Introduction
Sepsis is a life-threatening organ dysfunction
caused by dysregulated host response to infection (1,2).
Substantial efforts have been made to improve the outcome of
patients with sepsis; however, treatment of sepsis remains a big
challenge to clinicians (3). The
underlying mechanism of its pathogenesis has not been fully
elucidated (4,5). Recently, several studies have reported
that the disruption of the endothelial cell barrier induced high
vascular permeability, which serves a critical role in the
development of sepsis (6-8).
In addition to regulating axon guidance and cell
migration (9), the Slit/Robo
signaling pathway (in which the Slit ligand binds to the Robo
receptors) has also been demonstrated to serve an important role in
the development of organs (10,11).
The Slit protein is a secretory extracellular matrix protein, which
has three subtypes, Slit-1, -2 and -3(12). With respect to the Robo protein,
there are four subtypes, namely Robo-1, -2, -3 and -4(12); the Robo-4 protein is specifically
expressed in vascular endothelial (VE) cells and has been
associated with the genesis and development of blood vessels, and
serves an important role in maintaining the stability of VE cells
(6). The Slit2/Robo4 signaling
pathway was reported to be an endothelial permeability regulator,
which effectively regulates VE permeability (13). The Slit2/Robo4 pathway alleviates
the phagocytosis of VE-cadherin on the surface of VE cells mediated
by cytokines; thus, stabilizing VE cells and maintaining the
semi-permeable barrier of the microvessels (8). It is possible that the Slit2/Robo4
signaling pathway stabilizes the vasculature by enhancing
VE-cadherin localization to the cell surface. The maintenance of
the semi-permeable barrier between blood and peripheral tissues
represents a key component of therapy for sepsis (14,15).
Previous studies have shown that aminophylline, a
non-selective phosphodiesterase inhibitor, reduces endothelial cell
permeability (16-18).
However, the underlying mechanisms remain unclear. In the present
study, the mechanism of aminophylline in regulating the
permeability of endothelial cells via the Slit2/Robo4 signaling
pathway in a lipopolysaccharide (LPS)-induced inflammation model
was investigated.
Materials and methods
Reagents and cells
The human umbilical vein endothelial cells (HUVECs)
were purchased from the American Type Culture Collection. LPS
(Escherichia coli O111:B4) and Slit2 ELISA kit (cat. no.
HPA019511) were purchased from Sigma-Aldrich (Merck-KGaA), while
aminophylline was purchased from Shandong Xinhua Pharmaceutical
Co., Ltd. The Robo4 (cat. no. MAB50041), VE-cadherin (cat. no.
AF1002), fibronectin (cat. no. AF1918) and integrin (cat. no.
AF2045) primary antibodies were purchased from R&D Systems,
Inc. Alexa Fluor 488- (cat. no. sc-362257) and 594- (cat. no.
sc-362277) conjugated and GAPDH antibodies (cat. no. sc-47724) were
purchased from Santa Cruz Biotechnology, Inc. The N-terminal Slit2
(Slit2-N) protein was purchased from Abcam. FBS, trypsin,
penicillin/streptomycin and DMEM were purchased from Thermo Fisher
Scientific, Inc. All other chemicals were purchased from
Sigma-Aldrich (Merck KGaA).
Cell culture
The HUVECs (1x107) were cultured in
six-well culture plates with DMEM (supplemented with 10% FBS and 1%
penicillin/streptomycin) and grown to 80% confluence. For all
experiments, the cells were deprived of serum for 24 h before LPS
was added. After culturing for 24 h with different concentrations
of LPS (50, 100 and 200 µg/ml (at 37˚C), HUVECs were treated with
aminophylline (1 mM) for an additional 24 h (at 37˚C). All
experiments were repeated at least three times.
Immunofluorescence staining
The cells were fixed with 4% paraformaldehyde in PBS
for 15 min at room temperature, then washed with PBS and blocked
with 10% bovine serum in PBS for 1 h at room temperature.
Subsequently, the cells were incubated overnight at 4˚C with
primary antibodies against Robo4 (dilution 1:100) and VE-cadherin
(dilution 1:100). After washing with PBS adequately, the Alexa
Fluor 488- or 594-conjugated secondary antibodies (dilution 1:200)
were added and the samples were incubated at room temperature for 2
h in the dark. Subsequently, the cells were incubated with DAPI for
nuclei staining at room temperature for 15 min). Fluorescent images
were captured using a fluorescent microscope (magnification, x400)
(Nikon Corporation).
Western blot analysis
Western blot analysis was performed as previously
described (19). Briefly, the
protein concentrations were measured by BCA protein assay kit
(Thermo Scientific, Inc.) according to the manufacturer's
instructions. Equal amounts of total proteins (5-10 µg) were
subjected to 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) under reducing conditions and
transferred to nitrocellulose membrane (EMD Millipore). After
blocking with 5% skimmed milk in TBS with 0.01% Tween-20 for 1 h at
room temperature, the membranes were incubated with primary
antibodies against Robo4 (dilution 1:1,000), VE-cadherin (dilution
1:1,000), fibronectin (dilution 1:1,000), integrin (dilution
1:1,000) and GAPDH (dilution 1:1,000) overnight at 4˚C, followed by
treatment with the horseradish peroxidase (HRP)-conjugated
secondary antibody for 1 h at room temperature. Finally, the
protein expression level was detected with an enhanced
chemiluminescence kit (Bio-Rad Laboratories, Inc.). The relative
protein expression was analyzed by the software program Image J
1.42 (National Institutes of Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
RT-qPCR was performed with SYBR Green PCR real-time
PCR Master mix (Invitrogen; Thermo Fisher Scientific, Inc.), using
real-time cycle conditions of 95˚C and 5 min, followed by 40 cycles
of 95˚C, 30 sec and 60˚C, 1 min, as previously described (20). Data were analyzed through the
comparative threshold cycle (CT) method. Briefly, RNA was extracted
from the cells using TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions.
