Introduction
Thyroid carcinoma is a malignant tumor that is
derived from thyroid epithelial cells and is a common endocrine
malignancy (1). Compared with other
types thyroid cancer, the incident rate of papillary thyroid
carcinoma is highest, and is responsible for 90% of thyroid
cancers. However, early detection of papillary thyroid carcinoma
usually results in successful treatment and a favorable patient
prognosis (2,3). The incidence of papillary thyroid
carcinoma has increased in the previous few decades, and the 5-year
survival rate is 59% in the advanced stage, worldwide (4). Therefore, identifying novel drugs and
investigating the underlying mechanisms of this disease are
important for improving patient overall survival.
Emodin, which is also known as
1,3,8-trihydroxy-6-methy-anthraquinone, can be isolated from a
number of Chinese medicinal herbs, including Rheum and Polygonum
(5). A number of studies have
demonstrated that emodin inhibits growth in multiple cancer types,
including lung (5) and pancreatic
cancer (6), and hepatocellular
carcinoma (7). However, to the best
of our knowledge, no research on the effect of emodin on papillary
thyroid carcinoma has been previously performed. Previous reports
have indicated that emodin serves an inhibitory role via AMPK and
ERK in non-small cell lung cancer (8) and breast cancer (9).
AMPK has been revealed to serve an important role in
cellular metabolic processes, including nutrient deprivation,
mitochondrial dysfunction and oxidative stress (10). AMPK signaling has been indicated to
be associated with cell differentiation, proliferation and the
promotion of oncogenes and tumor growth, such as colorectal cancer
and lung cancer (11,12). However, whether AMPK serves a role
in activation or inhibition during emodin treatment in papillary
thyroid carcinoma requires clarification.
In the current study, whether emodin exerts a role
in inhibiting the development of papillary thyroid carcinoma via
AMPK was investigated further.
Materials and methods
Cell lines
TPC-1 cells were obtained from the China Center for
Type Culture Collection (Wuhan, China). TPC-1 cells were cultured
in RPMI-1640 medium (Thermo Fisher Scientific, Inc.) supplemented
with 10% FBS (Thermo Fisher Scientific, Inc.), 100 U/ml penicillin
and 100 mg/ml streptomycin (Thermo Fisher Scientific, Inc.). Cells
were cultured at 37˚C in 5% CO2.
Cell grouping
To study the effect of emodin on the proliferation
of TPC-1 cells, 10, 25 and 50 µM emodin were added to TPC-1 cells
for 24, 48 and 72 h at 37˚C, respectively.
To further study the underlying mechanism of emodin
on TPC-1 cells, cells were treated with different concentrations of
emodin (0, 5, 10, 20, 40 or 80 µm) for 48 h at 37˚C, and the
IC50 was calculated to be 49.01 µM using SPSS 20.0 (IBM
Corp.). Thus, emodin at a concentration of 50 µM was used for
subsequent experiments. The cells were divided into the control
group, emodin treatment group (Emodin), AMPK activator group
(AICAR), and emodin + AMPK inhibitor group (Combine). The cells of
the control group were treated with PBS. In the emodin group, 50 µM
emodin was added to cells for 48 h at 37˚C. Cells in the AICAR
group were treated with 2 mM AMPK activator (AICAR, Cell Signaling
Technology, Inc.) for 48 h at 37˚C (13). In the combination group, TPC-1 cells
were treated with 50 µM emodin and 10 µM AMPK inhibitor
(Dorsomorphin; cat. no. HY-13418A; MedChemExpress) for 48 h at
37˚C, as previously described (13). After 48 h, the cells were collected
and analyzed using Cell Counting Kit-8 (CCK-8), flow cytometry and
western blot analysis.
CCK-8 assay
TPC-1 cells (2x104 cells/ml) in each
group were seeded into 96-well plates and cultured at 37˚C at 5%
CO2. After incubation for 24, 48 and 72 h, cells were
incubated with 10 µl CCK-8 (Dojindo Molecular Technologies, Inc.)
at 37˚C for 4 h according to the manufacture's protocol. The
supernatant was carefully discarded and the absorbance of each hole
was measured at 450 nm. Cell viability=[Treatment (OD)]-Blank
(OD)/[Control (OD)-Blank (OD)].
Flow cytometry
Following incubation for 48 h, a AnnexinV-FITC/PI
cell apoptosis detection kit (Meilunbio) was used to measure
apoptotic cells. The cells (2x105) were centrifuged at
800 x g for 5 min at 4˚C, collected and washed with PBS twice at
4˚C. The cells (1x106/ml) were resuspended with 250 µl
1X binding buffer. The cells (100 µl) were subsequently added to a
pipe with 5 µl Annexin V-FITC and 10 µl propidium iodide (PI; 20
µg/ml). The mixture was cultured at 25˚C for 15 min in darkness.
The apoptotic rate was analyzed using a flow cytometer
(Cytoflex3L8C; Beckman Coulter, Inc.) and the results were analyzed
using Cell Quest software version 2.0 (BD Biosciences).
Animals
A total of 36 specific pathogen free Balb/c female
nude mice, (age, 5 weeks; weight, 18-20 g) were purchased from
Beijing Weitong Lihua Experimental Animal Technology Co., Ltd.
[license no. scxk (Jing) 20160006]. Food, water and bedding
materials were sterilized, and the animals were provided with food
and water ad libitum. Mice were housed in specifc pathogen
free conditions, at 22-26˚C with 40-70% humidity and 12 h
light/dark cycles.
