Introduction
Stroke ranks fifth among all causes of human death
and most patients have a stroke survive with serious long-term
disability (1,2). Ischemic stroke caused by blocked blood
supply is observed in ~87% of all patients with stroke. At present,
thrombolytic therapy with recombinant tissue plasminogen activator
is recommended by guidelines for the treatment of grade A stroke in
numerous countries. However, the effectiveness of this therapy
depends on the limited time window of stroke symptoms. Currently,
post stroke rehabilitation is still an effective way to treat
stroke (3). Thus, restoring blood
flow and recovering injured neurons are considered to be crucial
methods for the treatment of stroke.
Sodium tanshinone IIA sulfonate (STS) is a
water-soluble substance obtained by sulfonating the diterpenoid
quinone compound tanshinone IIA, which is isolated from Salvia
miltiorrhiza, a plant of the Labiatae family (4). Tanshinone IIA has poor water
solubility; therefore, STS was developed to increase
bioavailability and has been successfully used to treat patients
with cardiovascular diseases, cerebrovascular disease and nervous
system diseases such as Alzheimer's disease and cerebral ischemia
(5-7).
STS may improve myocardial injury mainly by inhibiting oxidative
stress (8), inhibiting inflammatory
actions (9) and promoting
angiogenesis (10). Studies have
indicated that STS protects against brain injury, promotes nerve
regeneration (11) and ameliorates
blood-brain barrier (BBB) damage caused by ischemia (12). STS is able to promote the integrity
(13) and angiogenesis of vascular
endothelial cells (14). However,
it has remained elusive whether STS has an effect on promoting
angiogenesis in cerebral ischemic injury.
After stroke, nerve cells in the ischemic penumbra
are impaired but they are still able to survive for a short
duration, which is a reversible state (15,16).
The survival and functional recovery of brain tissue after ischemic
stroke depends on cerebral ischemic reperfusion and the
establishment of a functional collateral circulation (13). Extensive experimentation has proven
that the angiogenic response has an important role in the recovery
of neurological function in the delayed phases of stroke disease
(17,18). Angiogenesis may improve tissue
microperfusion in ischemic boundary regions and reduce infarct
volumes (19,20). Studies have revealed that
angiogenesis not only provides a sufficient supply of oxygen and
nutrients for the repair of neurons, but also offers a favorable
microenvironment for neuron regeneration after brain damage
(21). These features are
beneficial for long-term functional recovery. Therefore,
enhancement of angiogenesis is one of the strategies for
facilitating functional recovery from acute injury to delayed
repair period (22).
Vascular endothelial growth factor (VEGF) represents
one of the most important modulators in promoting postischemia
neurovascular remodeling (23,24).
Ischemia stimulates VEGF expression in the brain (25), thereby promoting the formation of
new cerebral blood vessels (26).
VEGF may exert mitogenic and antiapoptotic effects on endothelial
cells, which increase vascular permeability and promote cell
migration (27). VEGF exerts its
biological functions through two closely related tyrosine kinase
receptors, VEGF receptor (VEGFR)1 and VEGFR2(28). Previous studies have indicated that
VEGF has a significant role in promoting postischemia neurovascular
recovery by regulating VEGFR2 signaling (23). Angiopoietin-1 (Ang-1) is also an
important factor in promoting angiogenesis. It has been suggested
that Ang-1 may enhance the effects of other angiogenic cytokines
and maintain the integrity of the neovascularization structure
(29).
In the present study, the protective effects of STS
on middle cerebral artery occlusion reperfusion injury and
angiogenesis were investigated in Sprague Dawley (SD) rats and
edaravone was used as a positive control. Furthermore, the
mechanisms by which STS promotes angiogenesis in brain tissue was
preliminarily explored.
Materials and methods
Animals
A total of 92 adult male SD rats (7-8 weeks, 280-320
g) were purchased from Sino-British SIPPR/BK Lab. Animal Co., Ltd..
The rats were individually caged in a climate-controlled room
(20-26˚C, relative humidity of 40-70%) and maintained on a 12-h
light/dark cycle, and they were allowed free access to water and
food. All of the animal experiments were approved by the Ethics
Committee of the animal experiment center of Shanghai University of
Traditional Chinese Medicine (no. PZSHUTCM19010411) and experiments
were performed in accordance with the guidelines of the regulation
for the administration of affairs concerning experimental animals
of China enacted in 1988. All efforts were made to minimize the
number of animals experiencing stress and pain.
The rats were randomly assigned to four groups
containing 20 animals each: i) Sham group (treated with saline);
ii) vehicle group [middle cerebral artery occlusion
(MCAO)/reperfusion model treated with saline]; iii) STS group
(MCAO/reperfusion model treated with STS); and iv) positive control
group (MCAO/reperfusion model treated with edaravone). STS was
purchased from Xi'an Tongze Biotechnology Co., Ltd. and edaravone
was obtained from Zhejiang Shengtong Biotechnology Co., Ltd..
Different concentrations of STS and edaravone (4 animals each dose)
were screened as follows: STS [5, 15 or 30 mg/kg body weight dose
selection was based on previous references (30,31)]
and edaravone (5 or 30 mg/kg body weight dose selection was based
on the maximum and minimum doses of STS) dissolved in saline was
intraperitoneally injected 2 h after reperfusion and then once a
day for 3 days.
MCAO and reperfusion in rats
All SD rats were anesthetized with isoflurane prior
to the experiments (3% of induction dose and 2.5% of maintenance
dose). Focal ischemic infarction was induced by occlusion of the
left MCA as described previously (32). In brief, an incision was performed
in the midline of the rat neck to isolate the common carotid
artery, external carotid artery and internal carotid artery. The
common carotid artery and the external carotid artery were then
ligated with a 4-0 nonabsorbent suture and a monofilament nylon
suture was inserted from the common carotid artery to block the
blood flow of the middle cerebral artery. Subsequently, the skin
incision was closed and the anesthesia was stopped. After ischemia
for 2 h, the nylon suture on the carotid arteries was removed and
the rats were returned to their cages to allow blood reperfusion
for 72 h. During the whole operation, the rats were kept in a 37˚C
environment. After the skin incision was closed, the anesthesia was
stopped and the rats were returned to their cages. The mental
status of rats was closely observed by the experimenter on the
cage-side once a day during the experimental period, and euthanasia
was performed when the rats developed soft paralysis of the limbs
with no spontaneous activity. This only occurred in a few rats, and
the rats without the aforementioned conditions were used for
subsequent experiments and the brain tissues were harvested after
anesthesia at the end of the experiment. The sham operation group
of rats received the same operation except for nylon suture
insertion.
Staining of infarcted brain
tissues
On the third day after the operation, rats were
anesthetized with 2.5% isoflurane and the brain was removed for
2,3,5-triphenyltetrazolium chloride (TTC) staining (four rats for
each group). The brain tissues were sliced into 1.0 mm-thick
coronal sections, incubated in 2% TTC solution (Sigma-Aldrich;
Merck KGaA) for 15 min at 37˚C and then fixed in 4%
paraformaldehyde for 3 min. The red area represented normal brain
tissue and the white area represented infarct tissue. The
infarction area was calculated in a blinded manner with the
PhotoShop CS6 (13.0) analysis system (Adobe®). The
results are expressed as the percentage of infarcted area/total
brain tissue area in the coronal sections.
Neurological testing
Neurobehavioral function assessments were performed
on the third day after MCAO/reperfusion according to a previous
protocol by Longa et al (33). The standard of the assessment was as
follows: 0, no neurological functional impairment; 1, failure to
extend the contralateral forelimb; 2, circling to the contralateral
side; 3, contralateral toppling of walking due to brain injury; and
4, no spontaneous walking and exhibition of a depressed level of
consciousness. The testing was performed in a blinded manner
regarding the group identity and there were four rats in each
group.
