1. Introduction
Nearly 10% of all global diseases are due to
pregnancy complications (1).
Recurrent pregnancy loss (RPL), pre-eclampsia (PE), fetal growth
restriction (FGR), and preterm delivery are examples of ‘great
obstetrical syndromes’ (GOS) (2).
Placental dysfunction is the most common pathogenesis of GOS
(3). The placenta starts to form
once implantation begins. The extravillous trophoblast cells (EVTs)
form the endometrium and move into the decidua. The process of
decidualization requires the morphological and functional changes
of uterine stromal cells and involvement of immune cells. Immune
cells account for nearly 40% of the decidual cells (4). The immune cells located in the
myometrium and decidua come into contact with EVT cells. The
removal of immune cells from the area of implantation or impeding
their signaling pathway could lead to miscarriages (5,6).
From the window of implantation to the first
trimester, most immune cells at the maternal-fetal interface are
uterine natural killer (uNK) cells (70%) (1). An increasing number of studies have
focused on the importance uNK cells in preserving the pregnancy
(7-9).
In normal human pregnancies, the effects of uNK cells involve
angiogenesis, promoting the remodeling of uterine spiral artery,
and improving the invasion of trophoblast cells (10-13).
Moreover, uNK cells supply killer immunoglobulin-like receptors
(KIRs), which come into contact with human leukocyte antigen-C
(HLA-C) ligands expressed by EVTs. The parents influence the HLA-C
genotypes in trophoblasts (14).
Since KIR and HLA-C are both polymorphic, the bindings of KIR and
HLA-C differ from one pregnancy to another. Numerous studies have
investigated the association between GOS and KIR/HLA-C combination
[for example, RPL (15-19),
PE (20-23),
and FGR (14)]. However, the
outcomes have not been conclusive.
Studies have indicated the deficiency of activation
as the reason for RPL, which is influenced by the improper binding
of KIR/HLA-C (16.24,25). The KIR/HLA-C combination modulates the
placental development physiologically. It contradicts the
traditional idea that the mother has an immune defense response to
the fetus (20). Specific KIR/HLA-C
binding occurs between maternal NK cells and EVT cells in the
decidua. This binding addresses activated tissue-remodeling issues
occurring in implantation and placentation. The present review
aimed to reveal the association between GOS and a combination of
KIR/HLA-C, which may aid high-risk pregnancy screening, strengthen
the treatment of pregnancy complications, and reduce the frequency
of adverse maternal and fetal outcomes. Related articles were
searched independently in PUBMED by two academics using the medical
subject headings: ‘KIR’, ‘HLA-C’, ‘recurrent pregnancy loss’,
‘recurrent miscarriages’, ‘pre-eclampsia’, ‘fetal growth
restriction’, or ‘pregnancy’. All articles were published from
January 2003 to November 2020 in English since there are few
related articles published before 2003. If a relationship exists
between the HLA-C haplotype and KIR, it may be useful to use the
KIR genotype and HLA-C molecule to select gametes from donors with
favorable combinations.
2. uNK cells and their receptors
During the first trimester of pregnancy, the
decrease of trophoblast invasion and poor recasting of uterine
spiral artery affect the blood flow in placenta, resulting in
pregnancy loss. NK cells are vital for the placental formation and
fetal development during implantation and the initial stage of
pregnancy. NK cells contain CD16 markers, which mediate
antibody-dependent cell-mediated cytotoxicity (ADCC). In general,
CD56dim NK cells belong to CD16+.
CD16+CD56dim NK cells have cytotoxicity,
while CD16-CD56bright NK cells exhibit
immunomodulatory effects (26).
Nearly 90% of uNK cells belong to the latter type. Hence, uNK cells
do not harm the fetus (27).
Numerous possibilities have been proposed about the origin of
decidual NK (dNK) cells; i) peripheral NK cells flow into the
uterus; ii) uterine stem cells differentiate into dNK cells; iii)
from bone marrow; iv) endometrial NK cells in the lining form dNK
cells by development and transformation (28). The dNK cells can produce numerous
cytokines, such as granulocyte-macrophage colony-stimulating factor
(GM-CSF), chemokine C motif ligand (XCL) 1, angiopoietin (Ang) 1,
Ang 2, vascular endothelial growth factor (VEGF), transforming
growth factor-β1 (TGF-β1), pleiotrophin and osteoglycin, which play
essential roles in sustaining uterine blood flow and fetal
development (10,13,29-32).
NK cell receptors include the immunoglobulin-like
transcripts (LILRB), C-type lectin heterodimer family (CD94:NKGs)
and KIRs (33). The inhibiting
NKG2A and activated NKG2C receptors are classified under the family
of CD94:NKGs, which could distinguish and combine with HLA-E
(34). HLA-G can discern the
inhibiting receptor LILRB1(35).
EVT cells supply class I HLA-C, non-classic HLA-G, and HLA-E
antigens (36,37). However, only the polymorphic HLA-C
ligands combine with KIRs expressed by uNK cells (33). The process of placentation is
affected by KIRs and HLA-C allotypes (15).
3. NK education and KIR/HLA-C
combination
Previously, studies involving mice (38) and humans (39-41)
have indicated that NK cells may experience ‘education’ and
‘licensing’ with the combined specific KIR and cognate HLA-C
allotypes to be effective. One likely scenario includes KIRs from
the mother interacting with her own HLA-C allotypes during the
growth of uNK cells, following this, ‘educating’ or ‘licensing’ the
uNK cells of the mother and modifying the method of interaction
with HLA-C allotypes of her child in the formation of the placenta
(14,42). However, how dNK cells educate,
license, or modulate in the uterus is yet to be established.
Considering NK cell education experiments, it has been revealed
that specific CD56bright NK cells react to interleukin
(IL)-12 or IL-15, and the licensing function was not strong in
resting NK cells, which implies that certain scenarios may neglect
the licensing needs (39). The dNK
cells exhibit an abundance of CD56(43), and release receptors and respond to
IL-15(44). Evidently, invasive
trophoblast cells are likely to increase the accumulation of dNK
cells to the maternal-fetal interface via the production of
different cytokines (13,45,46).
Regarding the education mechanism of NK cells in the association
between GOS and combined KIR/HLA-C, certain inhibiting KIRs and
HLA-C molecules from the father may lead to pregnancy
complications.
4. KIR family and HLA-C in pregnancy
The expression levels of KIR members partially
influence the response of NK cells. The polymorphic KIRs include
both activating and inhibitory receptors located on chromosome
19q13.4.11 in humans (47). KIRs
are extraordinarily polymorphic, and the types of genes located in
the KIR locus change prominently across different people and
regions. The phenotype of dNK cells renders them more likely to
express higher levels of KIRs specific for HLA-C molecules than
what appears in peripheral blood for the same individual (48). For each pregnancy, the maternal KIR
genes could be AA or Bx (47). KIR
AA haplotype is short of the majority of activating KIRs, except
KIR2DS4. However, the KIR Bx haplotype contains multiple activating
KIRs (49,50). AB and BB haplotypes are both
included in KIR Bx genotype (51).
HLA-C molecules are ligands for KIRs, which are sorted into two
types. HLA-C molecule with the C1 epitope (with asparagine at
location 80) combines with inhibiting KIR2DL2 and KIR2DL3. However,
HLA-C molecule with C2 epitope (with lysine at location 80)
connects with inhibiting KIR2DL1 and activating KIR2DS1. C2 is a
more potent ligand than C1(52).
