Introduction
Rheumatoid arthritis (RA) is a systemic autoimmune
disease that mainly causes chronic inflammation in synovial tissues
(1,2). Accumulating evidence suggests that
fibroblast-like synoviocytes (FLS) play a critical role in the
pathogenesis of RA, particularly in the erosion of cartilage and
bone (3,4). Stable activated RA-FLS exhibit tumor
cell-like phenotypes, such as overproduction of inflammatory
cytokines, excessive proliferation, aggressive migration and
invasion (5,6). Therefore, regulating the migration and
invasion of RA-FLS may be useful for ameliorating joint destruction
in RA.
Platelet-derived extracellular vesicles (PEVs) are
heterogeneous vesicles, sized 0.1-1.0 µm, that are released from
platelet membranes and have been attracting substantial attention
(7). PEVs may play a role in
several pathological conditions, such as ischemic stroke,
cardiovascular diseases, cancer and inflammatory diseases (8-10).
It has been reported that the level of circulating PEVs is
significantly elevated in various autoimmune diseases, such as RA,
Sjogren's syndrome, systemic lupus erythematosus and
antiphospholipid syndrome (11-13).
Notably, the numbers of PEVs are increased in both the peripheral
blood and joint cavity of patients with RA, and they are associated
with disease activity, indicating that PEVs are closely associated
with the occurrence and development of RA (14,15).
Increasing evidence suggest that PEVs not only deliver several
bioactive molecules, including chemokines, enzymes and inflammatory
mediators, but also induce monocytes and endothelial cells to
release more inflammatory mediators to aggravate inflammatory
processes (16-18).
In addition, PEVs promote the proliferation, angiogenesis and
migration and invasion of tumor cells by increasing the expression
of MMPs (19-21).
However, the role of PEVs in the pathogenesis of RA remains
unclear.
Our previous study demonstrated that PEVs promote
the migration and invasion of RA-FLS (22). The present study investigated the
specific protein composition of PEVs by proteomics analysis and
examined the chemokine contents of PEVs, such as C-C motif
chemokine ligand 5 (CCL5), C-X-C motif chemokine ligand (CXCL)4 and
CXCL7. In addition, it was investigated whether SB225002, an
antagonist of C-X-C motif chemokine receptor 2 (CXCR2; a CXCL7
receptor), could inhibit the migration and invasion of RA-FLS
induced by PEVs, and whether these effects are mediated via
suppression of IκB and NF-κB phosphorylation (23). The aim was to determine whether
SB225002 can inhibit the motility of RA-FLS induced by PEVs, which
may be a potential therapeutic target for RA.
Materials and methods
Cell culture
Human RA-FLS were purchased from Jennio Biotech Co.,
Ltd. and maintained in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 15% FBS (HyClone; Cytiva), 100 U/ml penicillin
and 100 µg/ml streptomycin (Invitrogen; Thermo Fisher Scientific,
Inc.), at 37˚C with 5% CO2. Primary RA-FLS from passages
3-6 were used in our experiments.
PEVs preparation and component
analysis
Platelet-rich plasma (PRP), purchased from Red Cross
Blood Station (Yangzhou, China), was centrifuged at 1,000 x g for 5
min at room temperature. Washed platelets were prepared from PRP
and resuspended in modified Tyrode's buffer (HyClone; Cytiva). PEVs
were subsequently harvested by stimulating the platelets in washing
buffer (1 mM CaCl2, 2 mM MgCl2 and 10 µM ADP)
for 30 min at 37˚C with gentle agitation, removing platelet
aggregates at 3,000 x g for 30 min, followed by centrifugation at
15,000 x g for 1 h at 4˚C. Subsequent verification of PEVs was
assessed using PE-labeled anti-CD41 via flow cytometric analysis
(FACS CantoⅡ; Becton, Dickinson and Company), and the relative PEVs
concentration was quantified using the BCA method. Subsequently,
liquid chromatography with tandem mass spectrometry (LC-MS-MS)
detection and component analysis were performed by Shanghai Applied
Protein Technology Co., Ltd.
Bioinformatics
Gene Ontology (GO) and Kyoto Encyclopedia of Genes
and Genomes (KEGG) analysis were performed using the David 6.8
online tool (http://david.ncifcrf.gov/). GO analysis was
constituted with three domains: Biological process, cellular
component and molecular function. KEGG analysis was performed to
explore the signaling pathways of the differentially expressed
proteins.
Immunofluorescence staining
RA-FLS were cultured in complete DMEM supplemented
with 15% FBS with or without 50 µg/ml PEVs for 24 h, fixed with 4%
paraformaldehyde for 15 min at room temperature and permeabilized
with 0.5% Triton X-100 for 15 min at room temperature. The actin
cytoskeleton was visualized following incubation with
rhodamine-conjugated phalloidin (Sigma-Aldrich; Merck KGaA) for 2 h
in the dark. Nuclei were counterstained with DAPI (Beyotime
Institute of Biotechnology) for 10 min at 37˚C. Following thorough
washing with PBS, coverslips were mounted on glass slides and
micrographs were captured under a fluorescence microscope
(magnification, x100).
Cell viability assay
RA-FLS were seeded into 96-well plates at a density
of 5x103 cells/well for 24 h and subsequently treated
with different concentrations of several chemokine receptor
antagonists: BX471 (50, 100 and 150 nM; CCR1 antagonist,
Sigma-Aldrich; Merck KGaA), AMG487 (0.5, 1 and 2 µM; CXCR3
antagonist, Sigma-Aldrich; Merck KGaA) and SB225002 (0.1, 0.2 and
0.4 µM; CXCR2 antagonist, Sigma-Aldrich; Merck KGaA). Following
incubation for 24 h at 37˚C, 10 µl Cell Counting Kit-8 (CCK-8;
Absin Bioscience, Inc.) reagent was added to each well and the
optical density was measured at a wavelength of 450 nm.
