Introduction
Diabetes, a debilitating disease that threatens the
health of individuals globally, is becoming a public health issue
concerning scientists worldwide since 2015 (1,2). The
primary pathological mechanisms underlying the development of
diabetes include glycometabolism and lipid metabolism dysregulation
(3,4). If phosphorylation of glucose
decreases, the signaling pathways of glycolysis, pentose phosphate
and tricarboxylic acid cycle will be blocked (5,6). The
synthesis of glycogen is reduced and decomposition increases,
resulting in a decline of the body's ability to utilize and take up
glucose (7-9),
leading to a glycometabolism disorder (10,11).
When the ability to utilize and take up glucose from fat tissues
declines, the concentration of triglycerides in the plasma
increases and the synthesis of fat reduces (7,12),
resulting in a lipid metabolism disorder (13). The main indexes of lipid metabolism
include fatty acid synthetase, acetyl-CoA carboxylase, acyl-CoA
oxidase, and carnitine palmityl transferase (14,15).
There are two types of diabetes, type I and II
(T2DM). The number of patients with T2DM makes up 80-90% of all
patients with diabetes in Thailand in 2018(16). T2DM is a long-term disease caused by
metabolic disorders as a result of heredity and environmental
factors, which results in a functional defect of islet β cells and
the secretion of insulin (17,18).
In this case, the metabolic disorders involve carbohydrates,
proteins, fat, electrolytes and water. Currently, treatments for
diabetes include hypoglycemic drugs, including insulin, insulin
analogues and sulfonylurea drugs (19-21).
However, these hypoglycemic drugs can cause numerous side effects,
including liver and kidney injury and insulin resistance (22). Certain Traditional Chinese Medicines
are being studied as potential therapeutics for diabetes to
increase drug efficacy and decrease side effects.
Ganoderma lucidum (G. lucidum)
polysaccharide (GLP) is one of the effective ingredients in the
mushroom G. lucidum (23,24).
It has been reported that GLP significantly decreased the
concentration of blood glucose, promoted the secretion of insulin,
improved glucose tolerance, regulated the concentration of fat in
the blood and postponed the development of diabetic complications
(3,25). However, the underlying mechanisms
remain unknown. Currently, family with sequence similarity 3
(FAM3C), which is composed of FAM3A-D, has been reported to be
associated with the regulation of glucometabolism and lipid
metabolism (26,27). Chen et al (28) reported that the FAM3C-heat shock
factor 1 (HSF1)-calmodulin (CaM) signaling pathway was
significantly inhibited in the liver of a streptozotocin
(STZ)-induced type I diabetic mouse model. The expression of
glycogen was inhibited, and the high concentration of blood glucose
of the mice was significantly improved by overexpressing FAM3C and
activating the HSF1-CaM-AKT signaling pathway. Furthermore, Zhang
et al (27) demonstrated
that as a novel hepatocyte factor, FAM3C significantly inhibited
the HSF1-CaM-AKT signaling pathway, by which gluconeogenesis and
lipogenesis were suppressed.
In the current study, the hypoglycemic and
hypolipidemic effects of GLP were investigated in a T2DM db/db
mouse model, along with an investigation of the effects of GLP on
the FAM3C-HSF1-CaM signaling pathway in order to determine the
underlying mechanism of GLP in the lipid metabolism dysregulation
in diabetes.
Materials and methods
Ethics approval
All animal experiments performed in the current
study were authorized by the Ethics Committee of Zhejiang Chinese
Medical University and carried out according to the guidelines for
the Care and Use of Laboratory Animals and to the Principles of
Laboratory Animal Care and Protection (approval no.
ZSLL-2018-172).
Animals
A total of 24 adult male (age, 6 weeks; weight,
18-24 g) idiopathic T2DM model mice (db/db) and 6-+/db male mice
were purchased from Vital River Laboratories Co., Ltd.. The housing
room was maintained between 20.0-26.1˚C, with an average humidity
between 30-70% and under a 12-h/12-h light/dark cycle. The db/db
mice were fed a high-fat diet for 2 weeks and -+/db mice were fed
an ordinary diet as a control. All the animals had free access to
food and water. Following this, 0.08 ml blood was collected from
the tail vein at the endpoint to detect the concentration of
glucose in the blood. The animals with concentrations of blood
glucose >11.1 mmol/l were selected as T2DM model mice.
Preparation of GLP
A total of ~5 g sporoderm-broken spores of G.
lucidum were settled into 100 ml ultrapure water. The lipids
were removed and centrifuged at a speed of 1,000 x g at 70˚C for 12
h. Following this, the mixture was centrifuged at 1,200 x g for 15
min to remove the insoluble substances and purified using the
Sevage method (29). The solution
was then centrifuged (300 x g, 10 min, 4˚C) to collect the
supernatant. Finally, the supernatant was centrifuged (300 x g, 10
min, 4˚C) again and freeze-dried using a H051 freeze dryer
(LaboGene) to obtain GLP for subsequent experiments.
Groups and dosing
The animals were divided into five groups. The
6-+/db male mice were used as the wild-type group. The other 24
T2DM model mice were divided into the following groups: i) Model
control group; ii) low dose GLP group (GLP 100 mg/kg/day); iii)
high dose GLP group (GLP 400 mg/kg/day); and iii) melbine (MET,
Tianjin Zhongxin Pharmaceutical Group Co., Ltd.) group (MET 300
mg/kg/day, which was used as the positive control). A total of six
mice were allocated per group. GLP was dissolved in 0.5% sodium
carboxyl methyl cellulose (Shandong Senmei Biological Technology
Co., Ltd.) and MET was dissolved in water. Each animal was dosed by
oral gavage every day for 8 weeks and the daily dose volume was
adjusted according to the body weight of each mouse. Animal health
and behavior were monitored every day.
Glycosylated hemoglobin and clinical
chemistry analysis
Following 8 weeks of treatment, levels of
glycosylated hemoglobin (HbAlc) were measured with an analyzer
(Roche Diagnostics) using whole blood obtained from the tail vein
(0.08 ml/animal) from anesthetized mice (1-2% inhalant isoflurane).
Blood (~0.08 ml) was collected from the tail vein of each mouse.
Blood was centrifuged at 1,000 x g at 4˚C for 15 min. The serum was
collected and stored at -20˚C. Following this, the concentrations
of fasting blood glucose (FBG), alanine aminotransferase (ALT),
total cholesterol (TC), glutamine transaminase (TG), high-density
lipoprotein cholesterol (HDL-C), low-density lipoprotein
cholesterol (LDL-C), serum creatinine (Scr) and urea nitrogen (BUN)
serum levels were detected using a 7020 full automatic biochemical
analyzer (Hitachi, Ltd.). The concentrations of uric acid (UA),
uric creatinine (Ucr) and urine microalbumin (U-LAB) in the urine
of mice were detected by an automatic biochemical analyzer
(Automatic Biochemical Analyzer 7180; Hitachi, Ltd.).
Hematoxylin and eosin (H&E)
staining
Following dosing for 8 weeks, animals were
sacrificed by 20-25% CO2 euthanasia. Following this, the
veterinarian verified death based on respiratory, heartbeat, pupil
and nerve reflex indicators. The liver of each mouse was removed
and placed into a plate filled with pre-cooled normal saline. After
tissues were fixed in 4% paraformaldehyde at room temperature for
15 min, they were dehydrated in 50, 70, 80, 95, 100 and 100%
alcohol, for 20 min each time. Subsequently, the dehydrated tissue
was placed in the following solutions: Εthanol + xylene (1:1) for 2
h, xylene I and II for 10 min each, and then placed in xylene +
paraffin (1:1) for 2 h, paraffin I (1 h) and paraffin II (2 h) to
immerse. After embedding in an ice box, 4-µm paraffin slices were
made, which were then dewaxed with xylene and gradient alcohol, and
stained with hematoxylin and eosin at room temperature for 30 min
for observation under a light microscope (x400 magnification;
Olympus Corporation).