Despite both HLA-C1 and HLA-C2 combining with KIR genes, the degree
of the downstream uNK activity is fundamentally affected through C1
and C2 zygosity in a specific pregnancy (53).
The KIR genotype appears comparatively later during
the growth of NK cells. These NK cells are induced by IL-15, which
is secreted by stromal cells to respond to progesterone in the
uterus (38). The uNK cells may
originate from the uterine progenitor cells instead of peripheral
blood NK cells. Hence, the KIR gene is changeable through
semaphores appearing in specific locations (54). The number of dNK cells during early
pregnancy exhibits a higher percentage of NK cells delivering KIRs
that can combine with HLA-C compared with the peripheral NK cells
and with uNK cells that originated from the endometrium of
non-pregnant females (55),
implying that HLA-C molecules on EVTs may be crucial for NK cell
education in the uterus.
Pregnancy is a unique physiological status, where
the particular non-self HLA molecules of KIRs could encounter their
cognate ligands. Polymorphic KIR/HLA-C binding has evolved
partially due to birth pressures occurring with human evolution and
the enlargement of brain tissues (42). Some bindings of KIR/HLA-C could be
unfavorable for a normal pregnancy than others. Activating KIRs
have protective effects during pregnancy complications, however,
people with activating KIRs are susceptible to other autoimmune
diseases (56). Insufficient
activation of NK cells may cause adverse maternal and neonatal
outcomes (57). If uNK cells gain
an activating signal in a female having a KIR B haplotype
(including KIR2DS1) and HLA-C2, the secretion of GM-CSF increases,
which can enhance the ability of migration and invasion of
trophoblast cells in cell lines (13).
5. KIR/HLA-C combination and GOS
KIR/HLA-C combination and RPL
RPL refers to the loss of embryos before twenty
weeks of gestation in three or more pregnancies; however, the
American Society for Reproductive Medicine (ASRM) defines it as two
times or more (58). RPL is caused
by numerous factors, such as chromosomal abnormalities, genetic
factors (26), anatomical factors,
endocrine abnormalities, and immune factors. Amidst these, the
etiology of certain patients is unknown and may be related to
autoimmune abnormalities. The correlation between KIR/HLA-C
combination and RPL is listed in Table
I, a few are inconsistent (14-17,19,25,26,59-65).
Reportedly, RPL females had an elevated rate of maternal KIR AA
haplotype and fetal HLA-C2 binding (14,15,60,66).
However, other empirical results were different (16-19).
A recent study reported a higher frequency of abortion after double
embryo transfers (DETs) in KIR AA females compared with KIR AB or
KIR BB (24). The genotype KIR AA
has an inhibitory effect, indicating the essentialness of
activation of uNK cells in early pregnancy. However, in a research
from northern India, it was revealed that RPL females had an
increased possibility of B haplotype rather than A haplotype
(53). Likewise, it was reported
that the KIR AA genotype has a protective effect on pregnant women
(16). However, there were some
shortcomings in this report; the sample size of the study was small
(n=40), and the abortion group was defined as a single spontaneous
abortion (16). Furthermore, Nowak
et al (17) indicated that
KIR AA and HLA-C1C2 females with HLA-C2C2 husbands had a higher
probability of a normal pregnancy. Pregnancies with C1/C1 fetuses
lead to a 2.5 times lower abortion rate relative to C2/C2
pregnancies (53). The higher rate
of successful pregnancies of C1/C1 babies did not emerge when the
transfer happened on an individual with a KIR B haplotype (53).
 | Table IExperiments on the association
between KIR/HLA-C combination and recurrent pregnancy loss. |
Table I
Experiments on the association
between KIR/HLA-C combination and recurrent pregnancy loss.
| Nationality | Authors
(Refs.) | Year of
publication | Experimental
group | Control group | Samples | Findings |
|---|
| Greek | Varla-Leftherioti
et al (59) | 2003 | RPL couples
(n=26) | Fertile couples
(n=26) | Peripheral
blood | RPL women had
decreased inhibiting KIR genes |
| Brazilian
Caucasian | Witt et al
(19) | 2004 | RPL women
(n=51) | Fertile women
(n=55) | NA | KIR genes did not
change between RPL patients and controls |
| Northern Irish | Flores et al
(60) | 2007 | RPL couples
(n=88) | Fertile couples
(n=30) and healthy controls (n=139) | NA | RPL patients had
decreased KIR2DL2; the KIR AA genotype was more common in RPL
women |
| Chinese | Wang et al
(61) | 2007 | RPL couples
(n=73) | Healthy couples
(n=68) | Peripheral
blood | RPL patients had
increased numbers of activating KIR genes including KIR2DS1 |
| European | Hiby et al
(15) | 2008 | RPL women (n=95)
and their partners (n=67) | Fertile women
(n=269) | Peripheral
blood | RPL women had a
higher frequency of KIR AA haplotype and decreased KIR2DS1 |
| Indian | Faridi et al
(25) | 2009 | RPL women
(n=205) | Healthy women
(n=224) | Peripheral
blood | RPL patients had
more activating KIR genes and they tended to be BB genotypes |
| South Brazilian
Caucasian | Vargas et al
(62) | 2009 | RPL couples
(n=68) | Fertile couples
(n=68) | Peripheral
blood | Females having more
activating KIR genes were three times more likely to develop
RPL |
| European | Hiby et al
(14) | 2010 | RPL women (n=115)
and their partners (n=81) | Fertile couples
(n=592) | Placental tissues
and maternal blood | RPL patients were
less frequent in women with a KIR B haplotype |
| Polish | Nowak et al
(17) | 2011 | RPL couples
(n=125) | Fertile couples
(n=117) | Peripheral
blood | Among KIR AA
females who had HLA-C2C2 partners, HLA-C1C2 females tended to have
a normal pregnancy |
| Turkish | Ozturk et al
(16) | 2012 | RPL women
(n=40) | Fertile women
(n=90) | Peripheral
blood | RPL patients tended
to be KIR Bx genotypes |
| Chinese | Wang et al
(63) | 2014 | RPL women
(n=30) | Fertile women
(n=30) | Decidual
tissues | There was decreased
frequency of CD56+ CD16− natural killer cell
staining for KIR2DL1/ S1 and KIR2DL2/S2/L3 in RPL women |
| American | Dambaeva et
al (64) | 2016 | RPL women (n=139)
and their partners (n=42) | HLA-C controls:
1,070 North American Caucasian Population (n=1,070) KIR controls:
American Caucasian population (n=255) | Peripheral
blood | KIR2DS1-positive
RPL females had increased HLA-C2 |
| Turkey | Elbaşı et al
(65) | 2020 | RPL couples
(n=25) | Healthy couples
(n=39) | Peripheral
blood | HLA-C2C2 were more
common in RPL partners; KIR2DL5 gene was more common in RPL
couples |
| Caucasian | Yang et al
(26) | 2020 | RPL patients
(n=160) and their partners (n=99) | HLA-C controls:
1,070 North American Caucasian Population (n=1,070) KIR controls:
American Caucasian population (n=255) | Women: peripheral
blood; Partners: buccal cells | KIR2DL2-positive
RPL females had lower HLA-C1C1, and their partners had increased
HLA-C2C2 compared with KIR2DL2 negative RPL patients |
A few empirical studies have indicated that RPL
females may lack proper KIR genes (e.g., KIR2DL1 and KIR2DL2) that
can combine with HLA-C in trophoblast cells and transfer inhibitory
signals for NK cell activity (59,60,67-71).