Xenograft model
TPC-1 cells were centrifuged at 800 x g for 5 min at
4˚C, washed with sterile PBS three times and diluted. A total of 40
µl cell suspension was added to 10 µl phenol blue stain and cells
were counted under a microscope. A cell suspension with a
concentration of 5x108 cells/ml was made. Cells (~0.2
ml/1x107 cell/ml) were inoculated subcutaneously under
the right rear armpit of mice. The nude mice were randomly divided
into four groups: Model group, emodin group, AICAR group and the
combined group. In the emodin group, mice were intraperitoneally
injected with emodin at 40 mg/kg (14). In AICAR group, mice were
intraperitoneally injected with emodin at 40 mg/kg (14) and AICAR at 50 mg/kg (15). In the combination group, mice were
intraperitoneally injected with emodin at 40 mg/kg and dorsomorphin
at 0.2 mg/kg (16). In each group,
mice were treated three times a week for a period of two weeks. In
the model group, mice were intraperitoneally injected with normal
saline. The tumor volume (V)=(axb2)/2 was calculated
every week. The tumor volume was observed every week for 4 weeks.
The health and behaviors of animals were monitored every day,
including diet, weight, mental states and death. There was no death
observed during the experiment. The maximum percentage of body
weight loss was 10.2% in animals from start to endpoint. After a
period of 28 days, 3% pentobarbital sodium (40 mg/kg) was
intraperitoneally injected, and the nude mice were sacrificed via
cervical dislocation. Five minutes after cardiac arrest, death was
confirmed. The tumor was subsequently separated and weighed.
Partial tumor sections were stored at -80°C for use in
western blot analysis, and partially tumor sections were fixed with
4% paraformaldehyde at 25˚C for 24 h for use in the TUNEL
assay.
TUNEL
Tumor sections were 4 µm thick. After regular
dewaxing with xylene and gradient ethanol, the sections were
assessed using a Situ TUNEL Apoptosis Detection kit (cat. no.
G001-2-1; Nanjing Jiancheng Bioengineering Institute) according to
the manufacturer's protocol. In brief, the sections were treated
with 50 µl TdT reaction solution for 60 min at 37˚C, then washed
with PBS for in triplicate. A total of 50 µl streptavidin-HRP
solution was added for 30 min at 37˚C in the dark, after which the
solution was washed three times with PBS for 3 times. Samples were
then treated with 80 µl DAB solution at room temperature for 10
min. At the end of staining procedure, the number of TUNEL positive
cells was observed under the microscope. A total of five fields of
view were randomly selected, and the number of TUNEL positive cells
was counted. The nuclei of apoptotic cells were stained brown.
Apoptotic index=(number of apoptotic positive cells/total cells)
x100%.
Western blot analysis
Total protein of the TPC-1 cells or tissues in each
group was extracted using a Tissue or Cell Total Protein Extraction
kit (cat. no. BC3710; Beijing Solarbio Science & Technology
Co., Ltd.) and the concentration of proteins was assessed using a
BCA protein quantification kit (cat. no. 23225; Thermo Fisher
Scientific, Inc.). Samples (50 µg/lane) were separated using 12%
SDS-PAGE (Bio-Rad Laboratories, Inc.) and transferred to a PVDF
membrane (Bio-Rad Laboratories, Inc.). Skim milk powder (5%) was
used to block membranes for 1 h at 4˚C. The primary antibodies were
diluted with 5% BSA and incubated with samples at 4˚C overnight.
The primary antibodies used were as follows: Rabbit anti-human
proliferating cell nuclear antigen (PCNA) antibody (1:700; cat. no.
orb386383; Biorbyt, Ltd.), anti-Cleaved caspase-3 antibody (1:500;
cat. no. ab49822; Abcam), caspase-3 antibody (1:500; cat. no.
ab13847; Abcam), anti-Cyclin D1 antibody (1:200; cat. no. ab16663;
Abcam), anti-phosphorylated (p-)AMPKa1 (Thr172) Antibody (1:1,000;
cat. no. orb99303; Biorbyt, Ltd.), anti-AMPKa1 Antibody (1:1,000;
cat. no. orb338932; Biorbyt, Ltd.), anti-ERK1/2 antibody (1:600;
cat. no. orb106403; Biorbyt, Ltd.), anti-p-ERK1/2 antibody (1:600;
Biorbyt, Ltd.), anti-MEK antibody (1:600; cat. no. orb38774;
Biorbyt, Ltd.), anti-p-MEK antibody (1:600; cat. no. orb106207;
Biorbyt, Ltd.) and GAPDH antibody (1:8,000; cat. no. orb555879;
Biorbyt, Ltd.). The secondary antibody was HRP-labeled anti-rabbit
IgG (1:1,800; cat. no. ab6721; Abcam) for 1 h at 4˚C. GAPDH was
used as the internal control. Blots were visualized on X-ray film
using the enhanced chemiluminescence detection system (Cytiva).
Densitometric analysis of the bands for the expression of protein
was performed using Image J 6.0 (National Institutes of
Health).