BBB permeability
Evans Blue was injected to evaluate BBB permeability
(four rats for each group). In brief, after 3 days of
MCAO/reperfusion, the animals were injected with 2% Evans blue
solution (4 ml/kg body weight) through the tail vein. The
circulation time (2, 4 and 24 h) of Evans blue solution was first
screened and 2 h of circulation was used in the present study
(34,35). Subsequently, the rats were
anesthetized with 3% isoflurane and treated with 150 ml of PBS for
transcardial perfusion. and the brain was removed to separate the
ischemic hemisphere. The brain tissue was then homogenized in 2 ml
DMSO and incubated at 50˚C for 2 h, followed by centrifugation
(10,000 x g) at 4C for 30 min. The supernatant was collected and
the absorbance was recorded at 620 nm using a microplate reader
(1510; Thermo Fisher Scientific, Inc.).
Brain edema
Brain tissues were collected on the third day after
MCAO/reperfusion and the olfactory bulb and the tissue at 2 mm in
front of the frontal pole were discarded. The remaining brain
tissue was cut into slices of 2 mm thickness for brain water
content determination (four rats for each group).
The specific method was as follows: A piece of
aluminum foil was weighed to determine its mass (mA),
the brain tissue was placed on the aluminum foil to measure the
total weight (mB) and the wet weight (WW) was calculated
as (mB-mA). The brain tissue was then wrapped
loosely with aluminum foil and placed in a constant-temperature
oven at 110˚C for 12 h (36). After
the tested brain tissue was returned to room temperature, its
weight (mC) was determined to calculate the dry weight
(DW). The formula for calculating the water content of brain tissue
was as follows: (WW-DW)/WW x100%.
Histological analysis
After 3 days of MCAO/reperfusion, the rats were
placed in the supine position after anesthesia with 3% isoflurane
and perfused with 60 ml of 4% paraformaldehyde for 5 min after
perfusion 150 ml of PBS (pH 7.4) for 10 min and then the brain was
rapidly removed. The intact brain tissue was fixed in 4%
paraformaldehyde at room temperature for 48 h, embedded in paraffin
and cut into 5-µm coronal sections. The sections were stained with
H&E (three rats for each group). The histological features of
the cerebral cortex and the striatum were observed under a light
microscope (Olympus Corporation).
Immunofluorescence staining
Paraffin brain sections were prepared for
immunofluorescence staining. The immunostaining procedure was
performed as previously described (37). Briefly, deparaffinize and rehydrate:
incubate sections in 3 changes of xylene, 10 min each. Dehydrate in
2 changes of pure ethanol for 5 min, followed by dehydrate in
gradient ethanol of 85 and 75% ethanol, respectively, 5 min each.
Wash in distilled water. Antigen retrieval: immerse the slides in
EDTA antigen retrieval buffer (Wuhan Servicebio Technology Co.,
Ltd.) and maintain at a boiling water for 35 min. Let air cooling.
Wash three times with PBS (pH 7.4) in a Rocker device, 5 min each.
and blocked with 10% goat serum (Beyotime Biotechnology Co., Ltd.)
at room temperature for 15 min. Sections were first incubated with
antibodies to CD31 (1:150 dilution; cat. no. GB12063; Wuhan
Servicebio Technology Co., Ltd.) and α-smooth muscle actin (SMA;
1:150 dilution; cat. no. GB111364; Wuhan Servicebio Technology Co.,
Ltd.) as indicators of vascular density (three rats for each
group), VEGF (1:150 dilution; sc-7269; Santa) as an indicator of
angiogenesis and neuronal nuclear protein (NeuN; 1:150 dilution;
ab177417; Abcam) to visualize mature neurons, overnight at 4˚C.
Subsequently, the sections were incubated with a suitable secondary
antibody (anti-mouse IgG Alexa Flour® 488 Conjugate;
cat. no. 4408; CST; 1:200 dilution and Cy™3-conjugated
Affinipure Donkey anti-rabbit IgG; 152679; Jackson Immuno Research;
1:350 dilution). Cell nuclei were stained with DAPI. The paraffin
sections were imaged with a Nikon fluorescence microscope (Olympus
Corporation) and images were quantified with ImageJ software1.50i
(NIH). All evaluations were performed by an investigator blinded to
the experiment (five rats for each group).
TUNEL staining
TUNEL staining of paraffin sections of brain tissues
was used to detect apoptotic cell death based on DNA fragmentation.
TUNEL assays were performed according to the manufacturer's
protocol (Roche Diagnostics). Slides were then counterstained with
DAPI. The paraffin sections were imaged using a Nikon fluorescence
microscope (Olympus Corporation) and images were quantitatively
evaluated with ImageJ software1.50i [National Institutes of Health
(NIH)]. Counting was performed in a blinded manner and there were
five rats for each group.
Immunohistochemical staining
Paraffinized brain sections were dewaxed and
dehydrated in xylene and ethanol solution, followed by antigen
retrieval and blocking with 10% goat serum (Beyotime Biotechnology
Co., Ltd.) at room temperature for 15 min. All sections were then
incubated with anti-VEGFR2 antibody (1:800 dilution; cat. no.
9698S; Cell Signaling Technology, Inc.) at 4˚C overnight.
Subsequently, the sections were washed in PBS three times and
incubated with Horseradish peroxidase (HRP)-conjugated goat
anti-rabbit IgG secondary antibodies (1:200 dilution; GB23303;
Wuhan Servicebio Technology Co., Ltd.) for 10 min at room
temperature. Next, the sections were visualized using a
diaminobenzidine kit (Wuhan Servicebio Technology Co., Ltd.) and
counterstained with hematoxylin. With a light microscope, an
investigator blinded to the experimental groups randomly chose
three separate tissue sections for each rat (four rats for each
group). The staining was analyzed using ImageJ analysis
software1.50i (NIH).
Reverse transcription-quantitative
(RT-q)PCR
Brain tissues from the penumbra were separated and
total RNA was extracted using TRIzol® reagent (CWBIO).
Subsequently, the RNA was reverse transcribed to cDNA with a
PrimeScript RT Reagent Kit (Takara Bio, Inc.) according to the
manufacturer's instructions. cDNA was amplified with
SYBR®-Green Real-time PCR Master Mix (Takara Bio, Inc.)
according to the manufacturer's instructions by RT-qPCR machine
(QuantStudio 6 Flex, Thermo Fisher Scientific, Inc.). The following
gene-specific primers: VEGF-A [forward (F),
5'-CGACAGAAGGGGAGCAGAAAG-3' and reverse (R),
5'-GCACTCCAGGGCTTCATCATT-3'], Ang-1 (F, 5'-GTCACTGCACAAAAGGGACA-3'
and R, 5'-GGCTTACAAGGATGGCGTTA-3'), VEGFR-2 (F,
5'-TAGCACGACAGAGACTGTGAGG-3' and R, 5'-TGAGGTGAGAGAGATGGGTAGG-3'),
CD31 (F, 5'-TTTCGCTGCCAAGCTGGCGT-3' and R,
5'-CCACCTGCACGCTGCACTTGAT-3'), β-actin (F,
5'-GGGACCTGACTGACTACCTC-3' and R, 5'-TCATACTCCTGCTTGCTGAT-3') and
basic fibroblast growth factor (bFGF) (F,
5'-GAGCGACCCACACGTCAAACTAC-3' and R,
5'-CAGCCGTCCATCTTCCTTCATAGC-3'). The following thermocycling
conditions were used for qPCR: Initial denaturation at 95˚C for 30
sec, denaturation (40 cycles) at 95˚C for 5 sec,
annealing/extending (40 cycles) at 60˚C for 30 sec. Melt curve at
95˚C for 15 sec, at 60˚C for 1 min and at 95˚C for 15 sec. β-actin
was used as a reference in the experiment. Relative mRNA expression
was normalized to β-actin levels and analyzed with the
2-ΔΔCq method (38).
Three rats were used for each group.
Statistical analysis
The neurological score data were presented as the
median and interquartile range and data were analyzed with the
Kruskal-Wallis test followed by Dunn's post-hoc test. All other
data were expressed as the mean ± standard deviation and
differences between groups were compared by one-way ANOVA followed
by Bonferroni's multiple-comparisons test. GraphPad 7.0 (GraphPad
Software, Inc.) was used for all statistical analyses.