The total RNA concentration and purity were assessed using UV
spectrophotometry (20). cDNA was
synthesized from total RNA using RT and the Superscript III First
Strand RT-PCR kit (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's instructions (19,20).
The primers used in the PCR were as follows: Slit2:
CCGAAGGTTACAGTGGCTTGTTCT (Forward), CCGCTGTCTTCATCTGTGGCAAT
(Reverse); GAPDH: GATGCTGGTGCTGAGTATGRCG (Forward),
GTGGTGCAGGATGCATTGCTCTGA (Reverse). The PCR products were amplified
and identified using 2% agarose gel electrophoresis. GAPDH served
as an internal control. The gene products were expressed as a
change in mRNA expression levels relative to GAPDH.
Transendothelial permeability
assay
The HUVECs were cultured in Transwell upper chambers
(8-µm pore size, Corning, Inc.) at 37˚C in a humidified incubator
with 5% CO2 and maintained until confluent. After 24 h
of culture with LPS, the HUVECs were treated with aminophylline for
an additional 24 h at 37˚C. Then, complete medium containing 1
mg/ml FITC-conjugated dextran or 0.5 µM HRP replaced the medium in
the upper Transwell chamber (incubation was 6 h and 37˚C) The
fluorescence in the lower chamber was measured using a TECAN GeNios
microplate reader (Tecan Group, Ltd.). The HRP was removed from the
lower chamber and analyzed spectrophotometrically at 492 nm with
buffer containing 0.5 mM tetramethylbenzidine. For normalization,
FITC-conjugated dextran and HRP concentration was calculated and
divided by the concentration of FITC-conjugated dextran and HRP at
baseline with no LPS added.
Statistical analysis
All the experimental data are presented as the mean
± standard error of the mean and were analyzed using a Student's
t-test or one-way ANOVA followed by a Tukey's multiple comparisons
test. Statistical analyses were performed using the SPSS v19.0.
software (IBM Corp.). P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression level of Slit2 and Robo4 in
LPS-induced inflammation
Firstly, the mRNA and protein expression level of