Wound healing assay
RA-FLS were seeded into 6-well plates at a density
of 1x105 cells/well for 12 h. Following incubation with
serum-free DMEM for 12 h at 37˚C, linear scratches in the cell
monolayer were generated using a 200-µl pipette tip when cell
confluence reached about 80-90%. Subsequently, RA-FLS were cultured
in serum-free DMEM supplemented with different concentration of
PEVs (0 and 50 µg/ml) and chemokine receptor antagonists for 24 h.
Images were captured under an inverted microscope (magnification,
x100; Eclipse Ti; Nikon Corporation).
Transwell migration and invasion
assay
Transwell chambers were used to assess cell
migration and invasion. For the Transwell migration assay,
2x104 cells were plated in the upper chambers of
Transwell plates (pore size, 8.0 µm; Corning, Inc.) in serum-free
DMEM for 12 h, followed by incubation with different concentrations
of PEVs (0 and 50 µg/ml) for 24 h. The lower chamber was
supplemented with DMEM containing 15% FBS as chemoattractant with
corresponding PEVs and chemokine receptor antagonists. The inserts
were removed after 24 h and the non-migratory cells were gently
wiped off with a cotton swab. After fixation with 100% methanol for
2 min, the migrated cells were stained with 10% Giemsa solution
(Vazyme Biotech Co., Ltd.) for 15 min at room temperature and
counted in eight randomly selected fields using an inverted
microscope (magnification, x100; Eclipse Ti; Nikon Corporation).
For the invasion assay, the inserts were precoated with 100 µl
Matrigel (100 µg/ml; BD Biosciences) 24 h prior to the experiment
and the basement membranes were hydrated for 1 h at 37˚C.
Western blotting
Total protein was extracted from RA-FLS using the
protein extraction kit (Vazyme Biotech Co., Ltd.) and protein
concentration was quantified using the BCA assay method. Proteins
were separated via 10% SDS-PAGE, transferred onto PVDF membranes
(MilliporeSigma) and blocked with 5% non-fat milk in 0.05%
TBS-Tween-20 for 1 h at room temperature. The membranes were
incubated with primary antibodies against IκB (1:1,000; cat. no.
4812; Cell Signaling Technology, Inc.), phosphorylated (p)-IκB
(1:1,000; cat. no. 2859; Cell Signaling Technology, Inc.), NF-κB
(1:1,000; cat. no. 8242; Cell Signaling Technology, Inc.), p-NF-κB
(1:1,000; cat. no. 3033; Cell Signaling Technology, Inc.) and GAPDH
(1:1,000; cat. no. 2118; Cell Signaling Technology, Inc.) overnight
at 4˚C. Following the primary antibody incubation, the membranes
were incubated with the HRP-conjugated goat anti-rabbit polyclonal
IgG secondary antibody (1:2,000; cat. no. 7074; Cell Signaling
Technology, Inc.). Protein bands were visualized by Pierce ECL Plus
Western Blotting substrate (Thermo Fisher Scientific, Inc.) and
subsequently analyzed using ImageJ software (version 1.51j8;
National Institutes of Health). GAPDH was used as the internal
control.
Statistical analysis
Statistical analysis was performed using SPSS 20.0
software (IBM Corp.). All experiments were performed in triplicate
and data are presented as the mean ± SD. The two-tailed paired
Student's t-test was used to compare the differences between two
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
Identification of chemokines in
PEVs
LC-MS-MS demonstrated that there were 5,256 proteins
in PEVs, and only proteins with >4 distinct peptides and >20%
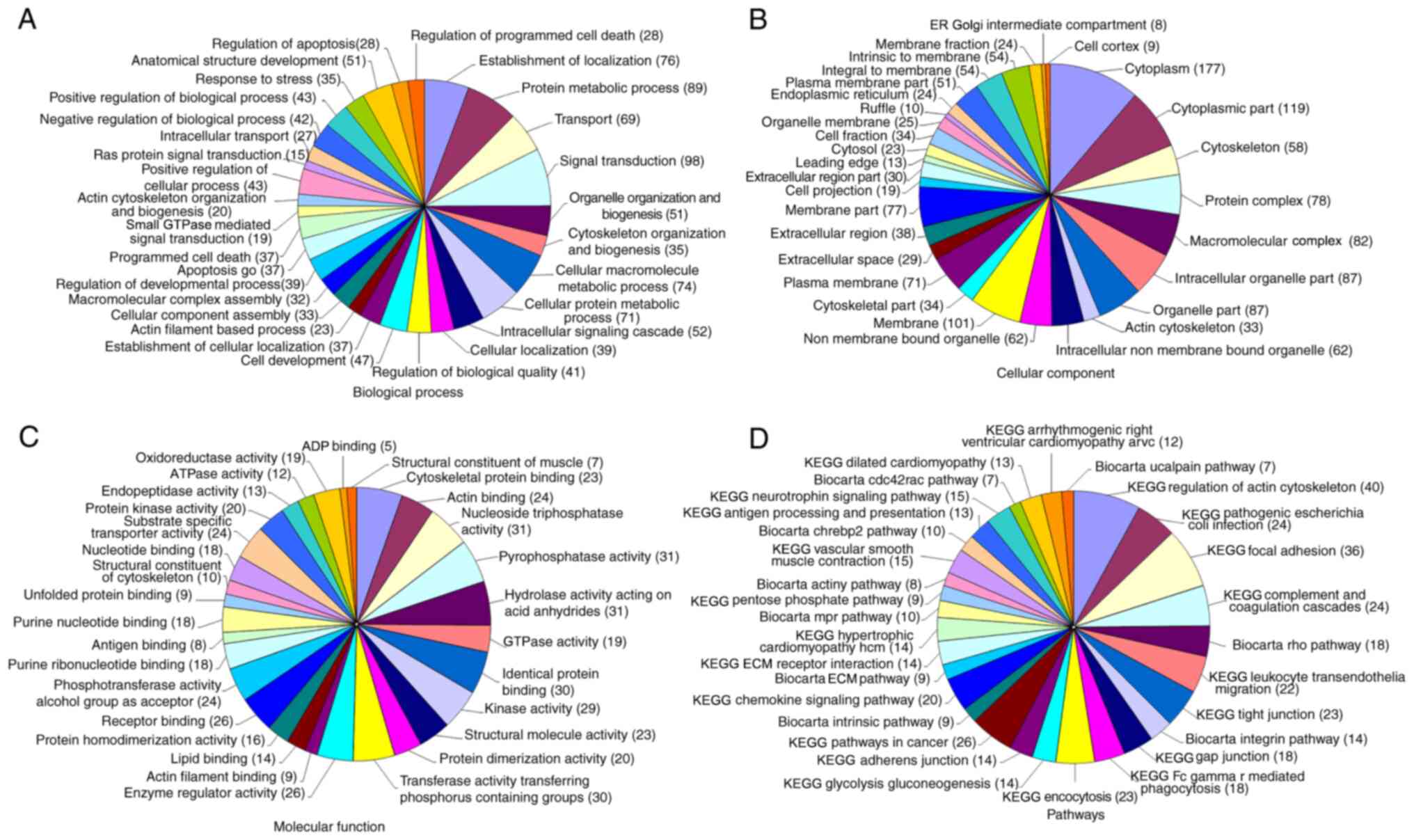
coverage were considered as significant (24,25).
