited migration. It
was demonstrated that the inhibitory effects of DHA on migration
were more significant than the effect on proliferation in the
present experiments.
TGF-β1 is primarily synthesized by platelets,
dendritic cells, epithelial cells, fibroblasts, lymphocytes and
macrophages/monocytes (24). TGF-β1
is released from cells or extracellular matrix-binding protein as
an inactive precursor. In the blood and tissues, TGF-β1 is almost
undetectable in its active form (25). DHA may regulate TGF-β1 activity by
increasing its synthesis and bioavailability, as reported in
previous studies (26).
TGF-β1 serves a pivotal role during angiogenesis
(11,27). It induces ALK1 activation, resulting
in increased migration, and activates ALK5, resulting in inhibition
of the migration of ECs (7). In
ECs, the coexistence of the ALK1/SMAD1/5 and the ALK5/SMAD2
signaling pathways is dependent on the pattern of genes activated
in a given circumstance (28). ALK1
signaling requires intact ALK5 for activation via TGF-β1(14). ALK5 is required in order to recruit
ALK1 into a heteromeric complex with type-II TGF-β receptor, and
ALK5 promotes ALK1 kinase activity and SMAD1/5 phosphorylation.
Additionally, SB-431542 has been demonstrated to abrogate both
TGFβ-induced SMAD2 and SMAD1/5 activation (14). More specifically, ALK5/SMAD2 is
directly antagonized by ALK1(14).
ALK1 negatively regulates TGF-β1 signaling by ALK5 through a
promoter construct, p3TP-Lux, which contains multiple copies of the
TGF-β1 response elements in human hepatoblastoma cell line (HepG2)
cells (29). Moreover,
overexpression of wild-type or constitutively active (ca) ALK1
inhibited TGF-β- or caALK5-induced12-luc reporter in
mouse embryonic endothelial and HepG2 cells (14). However, in the present study, no
significant changes in ALK1 expression levels in ECs treated with
DHA were observed. DHA treatment upregulated the expression levels
of ALK5 and increased the phosphorylation of SMAD2. In the presence
of SB431542, DHA did not activate the TGF-β1/ALK5/SMAD2 signaling
pathway or induce a decrease in EC migration. The current findings
suggested that DHA inhibited EC migration via the
TGF-β1/ALK5/SMAD2-dependent signaling pathway and may not influence
the ALK1 signaling pathways.
The TGF-β1/ALK5/SMAD2 signaling pathway has been
reported to be upregulated by artemisinin and its derivatives
(30). It may downregulate
different cytokines, such as the Rho small GTPases, cell division
cycle 42 and phosphoinositide 3-kinase, to inhibit EC sensing of
the motile stimuli, formation of protruding lamellipodia, cellular
extension, attachment and contraction in order to inhibit EC
migration (31-36).
In conclusion, the present results demonstrated that
treatment with DHA inhibited EC migration via the TGF-β1/ALK5/SMAD2
signaling pathway. These experiments may offer novel insights into
the anti-angiogenic effects of DHA against malignant tumors. DHA
may represent a novel drug that may improve the survival rate of
patients with malignant tumors.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81873473,
Traditional Chinese Medicine Research Projects of Shandong Province
(grant nos. 2019-0370 and 2015-285) and the Academic Promotion
Program of Shandong First Medical University (grant no.
2019QL014).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LG carried out the experimental work, the data
collection, interpretation and preparation of the manuscript. XW
carried out experimental work and the data collection and
interpretation. YH participated in the design and coordination of
experimental work. RS, LZ and FL participated in the study design,
data collection, analysis of data and preparation of the
manuscript. JL conceived the study design, the analysis and
interpretation of data, drafted the manuscript, and gave final
approval of the version to be published. LG and XW confirmed the
authenticity of all the raw data. All authors read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Carmeliet P: Mechanisms of angiogenesis
and arteriogenesis. Nat Med. 6:389–395. 2000.PubMed/NCBI View
Article : Google Scholar
|
|
2
|
Knighton DR, Silver IA and Hunt TK:
Regulation of wound-healing angiogenesis-effect of oxygen gradients
and inspired oxygen concentration. Surgery. 90:262–270.
