Introduction
Spinal cord injury (SCI) often results in permanent
disability, placing a subsequent emotional and financial burden on
patients, families and society (1,2).
Currently, SCI therapy includes corticosteroidal drugs, relieving
excessive spinal cord (SC) pressure and pain and rehabilitation
training; however, overcoming the limited self-repair capacity of
the SC has proven challenging (3,4). The
regeneration and functional recovery of the SC requires the
precisely regulated induction of multiple cellular processes,
including regrowth of injured axons, production of new axons,
re-establishment of precise synaptic connections and restoration of
axonal conduction by electrophysiological recovery and
remyelination (5). Thus, the
development of strategies for inducing neural regeneration
following SCI is a major aim of clinical neuroscience.
The mammalian SC possesses a population of
endogenous neural stem cells (NSCs) (6) that can differentiate into neurons,
astrocytes and oligodendrocytes in response to specific factors in
the SCI microenvironment and form new synaptic connections to
re-establish nerve transmission pathways (7,8). In
addition to axonal regrowth and repair following SCI,
re-establishment of myelination by oligodendrocytes is essential
for functional recovery (9).
Therefore, repair strategies utilizing NSCs must induce efficient
oligodendrocyte differentiation, axonal repair and/or the
production of new neurons.
Oscillating field stimulation (OFS) is considered a
promising therapeutic strategy for neural regeneration and has
demonstrated modest clinical efficacy (10). However, the underlying mechanisms
are still unknown, which hinders optimization for broader clinical
application. Previous research has shown that electric field
stimulation regulates Wnt protein expression and Wnt signaling
pathways (11). Wnt signaling
regulates NSC differentiation, which indicates that OFS-induced
modulation of Wnt signaling may be a potential therapeutic strategy
for SCI (12). However, whether OFS
can promote neurological recovery following SCI by regulating the
differentiation of endogenous NSCs has not been previously
reported.
In the current study, the capacity of OFS to enhance
endogenous NSC differentiation and promote functional recovery was
examined in a rat model of SCI. The present study aimed to provide
a theoretical basis for assessing the clinical value of OFS in the
treatment of SCI.
Materials and methods
Animals
A total of 72 adult female Sprague-Dawley rats (age,
~8 weeks; weight, 220±10 g) were purchased from Anhui Medical
University. The rats were raised in an animal room under controlled
ambient temperature (26˚C) and 60 % relative humidity on a 12-h
light/dark cycle with ad libitum access to food and water.
The rats were randomly allocated to three groups with 24 rats in
each group: Sham controls, SCI alone and SCI + OFS. The Sham group
underwent laminectomy without SCI surgery. The SCI and SCI + OFS
groups were subjected to laminectomy and experimental SCI. Rats in
the SCI group experienced the implantation of the stimulator,
though this was not activated so they did not receive OFS, while
rats in the SCI + OFS group underwent OFS as the intervention. All
surgical procedures and experiments were performed in accordance
with the National Institutes of Health Guide for the Care and Use
of Laboratory Animals (NIH Publications no. 80-23; revised 1996)
(13) and were approved by the
Animal Ethics Committee of Anhui Medical University (approval
reference no. LLSC20190736).
OFS device
The OFS device was provided by the Beijing Key
Laboratory of Bioelectromagnetics, Institute of Electrical
Engineering, Chinese Academy of Sciences. The parameters of
functionality, implantability and biocompatibility were considered
when selecting a device, so as to have the least negative impact on
the animals. The device was powered by a 3.0-V primary battery
(cat. no. CR1220; Panasonic Corporation) with an output current of
40 µA and an electric field strength of 400 µV/mm between positive
and negative electrodes. The structure and working mechanism of the
OFS device are described in a previous study (14).
Experimental SCI
The rats were anesthetized via intraperitoneal
injection of 300 mg/kg chloral hydrate and then fixed on an
operating table. Following hair removal from the back and skin
disinfection, a 3-cm longitudinal incision was made at the dorsal
midline overlying the lower thoracic (T) vertebrae. The SC at
T9-T10 was exposed by blunt dissection of the paravertebral muscles
and removal of spinous processes and laminae, while carefully
avoiding additional tissue damage. A 10 g New York University
Multicenter Animal Spinal Cord Injury Study impactor was used to
model SCI by striking the center of the exposed SC vertically from
a height of 3 cm (15).
Furthermore, in the SCI + OFS and SCI groups, OFS device electrodes
(25 mm x 7 mm) were sutured in the intervertebral muscles above and
below the injury site and the device was stitched to the back
epidermis. In all groups, the wounds were sutured layer-by-layer
using 4-0 silk.
Body temperature was maintained at 37˚C during the
operation using a heating lamp. All animals were subcutaneously
injected with 105 U/kg penicillin G once per day for 3
days post-surgery. No signs of peritonitis were observed in any
group following surgery. Manual bladder emptying was performed by
squeezing of the bladder every 8 h until rats resumed autonomous
voiding.
Behavioral analysis
The Basso-Beattie-Bresnahan (BBB) score is a widely
accepted tool for assessing recovery of motor function following
SCI (16). Rats were scored
multiple times a day at the indicated time points by observing
motor function and co-ordination of hind limbs on days 7, 14, 21,
28 and 35 post-surgery. Each rat was evaluated by two independent
observers, who were blinded to the animals' group assignment, and
the mean BBB scores were used in the final analysis.