*P<0.05, **P<0.01,
***P<0.001; #P<0.05,
##P<0.01, ###P<0.001;
&&P<0.01,
&&&P<0.001 indicates a significant
difference. * Indicates compared with the vehicle group, #
indicates compared with the control group. & indicates the STS
group compared with the control group.
Results
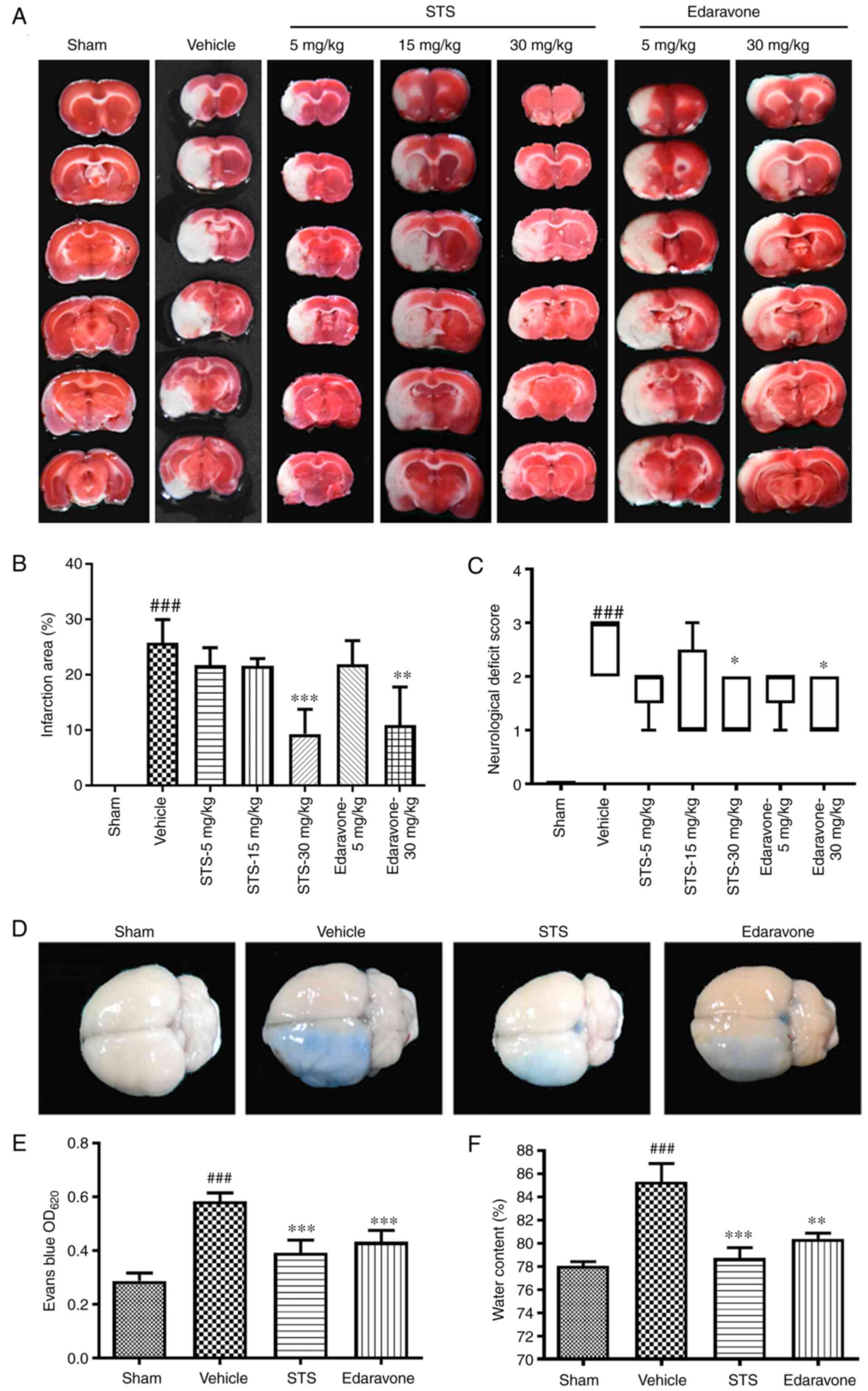
STS decreases infarct size and
neurological deficit scores
Cerebral ischemia injury was evaluated by
neurological function scores and measurement of infarct size in
rats. After 3 days of MCAO/reperfusion, the infarct area was
assessed by TTC staining. The infarct area significantly increased
in the vehicle group compared with the sham group. Compared with
that in the vehicle group, the infarct area in the STS treatment
group (30 mg/kg) and the edaravone group (30 mg/kg) was
significantly reduced (Fig. 1A and
B). The assessment of neurological
deficits was determined. The rats in the sham-operated group had no
neurological deficit. Conversely, the neurological deficit scores
of the vehicle rats significantly increased. STS significantly
ameliorated neurological function deficits at the dose of 30 mg/kg
at 3 days after surgery, as did edaravone (30 mg/kg) (Fig. 1C). The above results indicated that
the same dose of STS as that of edaravone (30 mg/kg) reduced
cerebral infarction and improved neurological function. Thus, in
subsequent experiments, STS and edaravone were used for further
study at a dose of 30 mg/kg.
STS preserves the integrity of the BBB
and reduces brain edema
To investigate whether STS has a protective effect
on the BBB, the extent of BBB disruption was detected by leakage of
Evans blue in the ipsilateral hemispheres after cerebral ischemia.
After 3 days of MCAO/reperfusion, in the vehicle group, the BBB
integrity was significantly decreased compared with the sham group.
The BBB integrity in the STS (30 mg/kg) and edaravone (30 mg/kg)
groups was significantly improved compared with that in the vehicle
group (Fig. 1D and E). The therapeutic effect of STS on the
MCAO/reperfusion model was also determined by the content of brain
edema. The cerebral edema content of the vehicle group was
significantly increased after injury compared with that in the sham
group, while that in the STS (30 mg/kg) and edaravone (30 mg/kg)
groups was significantly reduced compared with that in the vehicle
group (Fig. 1F). These results
indicated that STS attenuated pathological changes caused by
cerebral ischemia.