Compared with the normal controls, the levels of KIR2DL1/S1 on
CD56+CD16− NK cells in the deciduae of RPL
females were reduced (63). HLA-C1
is the ligand for KIR2DL2. Compared with healthy females, RPL
patients exhibited higher KIR2DL2 and HLA-C2/C2 or HLA-C2/x
combinations (65). Initially, the
authors established that KIRs in partners could not directly result
in a miscarriage; however, since the fetal KIR genotype is unclear,
distributed fetal cells with potentially unfavorable KIRs can
accelerate the ending course (65).
It was recently reported that KIR2DL2-positive RPL females
exhibited decreased molecules of HLA-C1 compared with
KIR2DL2-negative RPL females (P<0.05) (26). Reduced ligands for inhibiting KIRs
may cause inadequate inhibition of maternal uNK cells to the
trophoblast, thus giving rise to RPL.
Some studies have revealed an apparent increase in
the frequency of activating KIRs in RPL females (60-62,72).
Patients with elevated activation of KIRs were nearly three times
more susceptible to RPL (62).
KIR2DS2 and KIR2DS3 are both activating KIRs, which are hazardous
elements for RPL (73). The KIR
genotypes in 205 RPL women and 224 normal subjects were analyzed
(25). Accordingly, RPL patients
had more activating KIRs than normal controls (25). Similar results were obtained in the
Chinese Han population (61), the
authors suggested that relative to fertile females, there was a
lower rate of C2+ HLA-C molecules in RPL women (61). Reportedly, an elevated rate of
KIR2DS2 in HLA-C1 homozygous parents was associated with RPL
(16). Consequently, the
physiological reason underlying the high frequency of activating
KIR genotypes could be attributed to an extensive scope of diverse
activating KIR receptors making NK cells to discriminate more
activation ligands, which originated from fetal cells. This causes
an induced percentage of cytotoxic and apoptotic signals; these
fight the inhibiting function of receptors against semiallogeneic
fetuses at the maternal-fetal interface and end in abortion.
It has been suggested that KIR2DS1 assumes a
critical role in pregnancy, mainly because activation of dNK cells
through KIR2DS1 induces the production of soluble mediators, such
as GM-CSF. Reportedly, GM-CSF has been confirmed to be useful for
placentation and promoting fetal growth (74). Additionally, C2+ HLA-C
molecules were revealed to be more common among partners of
KIR2DS1+ women in the Caucasian population (64). One possible reason is that NK cell
education may lead to the hypo-responsiveness of
KIR2DS1+ deciduae, and inhibit the positive effect of
KIR2DS1 in a normal pregnancy. Since C2+ HLA-C molecules
are supposed to combine with KIR2DS1, it is hard to conclusively
elucidate the potential function of activating KIR2DS1 in the
absence of its ligand.
On the other hand, in the Brazilian population,
there was no difference between KIR genes and susceptibility to RPL
(19). Therefore, future studies on
the functionality of KIRs and their ligands should include a higher
sample size of RPL couples in a specific area compared with fertile
controls in the same region. Preferably, genotyping of embryos
should also be included.
KIR/HLA-C combination and PE
PE is a severe pregnancy complication; it is
characterized by hypertension (≧140/90 mmHg) and proteinuria
following twenty weeks of pregnancy. Additionally, the prevalence
of PE is 5-8% (75). Since the
invasion ability of the trophoblast is affected, inappropriate
combinations of KIR/HLA-C are reasons that cause PE. Reportedly, if
a woman carries the KIR AA genotype and her baby has HLA-C2 from
the father, then the possibility of PE in the pregnancy was
increased (20,76) (Table
II). This is because KIR AA/HLA-C2 binding may lead to
inadequate activation of uNK cells, affecting uterine artery
recasting and leading to PE. Furthermore, Hiby et al
(14) revealed a higher PE
incidence for females whose fetuses inherited another C2 from
males. For women with the KIR AA genotype, the pregnancy with fetal
HLA-C1C1 is more prone to PE than that of HLA-C1C2 or HLA-C2C2.
According to the NK education theory, maternal HLA-C2 molecules
educate KIR2DL1+ uNK cells to obtain functional
properties during period of rest rather than during an immune
response, and counteract the inhibitory input of fetal HLA-C2
molecules in pregnancy (14). A
method for estimating KIR2DL1 allotypes coded by the diverse KIR
genes has promoted the development of various anti-KIR antibodies
and genotyping to indicate that KIR2DL1 on the KIR A instead of the
KIR B haplotype is related to the higher probability of PE
(77). Mexican researchers
collected ten normal pregnant decidua tissues and nine PE decidua
samples during cesarean section (22). It was revealed that there were more
inhibitory KIRs in deciduae of PE patients than those in normal
pregnancies (22). Due to the lack
of activating KIRs, the activation signals cannot be provided
concurrently. Thus, NK cells in these females exhibited a decreased
physiological effect, without adequate support for placental
development.
| Table IIExperiments on the relationship
between KIR/HLA-C combination and PE. |
Table II
Experiments on the relationship
between KIR/HLA-C combination and PE.
| Nationality | Authors
(Refs.) | Year of
publication | Experimental
group | Control group | Samples | Findings |
|---|
| British | Hiby et al
(20) | 2004 | PE patients and
their fetuses (n=200) | Normal pregnant
women and their fetuses (n=201) | Maternal blood,
cord samples or neonatal mouth swabs | The female with an
AA genotype and a fetus with the HLA-C2 molecule tended to have
PE |
| British | Hiby et al
(14) | 2010 | PE couples (n=742)
and their fetuses (n=733) | Normal pregnant
couples (n=592) and their fetuses (n=423) | Maternal blood,
cord samples or neonatal mouth swabs | There was a higher
PE incidence for females whose fetuses inherited another C2 from
males |
| Mexican | Sanchez- Rodriguez
et al (22) | 2011 | PE patients
(n=9) | Normal pregnant
women (n=10) | Decidual
samples | There were more
inhibitory KIRs in deciduae of PE patients than those in normal
pregnancies |
| Ugandan | Nakimuli et
al (21) | 2015 | PE patients
(n=254) | Normal pregnant
women (n=484) | Maternal blood,
cord samples | KIR2DS5 on Cen-B of
Africans protected pregnancies from PE |
| Chinese Han
population | Long et al
(79) | 2015 | PE patients
(n=271) | Normal pregnant
women (n=295) | Maternal blood | Reduced KIR
activation was observed in PE women |
| British | Huhn et al
(77) | 2018 | PE patients
(n=693) | Normal pregnant
women (n=679) | Maternal blood | KIR2DL1A was
associated with increased PE risk |
| Danish | Larsen et al
(84) | 2019 | Severe PE patients
(n=259) | Normal pregnant
women (n=259) | Maternal and
neonatal blood | A relationship was
not found between maternal KIR genotypes and HLA-C in the
fetuses |
| Ethiopian | Kelemu et al
(76) | 2020 | PE patients
(n=131) | Normal pregnant
women (n=157) | Maternal blood,
cord samples | There was a
significant association between KIR AA genotype and PE |
Reduced KIR activation was observed in PE women
(78,79). In Europe and Ethiopia, the
protective function of the maternal B haplotype is regulated by an
activating C2 receptor, called KIR2DS1 (20,76,80).