GO enrichment analysis with respect to these proteins in PEVs was
performed to determine the biological process, cellular component
and molecular function. KEGG pathway annotation demonstrated that
these proteins were enriched in the ‘regulation of cytoskeleton
actin’, ‘focal adhesion’ and ‘chemokine signaling pathways’,
suggesting that PEVs may participate in several pathophysiological
processes (Fig. 1). Notably, three
of the chemokines in PEVs (CCL5, CXCL4 and CXCL7) were involved in
chemokine signaling pathways.
PEVs promotes reorganization of the
actin cytoskeleton of RA-FLS
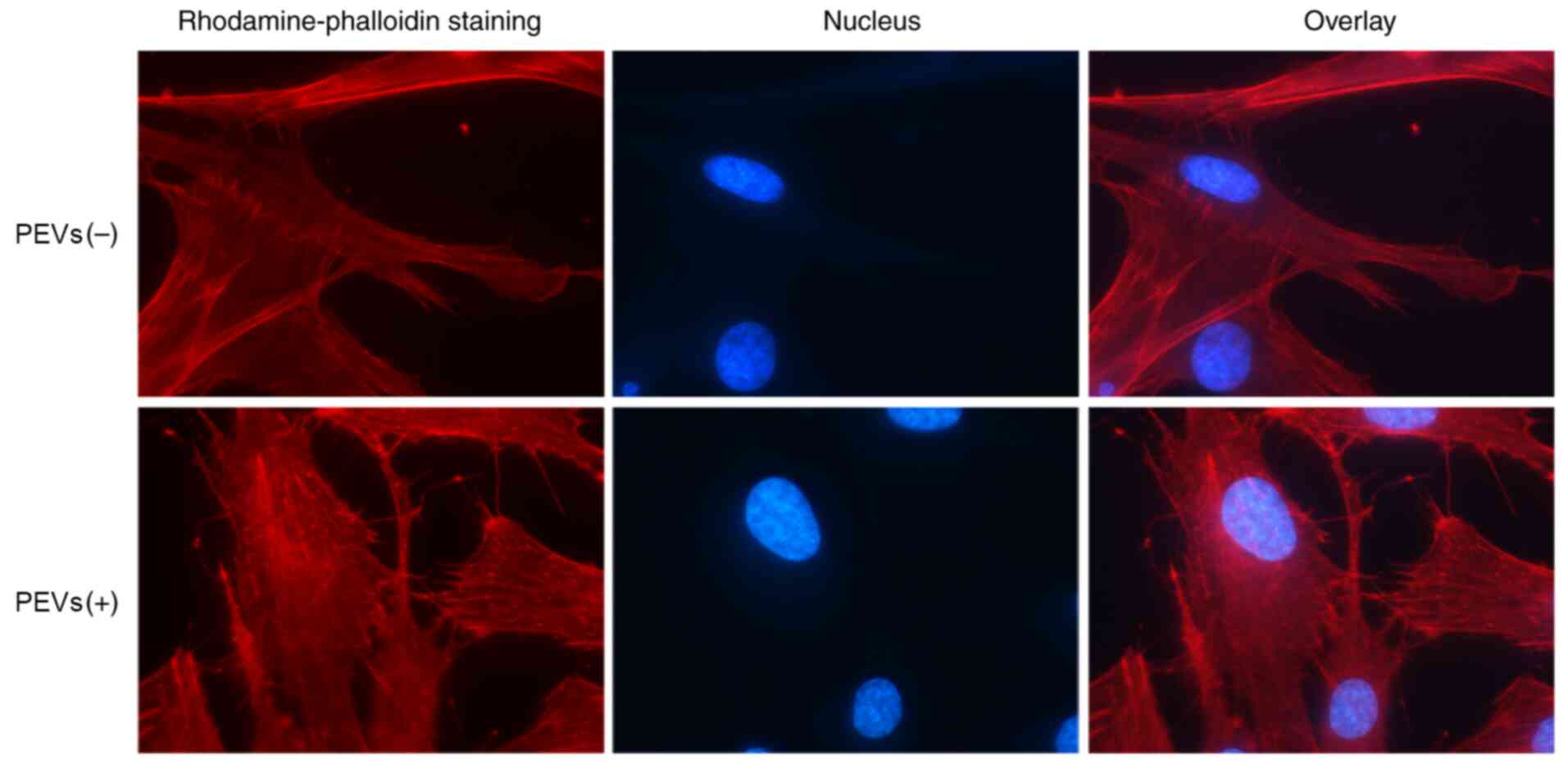
It has been reported that PEVs can promote the
motility of RA-FLS. Considering that the dynamic reorganization of
the actin cytoskeleton is critical for directional cell migration
(22,26), additional fluorescent phalloidin
staining was performed in the present study to determine whether
cytoskeletal changes are induced by PEVs. As shown in Fig. 2, treatment with PEVs increased the
number of fibers in cells, and facilitated lamellipodia and
filopodia formation at the leading edge of migrating cells,
suggesting that PEVs indeed promote cell invasion and
migration.