Slit2 and Robo4, respectively, in the HUVECs following LPS-induced
inflammation was investigated. Immunofluorescent staining and
western blot analysis revealed that the Robo4 protein expression
level was decreased following LPS-induced inflammation in a
dose-dependent manner (Fig. 1A-F).
In addition, the expression levels of other cell surface markers,
such as fibronectin and integrin, did not change under LPS
stimulation (Fig. 1E). RT-qPCR
analysis revealed that the Slit2 mRNA expression level was
significantly downregulated in HUVECs after LPS stimulation
(Fig. 1G).
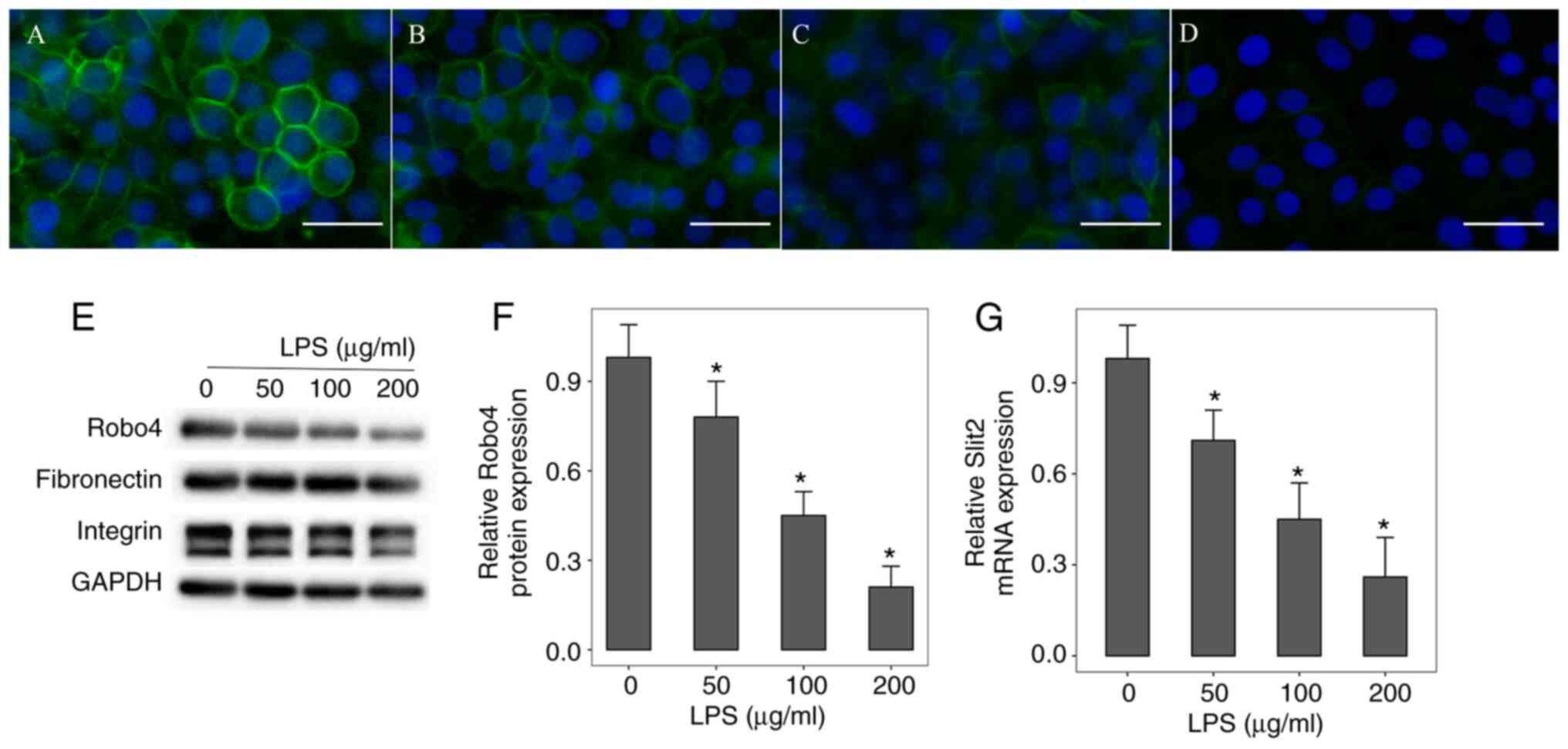 | Figure 1Expression of Slit2 and Robo4 in
LPS-induced inflammation. HUVECs were incubated with (A) 0, (B) 50,
(C) 100 or (D) 200 µg/ml LPS for 24 h. Immunofluorescent staining
revealed that Robo4 protein expression level was decreased in a
dose-dependent manner. Scale bar, 50 µm. Original magnification,
x400. Green indicates Robo4, while blue indicates DAPI. (E and F)
Western blot analysis of Robo4, fibronectin and integrin protein
expression level. (G) Reverse transcription-quantitative PCR
analysis of Slit2 mRNA expression levels. *P<0.05 vs.
control group. The data are presented as the mean ± standard error
of the mean. All experiments were repeated at least three times.
LPS, lipopolysaccharide; DAPI, 4',6-diamidino-2 phenylindole;
HUVECs, human umbilical vein endothelial cells. |
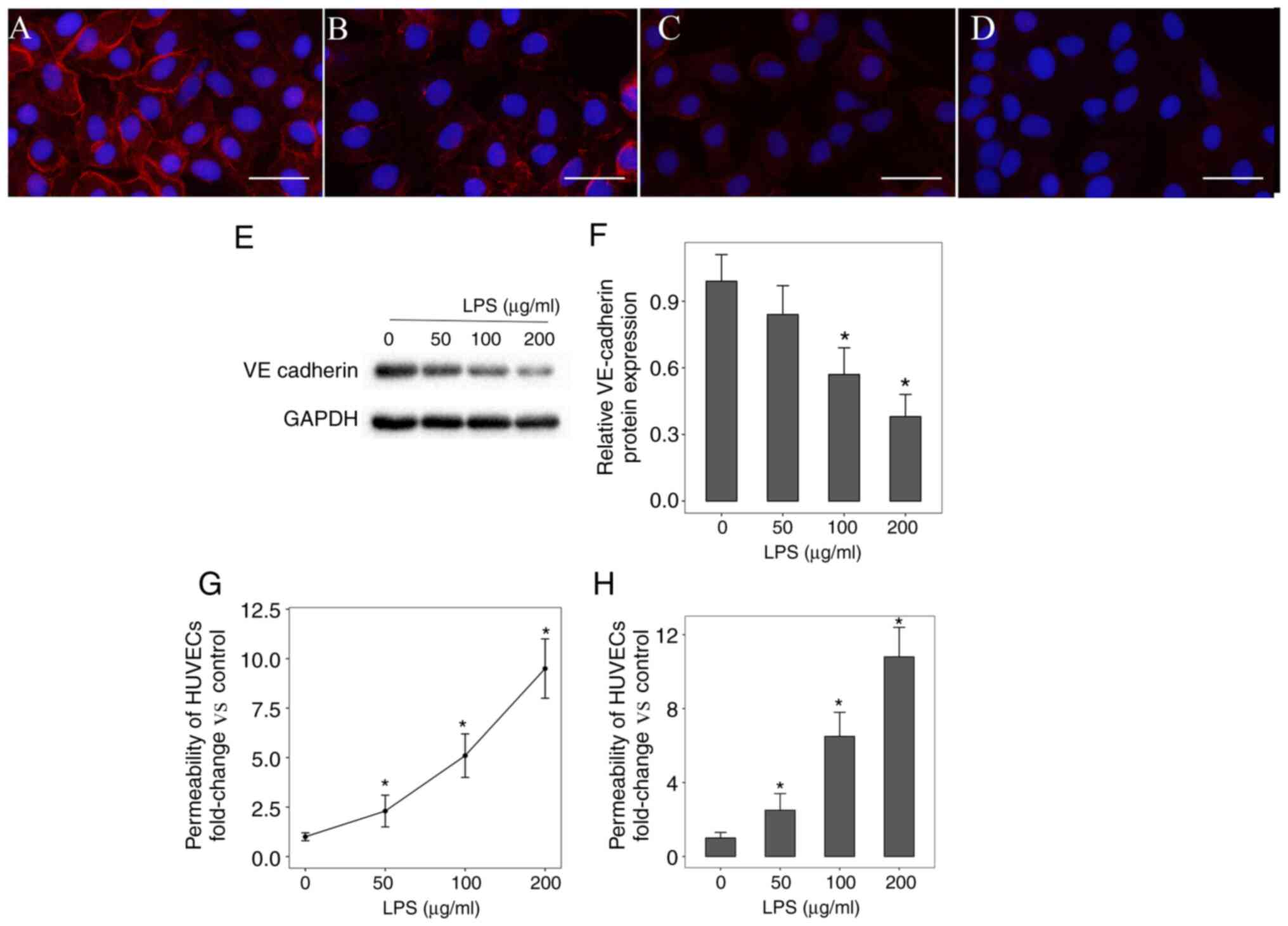
Integrity and stability of HUVECs in
LPS-induced inflammation
Subsequently, the VE-cadherin protein expression
level and permeability of HUVECs following LPS-induced inflammation
was investigated. Immunofluorescent staining and western blot
analysis revealed that VE-cadherin protein expression level was
significantly downregulated compared with the Robo4 protein
expression level (Fig. 2A-F). The
Transwell permeability assay revealed that LPS significantly
increased HUVECs permeability in a dose-dependent manner (Fig. 2G and H).