Double immunofluorescence (IF)
staining and microscopy
From each group, 4 rats were sacrificed on days 3, 7
and 14 post-surgery with excessive pentobarbital sodium anesthesia
(150 mg/kg) and a 1-cm SC section centered at the injury site was
removed. Excised spinal tissue was fixed in 4% paraformaldehyde at
room temperature for 12 h, embedded in paraffin and cut into serial
10-µm thick transverse sections. Tissue sections were immersed in
EDTA (cat. no. G1206; Servicebio Technology, Co., Ltd.) at 60˚C for
25 min to promote antigen repair. After rinsing in PBS, sections
were blocked in 5 % bovine serum albumin (BSA; G5001; Servicebio
Technology Co., Ltd.) at room temperature for 30 min and then
incubated with primary antibodies against β-tubulin III (1:200;
cat. no. T8578; Sigma-Aldrich; Merck KGaA) and neuron-glial antigen
2 (NG2; 1:200; cat. no. ab50009; Abcam) in QuickBlock™ primary
antibody dilution buffer (cat.no. P0262; Beyotime Institute of
Biotechnology) at 4˚C overnight. Each section was washed with PBS
three times and incubated with horseradish peroxidase-labeled goat
anti-mouse IgG (1:200; cat. no. GB23301; Servicebio Technology,
Co., Ltd.) in QuickBlock secondary antibody dilution buffer for
immunofluorescence (cat. no. p0265; Beyotime Institute of
Biotechnology) for 1 h at room temperature. Sections were stained
with FITC-conjugated donkey anti-goat IgG (1:100; cat. no. GB22404;
Servicebio Technology, Co., Ltd.) in Quickblock secondary antibody
dilution buffer at room temperature for 10 min and then immersed in
EDTA and heated in a microwave oven to remove the primary and
secondary antibodies combined on the tissue. Subsequently, sections
were incubated with the second primary antibody against Nestin
(1:100; cat. no. ab6142; Abcam) at 4˚C in Quickblock primary
antibody dilution buffer overnight and stained with Cy3-conjugated
goat anti-mouse IgG (1:200; cat. no. GB21301; Servicebio
Technology, Co., Ltd.) for 1 h at room temperature. Finally, nuclei
were counterstained with DAPI (cat. no. G1012; Servicebio
Technology, Co., Ltd.) for 10 min at room temperature. Images were
acquired using a fluorescent microscope (Eclipse C1; Nikon
Corporation) at x400 magnification. Each group had 36 sections, and
3 visual fields were randomly selected from each section and
quantified using Image-Pro Plus software (version 6.0; Media
Cybernetics, Inc.).
Hematoxylin and eosin (H&E)
staining
Rats from all experimental groups were sacrificed at
14 days after surgery and a 1-cm segment of SC section centered on
the site of injury was removed. Tissues were immediately fixed in
4% paraformaldehyde at room temperature for 24 h, followed by
paraffin embedding. Tissues from all groups were cut into 5-µm
thick transverse sections for H&E staining as follows: Sections
were stained with Hematoxylin (cat. no. H8070; Beijing Solarbio
Science & Technology Co., Ltd.) at room temperature for 5 min
and washed with distilled water for 5 min. Then, tissue sections
were immersed in 1% hydrochloric acid alcohol at room temperature
for 5 sec and washed with distilled water for 3 min. Each section
was immersed in 0.5% ammonia (cat. no. G1821; Beijing Solarbio
Science & Technology Co., Ltd.) at room temperature for 10 min
and washed with distilled water for 5 min. Subsequently, sections
were stained with Eosin Y solution (cat. no. G1100; Beijing
Solarbio Science & Technology Co., Ltd.) at room temperature
for 3 min. From each group 12 sections were stained and observed
using a fluorescent microscope (Eclipse E100; Nikon Corporation) at
x100 magnification for morphological changes (3 visual fields were
randomly selected from each section).
Nissl staining
Following sacrifice, a 1-cm SC section centered on
the injury site was removed at 14 days post-surgery, immediately
fixed in 4% paraformaldehyde at room temperature for 24 h, embedded
in paraffin and cut into 5-µm thick transverse sections. From each
group 12 sections were stained. Nissl staining was performed
according to the manufacturer's protocol (cat. no. G1434; Beijing
Solarbio Science & Technology Co., Ltd.) for 30 min at room
temperature. Sections were observed under a fluorescent microscope
(Eclipse E100; Nikon Corporation) at x100 magnification to evaluate
morphological changes (3 visual fields were randomly selected from
each section).
Uranium-lead staining and transmission
electron microscopy (TEM)
Following sacrifice, 1-cm SC sections centered on
the site of injury were removed at 14 days post-surgery and
immediately fixed in 2.5% glutaraldehyde at room temperature for 12
h, followed by immersion in 1% osmium acid at room temperature for
2 h. Following dehydration, tissues were embedded in pure epoxy
resin and cut into 70-nm thick sections using an ultra-thin slicer
(UC-7; Leica Microsystems, Inc.). Finally, sections were stained
with uranium and lead at room temperature for 30 min, and observed
under a TEM (JEM1400; JEOL, Ltd.) to assess myelin sheath
morphology. There were 12 sections for each group and 3 visual
fields were randomly selected from each section.
Statistical analysis
Data are presented as mean ± standard error of mean
(SEM) of 3 independent experiments. Multiple group means were
compared using two-way ANOVA with or without repeated measures or
one-way ANOVA all of which were followed by Tukey's post hoc tests
for multiple pair-wise comparisons. Pairs of means were compared
using the Student's t-test. SPSS software (version 19.0; SPSS Inc.)
was used for all statistical analyses. P<0.05 was considered to
indicate a statistically significant difference.
Results
OFS treatment improves functional
recovery following SCI
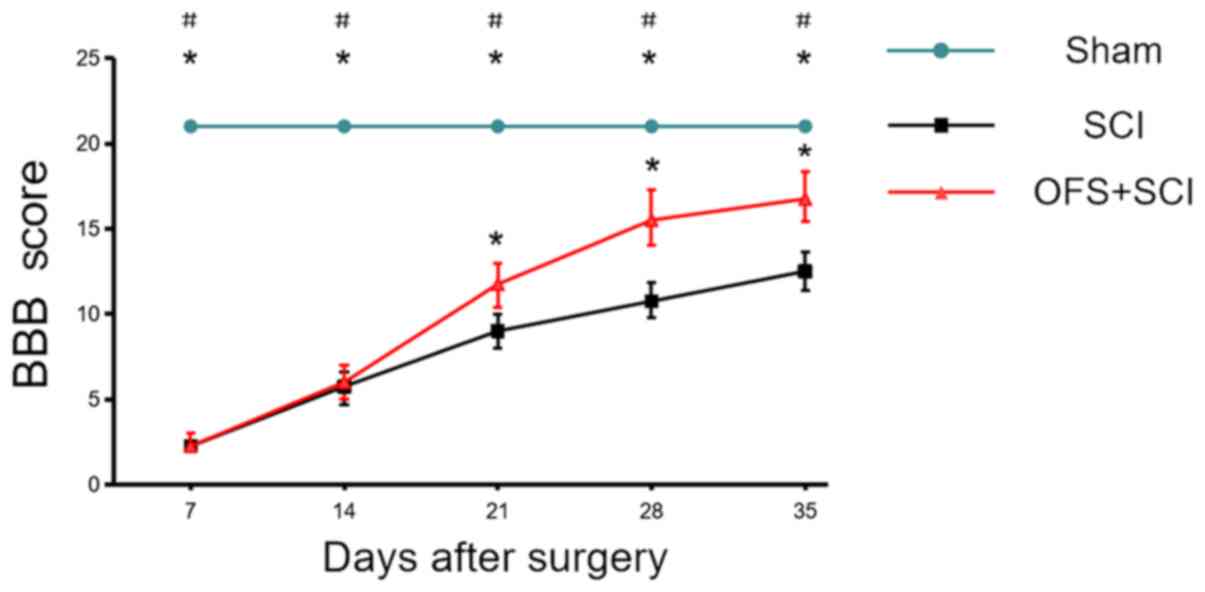
Hind limb movement and coordination were evaluated
using BBB scores to evaluate the efficacy of OFS for SCI treatment.