Effects of chemokine receptor
antagonists on the migration and invasion of RA-FLS induced by
PEVs
Considering that chemokines play important roles by
binding to their respective receptors (CCL5 receptor CCR1, CXCL4
receptor CXCR3 and CXCL7 receptor CXCR2), the following
corresponding chemokine receptor antagonists were investigated in
the present study: BX471 (CCR1 antagonist), AMG487 (CXCR3
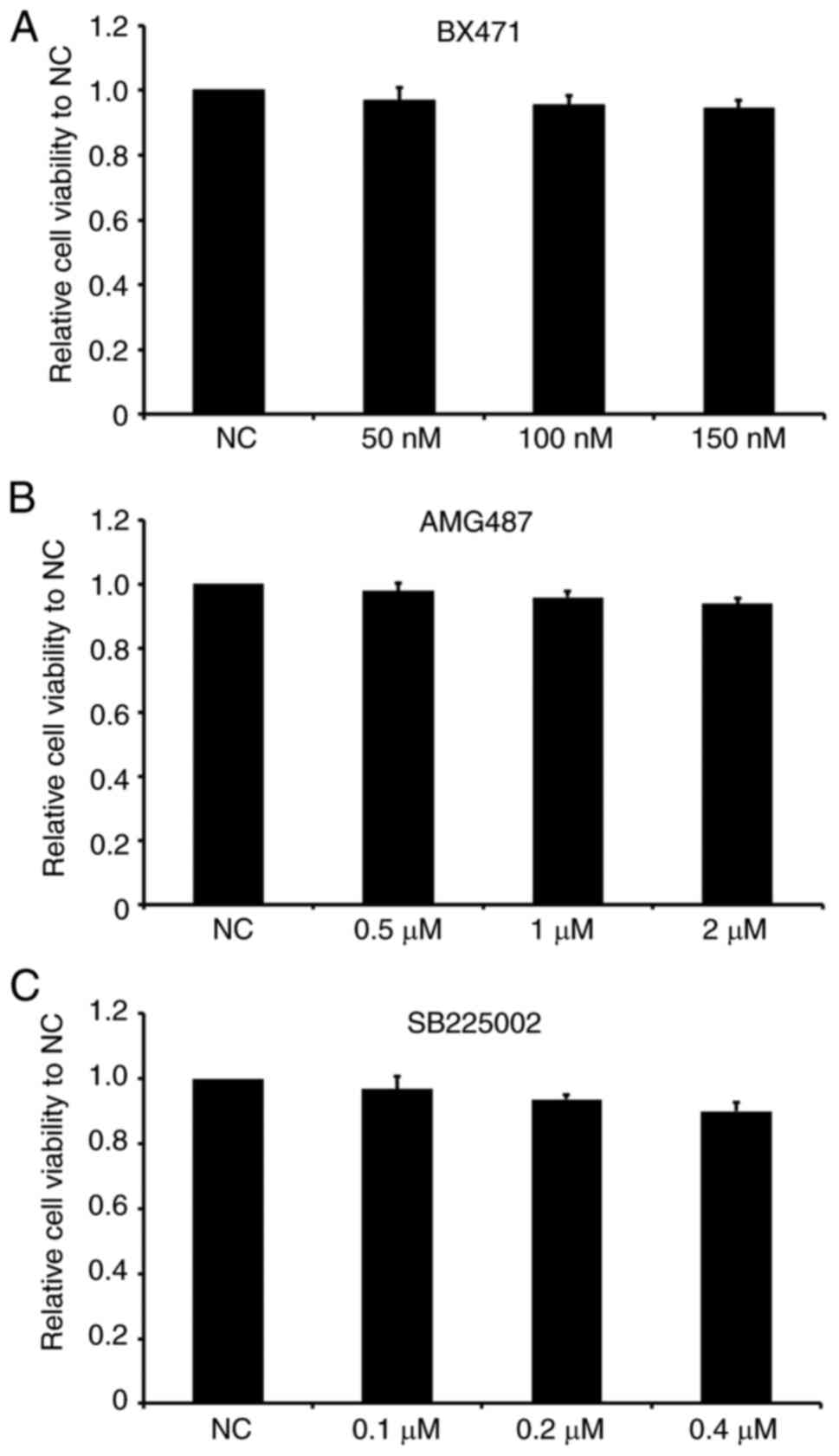
antagonist) and SB225002 (CXCR2 antagonist). First, the effects of
the three antagonists on the viability of RA-FLS were investigated.
As presented in Fig. 3, different
concentrations of BX471 (50, 100 and 150 nM), AMG487 (0.5, 1 and 2
µM) and SB225002 (0.1, 0.2 and 0.4 µM) exerted no significant
effects on cell viability compared with the control group,
suggesting that the chemokine receptor antagonists themselves did
not affect the viability of RA-FLS within the range of detected
concentrations.