It stimulates uNK cells that increase the angiogenesis and immune
reaction, which induces a successful pregnancy (81). KIR2DS4 and KIR2DS5 exhibited
protective effects from PE development in Caucasians and Africans,
respectively (21,30,82).
Clinically, in some patients, the resistance index of the uterine
artery increased, which indicated poor uterine artery recasting,
and the levels of KIR2DL/S1, 3, and 5 were simultaneously reduced
on the portion of dNK cells (78).
The change of blood flow in the uterine artery reflects the
clinical application of combined KIR/HLA-C in PE (83). In a Chinese study with a large
sample size, it was revealed that PE women exhibited decreased
activating receptors, including KIR2DS2, KIR2DS3, and
KIR2DS5(83). Notably, the total
number of activating KIRs in PE patients was less than that in
controls (P=0.03) (83).
However, it has been established that Caucasian
partners are more likely to have HLA-C2 molecules than Japanese
men. Therefore, Japanese women having children with Caucasian
partners are more likely to develop PE than those with Japanese
men. Nevertheless, a previous study has reported a 1.54% chance of
developing PE with a Caucasian partner, compared with 2.67% with a
Japanese partner for a Japanese woman (78). A recent study in Denmark involved
259 patients with PE or eclampsia and 259 normal controls (84). Peripheral blood samples were
obtained from both mothers and newborns (84). A relationship was not revealed
between maternal KIR genotypes and HLA-C in the fetuses (84). Since the babies had additional
HLA-C2 molecules compared with the mothers, there was no report of
any difference in maternal KIR AA genotype between the experimental
and control groups (84).
KIR/HLA-C combination and FGR
Hiby et al conducted a trial involving 118
FGR patients (fetal weight less than or equal to the 5th
percentage) (14). Allegedly, the
binding of maternal KIR AA genotype and fetal HLA-C2 was related to
an induced possibility of developing FGR (14). This trend was consistent with PE
previously discussed (14).
6. KIR/HLA-C combination and birth
weight
Human birth weight is influenced by stable selection
(42). Macrosomia may cause
obstructive dystocia, shoulder dystocia, and postpartum hemorrhage,
affecting maternal and neonatal health (42). Often, a fetus that is too small will
find it difficult to survive (42).
An abundance of research has indicated that this process is
regulated by the KIR genotype and their relevant HLA-C ligands on
invasive trophoblast cells (42).
Females having a KIR AA genotype and a baby with a paternal HLA-C2
molecule have restrained production of cytokines by uNK cells,
which results in delivering a small fetus (85). Since the combination of KIR2DS1 and
HLA-C2 can induce cytokine production and promote trophoblast
invasion, the female having a KIR2DS1 gene and an HLA-C2 baby tends
to have a large fetus (74,86). If the fetus is HLA-C1C1, the
function of maternal KIR is invalid, and this correlation does not
exist (14,74), which is attributable to the strict
peculiarity and obvious restraining granted by KIR2DL1/HLA-C2
bindings relative to the weaker KIR2DL2/3 bindings with HLA-C
alleles (87). In summary, in the
placentation region, all effects of HLA-C in pregnancy are
regulated by paternal HLA-C2, and the maternal KIR genotype decides
the size of the baby (Fig. 1).
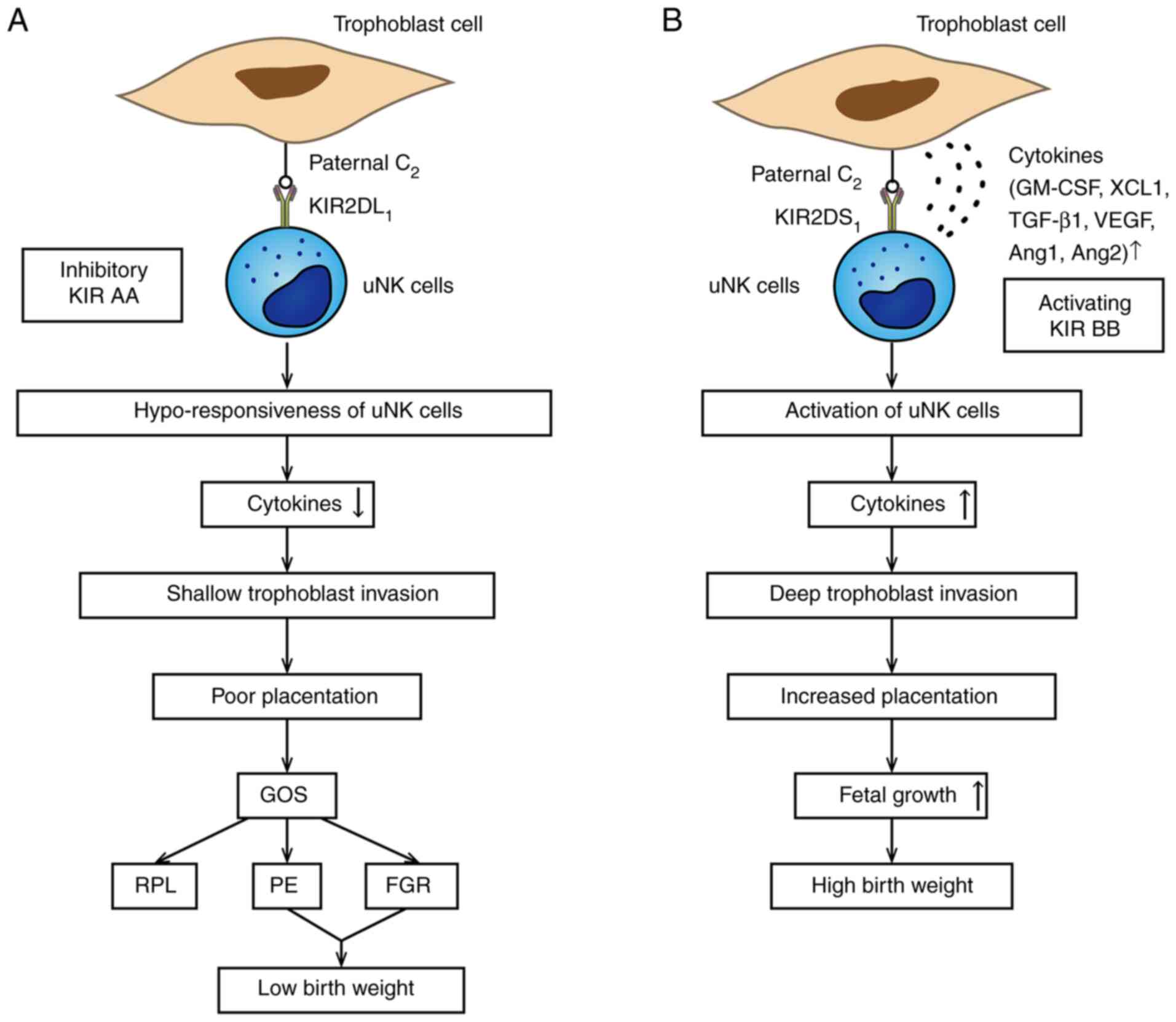 | Figure 1Association between KIR/HLA-C
combination and birth weight. (A) In this situation, the neonate
inherits an HLA-C2 molecule from the father. The mother is KIR AA
genotype; KIR2DL1 strongly combines to trophoblast HLA-C2 alleles
leading to hypo-responsiveness of uNK cells, which is related to
the poor placentation and leads to low birth weight in GOS. (B)
Binding of KIR2DS1/HLA-C2 is associated with high birth weight. The
KIR BB haplotype includes the activating KIR2DS1. Once this
happens, uNK cells are activated to secrete plenty of cytokines,
such as GM-CSF, which could enhance the placentation and lead to
higher birth weight. KIR, killer-cell immunoglobulin-like receptor;
HLA, human leukocyte antigen; uNK, uterine natural killer; GOS,
great obstetrical syndromes; RPL, recurrent pregnancy loss; PE,
preeclampsia; FGR, fetal growth restriction; GM-CSF,
granulocyte-macrophage colony-stimulating factor; XCL1, chemokine C
motif ligand 1; TGF-β1, transforming growth factor-β1; VEGF,
vascular endothelial growth factor; Ang 1, angiopoietin 1. |
There are significant differences in the
trophoblasts of mice and human beings. Compared with human
trophoblasts, the invasion of trophoblasts into deciduae is
shallower in mice, and the mice do not express non-classical MHC
(88). Murine NK receptors are part
of the Ly49 family of receptors. In the mice experiment, when the
additional MHC molecule H-2Dd was added, the vessel
remodeling ability was decreased, compared with the same type of
mice deficient only for H-2Dd (89). H-2Dd combines with the
inhibitory receptor Ly49A and can suppress additional uNK cells.