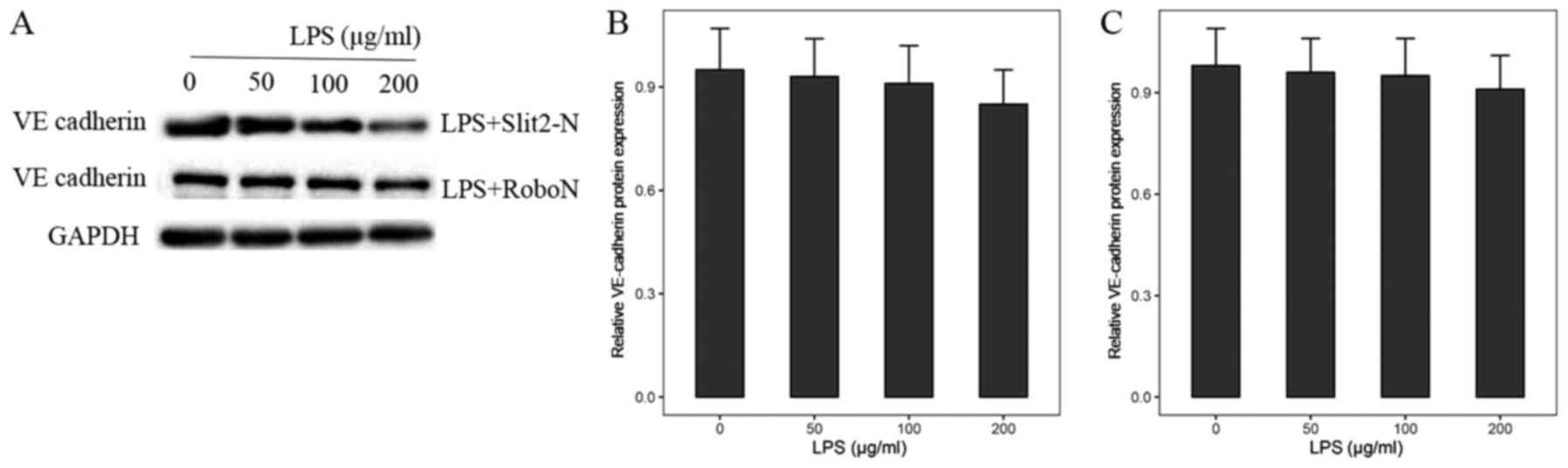
Slit2/Robo4 pathway modulates
VE-cadherin protein expression level in LPS-induced
inflammation
Based on the aforementioned results, the association
between the Slit2/Robo4 signaling pathway and the VE-cadherin
protein expression level in LPS-induced inflammation was
investigated. The HUVECs stimulated by LPS were incubated with 20
ng/ml Slit2-N for 24 h. Western blot analysis showed that the
protein expression level of VE-cadherin was significantly
upregulated (Fig. 3A and B). When the Robo4 receptor inhibitor,
RoboN was added, the VE-cadherin protein expression level did not
significantly change following LPS-induced inflammation (Fig. 3A and C).