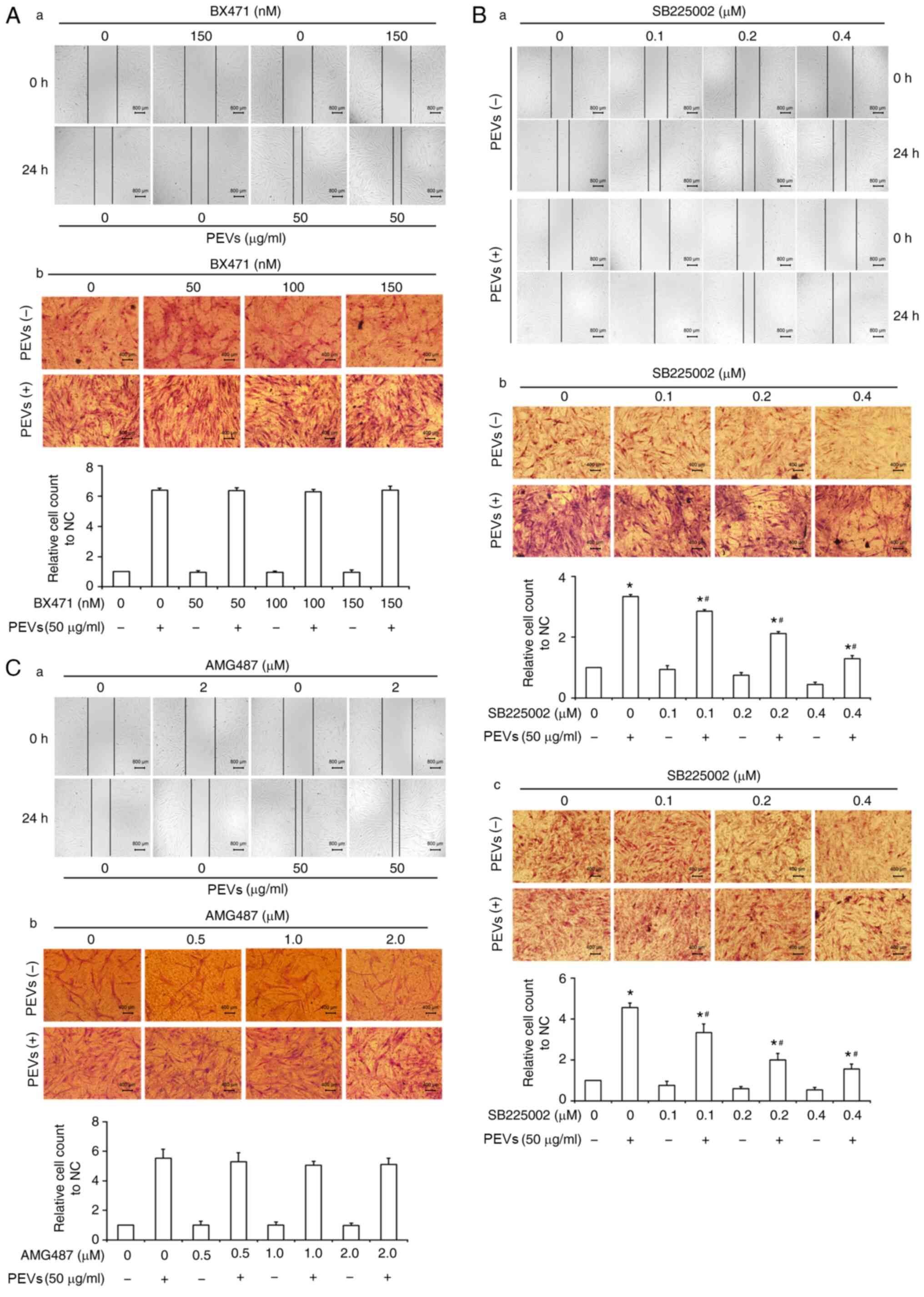
The migration and invasion of RA-FLS are crucial
characteristics that are associated with cartilage and bone erosion
during RA (5,6). To assess the effects of the three
chemokine receptor antagonists on the invasive and migratory
abilities of RA-FLS in the presence or absence of PEVs, wound
healing assay, and Transwell migration and invasion assays were
performed. As presented in Fig. 4,
PEVs significantly promoted the migration of RA-FLS, which is
consistent with previous findings (22). However, SB225002 was demonstrated to
partially antagonize the migration of RA-FLS induced by PEVs,
whereas no significant effects of BX471 or AMG487 on the migration
of RA-FLS were observed, in the presence or absence of PEVs. The
effect of SB225002 on the invasion of RA-FLS in the presence or
absence of PEVs was also investigated. As expected, the results of
the Transwell invasion assay demonstrated that SB225002 partially
antagonized the invasion of RA-FLS induced by PEVs, suggesting that
PEVs may affect the motility of RA-FLS via a CXCR2-mediated
signaling pathway.
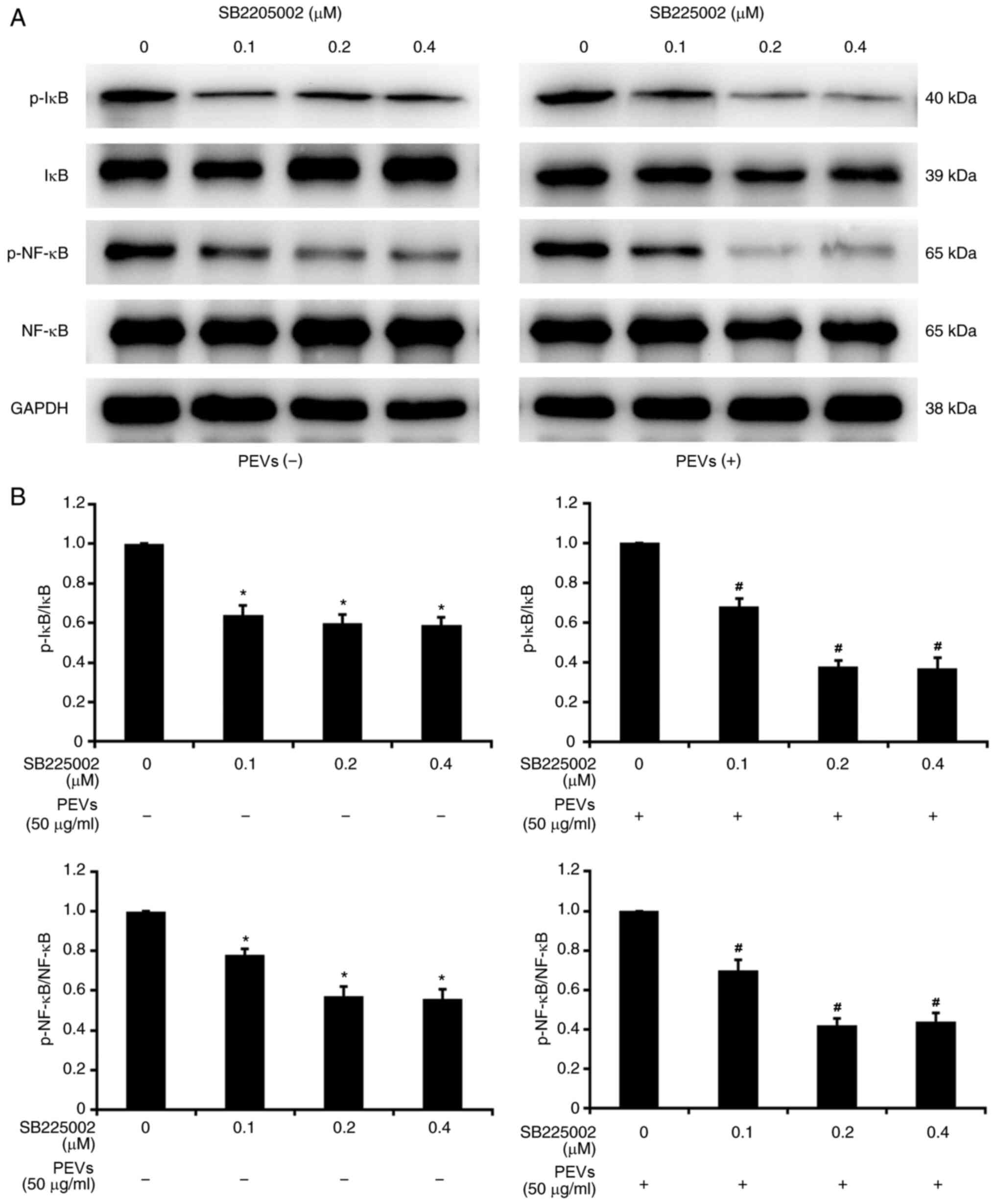
PEVs activated the CXCR2-mediated
NF-κB pathway in RA-FLS
It has been reported that PEVs may promote the
migration and invasion of RA-FLS by upregulating MMP-1 expression
via activation of ERK/NF-κB signaling (22). Considering that SB225002 partially
inhibits migration and invasion of RA-FLS induced by PEVs, as
mentioned above, it was next investigated whether PEVs activate
NF-κB via the CXCR2-mediated signaling pathway. As presented in
Fig. 5, SB225002 at different
concentrations (0.1, 0.2 and 0.4 µM) markedly decreased the
phosphorylation of IκB and NF-κB in RA-FLS. Of note, this tendency
was more significant following treatment with PEVs, suggesting that
SB225002 can inhibit the activating effect of PEVs on NF-κB
signaling in RA-FLS. Taken together, these results suggest that
PEVs may regulate the migration and invasion of RA-FLS via
CXCR2-mediated activation of the NF-κB pathway.
Discussion
RA-FLS, the dominant non-immune cells of synovial
tissues in patients with RA, contribute to the development of
synovitis, pannus formation and joint destruction via multiple
mechanisms (3). Increasing evidence
suggests that migration and invasion of RA-FLS play important roles
in RA initiation and progression (4-6).
As regards the promoting effect of PEVs on the motility of RA-FLS,
the present study demonstrated that PEVs regulate the actin
cytoskeletal reorganization in RA-FLS, which further verified the
active role of PEVs in cell motility, consistent with previous
findings (22). To identify the