Oil Red O staining
The liver of each mouse was frozen at -20˚C and cut
into 5-10 µm sections. The slides were air dried for 30-60 min at
room temperature and fixed in ice cold 10% formalin for 5-10 min at
4˚C. Following this, the slides were air dried again for another
30-60 min and placed in absolute propylene glycol for 2-5 min to
avoid carrying water into the Oil Red O. The slides were stained in
pre-warmed Oil Red O solution for 8-10 min in a 60˚C oven and
differentiated in 85% propylene glycol for 2-5 min. Distilled water
were used to rinse the slides. Then, the slides were stained at
room temperature in Gill's hematoxylin for 30 sec and washed
thoroughly in running tap water for 3 min. Lastly, the slides were
placed in distilled water. Lipid droplets were observed under a
light microscope (x200 magnification; Olympus Corporation).
Immunohistochemistry
Liver tissues were embedded in paraffin (fixed in 4%
paraformaldehyde for 20 min at room temperature), sectioned (3-µm
thickness), blocked for 1 h with 2% normal horse serum
(Sigma-Aldrich; Merck KGaA) at room temperature and incubated with
FAM3C antibodies (1:500; cat. no. PA5-83654; Thermo Fisher
Scientific, Inc.). Tissues were incubated at 37˚C for 1-2 h and
washed twice for 5 min with PBS. A HRP-conjugated antibody
(1:2,000; cat. no. 31466; Thermo Fisher Scientific, Inc.) was added
and incubated for 30 min at room temperature. A total of 1 ml
3,3'-diaminobenzidine (DAB) Plus substrate (Sigma-Aldrich; Merck
KGaA) was mixed with 1-2 drops of DAB Plus Chromogen
(Sigma-Aldrich; Merck KGaA) and pipetted onto the slides. Following
incubation for 3-15 min, the slides were washed with water and
observed with a fluorescence microscope (x200 magnification;
Olympus Corporation).
Reverse transcription-quantitative PCR
(RT-qPCR) for miR-34a expression
Following dosing for 8 weeks, animals were
sacrificed by 20-25% CO2 euthanasia. The liver of each
mouse was removed, and total RNA was collected from the tissues
using an RNA Extraction kit (Takara Bio, Inc.), according to the
manufacturer's protocols. Extracted RNA was quantified using a
NanoDrop spectrophotometer (NanoDrop Technologies; Thermo Fisher
Scientific, Inc.). A cDNA Synthesis kit (Takara Biotechnology Co.,
Ltd.) was subsequently used to synthesize cDNA according to the
manufacturer's instructions. SYBR Premix Ex Taq™ (Takara
Bio, Inc.) with an Applied Bio-Rad CFX96 Sequence Detection system
(Applied Biosystems; Thermo Fisher Scientific, Inc.) was used for
RT-qPCR. The thermocycling conditions were as follows: Initial
denaturation at 92˚C for 4 min, followed by 40 cycles of 90˚C for
15 sec and 60˚C for 30 sec. The expression levels of FAM3C, HSF1
and CaM were determined by the threshold cycle and relative
expression levels were calculated using the 2-ΔΔCq
method (30). The expression of
GAPDH in the tissues was used as the negative control. Three
independent assays were performed. The primers for each protein are
presented in Table I.
 | Table ISequences of primers for FAM3C, HSF1,
CaM and GAPDH. |
Table I
Sequences of primers for FAM3C, HSF1,
CaM and GAPDH.
| Primer | Sequences
(5'-3') | Primer length
(bp) |
|---|
| FAM3C | F:
CCACCACAGAAGACCCAGTT | 20 |
| | R:
AACCAAACTCACGGATGAGG | 20 |
| HSF1 | F:
CCAGCAGCAAAAAGTTGTCA | 20 |
| | R:
CTGGTGACAGCATCAGAGGA | 20 |
| CaM | F:
GTTTGGGTGTGTGACTCTGG | 21 |
| | R:
GTTCTGTGAAATCTTCCGGG | 20 |
| GAPDH | F:
CAATGACCCCTTCATTGACC | 20 |
| | R:
GAGAAGCTTCCCGTTCTCAG | 20 |
Western blotting
The livers were isolated from the animals following
dosing for 8 weeks. A Nuclear and Cytoplasmic Protein Extraction
kit (Beyotime Institute of Biotechnology) was used to isolate
proteins from tissues. Protein concentration was determined using a
BCA kit (Guangzhou Youdi Biotechnology Co., Ltd.). Following this,
~30 µg/lane of protein was separated via SDS-PAGE on a 12% gel, and
subsequently transferred to a PVDF membrane (EMD Millipore). The
membrane was blocked with 5% non-fat dry milk in Tris-buffered
saline with 0.1% Tween-20 (pH 7.4) for 1 h at room temperature, and
then incubated overnight at room temperature with primary rabbit
anti-human antibodies for FAM3C (1:1,000; cat. no. ab56065; Abcam),
HSF1 (1:1,000; cat. no. ab61382; Abcam), CaM (1:1,000; cat. no.
ab52476; Abcam), phosphorylated (p)-AKT (1:1,000; cat. no. ab38449;
Abcam), AKT (1:1,000; cat. no. ab18785; Abcam) and GAPDH (1:1,000;
cat. no. ab8245; Abcam) purchased from Abcam. Membranes were then
incubated with HRP-conjugated antibody against rabbit IgG (1:5,000;
cat. no. ab6721; Abcam) at room temperature for 2 h. Blots were
incubated with enhanced chemiluminescence reagent (Beyotime
Institute of Biotechnology) and exposed on a Tanon 5200
Chemiluminescent Imaging System (Tanon Science & Technology
Co., Ltd.) to detect protein bands. Three independent assays were
performed. Protein bands were scanned and quantified using a
ChemiDoc MP Image analysis system (cat. no. 170-8280; Bio-Rad
Laboratories, Inc.).
Statistical analysis
Statistically significant differences for continuous
variables were determined using a one-way ANOVA followed by a
Dunnett's post hoc test for normally distributed data. All testing
was performed using GraphPad Prism software (version 5; GraphPad
Software, Inc.). P<0.05 was considered to indicate a
statistically significant difference. Three independent assays were
performed on each assay type.