In the Sham group, there was no significant difference in BBB score
at each time point (P>0.05). Rats receiving OFS following SCI
demonstrated markedly higher scores compared with the SCI group
rats starting from the 3rd week post-surgery (P<0.05; Fig. 1).
OFS promotes the differentiation of
endogenous NSCs into neurons
IF double labeling for β-tubulin III and the neural
stem/progenitor cell marker Nestin was performed at days 3, 7 and
14 post-surgery to investigate whether OFS regulates the
differentiation of endogenous NSCs into neurons. There were more
Nestin and β-tubulin III positive cells in the SC sections from the
SCI + OFS group compared with the SCI and Sham groups at each
measurement time point (Fig. 2A),
with the Nestin and β-tubulin III positive cell number peaking in
the SCI + OFS group on the 7th day post-surgery compared with the
SCI group (P<0.05; Fig. 2B).
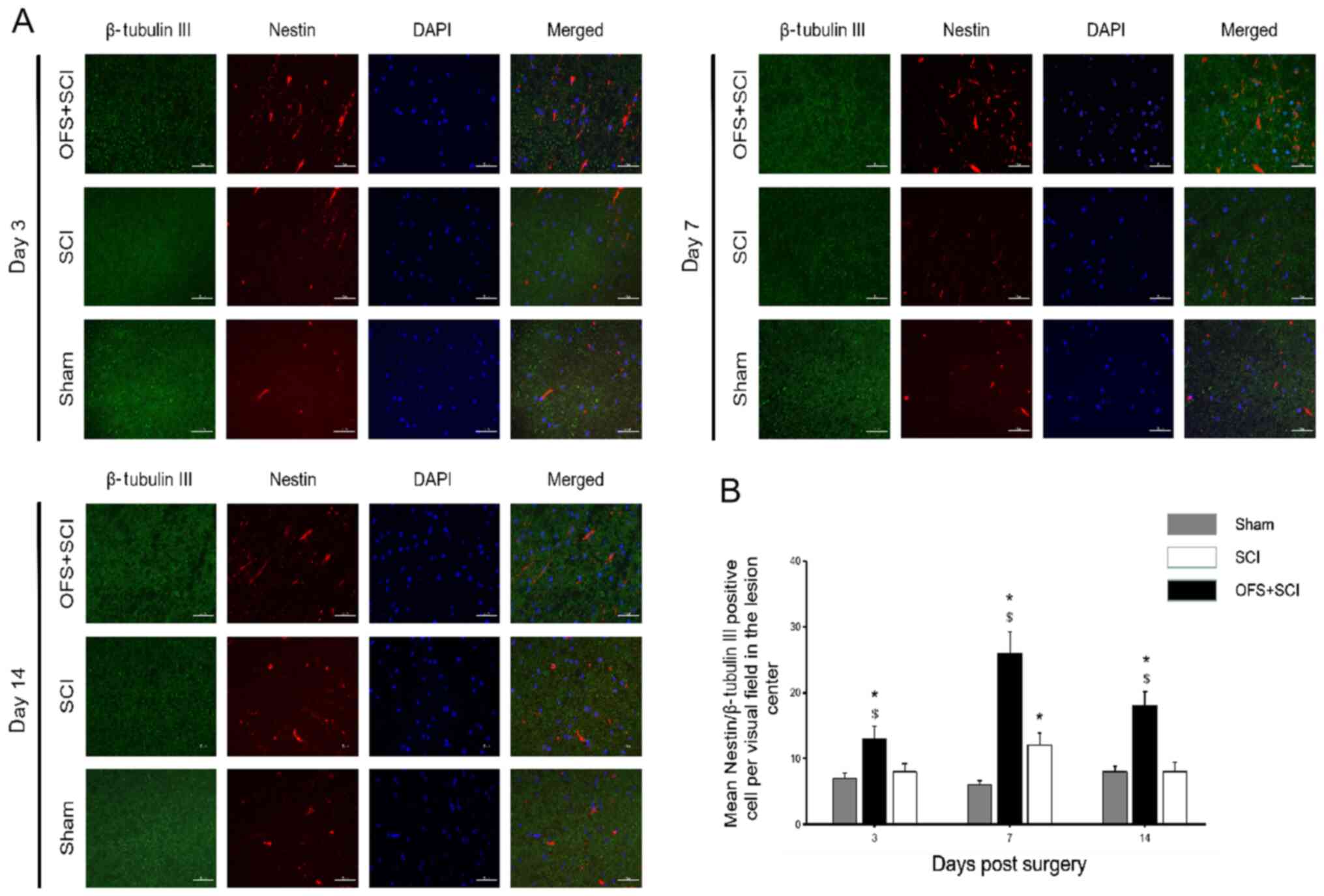 | Figure 2OFS treatment increases the
differentiation of endogenous neural stem cells into neurons. (A)
Immunofluorescence double-staining was used to identify the
expression of Nestin and β-tubulin Ⅲ among groups at day 3, 7 and
14 post-surgery. (B) Quantification of Nestin and β-tubulin Ⅲ
positive cells in the lesion site in each group at each time point.