main active contents and determine the molecular mechanism through
which PEVs regulate the motility of RA-FLS, LC-MS-MS analysis was
performed to identify the proteins of PEVs, which included several
significant chemokines, such as CCL5, CXCL4 and CXCL7. Previous
studies have demonstrated that these three chemokines are involved
in chemokine signaling pathways that are closely associated with
cell migration and invasion (27-29).
According to GO analysis, the biological processes these chemokines
participate in principally include ‘signal transduction’,
‘transport’, ‘establishment of localization’, ‘regulation of
developmental process’ and ‘negative regulation of biological
process’; the molecular functions include ‘substrate-specific
transporter activation’ and ‘receptor binding’. KEGG annotation
revealed that these chemokines are mainly involved in the
‘chemokine signaling pathway’.
Considering that chemokines play important roles in
autoimmune diseases, tumor-related inflammation and immunity, as
well as tumor growth and metastasis, it was hypothesized that the
chemokines in PEVs can affect the motility of RA-FLS (30-32).
In RA, CCL5 and its receptor, CCR1, are abundantly expressed in
synovial tissue and involved in monocyte and T lymphocyte
recruitment to the joints (33).
CXCL4/CXCR3 may be involved in lymphocyte chemotaxis to target
organs in patients with systemic lupus erythematosus, and have been
reported to be associated with disease activity (34). Notably, higher levels of synovial
CXCL4 and CXCL7 have been detected in early RA compared with
resolving arthritis or established RA (35). Taken together, these results suggest
that the chemokine/chemokine receptor axis may be a suitable target
for disease treatment. Chemokines and their receptors have been
implicated in inflammatory cell recruitment and angiogenesis, which
underlie the pathogenesis of RA (36). To verify whether PEVs modulate the
motility of RA-FLS via the chemokine/chemokine receptor pathway,
BX471 (CCR1 antagonist), AMG487 (CXCR3 antagonist) and SB225002
(CXCR2 antagonist) were selected in the present study. None of
these antagonists exerted significant effects on the viability of
RA-FLS. The results from the wound healing and Transwell assays
demonstrated that BX471 and AMG487 were unable to block the
migration of RA-FLS induced by PEVs. Conversely, SB225002 partially
antagonized the migration and invasion of RA-FLS induced by PEVs,
suggesting that PEVs may promote the migration and invasion of
RA-FLS via a CXCR2-mediated signaling pathway.
NF-κB is activated by several agents, including
cytokines, oxidant free radicals, bacterial or viral products and
ultraviolet irradiation (37).