Results
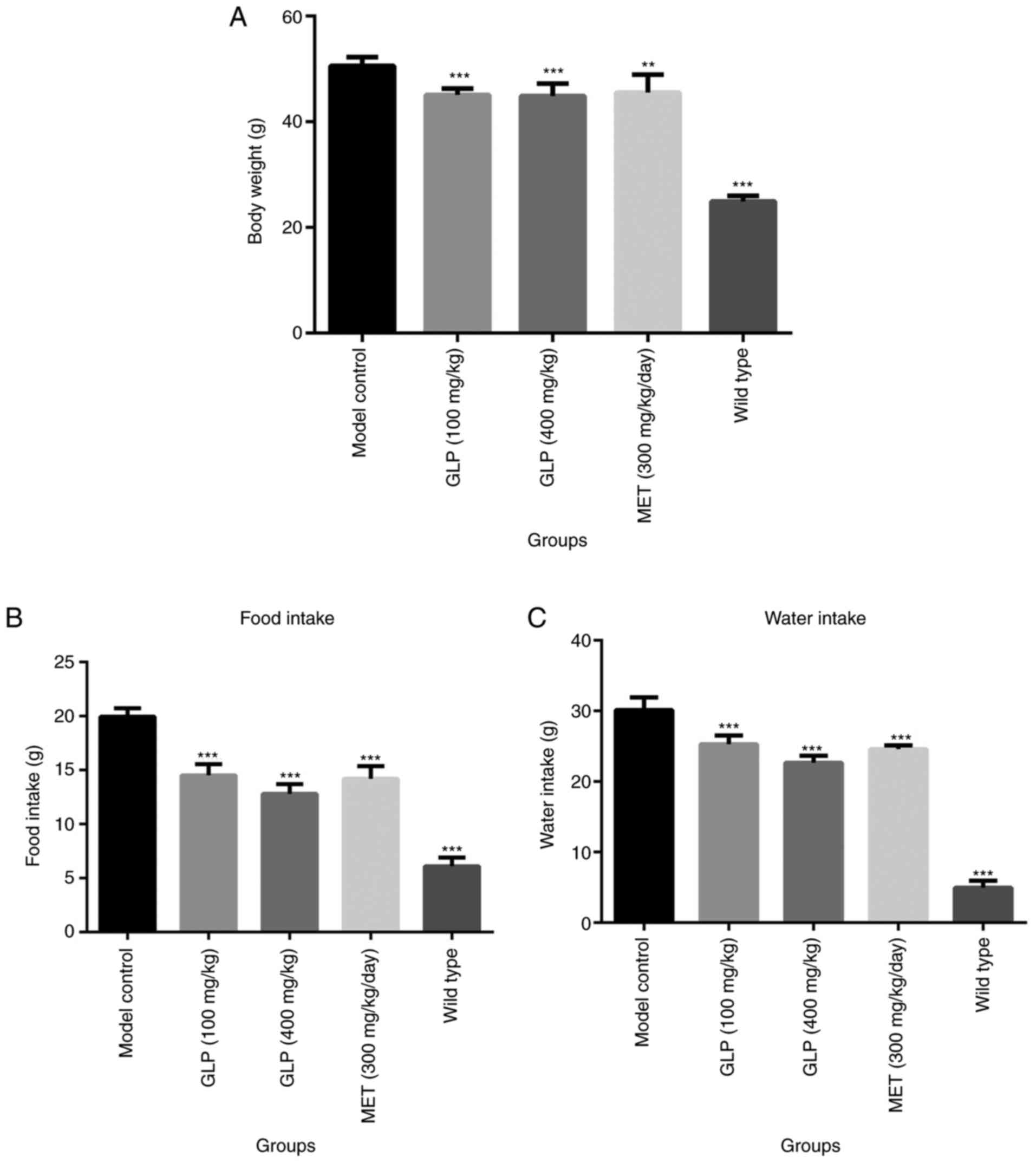
Body weight and food intake of T2DM
mice are influenced by GLP
The body weight of each mouse was recorded following
8 weeks of dosing. As presented in Fig.
1A, the body weights of the mice in the wild-type (P<0.001),
100 mg/kg GLP (P<0.001), 400 mg/kg GLP (P<0.001) and MET
(P<0.01) groups were all significantly lower compared with the
model control group. The results of food and water intake are
presented in Fig. 1B and C. Mice in the wild-type (P<0.001), 100
mg/kg GLP (P<0.001), 400 mg/kg GLP (P<0.001) and MET
(P<0.001) groups consumed significantly less food and water
compared with the model control group.
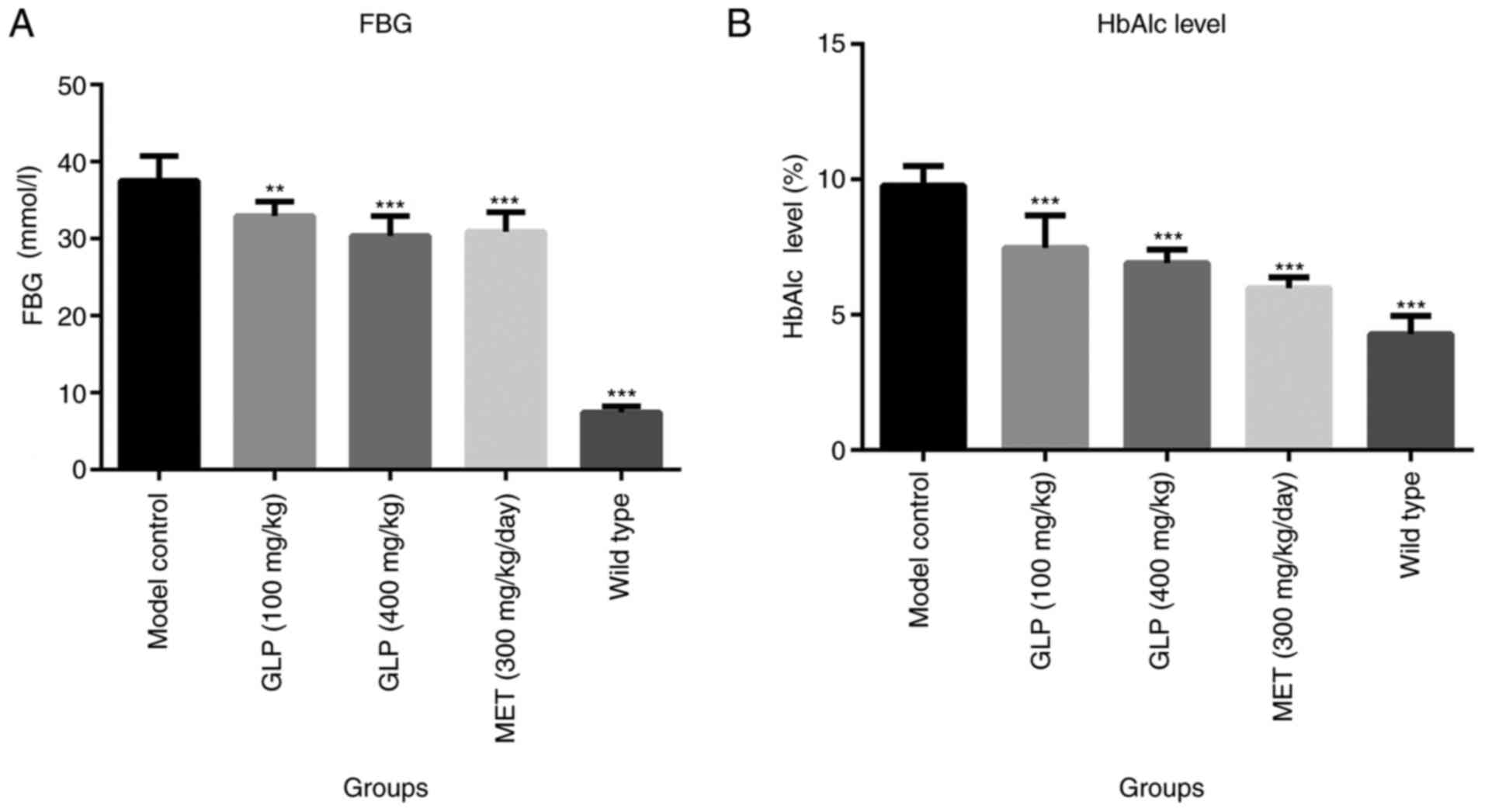
Effects of GLP on the concentration of
FBG and HbAlc levels of T2DM mice
The concentration of FBG was detected from tail vein
blood samples of T2DM mice. As demonstrated by Fig. 2A, the concentration of FBG in the
wild-type (P<0.001), 100 mg/kg GLP (P<0.01), 400 mg/kg GLP
(P<0.001) and MET (P<0.001) groups were all significantly
lower compared with the model control group. Furthermore, the level
of FBG in the 400 mg/kg GLP group was slightly lower compared with
the MET group. As presented in Fig.
2B, the HbAlc levels in the wild-type (P<0.001), GLP
(P<0.001) and MET groups (P<0.001) were significantly lower
compared with the model control group. Additionally, mice in the
400 mg/kg GLP group exhibited similar HbAlc levels to the MET
group.