Statistical analysis
The statistical data in the current study was
analyzed using SPSS 20.0 statistical software (IBM Corp.). The data
are presented as mean ± standard deviation. Among groups,
differences were analyzed using ANOVA and followed by Tukey's post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
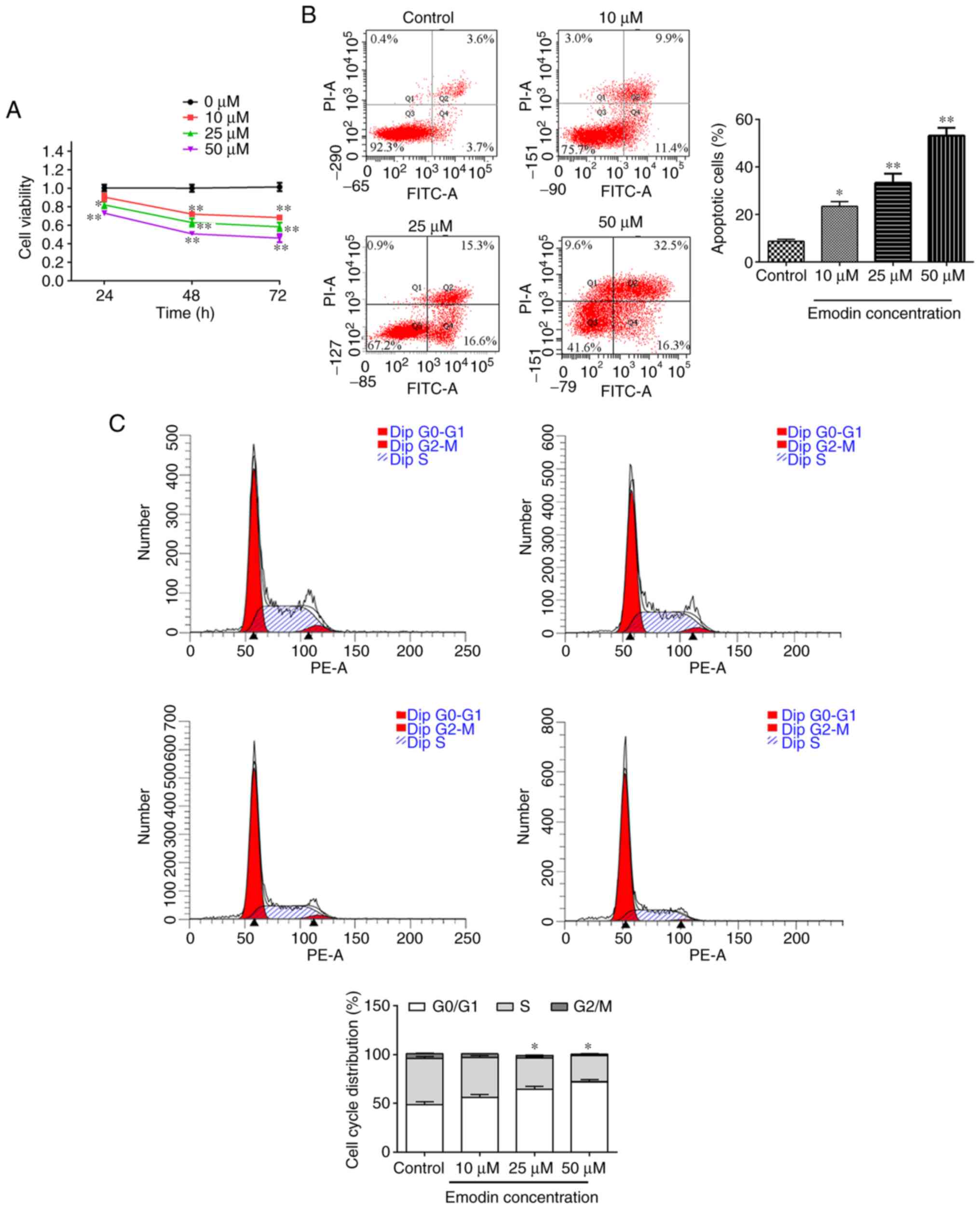
Emodin inhibits proliferation of
papillary thyroid cells
Compared with the control group, cell viability was
significantly decreased following treatment with varied doses of
emodin (Fig. 1A). Additionally, the
proportion of cells in the apoptosis and
G0/G1 phase increased significantly, and the
proportion of cells in the S phase decreased (P<0.05; Fig. 1B and C). These data indicated that emodin
inhibited TPC-1 cell proliferation and the cell cycle.
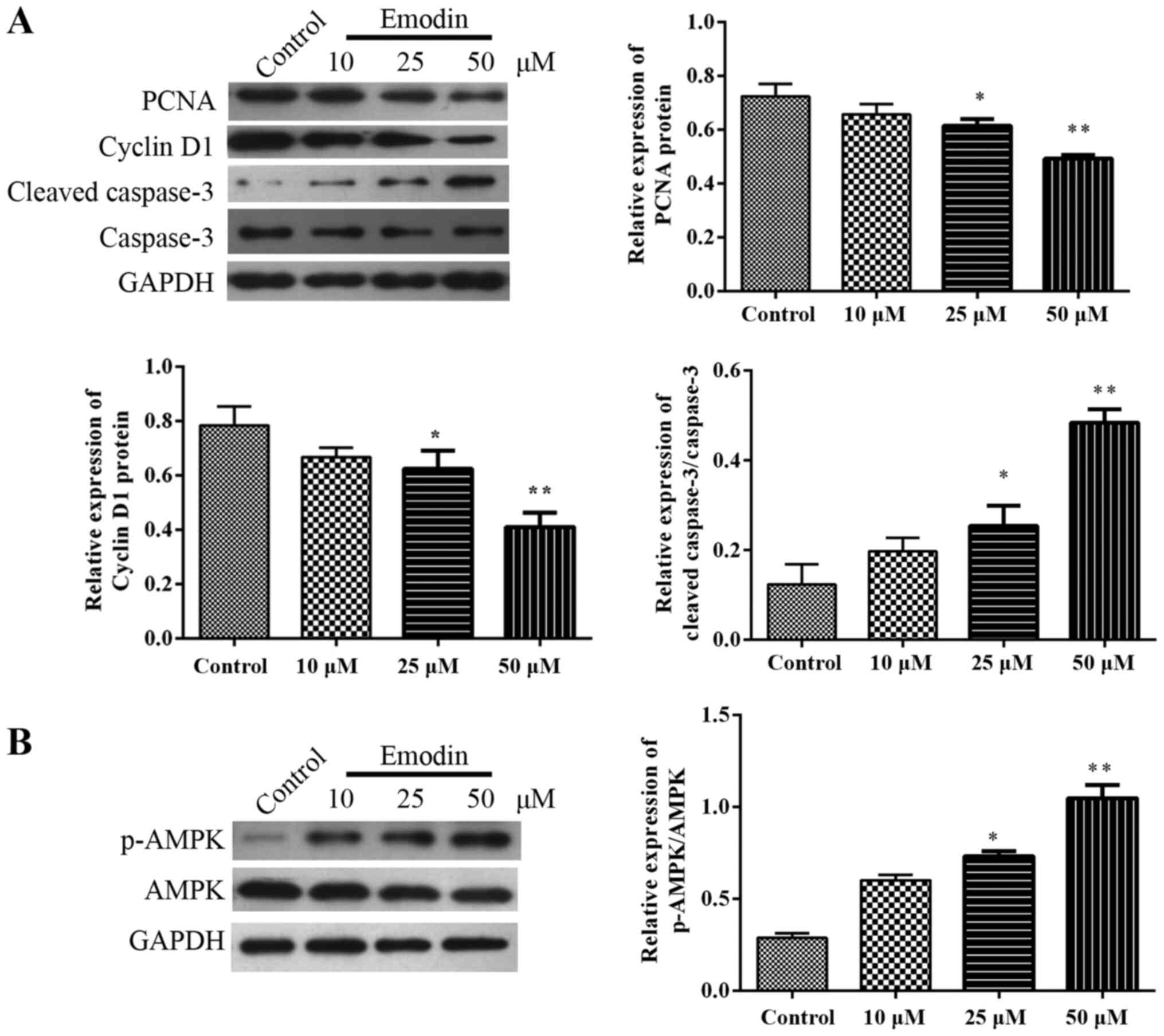
Emodin activates the phosphorylation
of the AMPK pathway
Fig. 2 indicated
that the expression levels of PCNA (Fig. 2A) and Cyclin D1 (Fig. 2A) protein were decreased, and the
expression of Cleaved-caspase-3/caspase-3 (Fig. 2A) and p-AMPK/AMPK protein (Fig. 2B) were upregulated following