Since PEVs presumably play a promoting role in the regulation of
motility of RA-FLS by activating NF-κB signaling, the present study
investigated whether the activation of NF-κB signaling was mediated
by CXCR2. As expected, the results confirmed that SB225002
decreased the phosphorylation of IκB and NF-κB in RA-FLS induced by
PEVs, rather than affecting CXCR2 expression. When the CXCL7/CXCR2
axis in RA-FLS is stimulated, the extracellular signal is
transmitted to the cytoplasm, which triggers the phosphorylation of
I-κB, which is degraded by the proteasome. Subsequently, NF-κB is
released and transferred into the nucleus, initiating transcription
of related genes, including inflammatory cytokines, chemokines and
MMPs, resulting in the malignant transformation and metastasis of
cells (38,39).
In conclusion, understanding the role of the main
active contents of PEVs in the occurrence and development of RA may
be crucial for exploring therapeutic targets. The findings of the
present study demonstrated that the CXCR2 antagonist exerted an
antagonistic effect against PEVs by decreasing IκB and NF-κB
phosphorylation in RA-FLS, indicating that CXCL7/CXCR2 may be a
potential therapeutic target for RA. However, further studies on
specific downstream factors of this signaling pathway and
verification in animal models are required to further elucidate the
role of PEVs in RA and develop novel therapeutic strategies.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant no. 81470070) and Nantong
Science and Technology Bureau (grant no. JC2020037).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ and WS were responsible for the concept and
design of the study. WW, ZD, GL and JY performed experiments and
data analysis. WW, WZ and CZ performed data interpretation,
presentation and writing of the manuscript. WW and YZ confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Smolen JS, Aletaha D, Barton A, Burmester
GR, Emery P, Firestein GS, Kavanaugh A, McInnes IB, Solomon DH,
Strand V and Yamamoto K: Rheumatoid arthritis. Nat Rev Dis Primers.
4(18001)2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Román-Fernández IV, García-Chagollán M,
Cerpa-Cruz S, Jave-Suárez LF, Palafox-Sánchez CA, García-Arellano
S, Sánchez-Zuno GA and Muñoz-Valle JF: Assessment of CD40 and CD40L
expression in rheumatoid arthritis patients, association with
clinical features and DAS28. Clin Exp Med. 19:427–437.
2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bartok B and Firestein GS: Fibroblast-like
synoviocytes: Key effector cells in rheumatoid arthritis. Immunol
Rev. 233:233–255. 2010.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Kawaguchi Y, Waguri-Nagaya Y, Tatematsu N,
Oguri Y, Kobayashi M, Nozaki M, Asai K, Aoyama M and Otsuka T: The
Janus kinase inhibitor tofacitinib inhibits TNF-α-induced
gliostatin expression in rheumatoid fibroblast-like synoviocytes.
Clin Exp Rheumatol. 36:559–567. 2018.PubMed/NCBI
|
|
5
|
Schönfeld C, Pap T, Neumann E and
Müller-Ladner U: Fibroblasts as pathogenic cells in rheumatic
inflammation. Z Rheumatol. 74:33–38. 2015.PubMed/NCBI View Article : Google Scholar : (In German).
|
|
6
|
Müller-Ladner U, Pap T, Gay RE, Neidhart M
and Gay S: Mechanisms of disease: The molecular and cellular basis
of joint destruction in rheumatoid arthritis. Nat Clin Pract
Rheumatol. 1:102–110. 2005.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ponomareva AA, Nevzorova TA, Mordakhanova
ER, Andrianova IA, Rauova L, Litvinov RI and Weisel JW:
Intracellular origin and ultrastructure of platelet-derived
microparticles. J Thromb Haemost. 15:1655–1667. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Rosińska J, Łukasik M and Kozubski W: The
impact of vascular disease treatment on Platelet-Derived
Microvesicles. Cardiovasc Drugs Ther. 31:627–644. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Vismara M, Zarà M, Negri S, Canino J,
Canobbio I, Barbieri SS, Moccia F, Torti M and Guidetti GF:
Platelet-derived extracellular vesicles regulate cell cycle
progression and cell migration in breast cancer cells. Biochim
Biophys Acta Mol Cell Res. 1868(118886)2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Vajen T, Mause SF and Koenen RR:
Microvesicles from platelets: Novel drivers of vascular
inflammation. Thromb Haemost. 114:228–236. 2015.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Sellam J, Proulle V, Jüngel A, Ittah M,
Miceli Richard C, Gottenberg JE, Toti F, Benessiano J, Gay S,
Freyssinet JM and Mariette X: Increased levels of circulating
microparticles in primary Sjögren's syndrome, systemic lupus
erythematosus and rheumatoid arthritis and relation with disease
activity. Arthritis Res Ther. 11(R156)2009.PubMed/NCBI View
Article : Google Scholar
|
|
12
|
Olumuyiwa-Akeredolu OO, Page MJ, Soma P
and Pretorius E: Platelets: Emerging facilitators of cellular
crosstalk in rheumatoid arthritis. Nat Rev Rheumatol. 15:237–248.
2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Chaturvedi S, Cockrell E, Espinola R, His
L, Fulton S, Khan M, Li L, Fonseca F, Kundu S and McCrae KR:
Circulating microparticles in patients with antiphospholipid
antibodies: Characterization and associations. Thromb Res.
135:102–108. 2015.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Knijff-Dutmer EA, Koerts J, Nieuwland R,
Kalsbeek-Batenburg EM and van de Laar MA: Elevated levels of
platelet microparticles are associated with disease activity in
rheumatoid arthritis. Arthritis Rheum. 46:1498–1503.
2002.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Boilard E, Nigrovic PA, Larabee K, Watts
GF, Coblyn JS, Weinblatt ME, Massarotti EM, Remold-O'Donnell E,
Farndale RW, Ware J and Lee DM: Platelets amplify inflammation in
arthritis via collagen-dependent microparticle production. Science.