Clinical chemistry of T2DM mice is
altered by GLP
Following collection of the blood samples after 8
weeks of treatment, the levels of different clinical chemistry
indexes were detected in the serum and urine. As shown in Fig. 3, the concentration of ALT, TC, TG,
LDL-C, Scr and BUN in the serum of mice in the wild-type
(P<0.001), GLP (P<0.01) and MET groups (P<0.01) were
significantly lower compared with the model control group.
Additionally, the concentration of HDL-C in the serum of mice in
the wild-type (P<0.001), GLP (P<0.001 or P<0.05) and MET
groups (P<0.01) was significantly higher compared with the model
control group. The levels of UA, Ucr and U-LAB in the urine of mice
in wild-type (P<0.001), GLP (P<0.001) and MET groups
(P<0.01) were significantly lower compared with the model
control group.
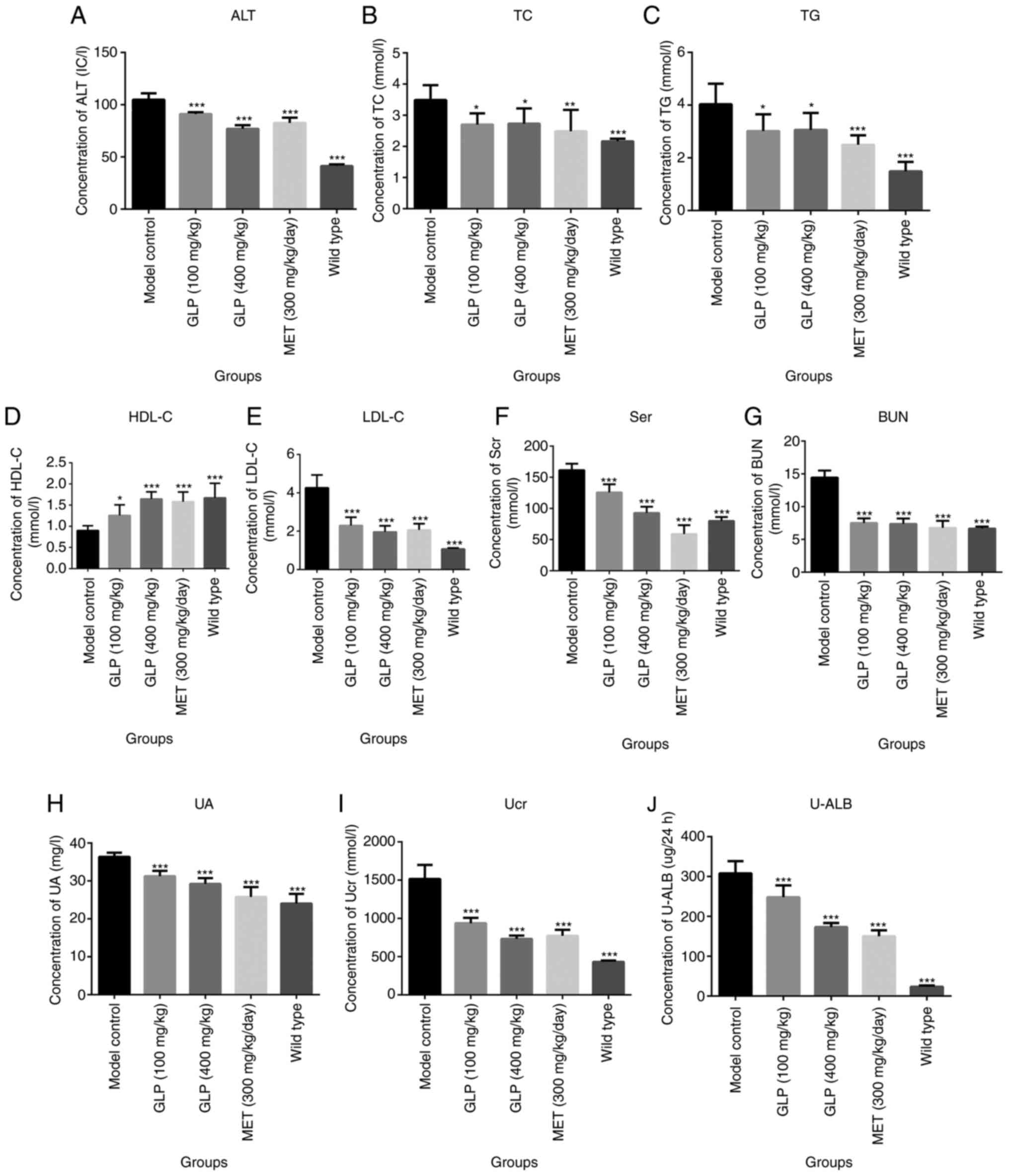 | Figure 3Clinical chemistry of type 2 diabetic
mice is altered by GLP. Concentrations of (A) ALT, (B) TC, (C) TG,
(D) HDL-C, (E) LDL-C, (F) Scr, (G) BUN, (H) UA, (I) Ucr and (J)
U-LAB. *P<0.05, **P<0.01 and
***P<0.001 vs. model control. ALT, alanine
aminotransferase; TC, total cholesterol; TG, glutamine
transaminase; HDL-C, high-density lipoprotein cholesterol; LDL-C,
low-density lipoprotein cholesterol; Scr, serum creatinine; BUN,
urea nitrogen; UA, uric acid; Ucr, uric creatinine; U-LAB, urine
microalbumin; GLP, Ganoderma lucidum polysaccharide; MET,
melbine. |
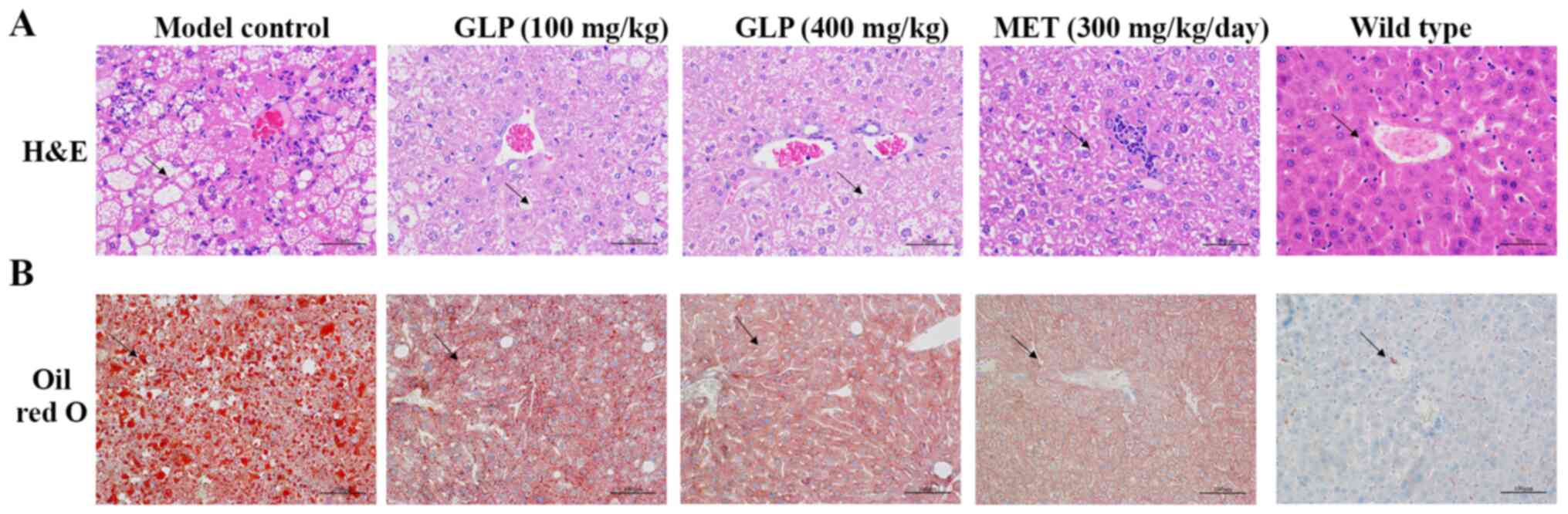
Hepatic steatosis in T2DM mice is
improved by GLP
H&E and Oil Red O staining were performed to
evaluate the effects of GLP on fatty liver in diabetic mice. The
results of light microscopy revealed that liver cells in the
wild-type group were arrayed neatly and there were almost no lipid
droplet vacuoles present (Fig. 4).