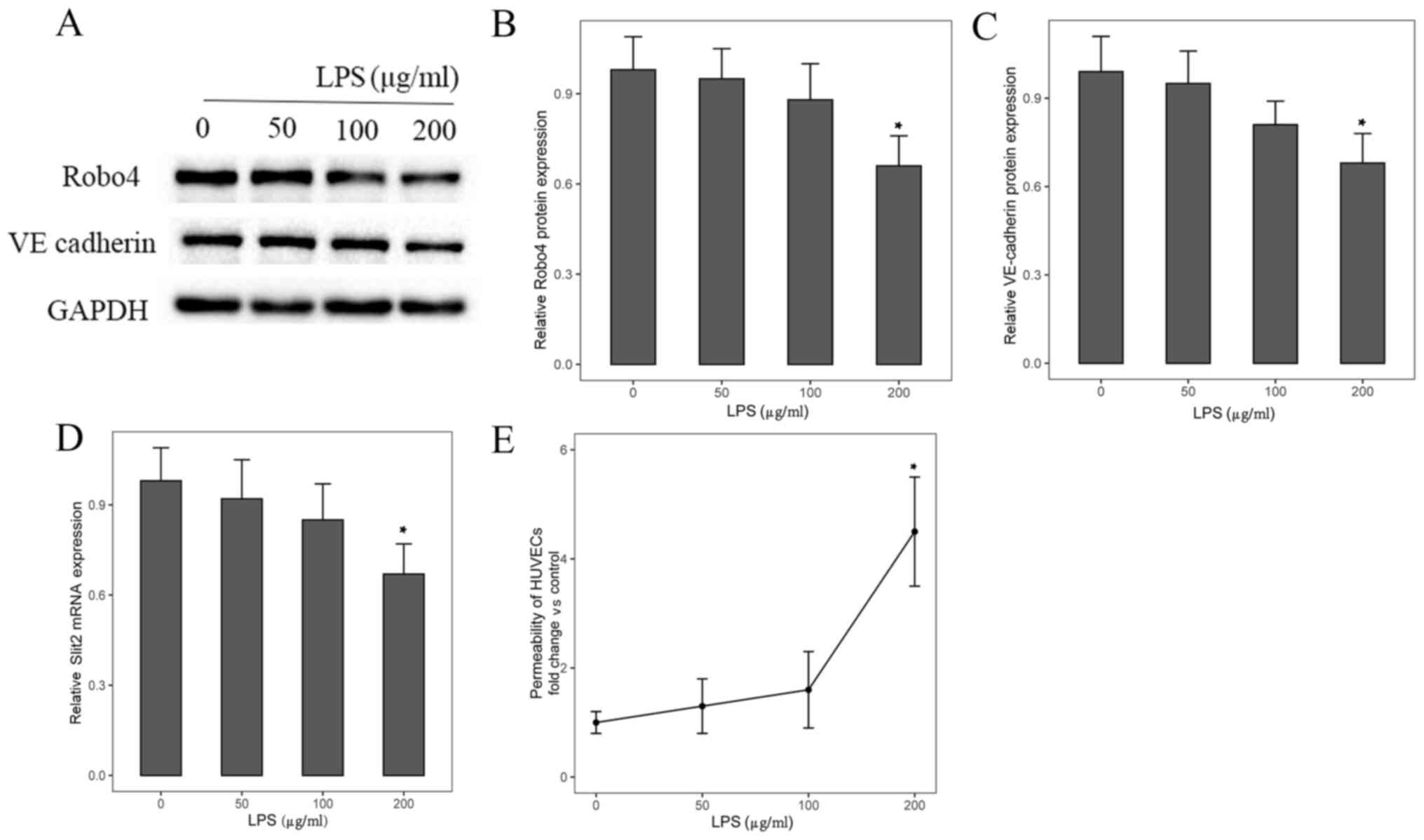
Aminophylline represses endothelial
permeability by upregulating VE-cadherin protein expression levels
via the Slit2/Robo4 signaling pathway
Next, the effect of aminophylline in LPS-induced
inflammation was investigated. To explore the effect of
aminophylline on endothelial cells permeability, three different
aminophylline concentrations (10 and 100 µm and 1 mM) were tested.
It was revealed that the 1 mM aminophylline significantly reduced
the permeability of endothelial cells (Fig. S1), so 1 mM aminophylline was used
for subsequent experiments in the current study. It was
demonstrated that aminophylline upregulated Robo4 and VE-cadherin
protein expression levels in HUVECs using immunofluorescent
staining (Fig. 4A-H) and western
blot analysis (Fig. 5A-C). RT-qPCR
revealed that Slit2 mRNA expression level was also upregulated in
HUVECs following aminophylline treatment (Fig. 5D). In addition, the Transwell
permeability assay revealed that aminophylline at 200 µg/ml
significantly alleviated endothelial cell permeability in
LPS-induced inflammation (Fig.
5E).
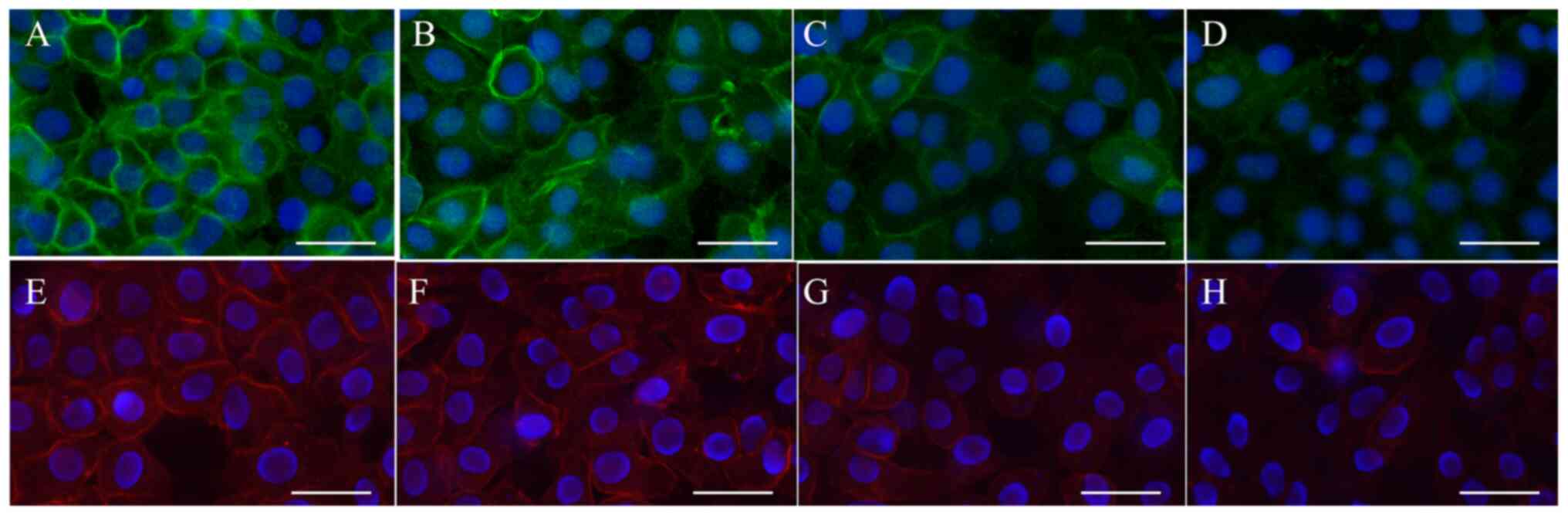 | Figure 4Robo4 and VE-cadherin protein
expression levels following aminophylline treatment. The HUVECs
were incubated with (A and E) 0 µg/ml LPS, (B and F) 50 µg/ml LPS +
aminophylline, (C and G) 100 µg/ml LPS + aminophylline and (D and
H) 200 µg/ml + aminophylline, both for 24 h, sequentially.