treatment with 25 and 50 µM emodin when compared with the control
group (P<0.05). These data suggested that emodin may serve a
role in the expression of the AMPK pathway and the inhibition of
TPC-1 cell proliferation and the cell cycle.
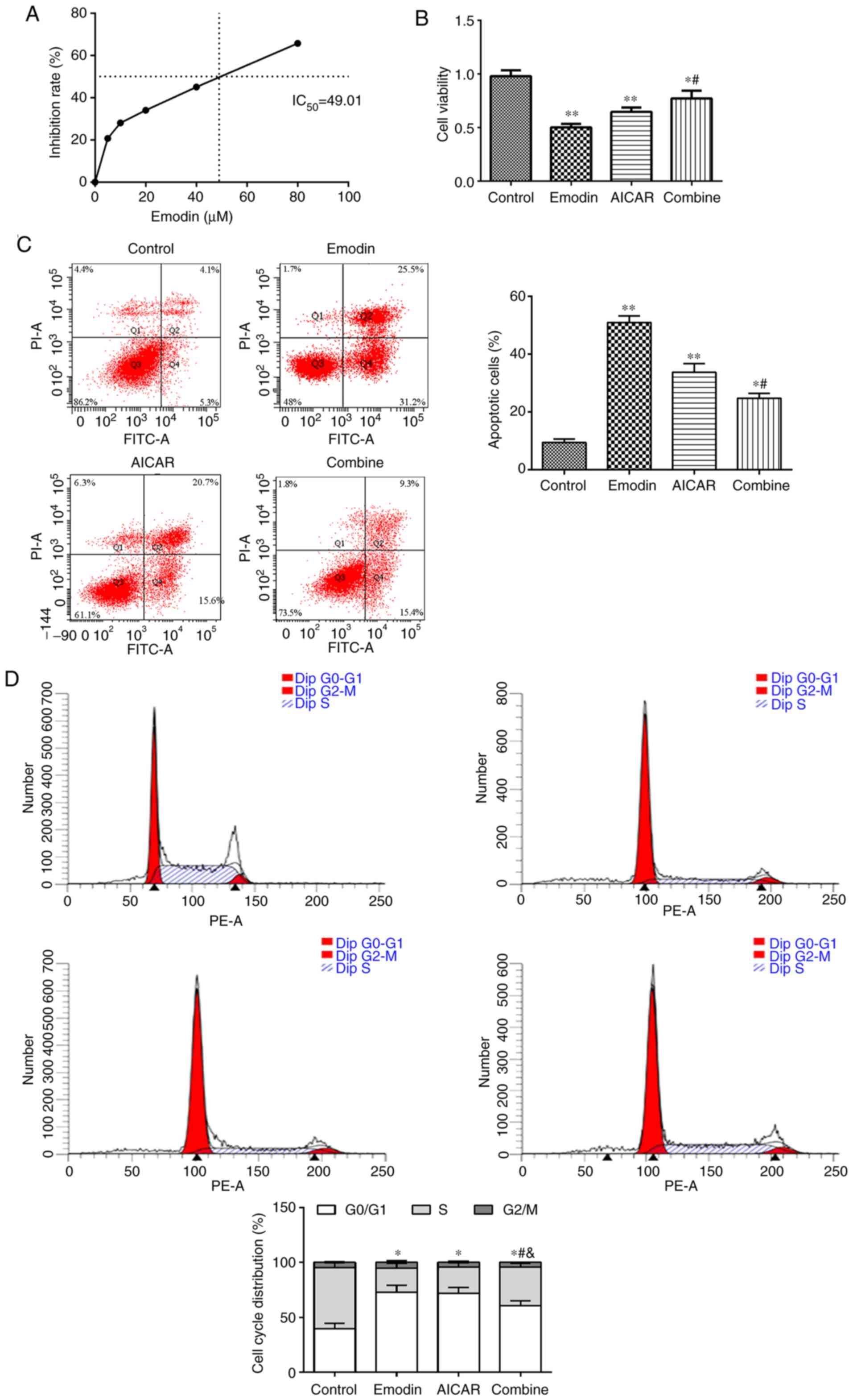
Emodin activates the AMPK pathway to
affect proliferation papillary thyroid cells
In order to elucidate whether emodin affects
proliferation and the cell cycle in TPC-1 cells via the AMPK
pathway, AMPK inhibitor (Dorsomorphin) and activator (AICAR) were
used to perform subsequent experiments. The experimental results
are presented in Fig. 3. The
IC50 of emodin was analyzed in Fig. 3A. When compared with the control
group, the proliferation of cells in the emodin and AICAR group
were reduced (Fig. 3A), the
apoptosis rate in TPC-1 cells was significantly increased and the
distribution was also markedly raised in the
G0/G1 phase (P<0.05; Fig. 3C and D). Compared with the emodin group, the
malignant biological behavior of the combined group was increased
(P<0.05).