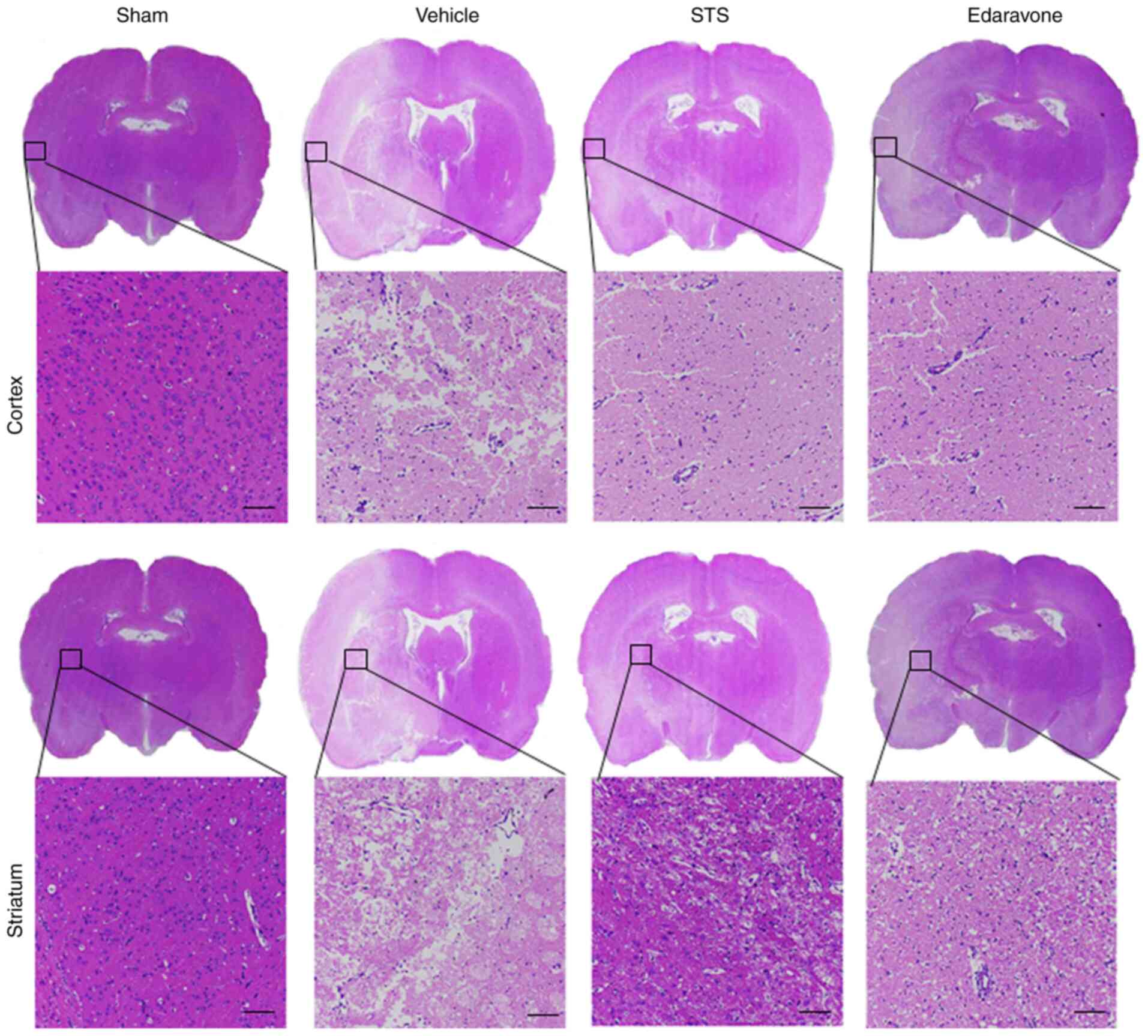
Histopathological changes associated
with STS treatment
To further verify the protective effects of STS on
the MCAO/reperfusion model, morphological changes were observed by
H&E staining after 3 days of MCAO/reperfusion. The brain tissue
was observed under a light microscope (Fig. 2). In the sham group, the structure
of the brain tissue was clear and intact with no obvious
pathological changes. The neurons in the cortex and striatum were
clear and arranged in order. No cell structure damage was observed
and no cytoplasmic vacuoles were detected. By contrast, the brain
pathological structure was significantly changed in the vehicle
group. The specific histopathological observations were as follows:
The structure of the brain tissue was loose and numerous vacuoles
in the cortex and striatum were present. Compared with that in the
sham group, the cell morphology was irregular and the cell number
was markedly decreased. In the STS (30 mg/kg) and edaravone (30
mg/kg) groups, the pathological changes in the cortex and striatum
were significantly alleviated. The loosening of the tissue
structure, shrinkage and cavitation of cells were obviously
reversed in the cortex and striatum and the number of remaining
cells was markedly increased compared with the vehicle group. Thus,
STS had an effect to improve the pathology of cerebral
ischemia/reperfusion injury similar to that of edaravone.
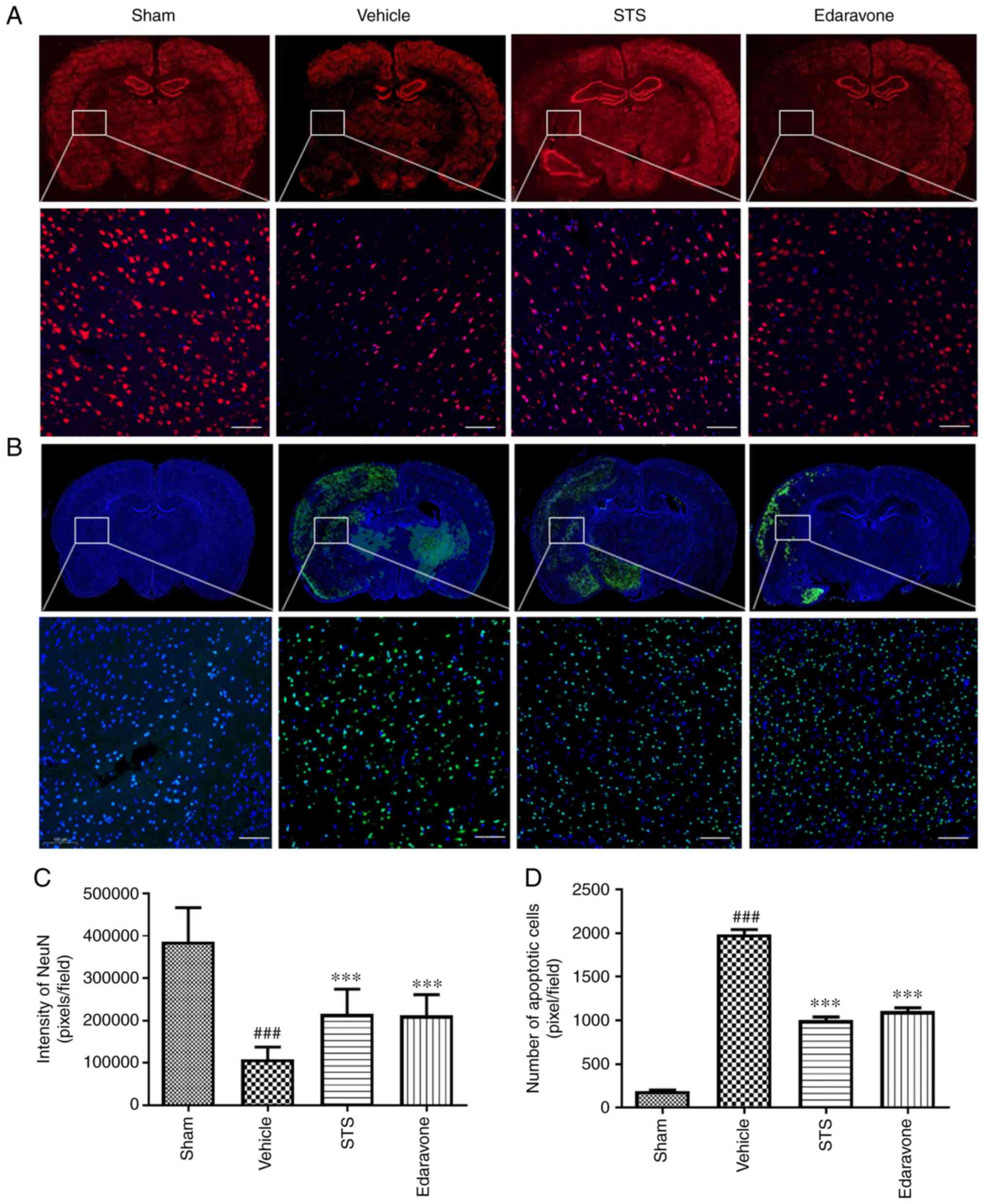
STS inhibits apoptotic cell death
To investigate the neuroprotective effects of STS
against cerebral ischemia/reperfusion injury through inhibiting
apoptotic cell death, NeuN protein, representing neurons in brain
tissue, was labeled with fluorescent antibodies (Fig. 3A) and TUNEL staining was performed
to detect apoptotic cell death in the penumbra area (Fig. 3B). As presented in Fig. 3A and C, numerous neurons were lost in the
vehicle-treated brain sections compared with the sham group, while
the survival rate of the cells in the STS (30 mg/kg) and edaravone
(30 mg/kg) groups was significantly higher than that in the vehicle
group. As presented in Fig. 3B and
D, there was significant apoptosis
in the vehicle compared with the sham group. The number of
apoptotic cells in the STS (30 mg/kg) and edaravone (30 mg/kg)
groups was significantly reduced compared with that in the vehicle
group.