Scale bar, 50 µm. DAPI staining, blue. Nestin staining, red.
β-tubulin Ⅲ staining, green. *P<0.05 vs. Sham group.
$P<0.05 vs. SCI group. Sham, controls that underwent
laminectomy only; SCI, group that underwent laminectomy and spinal
cord injury; OFS + SCI, group that underwent laminectomy, spinal
cord injury and OFS; OFS, oscillating field stimulation. |
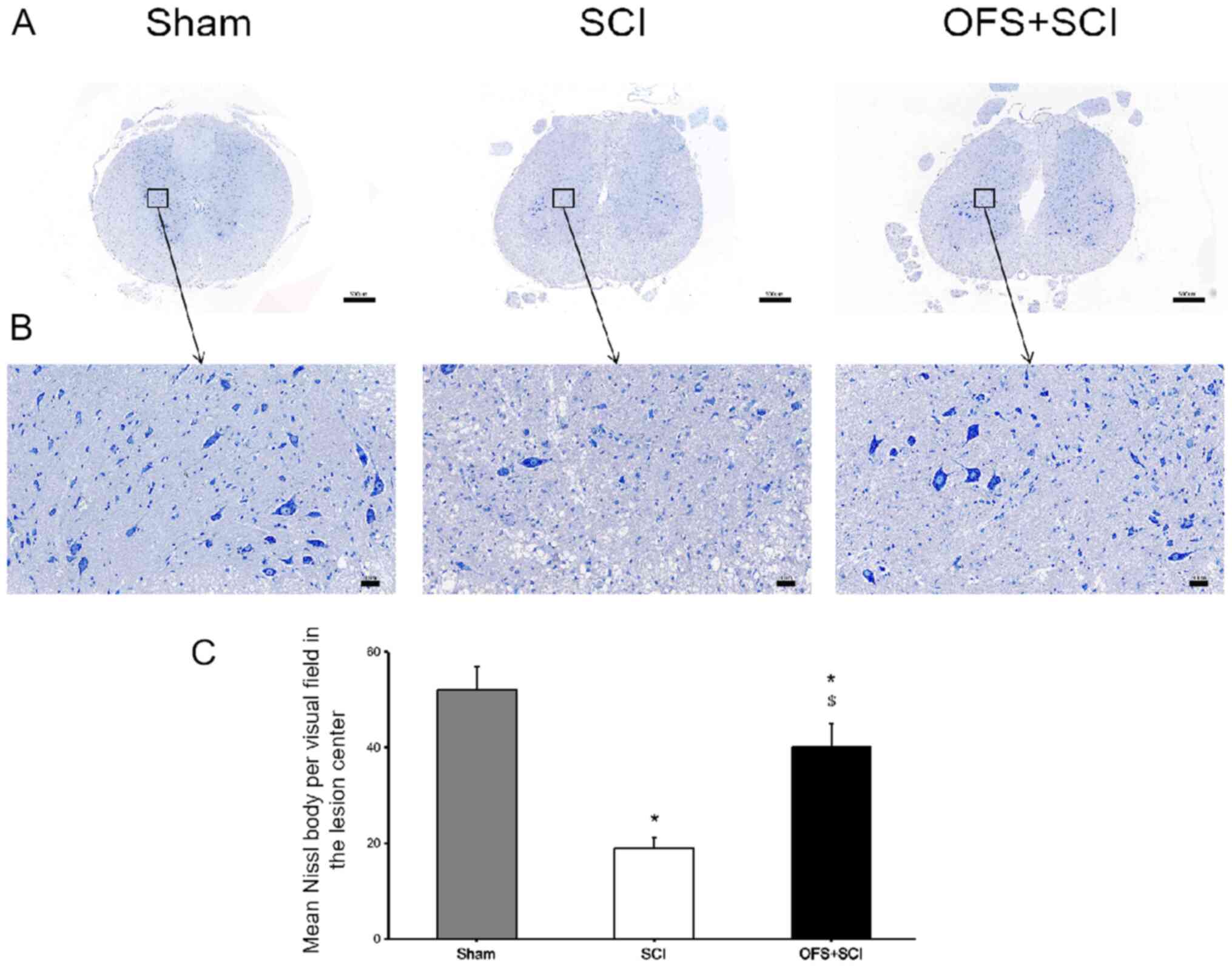
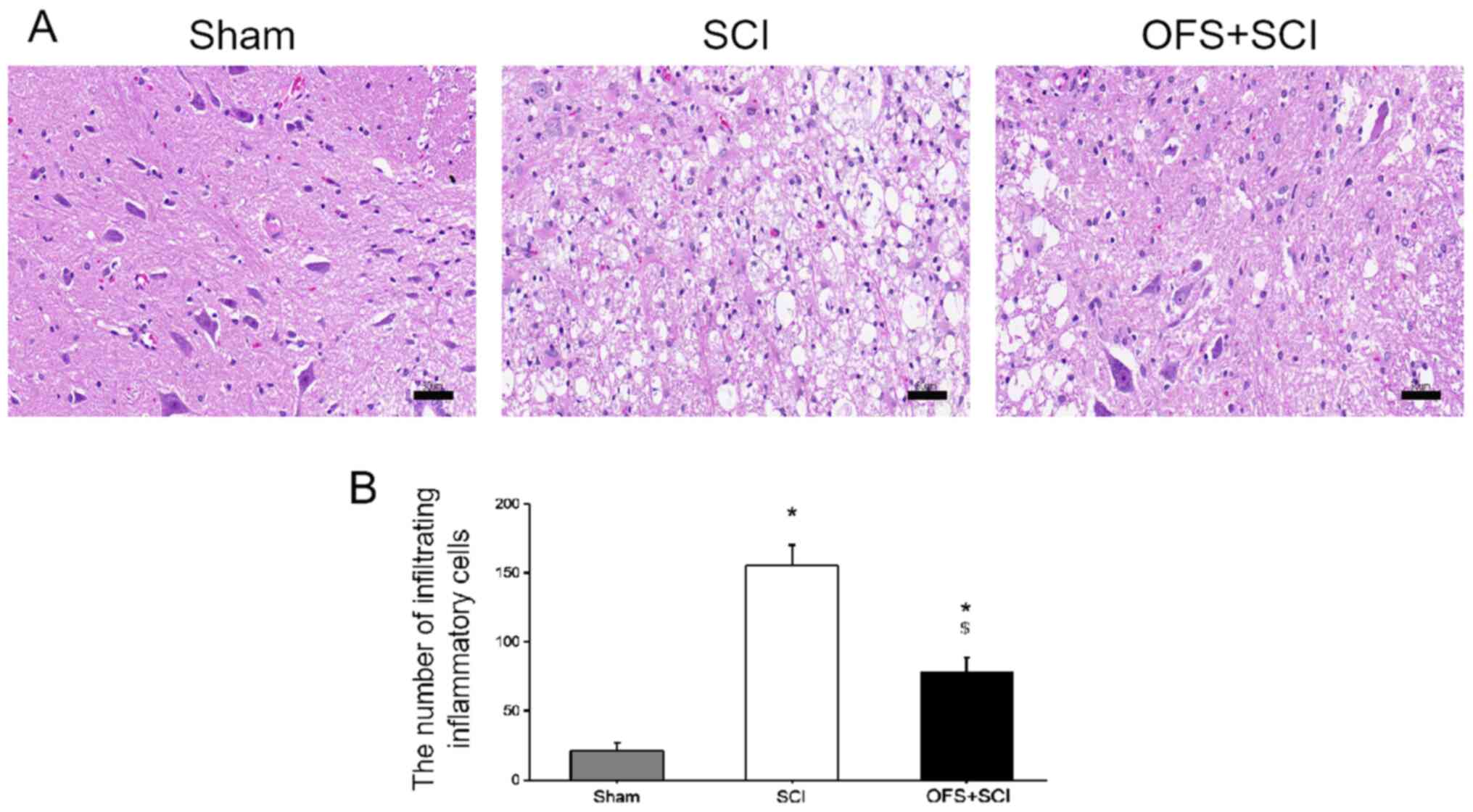
Nissl and H&E staining were also performed at
day 14 post-surgery to assess the level of tissue pathology. The
SCI group exhibited fewer Nissl bodies compared with the SCI + OFS
and Sham groups (P<0.05; Fig.
3). In addition, OFS treatment reduced inflammatory cell
infiltration compared with the SCI group (P<0.05; Fig. 4).
OFS promotes the differentiation of
endogenous NSCs into oligodendrocytes
Double IF staining was performed for Nestin and the