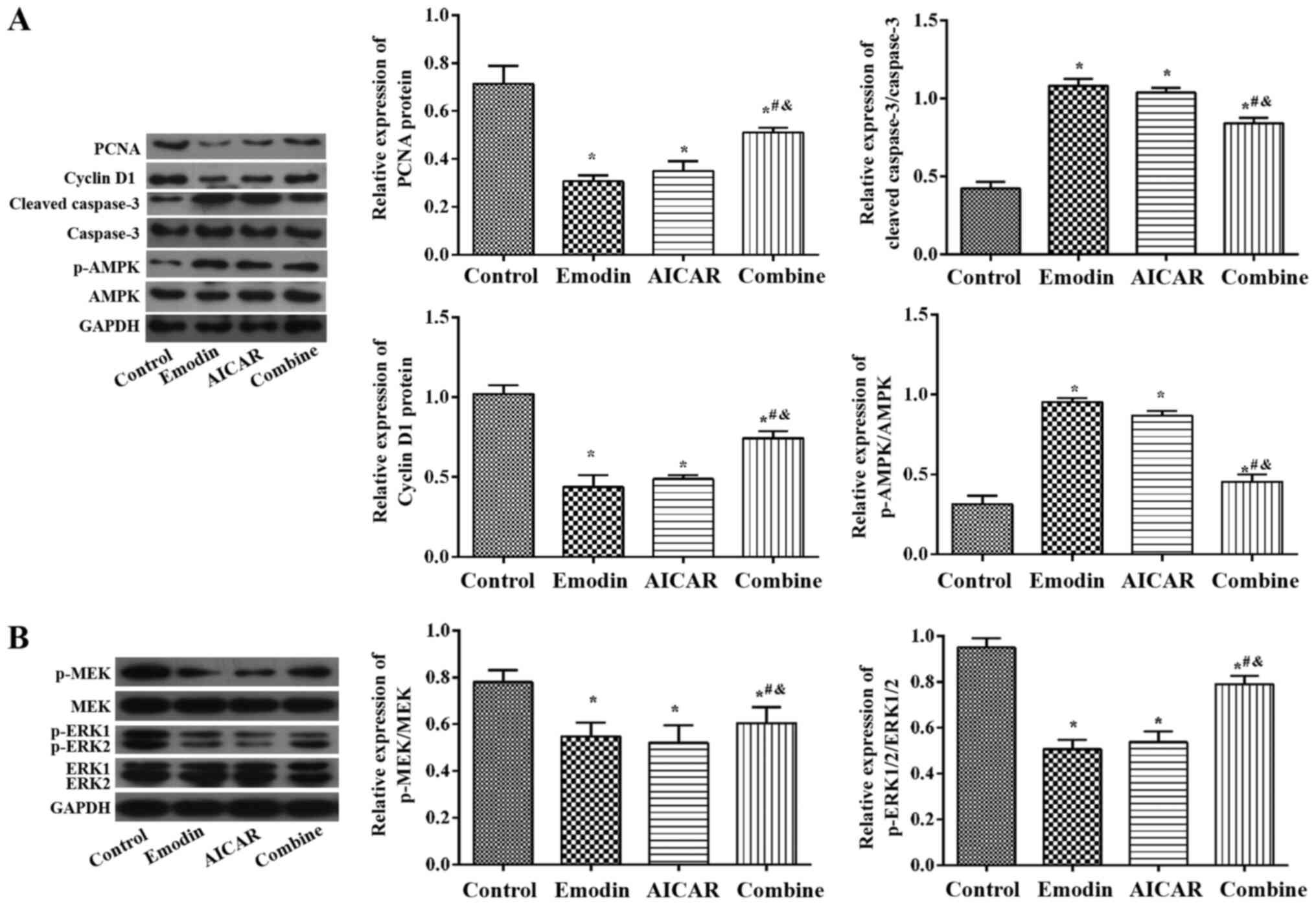
Emodin activates AMPK to affect the
MEK-ERK pathway
Fig. 4A indicated
that PCNA and Cyclin D1 proteins levels were downregulated, and the
levels of Cleaved-caspase-3/caspase-3 and p-AMPK/AMPK proteins were
upregulated in the emodin and AICAR group when compared with the
control group (P<0.05). However, there was no significant
difference between the two groups. Fig.
4B indicated that when compared with the control group,
p-ERK1/2 and p-MEK expression in the emodin and AICAR group were
decreased (P<0.05). In contrast to the emodin and AICAR group,
the two proteins in the combined group were markedly increased
(P<0.05). These results indicate that emodin activated AMPK to
downregulate the MEK-ERK pathway in TPC-1 cells.