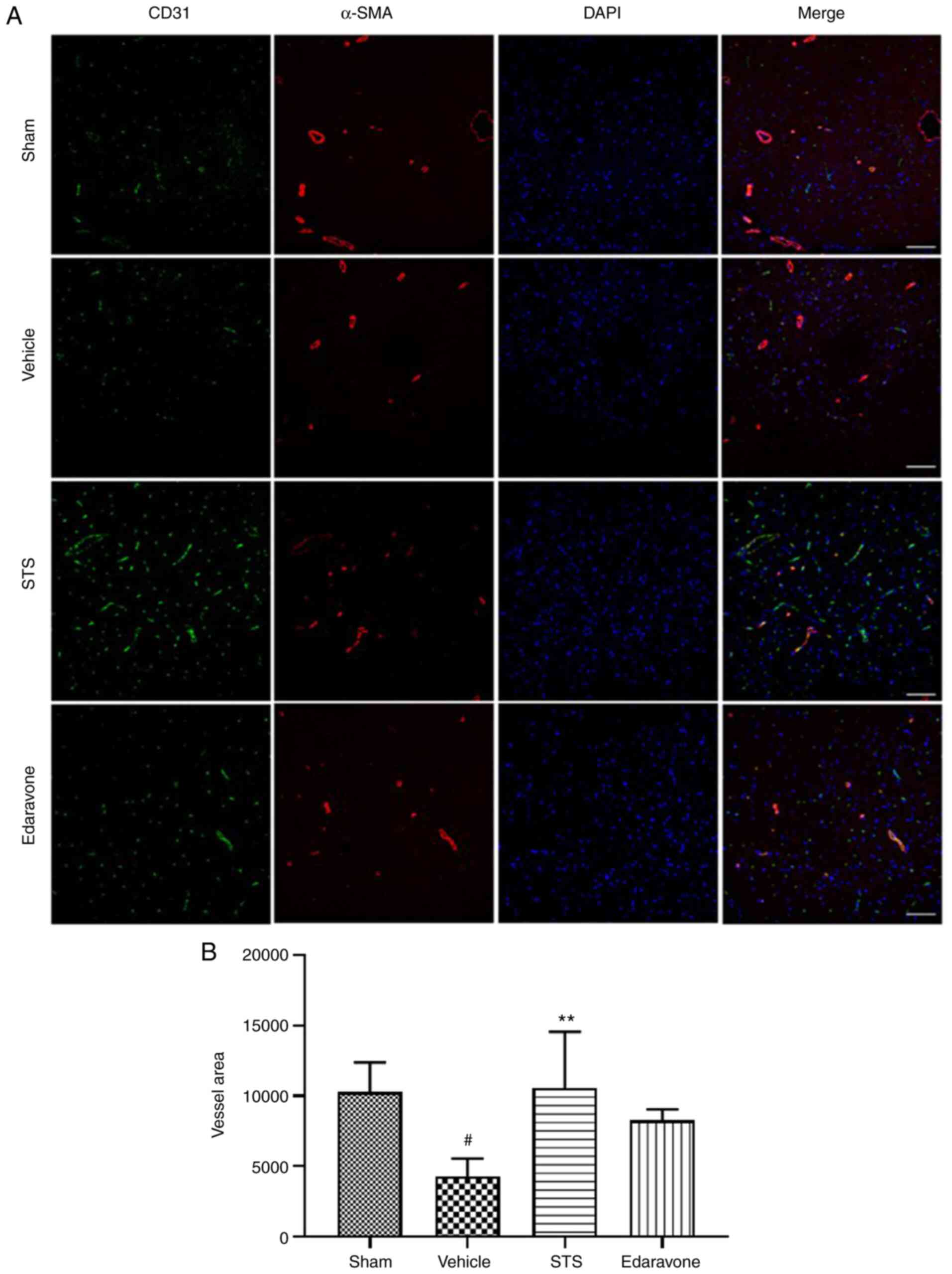
STS enhances vascular density in the
penumbra
Angiogenesis has an important role in ischemic
stroke-induced brain injury repair and long-term functional
recovery (39,40). Thus, in the present study, to
evaluate whether STS promotes vascular density in the penumbra
after 3 days of ischemia/reperfusion, the expression of the
endothelial cell marker CD31 and vascular smooth muscle cell marker
α-SMA was examined via immunofluorescence (Fig. 4). The vascular density was
significantly reduced in the vehicle group compared with that in
the sham operation group, while in the STS (30 mg/kg) group, the
vascular density was significantly increased compared with that in
the vehicle group. However, edaravone treatment (30 mg/kg) resulted
in no significant change in vascular density. The above results
indicated that STS was able to promote angiogenesis in the penumbra
after cerebral ischemia.
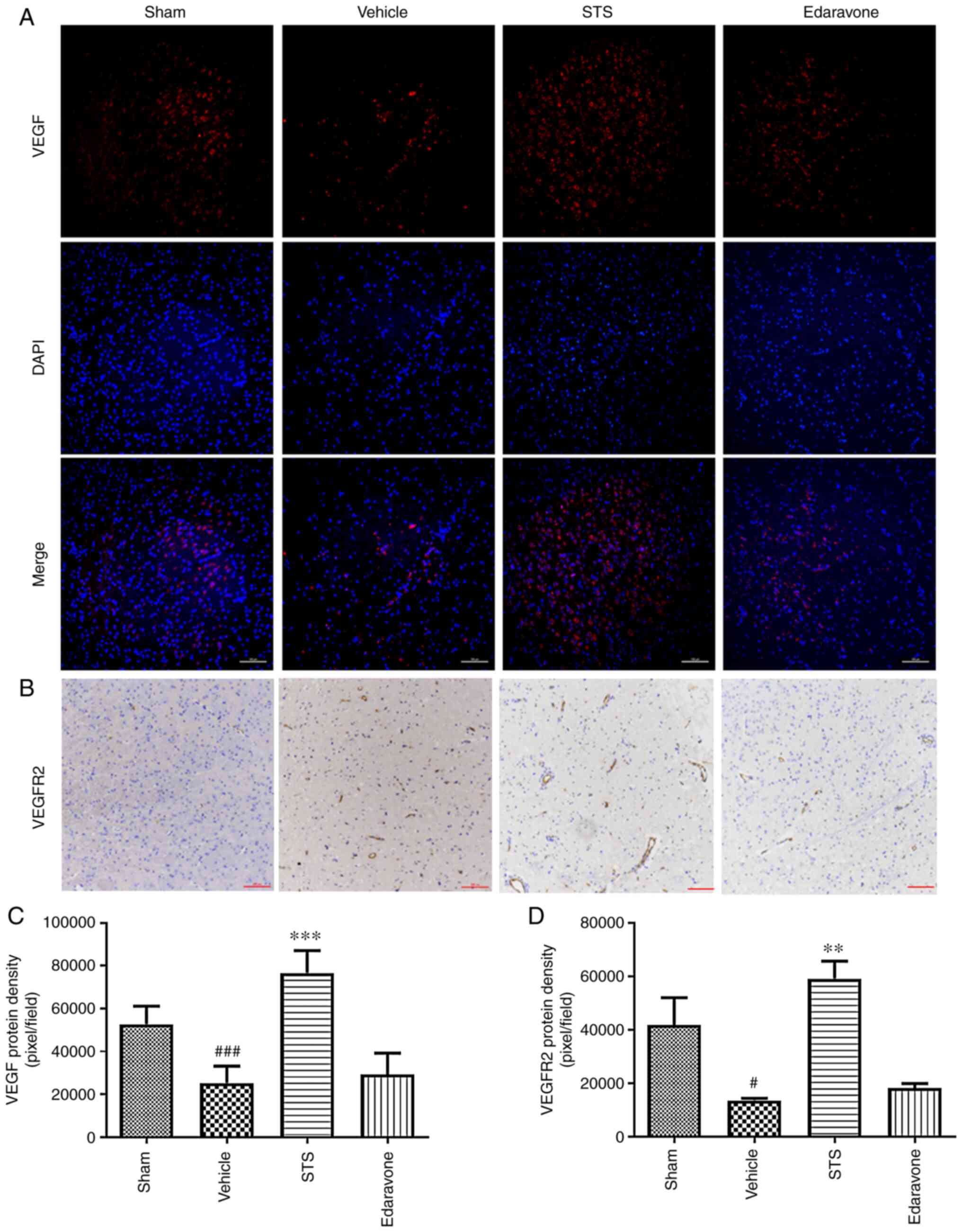
STS increases the protein expression
of VEGF and VEGFR2
VEGF/VEGFR2 signaling has the greatest effect to
promote neurovascular regeneration after ischemic stroke (23). In the ischemia/reperfusion model of
the present study, as presented in Fig.