oligodendrocyte precursor cell (OPC) marker NG2 at days 3, 7 and 14
post-surgery to examine the effects of OFS on NSC differentiation
into oligodendrocytes (Fig. 5A). IF
staining revealed a significantly greater number of Nestin and
NG2-positive cells in the SCI + OFS group compared with the SCI
group at all measurement points (P<0.05; Fig. 5B), with cell number peaking on the
7th day post-surgery.
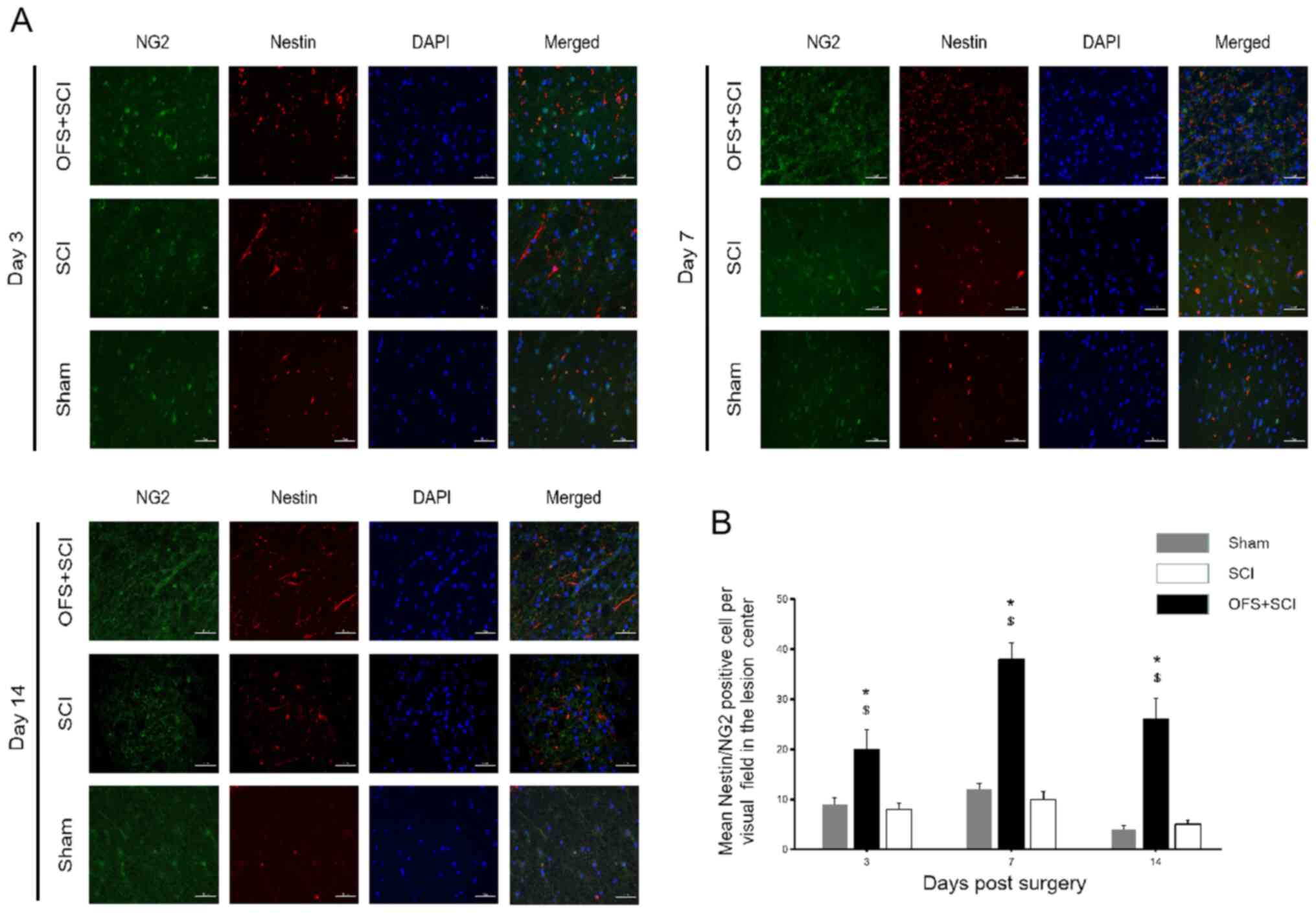 | Figure 5OFS treatment improves the
differentiation of endogenous NSCs into oligodendrocytes. (A)
Immunofluorescence double-staining was performed to identify the
expression of Nestin and NG2 among groups at day 3, 7 and 14
post-surgery. (B) Quantification of Nestin and NG2 positive cells
at the lesion site in each group at respective time points. Scale
bar, 50 µm. DAPI staining, blue. Nestin staining, red. NG2
staining, green. *P<0.05 vs. Sham group.