Noticeably, regardless of whether there was an H-2Dd
molecule from the father or not, the fetal growth was affected.
According to these results, some bindings of maternal NK receptors
and paternal (or maternal) MHC molecules could affect trophoblast
invasion and vascular recasting in mice. Therefore, it can be
concluded that in both human and mice, the over inhibition of the
function of uNK cells could result in fetal weight loss.
7. Future study directions
There are potentially critical clinical applications
in these empirical studies. Considering the RPL woman with a KIR AA
haplotype, the HLA-C molecules of the female and her partner are
usable as a basis for selecting the donor egg or sperm. According
to the epidemiology, donors homogeneous for HLA-C1 are considered
to be safe; meanwhile, C2/C2 partners or oocyte donors may be
harmful (14,90,91).
HLA-C and the KIR genotype are applicable in selecting third-party
gametes or surrogates (53).
Subsequently, these immunological indicators can be detected
through laboratory tests, which can be adopted to predict pregnancy
outcomes. By detecting the KIR genotype and HLA-C molecule, the
possible ligands can be inferred. The optimal binding will cause
activation of uNK cells and angiogenesis in placentation, leading
to a normal pregnancy. Females having an improper binding may
experience difficulties during their pregnancies. The
functionalities of these novel immune reactions have caused renewed
comprehension of how to protect the fetus and may be useful to
estimate and treat GOS. Detection of KIR genes in GOS patients
could potentially assume a vital role in diagnosing the alloimmune
etiology in these diseases. Studies conducted on a large-scale,
employing modified statistical model sets, including the
quantitative maternal-fetal genotype experiment, called a linear
mixed-effect model (86), would
enhance pregnancy outcomes. It entails statistically entering the
KIR copy number from the SNP genotype (92) or utilizing a full-length transcript
Smart-seq2 data (93).
Nevertheless, certain scholars claim that more
conclusiveness is needed before clinical application despite the
indications pointing towards credible evidence (94). For example, the binding of the KIR
AA genotype and fetal HLA-C2 exists in roughly 10% of Caucasian
pregnant females, making them twice as likely to have PE as normal
women. Despite elevating the risk, there is a weak clinical
application in this association. The only way to use this data
predictively is to combine it with other clinical risk factors and
serological indicators. KIR gene expression is highly polymorphic,
and other factors that can regulate NK cells. Moreover, there are
numerous ligands for activating receptors, and the mechanism of
modulating NK cells is not clearly outlined. Furthermore, the exact
manner of KIR/HLA-C combination in regulating NK cell education has
not been established. Therefore, novel technologies are useful to
distinguish the complex diversity of KIR and HLA-C. Sophisticated
laboratory experiments can help determine the specific role of NK
cells. Currently, studies detecting the function of uNK cells in
vitro have not been conducted. The function of uNK cells can
only be determined indirectly by detecting some cytokines, which
are generally used to detect the functionality of peripheral blood
NK cells. Mouse models can help understand the function of uNK
cells. Some transgenic mice containing specific KIR and HLA-C
molecules can help study the effect of distinct binding modes in
vivo.
8. Conclusion
Medawar's theory states that immune cells in
peripheral blood and the uterus need to be suppressed to form a
successful pregnancy (95).
However, KIR genotypes and HLA-C molecules reexamine the conditions
for a successful pregnancy. Sufficient activation of uNK cells is
crucial to maintain a normal pregnancy. Emphasis should be given on
solving the following problems: Adopting accurate methods to
determine KIR genotypes (e.g., by high-throughput genotyping),
conforming with uniform clinical inclusion criteria, understanding
the interaction between EVT and uNK cells. Moreover, the
interaction between KIR/HLA-C and GOS should be studied by
targeting women in a specific region since it is difficult to
compare studies in different areas. Thus, a large sample-sized
research in the same region, having a comparable control
population, accurate detection of maternal KIRs, and both parents
and fetal HLA-C molecules may determine the function of KIR and
HLA-C in GOS.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported (grant no. 2020-MS-167)
by the Natural Science Foundation of Liaoning Province (China).
Availability of data and materials
Not applicable.
Authors' contributions
XY and TM conceived the study, performed the
literature search and analyzed the data. XY wrote the manuscript.
TM revised the work for important intellectual content. XY and TM
confirm the authenticity of all the raw data. Both authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lopes MRU, Danowski A, Funke A, Rego J,
Levy R and Andrade DCO: Update on antiphospholipid antibody
syndrome. Rev Assoc Med Bras. 63:994–999. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Egeland GM, Skurtveit S, Staff AC, Eide
GE, Daltveit AK, Klungsøyr K, Trogstad L, Magnus PM, Brantsæter AL
and Haugen M: Pregnancy-related risk factors are associated with a
significant burden of treated hypertension within 10 years of
delivery: Findings from a population-based norwegian cohort. J Am
Heart Assoc. 7(e008318)2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Brosens I, Pijnenborg R, Vercruysse L and
Romero R: The ‘Great Obstetrical Syndromes’ are associated with
disorders of deep placentation. Am J Obstet Gynecol. 204:193–201.
2011.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Vince GS, Starkey PM, Jackson MC, Sargent
IL and Redman CW: Flow cytometric characterisation of cell
populations in human pregnancy decidua and isolation of decidual
macrophages. J Immunol Methods. 132:181–189. 1990.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Abrahams VM, Straszewski-Chavez SL, Guller
S and Mor G: First trimester trophoblast cells secrete Fas ligand
which induces immune cell apoptosis. Mol Hum Reprod. 10:55–63.
2004.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Moffett-King A: Natural killer cells and
pregnancy. Nat Rev Immunol. 2:656–663. 2002.PubMed/NCBI View
Article : Google Scholar
|
|
7
|
Yang X, Gilman-Sachs A and Kwak-Kim J:
Ovarian and endometrial immunity during the ovarian cycle. J Reprod
Immunol. 133:7–14. 2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Colucci F: The role of KIR and HLA
interactions in pregnancy complications. Immunogenetics.