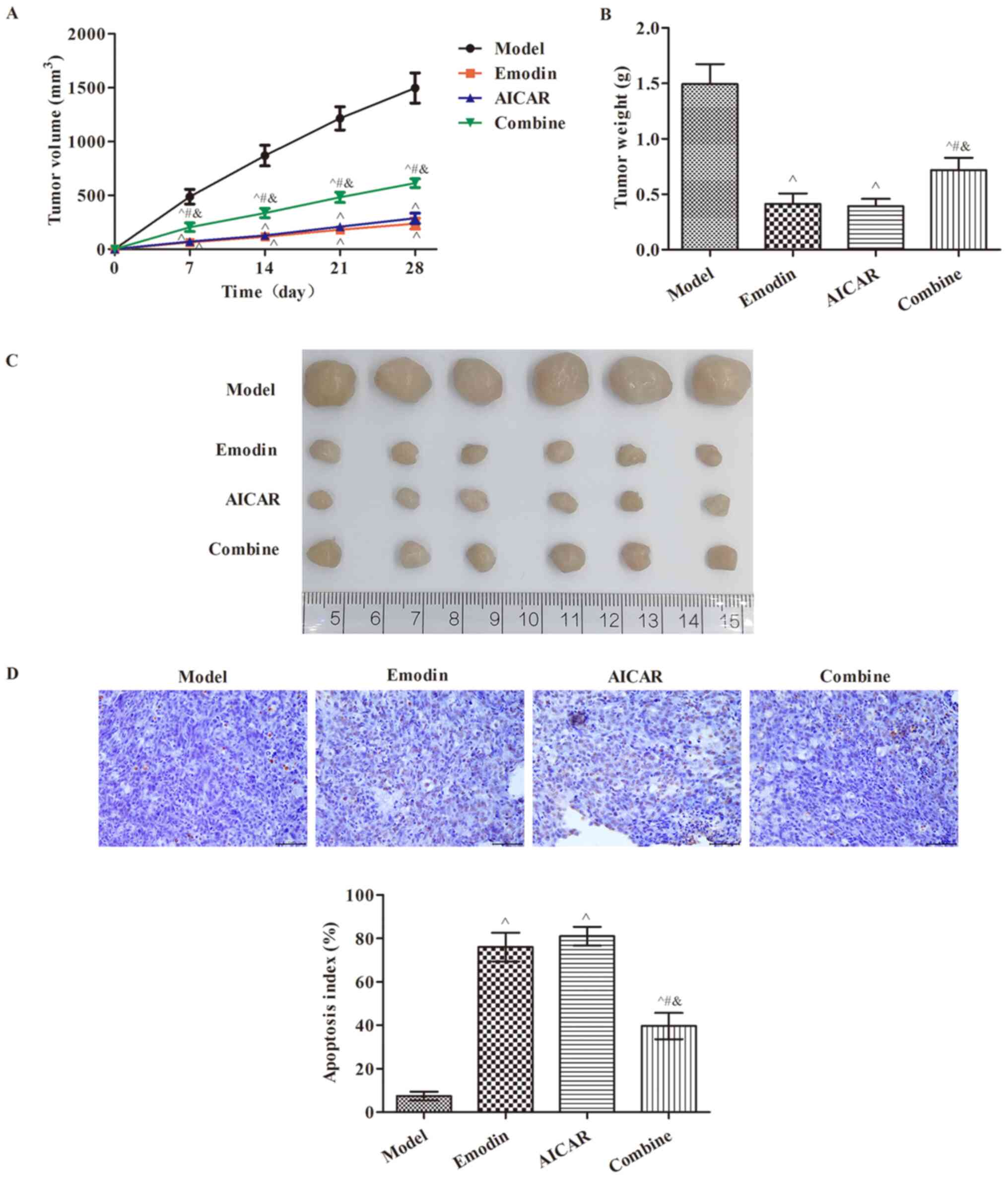
Emodin suppresses tumor growth via
activating AMPK
The mice model of tumor xenograft was established
using TPC-1 cells. Nude mice were randomly divided into the model,
emodin, AICAR and combined group. The tumor volume and tumor weight
in each group are presented in Fig.
5A-C. Consistent with the in vitro experiment, the
results revealed that emodin significantly suppressed tumor growth
in vivo when compared with the model mice (P<0.05). The
AMPK activator served a similar role to emodin, which suppressed
tumor growth. However, administration of the AMPK inhibitor
weakened the suppression of tumor growth caused by emodin
(P<0.05). Furthermore, the apoptosis index in tumor tissues of
each group indicated that emodin increased the number of apoptotic
cells compared with the model mice (P<0.05; Fig. 5D). These data suggested that emodin
may suppress papillary thyroid carcinoma via activating AMPK.
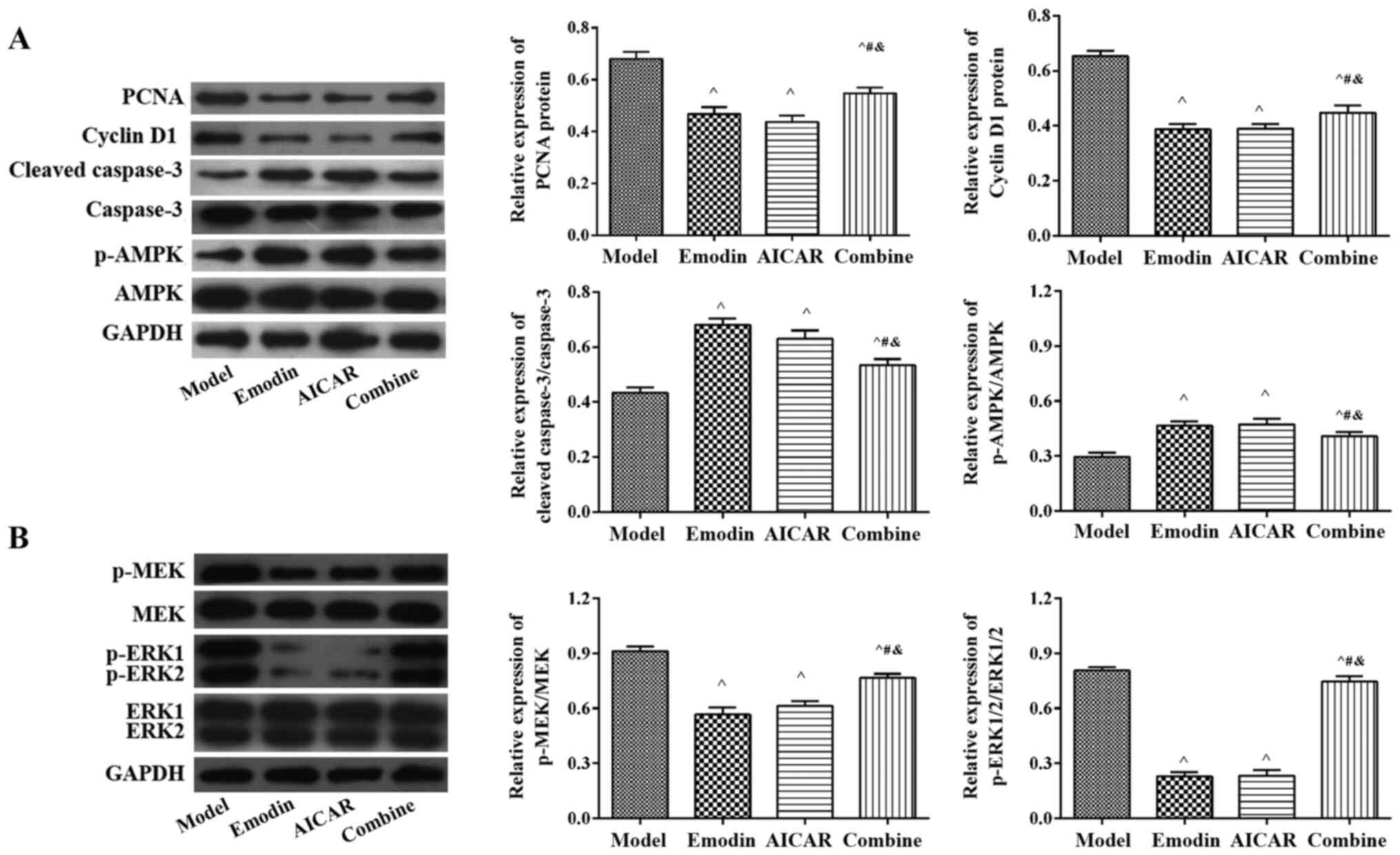
Emodin suppresses the MEK-ERK pathway
via activating AMPK
As presented in Fig.
6A, the expression of PCNA and Cyclin D1 significantly
decreased, while the expression of Cleaved-caspase-3/caspase-3 and
p-AMPK/AMPK increased in the emodin and AICAR group compared with
the model group (P<0.05). In addition, the expression of
p-ERK/ERK and p-MEK/MEK was significantly downregulated following
emodin or AICAR administration (P<0.05; Fig. 6B). In contrast to the emodin group,
the effect of emodin was significantly lower in the combined group
(P<0.05). These data revealed that emodin may suppress the
MEK-ERK pathway via activating AMPK.
Discussion
It is well known that emodin inhibits tumor growth
in a number of different cancer types, such as pancreatic cancer,
hepatocellular carcinoma and breast cancer (6,7,9). It
has also been reported that emodin can suppress PCNA expression in
some tumor cells, including those in cervical cancer (17). PCNA binds with human DNA polymerase
δ to serve a role in DNA duplication and repair, and is usually a
marker for cell proliferation and DNA replication (7). Consistent with a previous study
(17), the current study
demonstrated that emodin suppressed PCNA expression in TPC-1 cells.
Cell cycle interruption is a major feature of tumor transformation
and the status of the cell cycle represents a prognostic index for
cancer (18). Cyclin-D1, which is a
regulator of the G1/S cell cycle, serves an essential
role in tumor progression, contributes to tumor growth and resists