$P<0.05 vs. SCI group. NG2, neuron-glial antigen 2;
Sham, controls that underwent laminectomy only; SCI, group that
underwent laminectomy and spinal cord injury; SCI + OFS, group that
underwent laminectomy, spinal cord injury and OFS; OFS, oscillating
field stimulation. |
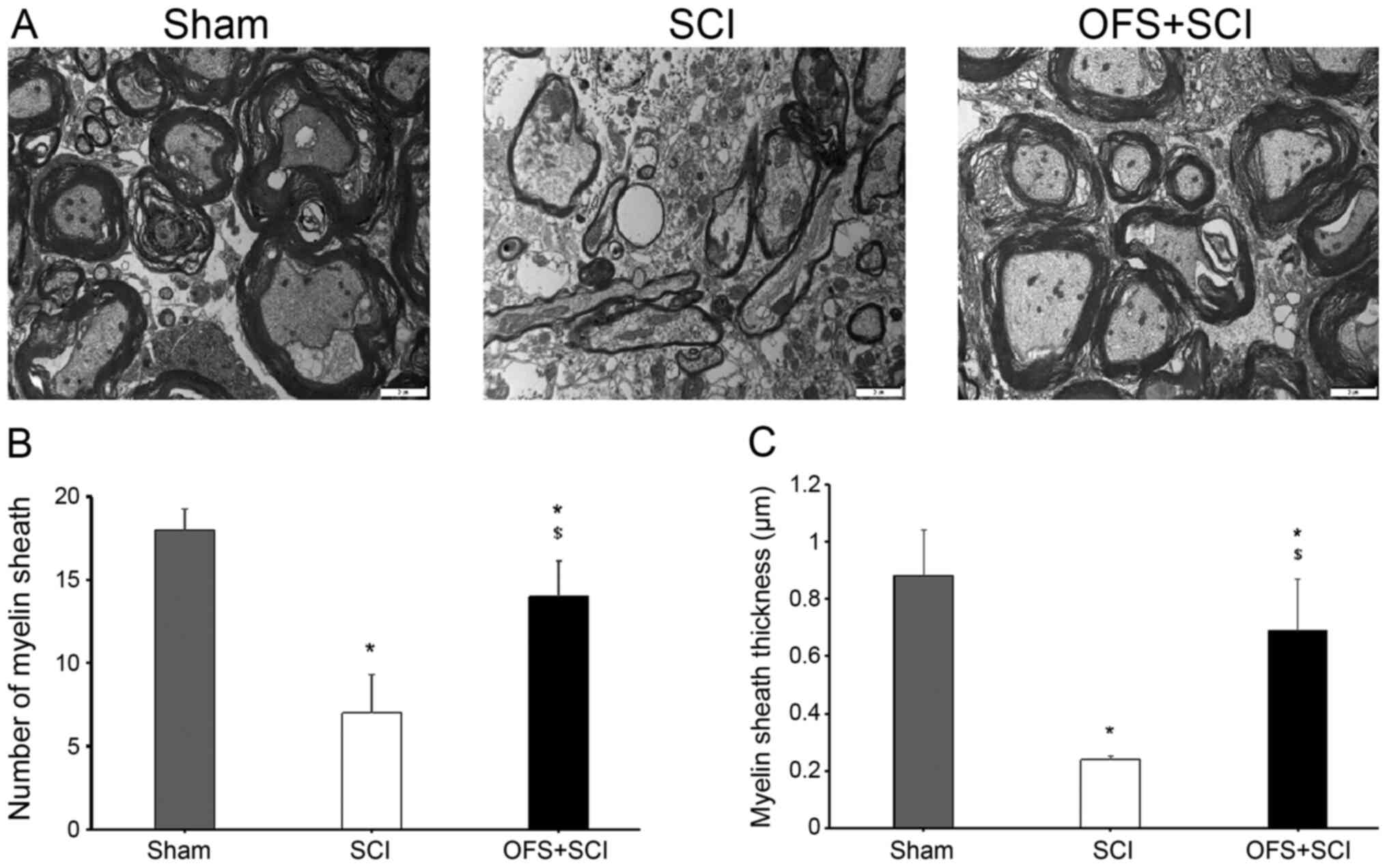
Myelin sheath morphology was examined using TEM to
assess the effects of OFS on remyelination following SCI. Compared
with the SCI group, the Sham and SCI + OFS group exhibited a
significantly greater number of myelin sheaths and thicker sheaths
at 14 days post-surgery (P<0.05; Fig. 6).
Discussion
Stem cell transplants have demonstrated promising
results for the treatment of SCI (17). However, cell rejection and poor
in vivo survival have limited the success of this strategy
(18,19). Recent studies have shown that
endogenous NSCs can improve the self-repair capacity of the SC,
indicating that endogenous NSCs may be potential therapeutic
targets for SCI treatment (20,21).
The differentiation of NSCs depends primarily on their
microenvironment following SCI, which is influenced by factors such
as edema, ischemia and activation of inflammatory response, and is
also an important factor affecting damage repair (22,23).
Therefore, regulation of directional differentiation of endogenous
NSCs may be the key to SCI repair.