69:557–565. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Moffett A and Colucci F: Uterine NK cells:
Active regulators at the maternal-fetal interface. J Clin Invest.
124:1872–1879. 2014.PubMed/NCBI View
Article : Google Scholar
|
|
10
|
Xiong S, Sharkey AM, Kennedy PR, Gardner
L, Farrell LE, Chazara O, Bauer J, Hiby SE, Colucci F and Moffett
A: Maternal uterine NK cell-activating receptor KIR2DS1 enhances
placentation. J Clin Invest. 123:4264–4272. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
11
|
Bulmer JN, Innes BA, Levey J, Robson SC
and Lash GE: The role of vascular smooth muscle cell apoptosis and
migration during uterine spiral artery remodeling in normal human
pregnancy. FASEB J. 26:2975–2985. 2012.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Li C, Houser BL, Nicotra ML and Strominger
JL: HLA-G homodimer-induced cytokine secretion through HLA-G
receptors on human decidual macrophages and natural killer cells.
Proc Natl Acad Sci USA. 106:5767–5772. 2009.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Hanna J, Goldman-Wohl D, Hamani Y, Avraham
I, Greenfield C, Natanson-Yaron S, Prus D, Cohen-Daniel L, Arnon
TI, Manaster I, et al: Decidual NK cells regulate key developmental
processes at the human fetal-maternal interface. Nat Med.
12:1065–1074. 2006.PubMed/NCBI View
Article : Google Scholar
|
|
14
|
Hiby SE, Apps R, Sharkey AM, Farrell LE,
Gardner L, Mulder A, Claas FH, Walker JJ, Redman CW, Morgan L, et
al: Maternal activating KIRs protect against human reproductive
failure mediated by fetal HLA-C2. J Clin Invest. 120:4102–4110.
2010.PubMed/NCBI View
Article : Google Scholar
|
|
15
|
Hiby SE, Regan L, Lo W, Farrell L,
Carrington M and Moffett A: Association of maternal killer-cell
immunoglobulin-like receptors and parental HLA-C genotypes with
recurrent miscarriage. Hum Reprod. 23:972–976. 2008.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ozturk OG, Sahin G, Karacor ED and
Kucukgoz U: Evaluation of KIR genes in recurrent miscarriage. J
Assist Reprod Genet. 29:933–938. 2012.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Nowak I, Malinowski A, Tchorzewski H,
Barcz E, Wilczynski JR, Banasik M, Grybos M, Kurpisz M, Luszczek W,
Majorczyk E, et al: HLA-C C1C2 heterozygosity may protect women
bearing the killer immunoglobulin-like receptor AA genotype from
spontaneous abortion. J Reprod Immunol. 88:32–37. 2011.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Faridi RM and Agrawal S: Killer
immunoglobulin-like receptors (KIRs) and HLA-C allorecognition
patterns implicative of dominant activation of natural killer cells
contribute to recurrent miscarriages. Hum Reprod. 26:491–497.
2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Witt CS, Goodridge J, Gerbase-Delima MG,
Daher S and Christiansen FT: Maternal KIR repertoire is not
associated with recurrent spontaneous abortion. Hum Reprod.
19:2653–2657. 2004.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Hiby SE, Walker JJ, O'Shaughnessy KM,
Redman CW, Carrington M, Trowsdale J and Moffett A: Combinations of
maternal KIR and fetal HLA-C genes influence the risk of
preeclampsia and reproductive success. J Exp Med. 200:957–965.
2004.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Nakimuli A, Chazara O, Hiby SE, Farrell L,
Tukwasibwe S, Jayaraman J, Traherne JA, Trowsdale J, Colucci F,
Lougee E, et al: A KIR B centromeric region present in Africans but
not Europeans protects pregnant women from pre-eclampsia. Proc Natl
Acad Sci USA. 112:845–850. 2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Sánchez-Rodríguez EN, Nava-Salazar S,
Mendoza-Rodríguez CA, Moran C, Romero-Arauz JF, Ortega E, Granados
J, Cervantes-Peredo A and Cerbón M: Persistence of decidual NK
cells and KIR genotypes in healthy pregnant and preeclamptic women:
A case-control study in the third trimester of gestation. Reprod
Biol Endocrinol. 9(8)2011.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Yang X, Yang Y, Yuan Y, Liu L and Meng T:
The roles of uterine natural killer (NK) cells and KIR/HLA-C
combination in the development of preeclampsia: A systematic
review. Biomed Res Int. 2020(4808072)2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Alecsandru D, Barrio A, Garrido N,
Aparicio P, Pellicer A, Moffett A and García-Velasco JA: Parental
human leukocyte antigen-C allotypes are predictive of live birth
rate and risk of poor placentation in assisted reproductive
treatment. Fertil Steril. 114:809–817. 2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Faridi RM, Das V, Tripthi G, Talwar S,
Parveen F and Agrawal S: Influence of activating and inhibitory
killer immunoglobulin-like receptors on predisposition to recurrent
miscarriages. Hum Reprod. 24:1758–1764. 2009.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Yang X, Yang E, Wang WJ, He Q, Jubiz G,
Katukurundage D, Dambaeva S, Beaman K and Kwak-Kim J: Decreased
HLA-C1 alleles in couples of KIR2DL2 positive women with recurrent
pregnancy loss. J Reprod Immunol. 142(103186)2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Sacks G: Enough! Stop the arguments and
get on with the science of natural killer cell testing. Hum Reprod.
30:1526–1531. 2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Yagel S: The developmental role of natural
killer cells at the fetal-maternal interface. Am J Obstet Gynecol.
201:344–350. 2009.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Abbas Y, Oefner CM, Polacheck WJ, Gardner
L, Farrell L, Sharkey A, Kamm R, Moffett A and Oyen ML: A
microfluidics assay to study invasion of human placental
trophoblast cells. J R Soc Interface. 14(20170131)2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Kennedy PR, Chazara O, Gardner L, Ivarsson
MA, Farrell LE, Xiong S, Hiby SE, Colucci F, Sharkey AM and Moffett
A: Activating KIR2DS4 is expressed by uterine NK cells and
contributes to successful pregnancy. J Immunol. 197:4292–4300.
2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Fu B, Zhou Y, Ni X, Tong X, Xu X, Dong Z,
Sun R, Tian Z and Wei H: Natural killer cells promote fetal
development through the secretion of growth-promoting factors.
Immunity. 47:1100–1113.e6. 2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Lash GE, Schiessl B, Kirkley M, Innes BA,
Cooper A, Searle RF, Robson SC and Bulmer JN: Expression of
angiogenic growth factors by uterine natural killer cells during
early pregnancy. J Leukoc Biol. 80:572–580. 2006.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Parham P and Moffett A: Variable NK cell
receptors and their MHC class I ligands in immunity, reproduction
and human evolution. Nat Rev Immunol. 13:133–144. 2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Lanier LL: NK cell recognition. Annu Rev
Immunol. 23:225–274. 2005.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Shiroishi M, Tsumoto K, Amano K,
Shirakihara Y, Colonna M, Braud VM, Allan DS, Makadzange A,
Rowland-Jones S, Willcox B, et al: Human inhibitory receptors
Ig-like transcript 2 (ILT2) and ILT4 compete with CD8 for MHC class
I binding and bind preferentially to HLA-G. Proc Natl Acad Sci USA.