5, the amount of VEGF protein in the vehicle group detected by
immunofluorescence was significantly reduced compared with that in
the sham group, while the expression of VEGF protein in the STS (30
mg/kg) group was significantly increased compared with that in the
vehicle group after 3 days of ischemia/reperfusion (Fig. 5A and C). The same trend and effect of STS (30
mg/kg) on VEGFR2 protein levels were obtained using
immunohistochemistry (Fig. 5B and
D). However, no obvious changes in
VEGF/VEGFR2 protein were observed after edaravone (30 mg/kg)
treatment (Fig. 5).
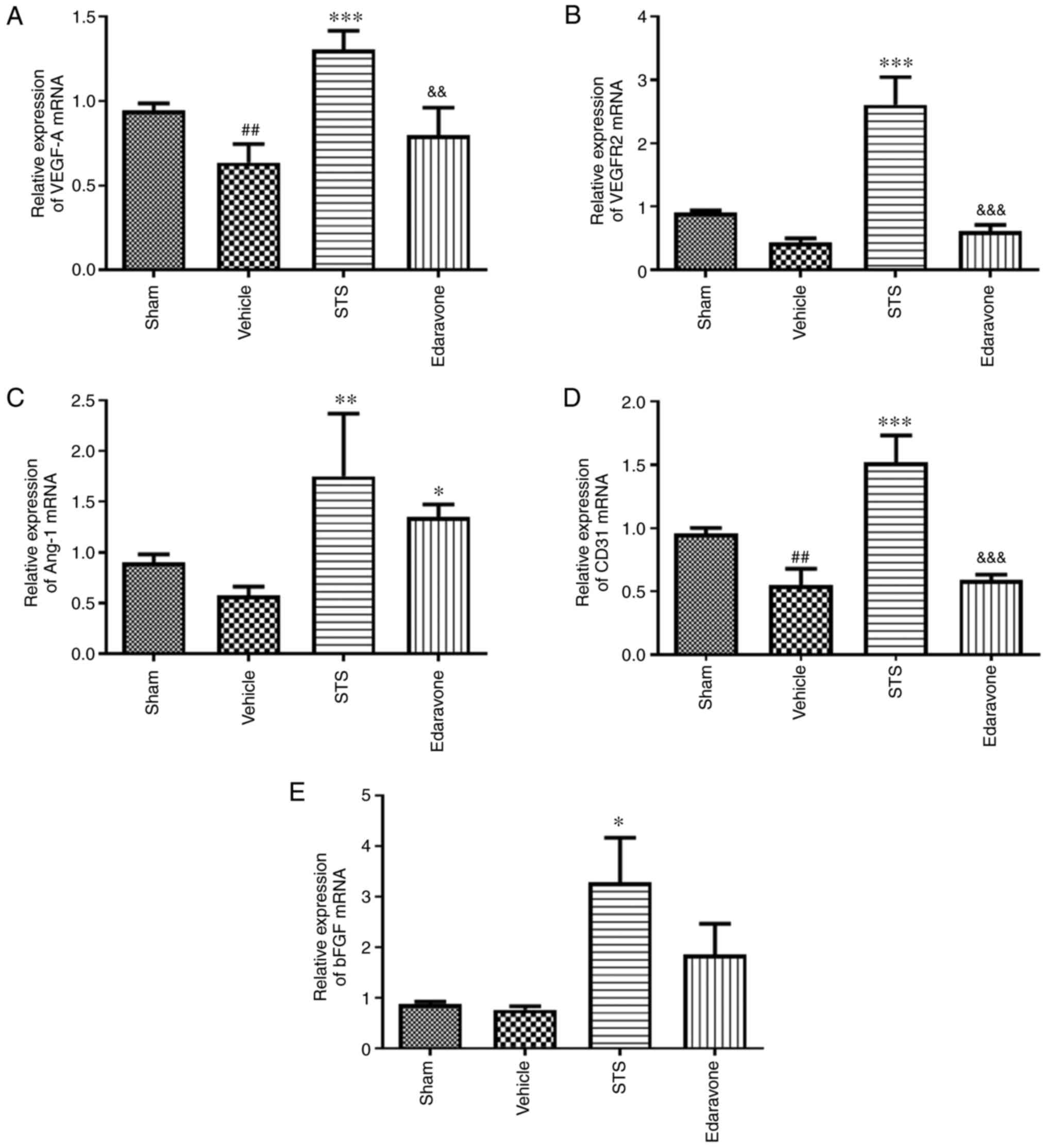
STS promotes mRNA expression of
angiogenesis-related factors
To determine the effect of STS on regulatory factors
of angiogenesis, the relative mRNA levels of the factors VEGF-A,
Ang-1, VEGFR-2, CD31 and bFGF in brain tissue were determined using
RT-qPCR. In the vehicle group, the mRNA expression of VEGF-A and
CD31 was significantly reduced, the mRNA expression of VEGFR2,
Ang-1 and bFGF was decreased, but not significantly compared with
that in the sham group. Treatment with STS (30 mg/kg) significantly
increased the mRNA expression of VEGF-A (Fig. 6A), VEGFR-2 (Fig. 6B), Ang-1 (Fig. 6C), CD31 (Fig. 6D) and bFGF (Fig. 6E) compared with that in the vehicle
group. There was no significant difference between the edaravone
group and vehicle group except for Ang-1 mRNA.
Discussion
Danshen separated from the dried root of Salvia
miltiorrhiza, a traditional Chinese medicinal herb, is widely
used for the prevention and treatment of various cardiovascular
diseases in the clinic, including angina pectoris, hyperlipidemia
and acute ischemic stroke (41,42).
Tanshinone IIA is an important monomer extracted from Danshen. To
improve the poor water solubility of tanshinone IIA, STS was
obtained by sulfonation to change the structure and increase the
bioavailability. The present study focused on the protective
effects of STS on brain damage induced by acute cerebral
ischemia/reperfusion in rats and demonstrated that STS relieved
cerebral injury after ischemia/reperfusion by promoting
angiogenesis.
A clinical study revealed that STS improved
neurologic functional outcomes for patients with acute ischemic
stroke by reducing BBB leakage and damage (12). Injury to the BBB allows blood-borne
cells, chemicals and fluids to flow into the brain, which may lead
to complications, such as inflammatory responses and cerebral edema
(43). In terms of clinical
symptoms, BBB destruction has been indicated to be closely related
to cerebral edema (44). In the
present model of cerebral ischemia/reperfusion, it was indicated
that STS was effective in reducing brain damage in rats with
cerebral ischemia/reperfusion, including attenuation of cerebral
infarction, protection of the integrity of the BBB and reduced
brain edema. More importantly, STS also decreased the neurological
deficit scores, reversed histopathological changes and reduced
apoptotic cell death. The magnitude of these protective effects of
STS on cerebral ischemia/reperfusion injury in rats was similar to
that of edaravone, a commonly used neuroprotective agent in the
clinic. In the present study, 5, 15 and 30 mg/kg doses of STS were
used for some preliminary screening, and 30 mg/kg of STS was the
most effective therapeutic dose. However, the best dose for
treatment remains to be determined.
To further elucidate the protective mechanism of STS
on cerebral ischemic/reperfusion injury in rats, the angiogenic
effect of STS was further studied. Angiogenesis has an important
role in the treatment of ischemic stroke (45). Pharmacological targeting of
microcirculatory dysregulations has been indicated to be improved
by angiogenesis (46) and STS has a
protective effect on microcirculatory disturbance (47). After a stroke, disruption of the BBB
occurs acutely whereas regeneration of cerebral microvessels
develops relatively late in ischemic brain (40), the body's own recovery is relatively
slow, among which the acceleration of angiogenesis may be expected
to enhance the prognosis (39), and
STS accelerates neovascularization of the ischemic zone. In the
present study, it was first verified that STS was able to increase
the vascular density in the ischemic brain area. The effect of STS
on the VEGF/VEGFR2 signaling pathway, which has a major role in
angiogenesis, was also explored. VEGF is a growth factor that
stimulates the proliferation of vascular endothelial cells and has
vital roles in neovascular remodeling in ischemic stroke (43). In addition, VEGF possesses
neurotrophic and neuroprotective activity and the direct
neurotrophic effect of VEGF was able to confer neuroprotective
activity independent of angiogenesis (48). However, whether STS promotes VEGF in
cerebral ischemia has not been reported in the literature, to the
best of our knowledge. The present study confirmed that STS
promoted the protein expression of VEGF and VEGFR2 in ischemic
brain tissue of rats, which may indicate that STS regulates
angiogenesis by activating the VEGF/VEGFR2 signaling pathway. The
protein expression of VEGF will be further detected with ELISAs in
the serum and supernatant of brain tissue homogenate in future
studies. In addition, the cell types expressing VEGF after STS
treatment have remained elusive. Certain studies have raised that
macrophages secrete various angiogenic growth factors (including
VEGF) and neurotrophic factors to promote angiogenesis, preserve
the cortical blood supply and improve neurological function in the
acute phase of cerebral ischemia/reperfusion (49,50).