In contrast, numerous lipid droplets were observed in the
hepatocytes of the model control group. The volumes of the
hepatocyte increased. In the GLP and MET groups, the volume of the
lipid droplets decreased compared with the model group (Fig. 4B), indicating that the hepatic
steatosis of T2DM mice was improved following treatment with GLP or
MET.
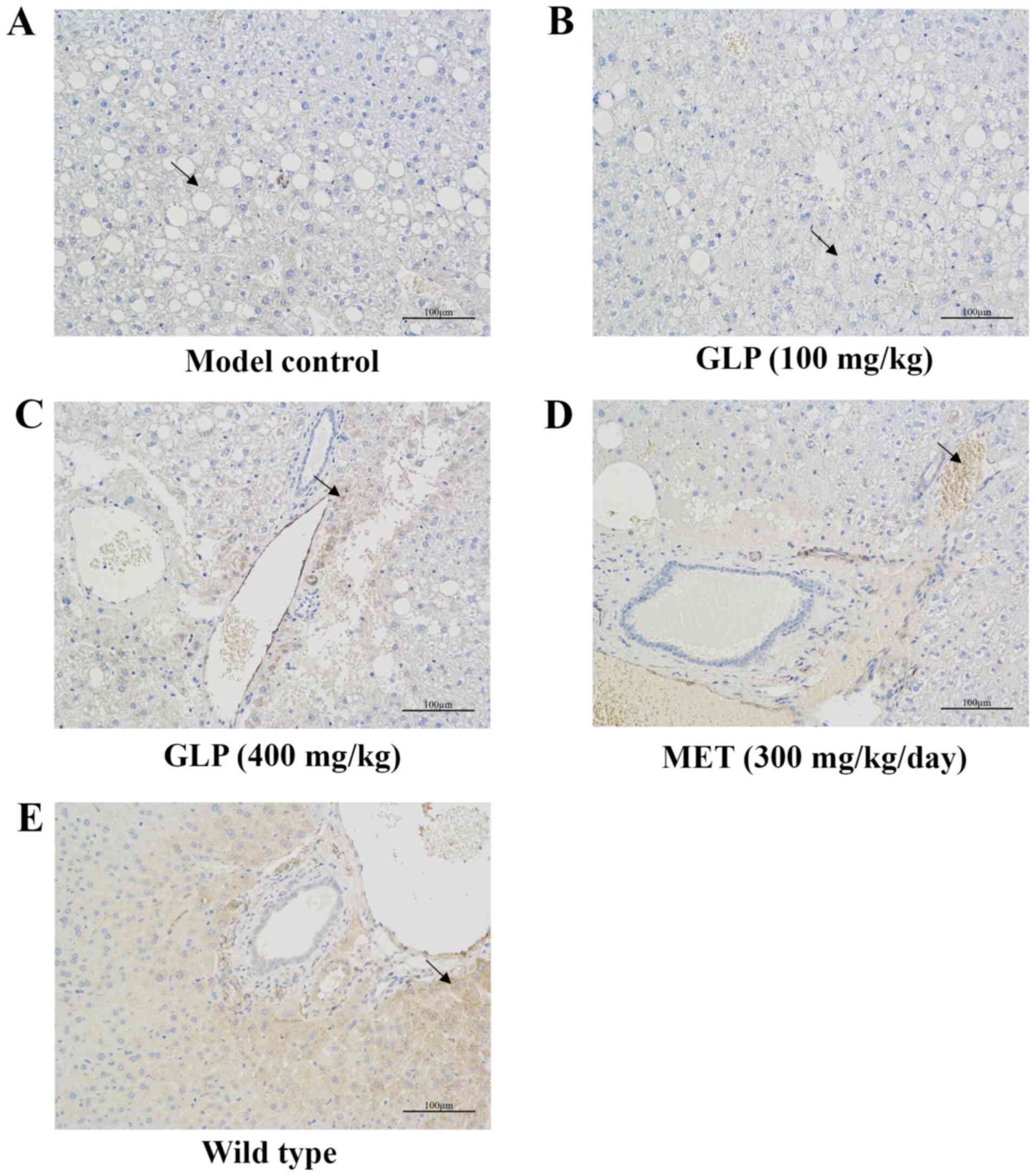
Effects of GLP on the expression
levels of the proteins FAM3C, HSF1, CaM and AKT in the liver of
T2DM mice as analyzed by immunohistochemistry, RT-qPCR and western
blotting
The results of immunohistochemistry are presented in
Fig. 5. In the images, brown
indicated FAM3C expression. The results indicated that the
expression of FAM3C was markedly higher in the livers of mice in
the wild-type, 400 mg/kg GLP and MET groups compared with model
control group, with no significant changes observed in the 100
mg/kg GLP group. Furthermore, the relative gene expression of
FAM3C, HSF1 and CaM in the liver of each mouse was detected by
RT-qPCR. The relative expression of FAM3C in the liver of mice in
the wild-type and the GLP groups (P<0.001 or P<0.05) was
significantly higher compared with the model control group
(Fig. 6). Notably, the expression
of FAM3C in the 400 mg/kg GLP group was higher compared with the
MET group. Additionally, HSF1 and CaM were highly expressed in the
liver of the 400 mg/kg GLP (P<0.001) and MET (P<0.001) groups
compared with the model group. Subsequently, the expression levels
of the proteins FAM3C, HSF1, CaM, AKT and p-AKT were detected by
western blotting. As demonstrated in Fig. 7, the expression levels of FAM3C,
HSF1, CaM and p-AKT/AKT in the liver of mice in the wild-type
(P<0.001), the 400 mg/kg GLP (P<0.001) and MET (P<0.001)
groups were significantly higher compared to the model control
group, with slightly or no significant changes observed in the 100
and 400 mg/kg GLP groups.