100:8856–8861. 2003.PubMed/NCBI View Article : Google Scholar
|
|
36
|
King A, Hiby SE, Gardner L, Joseph S,
Bowen JM, Verma S, Burrows TD and Loke YW: Recognition of
trophoblast HLA class I molecules by decidual NK cell receptors-a
review. Placenta. 21 (Suppl A):S81–S85. 2000.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Apps R, Murphy SP, Fernando R, Gardner L,
Ahad T and Moffett A: Human leucocyte antigen (HLA) expression of
primary trophoblast cells and placental cell lines, determined
using single antigen beads to characterize allotype specificities
of anti-HLA antibodies. Immunology. 127:26–39. 2009.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Kim S, Poursine-Laurent J, Truscott SM,
Lybarger L, Song YJ, Yang L, French AR, Sunwoo JB, Lemieux S,
Hansen TH and Yokoyama WM: Licensing of natural killer cells by
host major histocompatibility complex class I molecules. Nature.
436:709–713. 2005.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Anfossi N, André P, Guia S, Falk CS,
Roetynck S, Stewart CA, Breso V, Frassati C, Reviron D, Middleton
D, et al: Human NK cell education by inhibitory receptors for MHC
class I. Immunity. 25:331–342. 2006.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Yu J, Heller G, Chewning J, Kim S,
Yokoyama WM and Hsu KC: Hierarchy of the human natural killer cell
response is determined by class and quantity of inhibitory
receptors for self-HLA-B and HLA-C ligands. J Immunol.
179:5977–5989. 2007.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Fauriat C, Ivarsson MA, Ljunggren HG,
Malmberg KJ and Michaëlsson J: Education of human natural killer
cells by activating killer cell immunoglobulin-like receptors.
Blood. 115:1166–1174. 2010.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Moffett A and Colucci F: Co-evolution of
NK receptors and HLA ligands in humans is driven by reproduction.
Immunol Rev. 267:283–297. 2015.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Koopman LA, Kopcow HD, Rybalov B, Boyson
JE, Orange JS, Schatz F, Masch R, Lockwood CJ, Schachter AD, Park
PJ and Strominger JL: Human decidual natural killer cells are a
unique NK cell subset with immunomodulatory potential. J Exp Med.
198:1201–1212. 2003.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Verma S, Hiby SE, Loke YW and King A:
Human decidual natural killer cells express the receptor for and
respond to the cytokine interleukin 15. Biol Reprod. 62:959–968.
2000.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Drake PM, Gunn MD, Charo IF, Tsou CL, Zhou
Y, Huang L and Fisher SJ: Human placental cytotrophoblasts attract
monocytes and CD56(bright) natural killer cells via the actions of
monocyte inflammatory protein 1alpha. J Exp Med. 193:1199–1212.
2001.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Hanna J, Wald O, Goldman-Wohl D, Prus D,
Markel G, Gazit R, Katz G, Haimov-Kochman R, Fujii N, Yagel S, et
al: CXCL12 expression by invasive trophoblasts induces the specific
migration of CD16-human natural killer cells. Blood. 102:1569–1577.
2003.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Hsu KC, Chida S, Geraghty DE and Dupont B:
The killer cell immunoglobulin-like receptor (KIR) genomic region:
Gene-order, haplotypes and allelic polymorphism. Immunol Rev.
190:40–52. 2002.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Alecsandru D and García-Velasco JA: Why
natural killer cells are not enough: A further understanding of
killer immunoglobulin-like receptor and human leukocyte antigen.
Fertil Steril. 107:1273–1278. 2017.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Bashirova AA, Martin MP, McVicar DW and
Carrington M: The killer immunoglobulin-like receptor gene cluster:
Tuning the genome for defense. Annu Rev Genomics Hum Genet.
7:277–300. 2006.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Robinson J, Halliwell JA, McWilliam H,
Lopez R and Marsh SG: IPD-the immuno polymorphism database. Nucleic
Acids Res. 41(Database issue):D1234–D1240. 2013.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Gonzalez-Galarza FF, Christmas S,
Middleton D and Jones AR: Allele frequency net: A database and
online repository for immune gene frequencies in worldwide
populations. Nucleic Acids Res. 39(Database issue):D913–D919.
2011.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Winter CC, Gumperz JE, Parham P, Long EO
and Wagtmann N: Direct binding and functional transfer of NK cell
inhibitory receptors reveal novel patterns of HLA-C allotype
recognition. J Immunol. 161:571–577. 1998.PubMed/NCBI
|
|
53
|
Morin SJ, Treff NR, Tao X, Scott RT III,
Franasiak JM, Juneau CR, Maguire M and Scott RT: Combination of
uterine natural killer cell immunoglobulin receptor haplotype and
trophoblastic HLA-C ligand influences the risk of pregnancy loss: A
retrospective cohort analysis of direct embryo genotyping data from
euploid transfers. Fertil Steril. 107:677–683.e2. 2017.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Male V, Hughes T, McClory S, Colucci F,
Caligiuri MA and Moffett A: Immature NK cells, capable of producing
IL-22, are present in human uterine mucosa. J Immunol.
185:3913–3918. 2010.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Male V, Sharkey A, Masters L, Kennedy PR,
Farrell LE and Moffett A: The effect of pregnancy on the uterine NK
cell KIR repertoire. Eur J Immunol. 41:3017–3027. 2011.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Hilton HG and Parham P: Missing or altered
self: Human NK cell receptors that recognize HLA-C. Immunogenetics.
69:567–579. 2017.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Hong Y, Wang X, Lu P, Song Y and Lin Q:
Killer immunoglobulin-like receptor repertoire on uterine natural
killer cell subsets in women with recurrent spontaneous abortions.
Eur J Obstet Gynecol Reprod Biol. 140:218–223. 2008.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Practice Committee of the American Society
for Reproductive Medicine. Definitions of infertility and recurrent
pregnancy loss. Fertil Steril. 89(1603)2008.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Varla-Leftherioti M, Spyropoulou-Vlachou
M, Niokou D, Keramitsoglou T, Darlamitsou A, Tsekoura C,
Papadimitropoulos M, Lepage V, Balafoutas C and Stavropoulos-Giokas
C: Natural killer (NK) cell receptors' repertoire in couples with
recurrent spontaneous abortions. Am J Reprod Immunol. 49:183–191.