Next, the relationship between VEGF and macrophages will be further
investigated. In the present study, the mRNA expression levels of
angiogenesis-related factors in the rat brain after cerebral
ischemia were examined. The results indicated that the mRNA
expression levels of VEGF-A, Ang-1, VEGFR-2, CD31 and bFGF in the
rat brain after cerebral ischemia were reduced compared with the
Sham group. However, STS could reverse this phenomenon and promote
the expression of these angiogenesis-related factors compared with
the vehicle group. Edaravone had no significant effect on any of
these angiogenesis-related factors/processes apart from increasing
Ang-1 expression. Therefore, STS was proven to be particularly
effective in promoting angiogenesis following ischemic stroke.
In conclusion, the present study demonstrated that
STS promoted angiogenesis by regulating the protein expression of
VEGF and VEGFR2, and regulating the mRNA levels of
angiogenesis-related factors, which may contribute to the
angiogenesis-promoting effect of STS after cerebral ischemia,
leading to reduction of the cerebral infarction area, relief of
brain edema and improvement of BBB integrity, thereby promoting the
recovery of neurobehavior after experimental stroke.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81573579
and 82003800), the New Interdisciplinary Subject Funding Program
for Shanghai Traditional Chinese Medicine (grant no. E2-F18003) and
the Science and Technology Commission of Shanghai Municipality
(grant nos. 18401933500 and 20ZR1473200).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
JZ and YZ designed the study. JX, PZ and YC
performed the experiments. JX and YC analyzed the data. JX and YZ
drafted the manuscript and PZ and JZ revised the manuscript. YX and
PL helped with the data analysis. YZ and JX confirmed the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Ethics
Committee of the animal experiment center of Shanghai University of
Traditional Chinese Medicine (Shanghai, China; approval no.
PZSHUTCM19010411).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Benjamin EJ, Blaha MJ, Chiuve SE, Cushman
M, Das SR, Deo R, De Ferranti SD, Floyd JS, Fornage M, Gillespie C,
et al: Heart disease and stroke statistics-2017 update: A report
from the American heart association. Circulation. 135:e146–e603.
2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Benjamin EJ, Virani SS, Callaway CW,
Chamberlain AM, Chang AR, Cheng S, Chiuve SE, Cushman M, Delling
FN, Deo R, et al: Heart disease and stroke statistics-2018 update:
A report from the American heart association. Circulation.
137:e67–e492. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Winstein CJ, Stein J, Arena R, Bates BE,
Cherney LR, Cramer SC, Deruyter F, Eng JJ, Fisher BE, Harvey RL, et
al: Guidelines for adult stroke rehabilitation and recovery: A
guideline for healthcare professionals from the American heart
association/American stroke association. Stroke. 47:e98–e169.
2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Shang Q, Xu H and Huang L: Tanshinone IIA:
A promising natural cardioprotective agent. Evid Based Complement
Alternat Med. 2012(716459)2012.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Shang QH, Wang H, Li SM and Xu H: The
effect of sodium tanshinone IIA sulfate and simvastatin on elevated
serum levels of inflammatory markers in patients with coronary
heart disease: A study protocol for a randomized controlled trial.
Evid Based Complement Alternat Med. 2013(756519)2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Tang J, Zhu C, Li ZH, Liu XY, Sun SK,
Zhang T, Luo ZJ, Zhang H and Li WY: Inhibition of the spinal
astrocytic JNK/MCP-1 pathway activation correlates with the
analgesic effects of tanshinone IIA sulfonate in neuropathic pain.
J Neuroinflamm. 12(57)2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zhang H, Long M, Wu Z, Han X and Yu Y:
Sodium tanshinone IIA silate as an add-on therapy in patients with
unstable angina pectoris. J Thorac Dis. 6:1794–1799.
2014.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zhao YP, Wang F, Jiang W, Liu J, Liu BL,
Qi L and Zhou W: A mitochondrion-targeting tanshinone IIA
derivative attenuates myocardial hypoxia reoxygenation injury
through a SDH-dependent antioxidant mechanism. J Drug Target.
27:896–902. 2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Hu Q, Wei B, Wei L, Hua K, Yu X, Li H and
Ji H: Sodium tanshinone IIA sulfonate ameliorates ischemia-induced
myocardial inflammation and lipid accumulation in beagle dogs
through NLRP3 inflammasome. Int J Cardiol. 196:183–192.
2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Xu W, Yang J and Wu LM: Cardioprotective
effects of tanshinone IIA on myocardial ischemia injury in rats.
Pharmazie. 64:332–336. 2009.PubMed/NCBI
|
|
11
|
Morton JS, Andersson IJ, Cheung P, Baker
PN and Davidge ST: The vascular effects of sodium tanshinone IIA
sulphonate in rodent and human pregnancy. PLoS One.
10(e0121897)2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ji B, Zhou F, Han L, Yang J, Fan H, Li S,
Li J, Zhang X, Wang X and Chen X: Sodium tanshinone IIA sulfonate
enhances effectiveness Rt-PA treatment in acute ischemic stroke
patients associated with ameliorating blood-brain barrier damage.
Transl Stroke Res. 8:334–340. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhou ZY, Huang B, Li S, Huang XH, Tang JY,
Kwan YW, Hoi PM and Lee SMY: Sodium tanshinone IIA sulfonate
promotes endothelial integrity via regulating VE-cadherin dynamics
and RhoA/ROCK-mediated cellular contractility and prevents
atorvastatin-induced intracerebral hemorrhage in zebrafish. Toxicol
Appl Pharmacol. 350:32–42. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Li Z, Zhang S, Cao L, Li W, Ye YC, Shi ZX,
Wang ZR, Sun LX, Wang JW, Jia LT and Wang W: Tanshinone IIA and
Astragaloside IV promote the angiogenesis of mesenchymal stem
cell-derived endothelial cell-like cells via upregulation of Cx37,
Cx40 and Cx43. Exp Ther Med. 16:1847–1854. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Garcia JH, Wagner S, Liu KF and Hu XJ:
Neurological deficit and extent of neuronal necrosis attributable
to middle cerebral artery occlusion in rats. Statistical
validation. Stroke. 26:627–635. 1995.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Lo EH: A new penumbra: Transitioning from
injury into repair after stroke. Nat Med. 14:497–500.
2008.PubMed/NCBI View
Article : Google Scholar
|
|
17
|
Ma F, Morancho A, Montaner J and Rosell A:
Endothelial progenitor cells and revascularization following
stroke. Brain Res. 1623:150–159. 2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Jiang Y, Li L, Ma J, Zhang L, Niu F, Feng
T and Li C: Auricular vagus nerve stimulation promotes functional
recovery and enhances the post-ischemic angiogenic response in an
ischemia/reperfusion rat model. Neurochem Int. 97:73–82.
2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Han L and Li J, Chen Y, Zhang M, Qian L,
Chen Y, Wu Z, Xu Y and Li J: Human urinary kallidinogenase promotes
angiogenesis and cerebral perfusion in experimental stroke. PLoS
One. 10(e0134543)2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Hui Z, Sha DJ, Wang SL, Li CS, Qian J,
Wang JQ, Zhao Y, Zhang JH, Cheng HY, Yang H, et al: Panaxatriol
saponins promotes angiogenesis and enhances cerebral perfusion
after ischemic stroke in rats. BMC Complement Altern Med.