Immunofluorescent staining demonstrated that Robo4 and VE-cadherin
protein expression level were both upregulated following
aminophylline treatment. Scale bar, 50 µm. Original magnification,
x400. Green indicates Robo4, red indicates VE-cadherin and blue
indicates DAPI. VE, vascular endothelial; HUVECs, human umbilical
vein endothelial cells; LPS, lipopolysaccharide; DAPI,
4',6-diamidino-2 phenylindole. |
Discussion
The pathophysiological process of LPS-induced
inflammation is similar in nature to acute inflammation or sepsis
(21). The endothelial receptor,
Robo4 serves critical roles in endothelial cells, and the
Slit2/Robo4 signaling pathway is responsible for regulating the
expression level of the cell tight junction protein, VE-cadherin
and endothelial permeability (22,23).
In the present study was revealed that LPS treatment decreased
VE-cadherin protein expression levels and increased endothelial
permeability in vitro by downregulating the Slit2/Robo4
signaling pathway. Aminophylline alleviated LPS-induced endothelial
cell permeability, and upregulated Slit2, Robo4 and VE-cadherin
protein expression levels.
Robo4 is a membrane protein, located on the surface
of VE cells, and acts as a receptor for Slit2, which serves an
important role in stabilizing the vascular endothelium. Zhang et
al (24) found that targeting
the Slit2/Robo4 signaling pathway may protect the integrity of the
lymphatic barrier and reduce lymphatic endothelial
hyperpermeability. Shirakura et al (25) reported that Robo4 suppressed
vascular hyperpermeability and improved the survival of
Robo4-/- mice in a mouse model of inflammation. Robo4
was also responsible for stabilizing the VE-cadherin protein at
endothelial cell junctions, in an inflammation-induced
hyperpermeability model (25). A
transfusion-related acute lung injury study indicated that the
Slit2/Robo4 signaling pathway modulates endothelial cell
permeability, and regulates VE-cadherin protein expression level to
maintain endothelial barrier function (23). Similar results were also obtained in
the present study with an LPS-induced inflammation cell line model.
Nevertheless, another study revealed that Robo4 acted as a ligand
to bind UNC5B (a vascular Netrin receptor), and not to Slit2, which
may inhibit VEGF-induced vascular hyperpermeability (26).
Endothelial barrier dysfunction and capillary
leakage contribute to the pathological process of organ failure in
sepsis and in sepsis-related complications (27). It is well-known that cell-cell
junctions (primarily composed of VE-cadherin) are an important
component of the vascular barrier (28). Therapeutic strategies targeting
capillary leakage and endothelial cell dysfunction have been
considered to be potentially effective in improving the clinical
outcome of patients with sepsis (5).
Aminophylline acts as a non-selective
phosphodiesterase inhibitor, and has been demonstrated to have
effects on multiple systems, such as the pulmonary (29-31),
renal (32-34)
and cardiovascular systems (35). A
total of 30 years ago, Harada et al (36) demonstrated that aminophylline
attenuated the multiple organ albumin leaks in septic guinea pigs.
Previous studies, focusing on animal models of inflammatory
cytokine-induced acute lung injury, reported that pulmonary
vascular permeability was reduced following aminophylline
administration. These protective effects may be due to elevation of
intracellular cAMP/cGMP and/or inhibition of tumor necrosis, in
which pulmonary vascular endothelium leakage and pulmonary edema
are alleviated (37-40).
Notably, the results in the present study are slightly different
from previous studies. To the best of our knowledge, this is the
first study that demonstrated that aminophylline upregulated the
protein expression level of Slit2, Robo4 and VE-cadherin, and
decreased endothelial cell permeability, in HUVECs in a LPS-induced
inflammation model. The results from the present study suggest that
the protection of endothelial integrity and stability by
aminophylline may be mediated by VE-cadherin, which was modulated
via the Slit2/Robo4 signaling pathway. The maintenance of the
endothelial barrier function is a critical treatment strategy for
sepsis (6,13,15).
In addition, the results from the present study also indicated that
aminophylline might be a promising candidate for sepsis therapy, as
well as other vascular hyperpermeability diseases. However,
prospective studies are required to confirm the results of the
present study.
In conclusion, the present study revealed that LPS
downregulates the Slit2, Robo4 and VE-cadherin protein expression
levels during endotoxemia. The permeability of endothelial cells
was mediated by VE-cadherin via the Slit2/Robo4 signaling pathway.
Aminophylline reduced endothelial cell permeability during
LPS-induced inflammation. Taken together, aminophylline may
represent a promising candidate for modulating vascular
permeability induced by endotoxemia or sepsis.