327:580–583. 2010.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Puddu P, Puddu GM, Cravero E, Muscari S
and Muscari A: The involvement of circulating microparticles in
inflammation, coagulation and cardiovascular diseases. Can J
Cardiol. 26:140–145. 2010.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Italiano JE Jr, Mairuhu AT and Flaumenhaft
R: Clinical relevance of microparticles from platelets and
megakaryocytes. Curr Opin Hematol. 17:578–584. 2010.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Villar-Vesga J, Grajales C, Burbano C,
Vanegas-García A, Muñoz-Vahos CH, Vásquez G, Rojas M and Castaño D:
Platelet-derived microparticles generated in vitro resemble
circulating vesicles of patients with rheumatoid arthritis and
activate monocytes. Cell Immunol. 336:1–11. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Dashevsky O, Varon D and Brill A:
Platelet-derived microparticles promote invasiveness of prostate
cancer cells via upregulation of MMP-2 production. Int J Cancer.
124:1773–1777. 2009.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Janowska-Wieczorek A, Wysoczynski M,
Kijowski J, Marquez-Curtis L, Machalinski B, Ratajczak J and
Ratajczak MZ: Microvesicles derived from activated platelets induce
metastasis and angiogenesis in lung cancer. Int J Cancer.
113:752–760. 2005.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Barteneva NS, Fasler-Kan E, Bernimoulin M,
Stern JN, Ponomarev ED, Duckett L and Vorobjev IA: Circulating
microparticles: Square the circle. BMC Cell Biol.
14(23)2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wang W, Liu J, Yang B, Ma Z, Liu G, Shen W
and Zhang Y: Modulation of platelet-derived microparticles to
adhesion and motility of human rheumatoid arthritis fibroblast-like
synoviocytes. PLoS One. 12(e0181003)2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Grépin R, Guyot M, Giuliano S, Boncompagni
M, Ambrosetti D, Chamorey E, Scoazec JY, Negrier S, Simonnet H and
Pagès G: The CXCL7/CXCR1/2 axis is a key driver in the growth of
clear cell renal cell carcinoma. Cancer Res. 74:873–883.
2014.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Markov DA, Savkina M, Anikin M, Del Campo
M, Ecker K, Lambowitz AM, De Gnore JP and McAllister WT:
Identification of proteins associated with the yeast mitochondrial
RNA polymerase by tandem affinity purification. Yeast. 26:423–440.
2009.PubMed/NCBI View
Article : Google Scholar
|
|
25
|
Nadar M, Chan MY, Huang SW, Huang CC,
Tseng JT and Tsai CH: HuR binding to AU-rich elements present in
the 3' untranslated region of Classical swine fever virus. Virol J.
8(340)2011.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Sun BO, Fang Y, Li Z, Chen Z and Xiang J:
Role of cellular cytoskeleton in epithelial-mesenchymal transition
process during cancer progression. Biomed Rep. 3:603–610.
2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
An G, Wu F, Huang S, Feng L, Bai J, Gu S
and Zhao X: Effects of CCL5 on the biological behavior of breast
cancer and the mechanisms of its interaction with tumor-associated
macrophages. Oncol Rep. 42:2499–2511. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Quemener C, Baud J, Boyé K, Dubrac A,
Billottet C, Soulet F, Darlot F, Dumartin L, Sire M, Grepin R, et
al: Dual roles for CXCL4 chemokines and CXCR3 in angiogenesis and
invasion of pancreatic cancer. Cancer Res. 76:6507–6519.
2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Guo Q, Jian Z, Jia B and Chang L: CXCL7
promotes proliferation and invasion of cholangiocarcinoma cells.
Oncol Rep. 37:1114–1122. 2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Szekanecz Z and Koch AE: Successes and
failures of chemokine-pathway targeting in rheumatoid arthritis.
Nat Rev Rheumatol. 12:5–13. 2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Miyabe Y, Lian J, Miyabe C and Luster AD:
Chemokines in rheumatic diseases: Pathogenic role and therapeutic
implications. Nat Rev Rheumatol. 15:731–746. 2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Karin N and Razon H: Chemokines beyond
chemo-attraction: CXCL10 and its significant role in cancer and
autoimmunity. Cytokine. 109:24–28. 2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Haringman JJ, Smeets TJ, Reinders-Blankert
P and Tak PP: Chemokine and chemokine receptor expression in paired
peripheral blood mononuclear cells and synovial tissue of patients
with rheumatoid arthritis, osteoarthritis, and reactive arthritis.
Ann Rheum Dis. 65:294–300. 2006.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Im CH, Park JA, Kim JY, Lee EY, Lee EB,
Kim Y and Song YW: CXCR3 polymorphism is associated with male
gender and pleuritis in patients with systemic lupus erythematosus.
Hum Immunol. 75:466–469. 2014.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Yeo L, Adlard N, Biehl M, Juarez M,
Smallie T, Snow M, Buckley CD, Raza K, Filer A and Scheel-Toellner
D: Expression of chemokines CXCL4 and CXCL7 by synovial macrophages
defines an early stage of rheumatoid arthritis. Ann Rheum Dis.
75:763–771. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Szekanecz Z, Koch AE and Tak PP: Chemokine
and chemokine receptor blockade in arthritis, a prototype of
immune-mediated inflammatory diseases. Neth J Med. 69:356–366.
2011.PubMed/NCBI
|
|
37
|
DiDonato JA, Mercurio F and Karin M: NF-κB
and the link between inflammation and cancer. Immunol Rev.
246:379–400. 2012.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Dong YL, Kabir SM, Lee ES and Son DS:
CXCR2-driven ovarian cancer progression involves upregulation of
proinflammatory chemokines by potentiating NF-κB activation via
EGFR-transactivated Akt signaling. PLoS One.
8(e83789)2013.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhang Z, Tan X, Luo J, Cui B, Lei S, Si Z,
Shen L and Yao H: GNA13 promotes tumor growth and angiogenesis by
upregulating CXC chemokines via the NF-κB signaling pathway in
colorectal cancer cells. Cancer Med. 7:5611–5620. 2018.PubMed/NCBI View Article : Google Scholar
|