17(70)2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Petcu EB, Smith RA, Miroiu RI and Opris
MM: Angiogenesis in old-aged subjects after ischemic stroke: A
cautionary note for investigators. J Angiogenes Res.
2(26)2010.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Arai K, Jin G, Navaratna D and Lo EH:
Brain angiogenesis in developmental and pathological processes:
Neurovascular injury and angiogenic recovery after stroke. FEBS J.
276:4644–4652. 2009.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Li W, Fraser JL, Yu SP, Zhu J, Jiang Y and
Wei L: The role of VEGF/VEGFR2 signaling in peripheral
stimulation-induced cerebral neurovascular regeneration after
ischemic stroke in mice. Exp Brain Res. 214:503–513.
2011.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lee DH, Lee J, Jeon J, Kim KJ, Yun JH,
Jeong HS, Lee EH, Koh YJ and Cho CH: Oleanolic acids inhibit
vascular endothelial growth factor receptor 2 signaling in
endothelial cells: Implication for anti-angiogenic therapy. Mol
Cells. 41:771–780. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Wise GE and Yao S: Expression of vascular
endothelial growth factor in the dental follicle. Crit Rev Eukaryot
Gene Expr. 13:173–180. 2003.PubMed/NCBI
|
|
26
|
Krum JM, Mani N and Rosenstein JM:
Angiogenic and astroglial responses to vascular endothelial growth
factor administration in adult rat brain. Neuroscience.
110:589–604. 2002.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Melincovici CS, Boşca AB, Şuşman S,
Mărginean M, Mihu C, Istrate M, Moldovan IM, Roman AL and Mihu CM:
Vascular endothelial growth factor (VEGF)-key factor in normal and
pathological angiogenesis. Rom J Morphol Embryol. 59:455–467.
2018.PubMed/NCBI
|
|
28
|
Wang Y, Jin K, Mao XO, Xie L, Banwait S,
Marti HH and Greenberg DA: VEGF-overexpressing transgenic mice show
enhanced post-ischemic neurogenesis and neuromigration. J Neurosci
Res. 85:740–747. 2007.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Asahara T, Chen D, Takahashi T, Fujikawa
K, Kearney M, Magner M, Yancopoulos GD and Isner JM: Tie2 receptor
ligands, angiopoietin-1 and angiopoietin-2, modulate VEGF-induced
postnatal neovascularization. Circ Res. 83:233–240. 1998.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Wang L, Xiong X, Zhang X, Ye Y, Jian Z,
Gao W and Gu L: Sodium tanshinone IIA sulfonate protects against
cerebral ischemia-reperfusion injury by inhibiting autophagy and
inflammation. Neuroscience. 441:46–57. 2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Chen L, He W, Peng B, Yuan M, Wang N, Wang
J, Lu W and Wang T: Sodium tanshinone IIA sulfonate improves
post-ischemic angiogenesis in hyperglycemia. Biochem Biophys Res
Commun. 520:580–585. 2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zhou Z, Wei X, Xiang J, Gao J, Wang L, You
J, Cai Y and Cai D: Protection of erythropoietin against ischemic
neurovascular unit injuries through the effects of connexin43.
Biochem Biophys Res Commun. 458:656–662. 2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Longa EZ, Weinstein P, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Wang J, Zhang D, Fu X, Yu L, Lu Z, Gao Y,
Liu X, Man J, Li S, Li N, et al: Carbon monoxide-releasing
molecule-3 protects against ischemic stroke by suppressing
neuroinflammation and alleviating blood-brain barrier disruption. J
Neuroinflammation. 15(188)2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Li D, Lang W, Zhou C, Wu C, Zhang F, Liu
Q, Yang S and Hao J: Upregulation of microglial ZEB1 ameliorates
brain damage after acute ischemic stroke. Cell Rep. 22:3574–3586.
2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Gilmer LK, Roberts KN and Scheff SW:
Efficacy of progesterone following a moderate unilateral cortical
contusion injury. J Neurotrauma. 25:593–602. 2008.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Luan P, Xu J, Ding X, Cui Q, Jiang L, Xu
Y, Zhu Y, Li R, Lin G, Tian P and Zhang J: Neuroprotective effect
of salvianolate on cerebral ischaemia-reperfusion injury in rats by
inhibiting the caspase-3 signal pathway. Eur J Pharmacol.
872(172944)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Zhu Y, Yang L, Xu J, Yang X, Luan P, Cui
Q, Zhang P, Wang F, Li R, Ding X, et al: Discovery of the
anti-angiogenesis effect of eltrombopag in breast cancer through
targeting of HuR protein. Acta Pharm Sin B. 10:1414–1425.
2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Taguchi A, Soma T, Tanaka H, Kanda T,
Nishimura H, Yoshikawa H, Tsukamoto Y, Iso H, Fujimori Y, Stern DM,
et al: Administration of CD34+ cells after stroke
enhances neurogenesis via angiogenesis in a mouse model. J Clin
Invest. 114:330–338. 2004.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Zhang ZG, Zhang L, Jiang Q, Zhang RL,
Davies K, Powers C, Bruggen NV and Chopp M: VEGF enhances
angiogenesis and promotes blood-brain barrier leakage in the
ischemic brain. J Clin Invest. 106:829–838. 2000.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Zhou L, Zuo Z and Chow MS: Danshen: An
overview of its chemistry, pharmacology, pharmacokinetics, and
clinical use. J Clin Pharmacol. 45:1345–1359. 2005.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Chan P, Liu IM, Li YX, Yu WJ and Cheng JT:
Antihypertension induced by tanshinone IIA isolated from the roots
of Salvia miltiorrhiza. Evid Based Complement Alternat Med.
2011(392627)2011.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Gong P, Zhang Z, Zou C, Tian Q, Chen X,
Hong M, Liu X, Chen Q, Xu Z, Li M and Wang J: Hippo/YAP signaling
pathway mitigates blood-brain barrier disruption after cerebral
ischemia/reperfusion injury. Behav Brain Res. 356:8–17.
2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Giraud M, Cho TH, Nighoghossian N,
Maucort-Boulch D, Deiana G, Østergaard L, Baron JC, Fiehler J,
Pedraza S, Derex L and Berthezène Y: Early blood brain barrier
changes in acute ischemic stroke: A sequential MRI study. J
Neuroimaging. 25:959–963. 2015.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Navaratna D, Guo S, Arai K and Lo EH:
Mechanisms and targets for angiogenic therapy after stroke. Cell
Adh Migr. 3:216–223. 2009.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Ampofo E, Schmitt BM, Menger MD and
Laschke MW: Targeting the microcirculation by indole-3-carbinol and
its main derivate 3,3,'-diindolylmethane: Effects on angiogenesis,
thrombosis and inflammation. Mini Rev Med Chem. 18:962–968.
2018.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Zhu W, Lv Q, Chen H, Wang Z and Zhong Q:
Protective effect and mechanism of sodium tanshinone II A sulfonate
on microcirculatory disturbance of small intestine in rats with
sepsis. J Huazhong Univ Sci Technolog Med Sci.
31(441)2011.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Sun Y, Jin K, Xie L, Childs J, Mao XO,
Logvinova A and Greenberg DA: VEGF-induced neuroprotection,
neurogenesis, and angiogenesis after focal cerebral ischemia. J
Clin Invest. 111:1843–1851. 2003.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Liu J, Wang Y, Akamatsu Y, Lee CC, Stetler
RA, Lawton MT and Yang GY: Vascular remodeling after ischemic
stroke: Mechanisms and therapeutic potentials. Prog Neurobiol.
115:138–156. 2014.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Pedragosa J, Salas-Perdomo A, Gallizioli
M, Cugota R, Miró-Mur F, Briansó F, Justicia C, Pérez-Asensio F,
Marquez-Kisinousky L, Urra X, et al: CNS-border associated
macrophages respond to acute ischemic stroke attracting
granulocytes and promoting vascular leakage. Acta Neuropathol
Commun. 6(76)2018.PubMed/NCBI View Article : Google Scholar
|