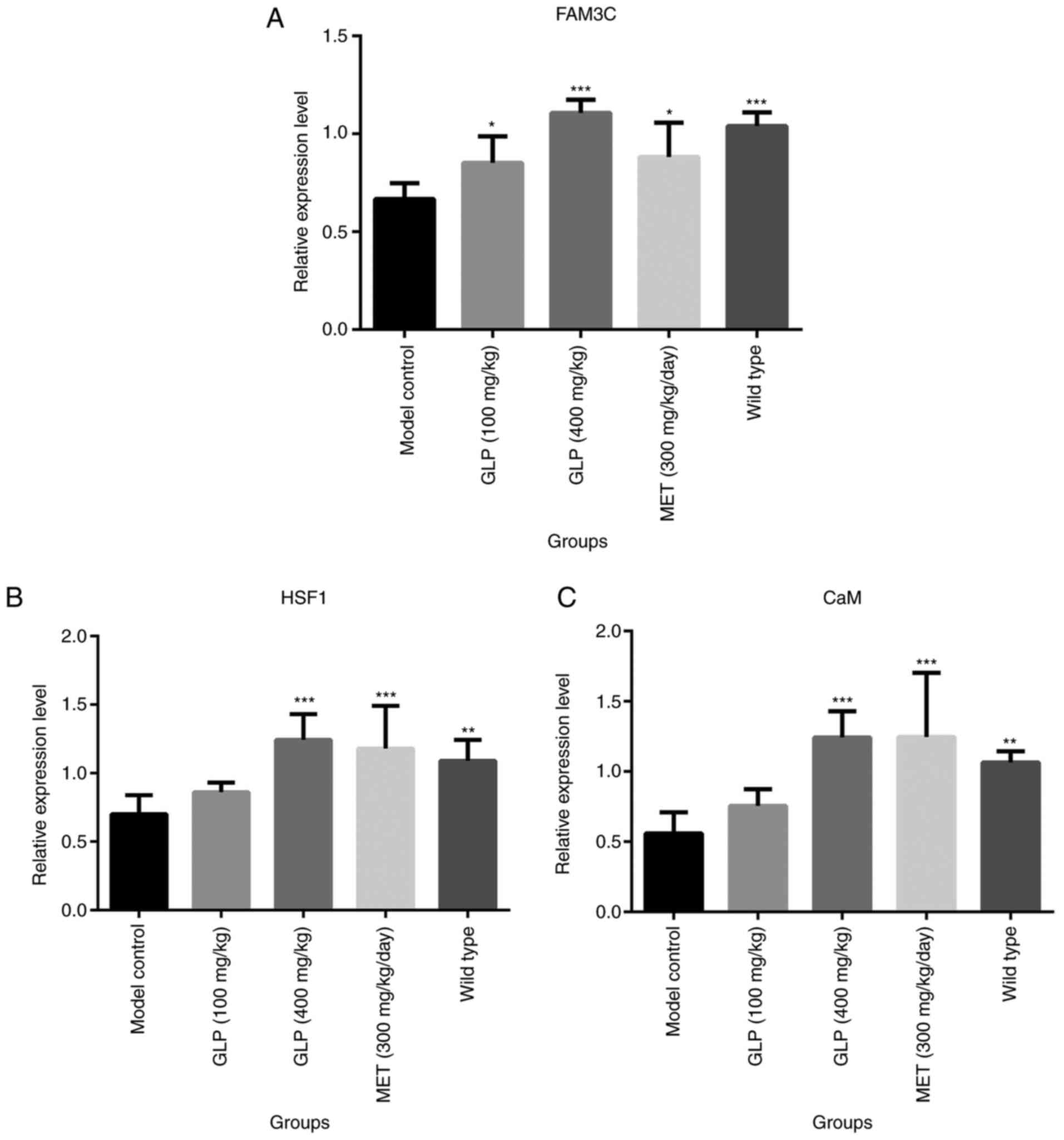 | Figure 6Effects of GLP on the expression of
FAM3C, HSF1, and CaM in liver tissues. Relative expression levels
of (A) FAM3C, (B) HSF1 and (C) CaM for each group, as determined by
reverse transcription-quantitative PCR. *P<0.05,
**P<0.01 and ***P<0.001 vs. model
control. FAM3C, family with sequence similarity 3; HSF1, heat shock
factor 1; CaM, calmodulin; GLP, Ganoderma lucidum
polysaccharide; MET, melbine. |
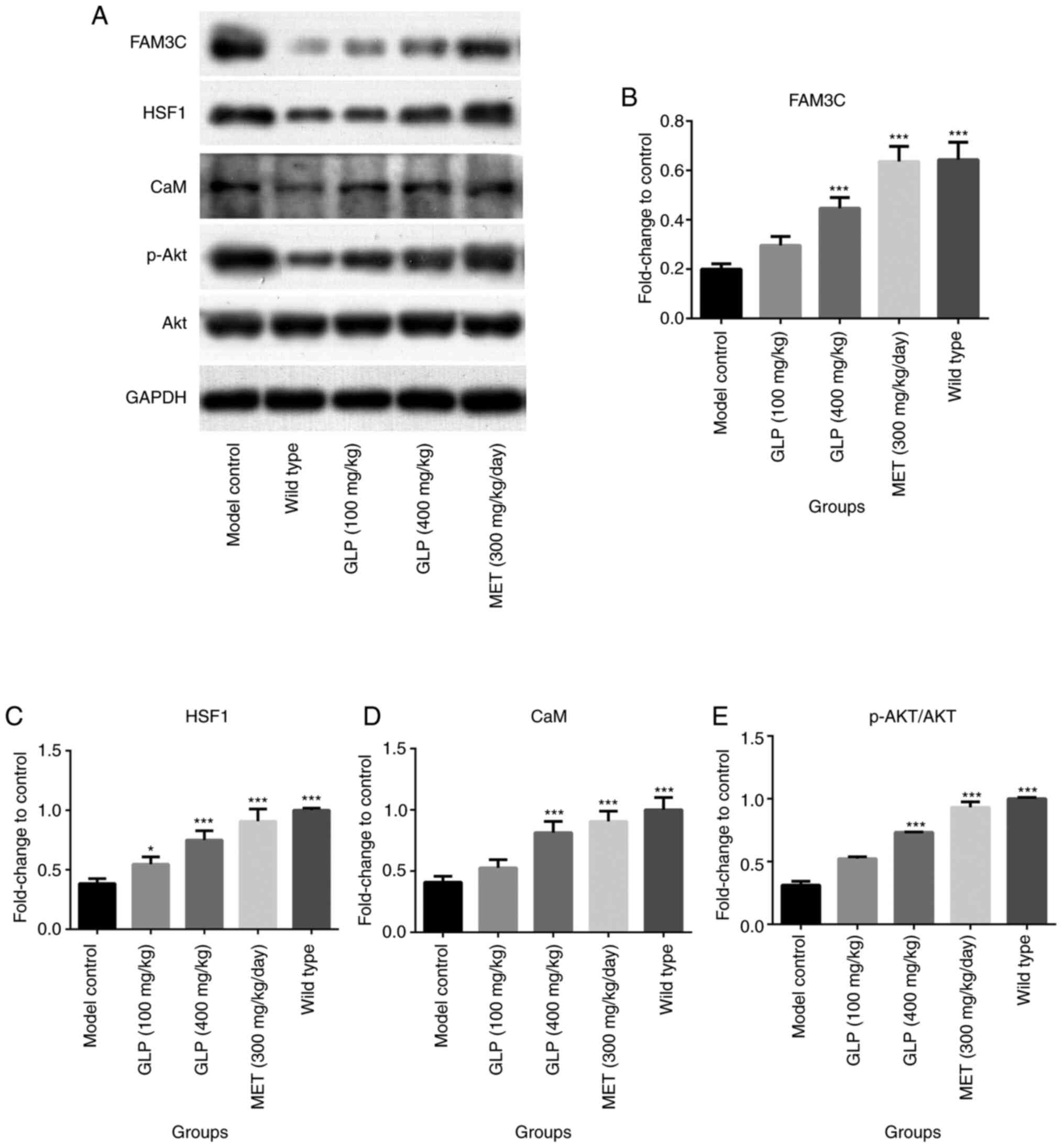 | Figure 7Effects of GLP on the levels of
FAM3C, HSF1, CaM, AKT and p-AKT in liver tissues. Expression levels
of the proteins (A) FAM3C, HSF1, CaM, AKT and p-AKT in the livers
of T2DM mice, as analyzed by western blotting. (B-E) Levels of each
protein, as quantified by gray scale analysis.
*P<0.05 and ***P<0.001 vs. model
control. FAM3C, family with sequence similarity 3; HSF1, heat shock
factor 1; CaM, calmodulin; p-, phosphorylated; GLP, Ganoderma
lucidum polysaccharide; MET, melbine. |
Discussion
G. lucidum is a mushroom that is used in
Traditional Chinese Medicine to enhance the immune system, regulate
blood glucose and control blood pressure (31). GLP was reported to be one of the
main ingredients extracted from G. lucidum (32,33).