Wnt signaling is an important regulator of NSC
proliferation, self-renewal and differentiation in the central
nervous system (CNS) (24,25). Various Wnt factors directly regulate
the proliferation and differentiation of endogenous NSC (12). Previous studies have reported that
Wnt1, Wnt2 and Wnt3a expression results in NSC proliferation in the
CNS (26,27). Furthermore, Wnt3a expression directs
the differentiation of mouse embryonic stem cells into dorsal
interneurons (28) and Wnt7a
expression promotes neuronal differentiation of NSCs in mice
(29). Additionally, it has been
revealed that electric field stimulation alters Wnt protein
expression and Wnt signaling, thus regulating the differentiation
of NSCs and affecting nerve regeneration and SCI repair. OFS, as a
neuroelectric technology, is an emerging neurostimulation modality
that has shown promise for facilitating repair after SCI (10,14).
The present study also demonstrated that OFS regulated the
differentiation of endogenous NSCs following SCI. However, the
potential mechanism needs further study.
To the best of our knowledge, the current study was
the first to identify that targeted OFS to the SCI site promotes
the differentiation of endogenous NSCs into neurons and
oligodendrocytes and accelerates recovery of hind limb function.
The BBB scores revealed substantially faster and improved locomotor
improvement in the SCI + OFS group compared with the SCI group with
no apparent adverse effects on survival of rats, indicating that
OFS is a feasible strategy for the treatment of SCI.
β-tubulin is one of the main components of the
cytoskeleton and the class III isoform is expressed exclusively in
neurons, where it contributes to morphological development and axon
orientation (30). NSCs promote SC
repair by enhancing neuronal viability, reducing neuroinflammation
and replenishing cells lost to apoptosis (31). Although axonal repair or
regeneration have been the main focus of SCI research, regeneration
of oligodendrocytes and remyelination are also critical for
recovery of neurological function after SCI (32). The present results suggested that
OFS enhanced the production of NSC-derived oligodendrocytes and
neurons in the early post-SCI phase as shown by Nestin and NG2
double labeling and Nestin and β-tubulin III double labeling,
respectively. Nissl staining identified more neurons in SCI rats
receiving OFS. Additionally, H&E staining demonstrated less
infiltration of inflammatory cells in the OFS group compared with
SCI group. Collectively, these results indicated that OFS therapy
promoted the differentiation of endogenous NSCs into viable
neurons.
Previous studies have identified OPCs in the mature
mammalian CNS that contribute to the repair of nerve fiber
demyelination (33-35).
It has also been shown that OFS can enhance the differentiation of
OPCs into oligodendrocytes and promote remyelination following SCI
in rats (14). In the current
study, it was found that Nestin and NG2-positive cell numbers at
the lesion site and Nestin and NG2 expression levels were
significantly increased as early as 3 days following SCI in the OFS
group. Uranium-lead staining and TEM directly demonstrated that OFS
increased the number of myelinated fibers and myelin sheath
thickness. Thus, OFS therapy improved the differentiation of
endogenous NSCs into oligodendrocytes, thereby promoting
remyelination of nerve fibers following SCI in rats.
A total of 19 Wnt proteins have been discovered in
mammals and some of these are expressed at various stages of CNS
development (29). In the process
of treating SCI, the degree of interaction between OFS and the Wnt
signaling pathway, as well as the specific Wnt proteins involved in
repair remain unknown. Therefore, these factors require further
investigation.
In conclusion, OFS promotes NSC differentiation into
neurons and oligodendrocytes following SCI in rats, thereby
accelerating axonal regeneration, remyelination and neurological
recovery. Therefore, it was speculated that OFS may be a safe and
effective strategy for the treatment of SCI, either alone or in
conjunction with NSC transplant.
Acknowledgements
Not applicable.
Funding
Funding: The current study was supported by the National
NaturalScience Foundation of China (grant no. 81471273 and
81671204), theExcellent Young Talents Support Program in
Universities(grant no. gxyqZD2017032) and the Natural science
research projects in Anhui Universities (grant no. KJ2020ZD23).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CF and JQ designed this research and wrote the
manuscript; CF and JS performed the experiments; LW and FG
collected and analyzed the data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
All surgical procedures and experiments were
performed in accordance with the National Institutes of Health
Guide for the Care and Use of Laboratory Animals (NIH Publications
no. 80-23; revised 1996) and were approved by the Animal Ethics
Committee of Anhui Medical University (approval reference no.
LLSC20190736). All applicable international, national and/or
institutional guidelines for the care and use of animals were
followed.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wyndaele M and Wyndaele JJ: Incidence,
prevalence and epidemiology of spinal cord injury: What learns a
worldwide literature survey? Spinal Cord. 44:523–529.
2006.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Singh A, Tetreault L, Kalsi-Ryan S, Nouri
A and Fehlings MG: Global prevalence and incidence of traumatic
spinal cord injury. Clin Epidemiol. 6:309–331. 2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Middleton JW, Dayton A, Walsh J, Rutkowski
SB, Leong G and Duong S: Life expectancy after spinal cord injury:
A 50-year study. Spinal Cord. 50:803–811. 2012.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Devivo MJ: Epidemiology of traumatic
spinal cord injury: Trends and future implications. Spinal Cord.
50:365–372. 2012.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Dyck SM and Karimi-Abdolrezaee S:
Chondroitin sulfate proteoglycans: Key modulators in the developing
and pathologic central nervous system. Exp Neurol. 269:169–187.
2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Sabelström H, Stenudd M and Frisén J:
Neural stem cells in the adult spinal cord. Exp Neurol. 260:44–49.
2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Grégoire CA, Goldenstein BL, Floriddia EM,
Barnabé-Heider F and Fernandes KJ: Endogenous neural stem cell
responses to stroke and spinal cord injury. Glia. 63:1469–1482.
2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Stenudd M, Sabelström H and Frisén J: Role
of endogenous neural stem cells in spinal cord injury and repair.
JAMA Neurol. 72:235–237. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Taveggia C, Feltri ML and Wrabetz L:
Signals to promote myelin formation and repair. Nat Rev Neurol.