Supplementary Material
Aminophylline dose and permeability
curve. Permeability of HUVECs in LPS-induced inflammation was
attenuate by aminophylline. HUVECs were incubated with 200 μg/ml
LPS for 24 h. Dextran permeability assay showed that aminophylline
significantly represses permeability in a dose-dependent manner.
*P<0.05 vs. control group. The data are presented as
the mean ± standard error of the mean. All experiments were
repeated at least three times. HUVECs, human umbilical vein
endothelial cells; LPS, lipopolysaccharide.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant. no. 81772054), Zhejiang
Medicines Health Science and Technology Program (grant. no.
2016KYB189) and the Wenzhou Science and Technology Bureau (grant.
nos. Y20170179 and Y20160114).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JX, JW, QC and XZ designed the study and performed
the experiments. RH and ZZ collected the data. ZW and YC analyzed
the data. QC and XZ prepared the manuscript. QC and JX confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Shankar-Hari M, Phillips GS, Levy ML,
Seymour CW, Liu VX, Deutschman CS, Angus DC, Rubenfeld GD and
Singer M: Sepsis Definitions Task Force: Developing a new
definition and assessing new clinical criteria for septic shock:
For the third international consensus definitions for sepsis and
septic shock (Sepsis-3). JAMA. 315:775–787. 2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Weng J, Wu H, Xu Z, Xi H, Chen C, Chen D,
Gong Y, Hua Y and Wang Z: The role of propionic acid at diagnosis
predicts mortality in patients with septic shock. J Crit Care.
43:95–101. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Cecconi M, Evans L, Levy M and Rhodes A:
Sepsis and septic shock. Lancet. 392:75–87. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Lelubre C and Vincent JL: Mechanisms and
treatment of organ failure in sepsis. Nat Rev Nephrol. 14:417–427.
2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Cross D, Drury R, Hill J and Pollard AJ:
Epigenetics in sepsis: Understanding its role in endothelial
dysfunction, immunosuppression, and potential therapeutics. Front
Immunol. 10(1363)2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Ni J, Lin M, Jin Y, Li J, Guo Y, Zhou J,
Hong G, Zhao G and Lu Z: Gas6 attenuates sepsis-induced tight
junction injury and vascular endothelial hyperpermeability via the
Axl/NF-κB signaling pathway. Front Pharmacol.
10(662)2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Gao S, Wake H, Gao Y, Wang D, Mori S, Liu
K, Teshigawara K, Takahashi H and Nishibori M: Histidine-rich
glycoprotein ameliorates endothelial barrier dysfunction through
regulation of NF-κB and MAPK signal pathway. Br J Pharmacol.
176:2808–2824. 2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Martinez-Quinones P, Komic A, McCarthy CG,
Webb RC and Wenceslau CF: Targeting endothelial barrier dysfunction
caused by circulating bacterial and mitochondrial N-formyl peptides
with deformylase. Front Immunol. 10(1270)2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Rothberg JM, Hartley DA, Walther Z and
Artavanis-Tsakonas S: Slit: An EGF-homologous locus of D.
Melanogaster involved in the development of the embryonic central
nervous system. Cell. 55:1047–1059. 1988.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Mommersteeg MT, Andrews WD, Ypsilanti AR,
Zelina P, Yeh ML, Norden J, Kispert A, Chedotal A, Christoffels VM
and Parnavelas JG: Slit-roundabout signaling regulates the
development of the cardiac systemic venous return and pericardium.
Circ Res. 112:465–475. 2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Fan X, Li Q, Pisarek-Horowitz A, Rasouly
HM, Wang X, Bonegio RG, Wang H, McLaughlin M, Mangos S, Kalluri R,
et al: Inhibitory effects of Robo2 on nephrin: A crosstalk between
positive and negative signals regulating podocyte structure. Cell
Rep. 2:52–61. 2012.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Jiang Z, Liang G, Xiao Y, Qin T, Chen X,
Wu E, Ma Q and Wang Z: Targeting the SLIT/ROBO pathway in tumor
progression: Molecular mechanisms and therapeutic perspectives.
Ther Adv Med Oncol. 11(1758835919855238)2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhao H, Anand AR and Ganju RK: Slit2-Robo4
pathway modulates lipopolysaccharide-induced endothelial
inflammation and its expression is dysregulated during endotoxemia.
J Immunol. 192:385–393. 2014.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Schnoor M, Ponce AG, Vadillo E, Pelayo R,
Rossaint J and Zarbock A: Actin dynamics in the regulation of
endothelial barrier functions and neutrophil recruitment during
endotoxemia and sepsis. Cell Mol Life Sci. 74:1985–1997.
2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Xiao F, Wang D, Kong L, Li M, Feng Z,
Shuai B, Wang L, Wei Y, Li H, Wu S, et al: Intermedin protects
against sepsis by concurrently re-establishing the endothelial
barrier and alleviating inflammatory responses. Nat Commun.
9(2644)2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Möller AD and Grände PO: Low-dose
prostacyclin is superior to terbutaline and aminophylline in
reducing capillary permeability in cat skeletal muscle in vivo.
Crit Care Med. 27:130–136. 1999.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Korn C, Neidlein R, Strein K and Wilhelms
OH: Comparison of the effects of ularitide acetate and other
bronchorelaxing substances on the thrombin-induced permeability
raise of human endothelial cell monolayers. Arzneimittelforschung.