Furthermore, it has been reported that GLP significantly decreased
the concentration of blood glucose, promoted insulin secretion,
improved glucose tolerance and regulated the concentration of blood
lipids (34). The results of the
present study demonstrated that following dosing with 100 and 400
mg/kg GLP, the FBG, HbAlc and certain diabetes-related clinical
chemistry indexes (ALT, TC, TG, LDL-C, Scr, BUN, UA, Ucr and U-LAB)
in T2DM mice were significantly improved compared with the model
control group. These results were consistent with previous reports
(35,36). Diabetes-associated symptoms were
reported to be significantly improved in diabetic rats by oral
dosing with GLP (37). The
concentrations of blood glucose and triglyceride and cholesterol in
the serum were decreased following dosing for 3 months (38). Additionally, Ma et al
(39) reported that blood glucose
levels and diabetic-related morbidity were significantly decreased
in multiple low-dose STZ-induced diabetic mice following dosing
with GLP. In the current study, the results of H&E and Oil Red
O staining data demonstrated that the lipid droplets in the
hepatocytes of mice shrank in the GLP groups compared with the
model control group, which indicated that fatty liver induced by
diabetes was significantly improved by GLP treatment. The present
study was accompanied with a positive control, MET, which has been
used to effectively treat T2DM (40). Notably, nearly all the indexes
measured showed that 400 mg/kg GLP exerted a similar anti-diabetic
effect as 300 mg/kg MET. The current study used a T2DM model to
evaluate the anti-diabetic effect of GLP and its effect on the
FAM3C-HSF1-CAM signaling pathway. Further verification of the
anti-diabetic effects of GLP is being investigated using different
types of animal models by the current authors, including GK/IRS-1
double transgenic mice and MKR transgenic mice.
It has been reported that in STZ-induced diabetic
mice, the expression of FAM3C is significantly decreased, which
results in the inactivation of the HSF1-CaM-AKT signaling pathway.
This increases the expression of the glycogen gene, and
gluconeogenesis and lipogenesis in the liver become dysregulated,
resulting in diabetes (27,28). In the present study, following
dosing with GLP for 8 weeks, the expression of FAM3C, HSF1, CaM,
AKT and p-AKT in the hepatocytes of mice were detected with
immunohistochemistry, RT-qPCR and western blotting. The results
indicated that the FAM3C-HSF1-CaM-AKT signaling pathway was
significantly activated following GLP treatment in T2DM mice, which
may explain the significant anti-diabetic effect of GLP.
Additionally, the results indicated that MET significantly
activated the FAM3C-HSF1-CaM-AKT signaling pathway to exert an
anti-diabetic effect. However, based on the current results, it is
difficult to ascertain that the effect of GLP is directly
associated with FAM3C-HSF1-CaM signaling. Fully understanding the
mechanism of the anti-diabetic effect of GLP is an aim of the
current authors. Further detailed investigation on the association
between the anti-diabetic effects of GLP and the FAM3C-HSF1-CaM
signaling pathway should be performed to elucidate the underlying
mechanism in the future, including investigations using
insulin-resistant in vitro models.
GLPs have previously been demonstrated to exert
anti-diabetic effects in multiple reports. Chen et al
(23) reported that GLPs
effectively lowered blood glucose levels and protected aortas in
rats. The underlying mechanism may be involved in the
downregulation of advanced glycation end-products and receptor for
advanced glycation end-products in aortal tissue. Additionally, it
has been reported that GLPs significantly decrease fasting serum
glucose levels in T2DM mice in a dose-dependent manner (36). The decrease in fasting serum glucose
levels may be associated with decreased mRNA expression level of
several key enzymes, such as hexokinase, involved in
gluconeogenesis and/or glycogenolysis (41). Furthermore, Xiao et al
(42) demonstrated that the
anti-diabetic effect of GLPs may be associated with the
downregulation of hepatic glucose-regulated enzyme mRNA levels via
AMPK activation. However, whether a diabetic-associated signaling
pathway was involved in the anti-diabetic effect of GLP remains
unknown.
In the present study, the significant anti-diabetic
effect of GLPs was verified and the FAM3C-HSF1-CaM signaling
pathway was revealed to be associated with this mechanism. Although
further, direct verifications are required, these results reported
a novel direction for the future investigation of the mechanism of
the anti-diabetic effect of GLPs. In conclusion, the present study
demonstrated that GLP exerted a significant effect on lipid
metabolism dysregulation in diabetic model mice, which may be
associated with the activation of the FAM3C-HSF1-CaM signaling
pathway.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
RP was involved in drafting the manuscript and data
analysis. JL and LW assisted with the acquisition of data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The current study declares that all animal
experiments were authorized by the Ethical Committee of Zhejiang
Chinese Medical University (Hangzhou, China) and were performed
according to the guidelines for the Care and Use of Laboratory
Animals and to the Principles of Laboratory Animal Care and
Protection.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhu J, Zhang X, Zhang X, Dong M, Wu J,
Dong Y, Chen R, Ding X, Huang C, Zhang Q and Zhou W: The burden of
ambient air pollution on years of life lost in Wuxi, China,
2012-2015: A time-series study using a distributed lag non-linear
model. Environ Pollut. 224:689–697. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Patil P, Mandal S, Tomar SK and Anand S:
Food protein-derived bioactive peptides in management of type 2
diabetes. Eur J Nutr. 54:863–880. 2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Garber AJ, Abrahamson MJ, Barzilay JI,
Blonde L, Bloomgarden ZT, Bush MA, Dagogo-Jack S, DeFronzo RA,
Einhorn D, Fonseca VA, et al: Consensus statement by the American
Association of clinical endocrinologists and American college of
endocrinology on the comprehensive type 2 diabetes management
algorithm-2019 executive summary. Endocr Pract. 25:69–100.
2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
He K, Shi JC and Mao XM: Safety and
efficacy of acarbose in the treatment of diabetes in Chinese
patients. Ther Clin Risk Manag. 10:505–511. 2014.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Abe M, Okada K, Maruyama T, Maruyama N and
Matsumoto K: Combination therapy with mitiglinide and voglibose
improves glycemic control in type 2 diabetic patients on
hemodialysis. Expert Opin Pharmacother. 11:169–176. 2010.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Kirschner LS, Qamri Z, Kari S and Ashtekar
A: Mouse models of thyroid cancer: A 2015 update. Mol Cell
Endocrinol. 421:18–27. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ali MY, Zaib S, Rahman MM, Jannat S, Iqbal
J, Park SK and Chang MS: Didymin, a dietary citrus flavonoid
exhibits anti-diabetic complications and promotes glucose uptake
through the activation of PI3K/Akt signaling pathway in
insulin-resistant HepG2 cells. Chem Biol Interact. 305:180–194.
2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Asano N, Yamashita T, Yasuda K, Ikeda K,
Kizu H, Kameda Y, Kato A, Nash RJ, Lee HS and Ryu KS:
Polyhydroxylated alkaloids isolated from mulberry trees (Morus
alba L.) and silkworms (Bombyx mori L.). J Agric Food
Chem. 49:4208–4213. 2001.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Chehade JM and Mooradian AD: A rational
approach to drug therapy of type 2 diabetes mellitus. Drugs.
60:95–113. 2000.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Lorenzo PI, Juárez-Vicente F,
Cobo-Vuilleumier N, Garcia-Dominguez M and Gauthier BR: The
diabetes-linked transcription factor PAX4: From gene to functional
consequences. Genes (Basel). 8(101)2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Omori K, Kobayashi E, Rawson J, Takahashi
M and Mullen Y: Mechanisms of islet damage mediated by pancreas
cold ischemia/rewarming. Cryobiology. 73:126–134. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Beattie GM, Leibowitz G, Lopez AD, Levine
F and Hayek A: Protection from cell death in cultured human fetal
pancreatic cells. Cell Transplant. 9:431–438. 2000.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Xue YF, Guo CZ, Hu F, Sun DM, Liu JH and
Mao SY: Molecular mechanisms of lipid metabolism disorder in livers
of ewes with pregnancy toxemia. Animal. 13:992–999. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Kaneko YK: Development and analysis of
novel therapeutic targets to improve pancreatic β-cell function in
type 2 diabetes. Yakugaku Zasshi. 136:1623–1629. 2016.PubMed/NCBI View Article : Google Scholar : (In Japanese).
|
|
15
|
Langley AK, Suffoletta TJ and Jennings HR:
Dipeptidyl peptidase IV inhibitors and the incretin system in type
2 diabetes mellitus. Pharmacotherapy. 27:1163–1180. 2007.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Somtimuang C, Olatunji OJ and Ovatlarnporn
C: Evaluation of in vitro α-amylase and α-glucosidase inhibitory
potentials of 14 medicinal plants constituted in thai folk
antidiabetic formularies. Chem Biodivers.