anticancer drugs (18). It was
therefore hypothesized that emodin would reduce the expression of
Cyclin-D1 expression in TPC-1 cells and in a xenograft model.
A number of studies have demonstrated that AMPK is a
cellular energy and nutritional status sensor in eukaryotic cells
that is associated with the mechanism targets of rapamycin
complex-1 (19-21).
AMPK is likely to be the earliest signaling pathway in a number of
evolutionary processes within eukaryotes (20). AMPK activation raises catabolism
production (ATP production) and represses anabolic rate (ATP
utilization). AMPK not only maintains the energy balance within the
cell, but also regulates energy metabolism throughout the body
(20). Due to its pivotal role in
maintaining energy homeostasis, AMPK has the potential to become a
underlying target for the treatment of metabolic diseases, which
makes it an interesting study target (21). In the present study, emodin was
indicated to be a cancer suppressor, and was revealed to suppress
the proliferation and cell cycle of TPC-1 cells, and to promote
cell apoptosis. Furthermore, the current study suggested that
emodin may activate the AMPK pathway to inhibit cell proliferation
and the cell cycle in human papillary thyroid carcinoma cells.
The MEK-ERK signaling cascade is activated by
tyrosine kinase, which is linked by its homologous growth factor
receptor under normal physiological conditions (22). MEK, which is a tyrosine-threonine
kinase, is being widely studied as a potential drug target
(22). A number of previous studies
have demonstrated that emodin may inhibit cancer proliferation in a
number of cancers via the ERK pathway, including in hepatocellular
carcinoma (23), non-small cell
lung cancer (8) and breast cancer
(9). The results of the current
study revealed that in human papillary thyroid carcinoma cells,
emodin decreased p-ERK1/2 and p-MEK expression by activating AMPK,
both in vitro and in vivo.
The association between emodin and AMPK pathway
remains to be further elucidated in future research. Although the
present study suggested that emodin may activate AMPK to inhibit
the activity of MEK-ERK pathway, the specific mechanism governing
this is still unclear and requires further study. In conclusion,
the results of the current study suggested that emodin inhibited
the proliferation of papillary thyroid cancer cells, and the
mechanism may be associated with the activation of AMPK.
Acknowledgements
Not applicable.