48:251–258. 1998.PubMed/NCBI
|
|
18
|
Foy T, Marion J, Brigham KL and Harris TR:
Isoproterenol and aminophylline reduce lung capillary filtration
during high permeability. J Appl Physiol Respir Environ Exerc
Physiol. 46:146–151. 1979.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Weng J, Tu M, Wang P, Zhou X, Wang C, Wan
X, Zhou Z, Wang L, Zheng X, Li J, et al: Amiodarone induces cell
proliferation and myofibroblast differentiation via ERK1/2 and p38
MAPK signaling in fibroblasts. Biomed Pharmacother.
115(108889)2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Weng J, Chen H, Wu H, Tu M, Wang Z, Chen
D, Wang Z and Chen C: Amiodarone induces epithelial-mesenchymal
transition in A549 cells via activation of TGF-β1. Drug Chem
Toxicol. 43:415–422. 2020.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Dickson K and Lehmann C: Inflammatory
response to different toxins in experimental sepsis models. Int J
Mol Sci. 20(4341)2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhang F, Prahst C, Mathivet T,
Pibouin-Fragner L, Zhang J, Genet G, Tong R, Dubrac A and Eichmann
A: The Robo4 cytoplasmic domain is dispensable for vascular
permeability and neovascularization. Nat Commun.
7(13517)2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Weng J, Zhou X, Xie H, Gao Y, Wang Z and
Gong Y: Slit2/Robo4 signaling pathway modulates endothelial
hyper-permeability in a two-event in vitro model of
transfusion-related acute lung injury. Blood Cells Mol Dis.
76:7–12. 2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zhang X, Yu J, Kuzontkoski PM, Zhu W, Li
DY and Groopman JE: Slit2/Robo4 signaling modulates HIV-1
gp120-induced lymphatic hyperpermeability. PLoS Pathog.
8(e1002461)2012.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Shirakura K, Ishiba R, Kashio T, Funatsu
R, Tanaka T, Fukada SI, Ishimoto K, Hino N, Kondoh M, Ago Y, et al:
The Robo4-TRAF7 complex suppresses endothelial hyperpermeability in
inflammation. J Cell Sci. 132(jcs220228)2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Koch AW, Mathivet T, Larrivée B, Tong RK,
Kowalski J, Pibouin-Fragner L, Bouvrée K, Stawicki S, Nicholes K,
Rathore N, et al: Robo4 maintains vessel integrity and inhibits
angiogenesis by interacting with UNC5B. Dev Cell. 20:33–46.
2011.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Goldenberg NM, Steinberg BE, Slutsky AS
and Lee WL: Broken barriers: A new take on sepsis pathogenesis. Sci
Transl Med. 3(88ps25)2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Darwish I and Liles WC: Emerging
therapeutic strategies to prevent infection-related microvascular
endothelial activation and dysfunction. Virulence. 4:572–582.
2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Mizus I, Summer W, Farrukh I, Michael JR
and Gurtner GH: Isoproterenol or aminophylline attenuate pulmonary
edema after acid lung injury. Am Rev Respir Dis. 131:256–259.
1985.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Lindenschmidt RC and Witschi H:
Attenuation of pulmonary fibrosis in mice by aminophylline. Biochem
Pharmacol. 34:4269–4273. 1985.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Nowak D, Rozniecki J, Ruta U, Bednarowicz
A and Izdebski J: The influence of aminophylline on human
neutrophils-possible protection of lung from proteolytic injury.
Arch Immunol Ther Exp (Warsz). 36:351–360. 1988.PubMed/NCBI
|
|
32
|
Bhatt GC, Gogia P, Bitzan M and Das RR:
Theophylline and aminophylline for prevention of acute kidney
injury in neonates and children: A systematic review. Arch Dis
Child. 104:670–679. 2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Seo K, Choi JW, Kim DW, Han D, Noh SJ and
Jung HS: Aminophylline effect on renal ischemia-reperfusion injury
in mice. Transplant Proc. 49:358–365. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Su X, Xie X, Liu L, Lv J, Song F, Perkovic
V and Zhang H: Comparative effectiveness of 12 treatment strategies
for preventing contrast-induced acute kidney injury: A systematic
review and bayesian network meta-analysis. Am J Kidney Dis.
69:69–77. 2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Matthay RA and Mahler DA: Theophylline
improves global cardiac function and reduces dyspnea in chronic
obstructive lung disease. J Allergy Clin Immunol. 78 (4 Pt
2):793–799. 1986.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Harada H, Ishizaka A, Yonemaru M, Mallick
AA, Hatherill JR, Zheng H, Lilly CM, O'Hanley PT and Raffin TA: The
effects of aminophylline and pentoxifylline on multiple organ
damage after escherichia coli sepsis. Am Rev Respir Dis.
140:974–980. 1989.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Hsu K, Wang D, Chang ML, Wu CP and Chen
HI: Pulmonary edema induced by phorbol myristate acetate is
attenuated by compounds that increase intracellular cAMP. Res Exp
Med (Berl). 196:17–28. 1996.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Sciuto AM, Strickland PT, Kennedy TP and
Gurtner GH: Postexposure treatment with aminophylline protects
against phosgene-induced acute lung injury. Exp Lung Res.
23:317–332. 1997.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Berthiaume Y: Effect of exogenous cAMP and
aminophylline on alveolar and lung liquid clearance in anesthetized
sheep. J Appl Physiol (1985). 70:2490–2497. 1991.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Sato K, Stelzner TJ, O'Brien RF, Weil JV
and Welsh CH: Pentoxifylline lessens the endotoxin-induced increase
in albumin clearance across pulmonary artery endothelial monolayers
with and without neutrophils. Am J Respir Cell Mol Biol. 4:219–227.
1991.PubMed/NCBI View Article : Google Scholar
|