1981.PubMed/NCBI
|
|
3
|
Reynolds LP, Killilea SD and Redmer DA:
Angiogenesis in the female reproductive system. FASEB J. 6:886–892.
1992.PubMed/NCBI
|
|
4
|
Weidner N, Carroll PR, Flax J, Blumenfeld
W and Folkman J: Tumor angiogenesis correlates with metastasis in
invasive prostate carcinoma. Am J Pathol. 143:401–409.
1993.PubMed/NCBI
|
|
5
|
Al-Abd AM, Alamoudi AJ, Abdel-Naim AB,
Neamatallah TA and Ashour OM: Anti-angiogenic agents for the
treatment of solid tumors: Potential pathways, therapy and current
strategies-A review. J Adv Res. 8:591–605. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Klagsbrun M and Moses MA: Molecular
angiogenesis. Chem Biol. 6:R217–R224. 1999.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Goumans MJ, Valdimarsdottir G, Itoh S,
Rosendahl A, Sideras P and ten Dijke P: Balancing the activation
state of the endothelium via two distinct TGF-beta type I
receptors. EMBO J. 21:1743–1753. 2002.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Crespo-Ortiz MP and Wei MQ: Antitumor
activity of artemisinin and its derivatives: From a well-known
antimalarial agent to a potential anticancer drug. J Biomed
Biotechnol. 2012(247597)2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Wei T and Liu J: Anti-angiogenic
properties of artemisinin derivatives (review). Int J Mol Med.
40:972–978. 2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Chen HH, Zhou HJ, Wang WQ and Wu GD:
Antimalarial dihydroartemisinin also inhibits angiogenesis. Cancer
Chemother Pharmacol. 53:423–432. 2004.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Pepper MS: Transforming growth
factor-beta: Vasculogenesis, angiogenesis, and vessel wall
integrity. Cytokine Growth Factor Rev. 8:21–43. 1997.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Goumans MJ, Lebrin F and Valdimarsdottir
G: Controlling the angiogenic switch: A balance between two
distinct TGF-b receptor signaling pathways. Trends Cardiovasc Med.
13:301–307. 2003.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Liu Z, Kobayashi K, van Dinther M, van
Heiningen SH, Valdimarsdottir G, van Laar T, Scharpfenecker M,
Löwik CW, Goumans MJ, Ten Dijke P and Pardali E: VEGF and
inhibitors of TGFbeta type-I receptor kinase synergistically
promote blood-vessel formation by inducing alpha5-integrin
expression. J Cell Sci. 122:3294–3302. 2009.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Goumans MJ, Valdimarsdottir G, Itoh S,
Lebrin F, Larsson J, Mummery C, Karlsson S and ten Dijke P: Activin
receptor-like kinase (ALK)1 is an antagonistic mediator of lateral
TGFbeta/ALK5 signaling. Mol Cell. 12:817–828. 2003.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Dong F, Zhou X, Li C, Yan S, Deng X, Cao
Z, Li L, Tang B, Allen TD and Liu J: Dihydroartemisinin targets
VEGFR2 via the NF-κB pathway in endothelial cells to inhibit
angiogenesis. Cancer Biol Ther. 15:1479–1488. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Martins MD, Silveira FM, Webber LP, Wagner
VP, Martins MAT, Squarize CH and Castilho RM: The impact of
photobiomodulation therapy on the biology and behavior of head and
neck squamous cell carcinomas cell lines. J Photochem Photobiol B.
209(111924)2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Guo L, Dong F, Hou Y, Cai W, Zhou X, Huang
AL, Yang M, Allen TD and Liu J: Dihydroartemisinin inhibits
vascular endothelial growth factor-induced endothelial cell
migration by a p38 mitogen-activated protein kinase-independent
pathway. Exp Ther Med. 8:1707–1712. 2014.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Derynck R, Zhang Y and Feng XH: Smads:
Transcriptional activators of TGF-beta responses. Cell. 95:737–740.
1998.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Hurwitz H, Fehrenbacher L, Novotny W,
Cartwright T, Hainsworth J, Heim W, Berlin J, Baron A, Griffing S,
Holmgren E, et al: Bevacizumab plus irinotecan, fluorouracil, and
leucovorin for metastatic colorectal cancer. N Engl J Med.