Funding
Funding: The current study was supported by the Project of
Yantai Science and Technology Plan (grant no. 2018SFGY113).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WL and DW carried out the experimental work and the
data collection and interpretation. WL, ML and DW participated in
the design and coordination of experimental work, and acquisition
of data. WL, DW and BL participated in the data collection,
analysis of data and preparation of the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The experiments were approved by The Animal
Protection and Use Committee of the Affiliated Yantai Yuhuangding
Hospital of Qingdao University (approval no. 2020-161).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sak SD: Variants of papillary thyroid
carcinoma: Multiple faces of a familiar tumor. Turk Patoloji Derg.
31 (Suppl 1):S34–S47. 2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Lam AK and Saremi N: Cribriform-morular
variant of papillary thyroid carcinoma: A distinctive type of
thyroid cancer. Endocr Relat Cancer. 24:R109–R121. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Ambrosi F, Righi A, Ricci C, Erickson LA,
Lloyd RV and Asioli S: Hobnail variant of papillary thyroid
carcinoma: A literature review. Endocr Pathol. 28:293–301.
2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Stoffer SS: Werner & Ingbar's The
Thyroid: A Fundamental and Clinical Text. 7th edition. Lippincott
Williams & Wilkins, Philadelphia, PA, 1996.
|
|
5
|
Su J, Yan Y, Qu J, Xue X, Liu Z and Cai H:
Emodin induces apoptosis of lung cancer cells through ER stress and
the TRIB3/NF-κB pathway. Oncol Rep. 37:1565–1572. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Li N, Wang C, Zhang P and You S: Emodin
inhibits pancreatic cancer EMT and invasion by up-regulating
microRNA-1271. Mol Med Rep. 18:3366–3374. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kim YS, Lee YM, Oh TI, Shin DH, Kim GH,
Kan SY, Kang H, Kim JH, Kim BM, Yim WJ and Lim JH: Emodin
sensitizes hepatocellular carcinoma cells to the anti-cancer effect
of sorafenib through suppression of cholesterol metabolism. Int J
Mol Sci. 19(3127)2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Tang Q, Wu J, Zheng F, Hann SS and Chen Y:
Emodin increases expression of insulin-like growth factor binding
protein 1 through activation of MEK/ERK/AMPKα and interaction of
PPARγ and Sp1 in lung cancer. Cell Physiol Biochem. 41:339–357.
2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Tseng HS, Wang YF, Tzeng YM, Chen DR, Liao
YF, Chiu HY and Hsieh WT: Aloe-emodin enhances tamoxifen
cytotoxicity by suppressing Ras/ERK and PI3K/mTOR in breast cancer
cells. Am J Chin Med. 45:337–350. 2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Tang Q, Zhao S, Wu J, Zheng F, Yang L, Hu
J and Hann SS: Inhibition of integrin-linked kinase expression by
emodin through crosstalk of AMPKα and ERK1/2 signaling and
reciprocal interplay of Sp1 and c-Jun. Cell Signal. 27:1469–1477.
2015.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Yu H, Xie Y, Zhou Z, Wu Z, Dai X and Xu B:
Curcumin regulates the progression of colorectal cancer via LncRNA
NBR2/AMPK pathway. Technol Cancer Res Treat.
18(1533033819870781)2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Chen D, Wu YX, Qiu YB, Wan BB, Liu G, Chen
JL, Lu MD and Pang QF: Hyperoside suppresses hypoxia-induced A549
survival and proliferation through ferrous accumulation via
AMPK/HO-1 axis. Phytomedicine. 67(153138)2020.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Samuels JS, Shashidharamurthy R and
Rayalam S: Novel anti-obesity effects of beer hops compound
xanthohumol: Role of AMPK signaling pathway. Nutr Metab (Lond).
15(42)2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Wu CC, Chen MS, Cheng YJ, Ko YC, Lin SF,
Chiu IM and Chen JY: Emodin inhibits EBV reactivation and represses
NPC tumorigenesis. Cancers (Basel). 11(1795)2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Tsai WL, Hsu CN and Tain YL: Whether AICAR
in pregnancy or lactation prevents hypertension programmed by high
saturated fat diet: A pilot study. Nutrients.
12(448)2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Hasanvand A, Amini-Khoei H, Hadian MR,
Abdollahi A, Tavangar SM, Dehpour AR, Semiei E and Mehr SE:
Anti-inflammatory effect of AMPK signaling pathway in rat model of
diabetic neuropathy. Inflammopharmacology. 24:207–219.
2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Guo JM, Xiao BX, Liu Q, Zhang S, Liu DH
and Gong ZH: Anticancer effect of aloe-emodin on cervical cancer
cells involves G2/M arrest and induction of differentiation. Acta
Pharmacol Sin. 28:1991–1995. 2007.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Hsu LC, Tu HF, Hsu FT, Yueh PF and Chiang
IT: Beneficial effect of fluoxetine on anti-tumor progression on
hepatocellular carcinoma and non-small cell lung cancer bearing
animal model. Biomed Pharmacother. 126(110054)2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Yasemin S, Peterson TR, Shaul YD,
Lindquist RA, Thoreen CC, Bar-Peled L and Sabatini DM: The Rag
GTPases bind raptor and mediate amino acid signaling to mTORC1.
Science. 320:1496–1501. 2008.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Hardie DG: AMPK-sensing energy while
talking to other signaling pathways. Cell Metab. 20:939–952.
2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Carling D: AMPK signalling in health and
disease. Curr Opin Cell Biol. 45:31–37. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Poulikakos PI, Zhang C, Bollag G, Shokat
KM and Rosen N: RAF inhibitors transactivate RAF dimers and ERK
signalling in cells with wild-type BRAF. Nature. 464:427–430.
2010.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Lin W, Zhong M, Yin H, Chen Y, Cao Q, Wang
C and Ling C: Emodin induces hepatocellular carcinoma cell
apoptosis through MAPK and PI3K/AKT signaling pathways in
vitro and in vivo. Oncol Rep. 36:961–967.
2016.PubMed/NCBI View Article : Google Scholar
|