6:276–287. 2010.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Shapiro S, Borgens R, Pascuzzi R, Roos K,
Groff M, Purvines S, Rodgers RB, Hagy S and Nelson P: Oscillating
field stimulation for complete spinal cord injury in humans: A
phase 1 trial. J Neurosurg Spine. 2:3–10. 2005.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wu Y, Collier L, Qin W, Creasey G, Bauman
WA, Jarvis J and Cardozo C: Electrical stimulation modulates Wnt
signaling and regulates genes for the motor endplate and calcium
binding in muscle of rats with spinal cord transection. BMC
Neurosci. 14(81)2013.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Piccin D and Morshead CM: Wnt signaling
regulates symmetry of division of neural stem cells in the adult
brain and in response to injury. Stem Cells. 29:528–538.
2011.PubMed/NCBI View
Article : Google Scholar
|
|
13
|
Li Z, Yao F, Cheng L, Cheng W, Qi L, Yu S,
Zhang L, Zha X and Jing J: Low frequency pulsed electromagnetic
field promotes the recovery of neurological function after spinal
cord injury in rats. J Orthop Res. 37:449–456. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Jing JH, Qian J, Zhu N, Chou WB and Huang
XJ: Improved differentiation of oligodendrocyte precursor cells and
neurological function after spinal cord injury in rats by
oscillating field stimulation. Neuroscience. 303:346–351.
2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Cho SR, Kim YR, Kang HS, Yim SH, Park CI,
Min YH, Lee BH, Shin JC and Lim JB: Functional recovery after the
transplantation of neurally differentiated mesenchymal stem cells
derived from bone marrow in a rat model of spinal cord injury. Cell
Transplant. 25(1423)2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Basso DM, Beattie MS, Bresnahan JC,
Anderson DK, Faden AI, Gruner JA, Holford TR, Hsu CY, Noble LJ,
Nockels R, et al: MASCIS evaluation of open field locomotor scores:
Effects of experience and teamwork on reliability. Multicenter
Animal Spinal Cord Injury Study. J Neurotrauma. 13:343–359.
1996.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Cofano F, Boido M, Monticelli M, Zenga F,
Ducati A, Vercelli A and Garbossa D: Mesenchymal stem cells for
spinal cord injury: current options, limitations, and future of
cell therapy. Int J Mol Sci. 20(2698)2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Galderisi U, Peluso G, Di Bernardo G,
Calarco A, D'Apolito M, Petillo O, Cipollaro M, Fusco FR and Melone
MA: Efficient cultivation of neural stem cells with controlled
delivery of FGF-2. Stem Cell Res (Amst). 10:85–94. 2013.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Hernández J, Torres-Espín A and Navarro X:
Adult stem cell transplants for spinal cord injury repair: Current
state in preclinical research. Curr Stem Cell Res Ther. 6:273–287.
2011.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Liu S and Chen Z: Employing Endogenous
NSCs to Promote Recovery of Spinal Cord Injury. Stem Cells Int.
2019(1958631)2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Yu JH, Seo J-H, Lee JY, Lee M-Y and Cho
S-R: Induction of neurorestoration from endogenous stem cells. Cell
Transplant. 25:863–882. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Sandner B, Prang P, Rivera FJ, Aigner L,
Blesch A and Weidner N: Neural stem cells for spinal cord repair.
Cell Tissue Res. 349:349–362. 2012.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Schwab ME: Repairing the injured spinal
cord. Science. 295:1029–1031. 2002.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Willert K, Brown JD, Danenberg E, Duncan
AW, Weissman IL, Reya T, Yates JR III and Nusse R: Wnt proteins are
lipid-modified and can act as stem cell growth factors. Nature.
423:448–452. 2003.PubMed/NCBI View Article : Google Scholar
|
|
25
|
David MD, Cantí C and Herreros J: Wnt-3a
and Wnt-3 differently stimulate proliferation and neurogenesis of
spinal neural precursors and promote neurite outgrowth by canonical
signaling. J Neurosci Res. 88:3011–3023. 2010.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Sousa KM, Villaescusa JC, Cajanek L, Ondr
JK, Castelo-Branco G, Hofstra W, Bryja V, Palmberg C, Bergman T,
Wainwright B, et al: Wnt2 regulates progenitor proliferation in the
developing ventral midbrain. J Biol Chem. 285:7246–7253.
2010.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Megason SG and McMahon AP: A mitogen
gradient of dorsal midline Wnts organizes growth in the CNS.
Development. 129:2087–2098. 2002.PubMed/NCBI
|
|
28
|
Murashov AK, Pak ES, Hendricks WA, Owensby
JP, Sierpinski PL, Tatko LM and Fletcher PL: Directed
differentiation of embryonic stem cells into dorsal interneurons.
FASEB J. 19:252–254. 2005.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Hirabayashi Y, Itoh Y, Tabata H, Nakajima
K, Akiyama T, Masuyama N and Gotoh Y: The Wnt/beta-catenin pathway
directs neuronal differentiation of cortical neural precursor
cells. Development. 131:2791–2801. 2004.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Binarová P and Tuszynski J: Tubulin:
structure, functions and roles in disease. Cells.
8(1294)2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Lu P, Wang Y, Graham L, McHale K, Gao M,
Wu D, Brock J, Blesch A, Rosenzweig ES, Havton LA, et al:
Long-distance growth and connectivity of neural stem cells after
severe spinal cord injury. Cell. 150:1264–1273. 2012.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Hesp ZC, Goldstein EZ, Miranda CJ, Kaspar
BK and McTigue DM: Chronic oligodendrogenesis and remyelination
after spinal cord injury in mice and rats. J Neurosci.
35:1274–1290. 2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Kang SH, Fukaya M, Yang JK, Rothstein JD
and Bergles DE: NG2+ CNS glial progenitors remain
committed to the oligodendrocyte lineage in postnatal life and
following neurodegeneration. Neuron. 68:668–681. 2010.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Guo YE, Suo N, Cui X, Yuan Q and Xie X:
Vitamin C promotes oligodendrocytes generation and remyelination.
Glia. 66:1302–1316. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Skaper SD: Oligodendrocyte precursor cells
as a therapeutic target for demyelinating diseases. Prog Brain Res.
245:119–144. 2019.PubMed/NCBI View Article : Google Scholar
|