350:2335–2342. 2004.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Ilhan-Mutlu A, Osswald M, Liao Y, Gömmel
M, Reck M, Miles D, Mariani P, Gianni L, Lutiger B, Nendel V, et
al: Bevacizumab prevents brain metastases formation in lung
adenocarcinoma. Mol Cancer Ther. 15:702–710. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Gao P, Wang LL, Liu J, Dong F, Song W,
Liao L, Wang B, Zhang W, Zhou X, Xie Q, et al: Dihydroartemisinin
inhibits endothelial cell tube formation by suppression of the
STAT3 signaling pathway. Life Sci. 242(117221)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Horwitz R and Webb D: Cell migration. Curr
Biol. 13:R756–R759. 2003.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Krzemień S and Knapczyk P: Current review
on the role of transforming growth factor beta (TGF-beta) in some
pathological disorders. Wiad Lek. 58:536–539. 2005.PubMed/NCBI(In Polish).
|
|
25
|
Huang FY, Mei WL, Li YN, Tan GH, Dai HF,
Guo JL, Wang H, Huang YH, Zhao HG, Zhou SL and Lin YY:
Toxicarioside A inhibits tumor growth and angiogenesis: Involvement
of TGF-β/endoglin signaling. PLoS One. 7(e50351)2012.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Kajdaniuk D, Marek B, Borgiel-Marek H and
Kos-Kudła B: Transforming growth factor β1 (TGFβ1) in physiology
and pathology. Endokrynol Pol. 64:384–396. 2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Jiang Y, Zhou X, Hu R and Dai A:
TGF-β1-induced SMAD2/3/4 activation promotes RELM-β transcription
to modulate the endothelium-mesenchymal transition in human
endothelial cells. Int J Biochem Cell Biol. 105:52–60.
2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Byfield SD and Roberts AB: Lateral
signaling enhances TGF-beta response complexity. Trends Cell Biol.
14:107–111. 2004.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Oh SP, Seki T, Goss KA, Imamura T, Yi Y,
Donahoe PK, Li L, Miyazono K, ten Dijke P, Kim S and Li E: Activin
receptor-like kinase 1 modulates transforming growth factor-beta 1
signaling in the regulation of angiogenesis. Proc Natl Acad Sci
USA. 97:2626–2631. 2000.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Li T, Chen H, Yang Z, Liu XG, Zhang LM and
Wang H: Evaluation of the immunosuppressive activity of artesunate
in vitro and in vivo. Int Immunopharmacol. 16:306–312.
2013.PubMed/NCBI View Article : Google Scholar
|
|
31
|
van Nieuw Amerongen GP, Koolwijk P,
Versteilen A and van Hinsbergh VW: Involvement of RhoA/Rho kinase
signaling in VEGF-induced endothelial cell migration and
angiogenesis in vitro. Arterioscler Thromb Vasc Biol. 23:211–217.
2003.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Qi JH and Claesson-Welsh L: VEGF-induced
activation of phosphoinositide 3-kinase is dependent on focal
adhesion kinase. Exp Cell Res. 263:173–182. 2001.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Lamalice L, Le Boeuf F and Huot J:
Endothelial cell migration during angiogenesis. Circ Res.
100:782–794. 2007.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Gong C, Stoletov KV and Terman BI: VEGF
treatment induces signaling pathways that regulate both actin
polymerization and depolymerization. Angiogenesis. 7:313–321.
2004.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Liu J, Wada Y, Katsura M, Tozawa H, Erwin
N, Kapron CM, Bao G and Liu J: Rho-associated coiled-coil kinase
(ROCK) in molecular regulation of angiogenesis. Theranostics.
8:6053–6069. 2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Castañares C, Redondo-Horcajo M,
Magán-Marchal N, ten Dijke P, Lamas S and Rodriguez-Pascual F:
Signaling by ALK5 mediates TGF-beta-induced ET-1 expression in
endothelial cells: A role for migration and proliferation. J Cell
Sci. 120:1256–1266. 2007.PubMed/NCBI View Article : Google Scholar
|