15(e1800025)2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Wu H, Liu J, Lou Q, Liu J, Shen L, Zhang
M, Lv X, Gu M and Guo X: Comparative assessment of the efficacy and
safety of acarbose and metformin combined with premixed insulin in
patients with type 2 diabetes mellitus. Medicine (Baltimore).
96(e7533)2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Xu J and Rajaratnam R: Cardiovascular
safety of non-insulin pharmacotherapy for type 2 diabetes.
Cardiovasc Diabetol. 16(18)2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Laoud A, Ferkous F, Maccari L, Maccari G,
Saihi Y and Kraim K: Identification of novel nt-MGAM inhibitors for
potential treatment of type 2 diabetes: Virtual screening, atom
based 3D-QSAR model, docking analysis and ADME study. Comput Biol
Chem. 72:122–135. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Men P, Li XT, Tang HL and Zhai SD:
Efficacy and safety of saxagliptin in patients with type 2
diabetes: A systematic review and meta-analysis. PLoS One.
13(e0197321)2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Min SH, Yoon JH, Hahn S and Cho YM:
Efficacy and safety of combination therapy with an α-glucosidase
inhibitor and a dipeptidyl peptidase-4 inhibitor in patients with
type 2 diabetes mellitus: A systematic review with meta-analysis. J
Diabetes Investig. 9:893–902. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Chiefari E, Arcidiacono B, Foti D and
Brunetti A: Gestational diabetes mellitus: An updated overview. J
Endocrinol Invest. 40:899–909. 2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Chen Y, Qiao J, Luo J, Wu F, Meng G, Chen
H, Zheng H and Xu J: Effects of Ganoderma lucidum
polysaccharides on advanced glycation end products and receptor of
aorta pectoralis in T2DM rats. Zhongguo Zhong Yao Za Zhi.
36:624–627. 2011.PubMed/NCBI(In Chinese).
|
|
24
|
Teng BS, Wang CD, Yang HJ, Wu JS, Zhang D,
Zheng M, Fan ZH, Pan D and Zhou P: A protein tyrosine phosphatase
1B activity inhibitor from the fruiting bodies of Ganoderma
lucidum (Fr.) Karst and its hypoglycemic potency on
streptozotocin-induced type 2 diabetic mice. J Agric Food Chem.
59:6492–6500. 2011.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Tamez-Pérez HE, Gonzalez-Guajardo EE and
Tamez-Peña AL: Re: Consensus statement by the American association
of clinical endocrinologists and American college of endocrinology
on the comprehensive type 2 diabetes management algorithm-2019
executive summary. Endocr Pract. 25(622)2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Shen L, Ao L, Xu H, Shi J, You D, Yu X, Xu
W, Sun J and Wang F: Poor short-term glycemic control in patients
with type 2 diabetes impairs the intestinal mucosal barrier: A
prospective, single-center, observational study. BMC Endocr Disord.
19(29)2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Zhang X, Yang W, Wang J, Meng Y, Guan Y
and Yang J: FAM3 gene family: A promising therapeutical target for
NAFLD and type 2 diabetes. Metabolism. 81:71–82. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chen Z, Wang J, Yang W, Chen J, Meng Y,
Feng B, Chi Y, Geng B, Zhou Y, Cui Q and Yang J: FAM3C activates
HSF1 to suppress hepatic gluconeogenesis and attenuate
hyperglycemia of type 1 diabetic mice. Oncotarget. 8:106038–106049.
2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zhao X, Li J, Liu Y, Wu D, Cai P and Pan
Y: Structural characterization and immunomodulatory activity of a
water soluble polysaccharide isolated from Botrychium
ternatum. Carbohydr Polym. 171:136–142. 2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Liu Y, Zhang C, Du J, Jia R, Cao L, Jeney
G, Teraoka H, Xu P and Yin G: Protective effect of Ganoderma
lucidum polysaccharide against carbon tetrachloride-induced
hepatic damage in precision-cut carp liver slices. Fish Physiol
Biochem. 43:1209–1221. 2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Rashad FM, Kattan MHE, Fathy HM, El-Fattah
DAA, Tohamy ME and Farahat AA: Recycling of agro-wastes for
Ganoderma lucidum mushroom production and Ganoderma
post mushroom substrate as soil amendment. Waste Manag. 88:147–159.
2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wan Y, Wang Q and Prud'homme GJ: GABAergic
system in the endocrine pancreas: A new target for diabetes
treatment. Diabetes Metab Syndr Obes. 8:79–87. 2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Singh P, Jayaramaiah RH, Agawane SB,
Vannuruswamy G, Korwar AM, Anand A, Dhaygude VS, Shaikh ML, Joshi
RS, Boppana R, et al: Potential dual role of eugenol in inhibiting
advanced glycation end products in diabetes: Proteomic and
mechanistic insights. Sci Rep. 6(18798)2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Xue H, Qiao J, Meng G, Wu F, Luo J, Chen
H, Zheng H and Xu J: Effect of Ganoderma lucidum
polysaccharides on hemodynamic and antioxidation in T2DM rats.
Zhongguo Zhong Yao Za Zhi. 35:339–343. 2010.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
36
|
Li HN, Zhao LL, Zhou DY and Chen DQ:
Ganoderma lucidum polysaccharides ameliorates hepatic
steatosis and oxidative stress in db/db mice via targeting nuclear
factor E2 (erythroid-derived 2)-related factor-2/heme oxygenase-1
(HO-1) pathway. Med Sci Monit. 26(e921905)2020.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Chen M, Xiao D, Liu W, Song Y, Zou B, Li
L, Li P, Cai Y, Liu D, Liao Q and Xie Z: Intake of Ganoderma
lucidum polysaccharides reverses the disturbed gut microbiota
and metabolism in type 2 diabetic rats. Int J Biol Macromol.
155:890–902. 2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Chang CJ, Lin CS, Lu CC, Martel J, Ko YF,
Ojcius DM, Tseng SF, Wu TR, Chen YY, Young JD and Lai HC:
Ganoderma lucidum reduces obesity in mice by modulating the
composition of the gut microbiota. Nat Commun.
6(7489)2015.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Ma HT, Hsieh JF and Chen ST: Anti-diabetic
effects of Ganoderma lucidum. Phytochemistry. 114:109–113.
2015.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Adeyemi WJ, Olayaki LA, Abdussalam TA,
Fabiyi TO, Raji TL and Adetunji AA: Co-administration of omega-3
fatty acids and metformin showed more desirable effects than the
single therapy on indices of bone mineralisation but not
gluco-regulatory and antioxidant markers in diabetic rats. Biomed
Pharmacother. 121(109631)2020.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Xiao C, Wu QP, Cai W, Tan JB, Yang XB and
Zhang JM: Hypoglycemic effects of Ganoderma lucidum
polysaccharides in type 2 diabetic mice. Arch Pharm Res.
35:1793–1801. 2012.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Xiao C, Wu Q, Zhang J, Xie Y, Cai W and
Tan J: Antidiabetic activity of Ganoderma lucidum
polysaccharides F31 down-regulated hepatic glucose regulatory
enzymes in diabetic mice. J Ethnopharmacol. 196:47–57.
2017.PubMed/NCBI View Article : Google Scholar
|