2003.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Flores AC, Marcos CY, Paladino N, Arruvito
L, Williams F, Middleton D and Fainboim L: KIR receptors and HLA-C
in the maintenance of pregnancy. Tissue Antigens. 69 (Suppl
1):S112–S113. 2007.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Wang S, Zhao YR, Jiao YL, Wang LC, Li JF,
Cui B, Xu CY, Shi YH and Chen ZJ: Increased activating killer
immunoglobulin-like receptor genes and decreased specific HLA-C
alleles in couples with recurrent spontaneous abortion. Biochem
Biophys Res Commun. 360:696–701. 2007.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Vargas RG, Bompeixe EP, Franca PP, Marques
de Moraes M and da Graca Bicalho M: Activating killer cell
immunoglobulin-like receptor genes' association with recurrent
miscarriage. Am J Reprod Immunol. 62:34–43. 2009.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Wang S, Li YP, Ding B, Zhao YR, Chen ZJ,
Xu CY, Fu YB and Wang XT: Recurrent miscarriage is associated with
a decline of decidual natural killer cells expressing killer cell
immunoglobulin-like receptors specific for human leukocyte antigen
C. J Obstet Gynaecol Res. 40:1288–1295. 2014.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Dambaeva SV, Lee DH, Sung N, Chen CY, Bao
S, Gilman-Sachs A, Kwak-Kim J and Beaman KD: Recurrent pregnancy
loss in women with killer cell immunoglobulin-like receptor KIR2DS1
is associated with an increased HLA-C2 allelic frequency. Am J
Reprod Immunol. 75:94–103. 2016.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Elbaşı MO, Tulunay A, Karagözoğlu H,
Kahraman S and Ekşioğlu-Demiralp E: Maternal killer-cell
immunoglobulin-like receptors and paternal human leukocyte antigen
ligands in recurrent pregnancy loss cases in Turkey. Clin Exp
Reprod Med. 47:122–129. 2020.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Alecsandru D, Garrido N, Vicario JL,
Barrio A, Aparicio P, Requena A and García-Velasco JA: Maternal KIR
haplotype influences live birth rate after double embryo transfer
in IVF cycles in patients with recurrent miscarriages and
implantation failure. Hum Reprod. 29:2637–2643. 2014.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Varla-Leftherioti M, Spyropoulou-Vlachou
M, Keramitsoglou T, Papadimitropoulos M, Tsekoura C, Graphou O,
Papadopoulou C, Gerondi M and Stavropoulos-Giokas C: Lack of the
appropriate natural killer cell inhibitory receptors in women with
spontaneous abortion. Hum Immunol. 66:65–71. 2005.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Clark DA, Chaouat G, Wong K, Gorczynski RM
and Kinsky R: Tolerance mechanisms in pregnancy: A reappraisal of
the role of class I paternal MHC antigens. Am J Reprod Immunol.
63:93–103. 2010.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Nowak I, Malinowski A, Tchorzewski H,
Barcz E, Wilczynski JR, Grybos M, Kurpisz M, Luszczek W, Banasik M,
Reszczynska-Slezak D, et al: Frequencies of killer
immunoglobulin-like receptor genotypes influence susceptibility to
spontaneous abortion. J Appl Genet. 50:391–398. 2009.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Yamada H, Shimada S, Kato EH, Morikawa M,
Iwabuchi K, Kishi R, Onoé K and Minakami H: Decrease in a specific
killer cell immunoglobulin-like receptor on peripheral natural
killer cells in women with recurrent spontaneous abortion of
unexplained etiology. Am J Reprod Immunol. 51:241–247.
2004.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Varla-Leftherioti M: The significance of
the women's repertoire of natural killer cell receptors in the
maintenance of pregnancy. Chem Immunol Allergy. 89:84–95.
2005.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Ntrivalas EI, Bowser CR, Kwak-Kim J,
Beaman KD and Gilman-Sachs A: Expression of killer
immunoglobulin-like receptors on peripheral blood NK cell subsets
of women with recurrent spontaneous abortions or implantation
failures. Am J Reprod Immunol. 53:215–221. 2005.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Akbari S, Shahsavar F, Karami R, Yari F,
Anbari K and Ahmadi SAY: Recurrent spontaneous abortion (RSA) and
maternal KIR genes: A comprehensive meta-analysis. JBRA Assist
Reprod. 24:197–213. 2020.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Hiby SE, Apps R, Chazara O, Farrell LE,
Magnus P, Trogstad L, Gjessing HK, Carrington M and Moffett A:
Maternal KIR in combination with paternal HLA-C2 regulate human
birth weight. J Immunol. 192:5069–5073. 2014.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Yang X and Meng T: MiR-215-5p decreases
migration and invasion of trophoblast cells through regulating CDC6
in preeclampsia. Cell Biochem Funct. 38:472–479. 2020.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Kelemu T, Erlandsson L, Seifu D, Hansson
E, Abebe M, Teklu S, Girma S, Traherne JA, Moffett A and Hansson
SR: Polymorphism in killer cell immunoglobulin-like receptors and
human leukocyte antigen-c and predisposition to preeclampsia in
Ethiopian pregnant women population. J Reprod Immunol.
141(103169)2020.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Huhn O, Chazara O, Ivarsson MA, Retière C,
Venkatesan TC, Norman PJ, Hilton HG, Jayaraman J, Traherne JA,
Trowsdale J, et al: High-resolution genetic and phenotypic analysis
of KIR2DL1 alleles and their association with pre-eclampsia. J
Immunol. 201:2593–2601. 2018.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Saito S, Takeda Y, Sakai M, Nakabayahi M
and Hayakawa S: The incidence of pre-eclampsia among couples
consisting of Japanese women and Caucasian men. J Reprod Immunol.
70:93–98. 2006.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Long W, Shi Z, Fan S, Liu L, Lu Y, Guo X,
Rong C, Cui X and Ding H: Association of maternal KIR and fetal
HLA-C genes with the risk of preeclampsia in the Chinese Han
population. Placenta. 36:433–437. 2015.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Gentle NL, Loubser S, Paximadis M, Puren A
and Tiemessen CT: Killer-cell immunoglobulin-like receptor (KIR)
and human leukocyte antigen (HLA) class I genetic diversity in four
South African populations. Hum Immunol. 78:503–509. 2017.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Higuma-Myojo S, Sasaki Y, Miyazaki S,
Sakai M, Siozaki A, Miwa N and Saito S: Cytokine profile of natural
killer cells in early human pregnancy. Am J Reprod Immunol.
54:21–29. 2005.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Nakimuli A, Chazara O, Byamugisha J,
Elliott AM, Kaleebu P, Mirembe F and Moffett A: Pregnancy,
parturition and preeclampsia in women of African ancestry. Am J
Obstet Gynecol. 210:510–520.e1. 2014.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Wallace AE, Whitley GS, Thilaganathan B
and Cartwright JE: Decidual natural killer cell receptor expression
is altered in pregnancies with impaired vascular remodeling and a
higher risk of pre-eclampsia. J Leukoc Biol. 97:79–86.
2015.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Larsen TG, Hackmon R, Geraghty DE and
Hviid TVF: Fetal human leukocyte antigen-C and maternal killer-cell
immunoglobulin-like receptors in cases of severe preeclampsia.
Placenta. 75:27–33. 2019.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Moffett A, Hiby SE and Sharkey AM: The
role of the maternal immune system in the regulation of human
birthweight. Philos Trans R Soc Lond B Biol Sci.
370(20140071)2015.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Clark MM, Chazara O, Sobel EM, Gjessing
HK, Magnus P, Moffett A and Sinsheimer JS: Human birth weight and
reproductive immunology: Testing for interactions between maternal
and offspring KIR and HLA-C genes. Hum Hered. 81:181–193.
2016.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Hilton HG, Vago L, Older Aguilar AM,
Moesta AK, Graef T, Abi-Rached L, Norman PJ, Guethlein LA,
Fleischhauer K and Parham P: Mutation at positively selected
positions in the binding site for HLA-C shows that KIR2DL1 is a
more refined but less adaptable NK cell receptor than KIR2DL3. J
Immunol. 189:1418–1430. 2012.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Madeja Z, Yadi H, Apps R, Boulenouar S,
Roper SJ, Gardner L, Moffett A, Colucci F and Hemberger M: Paternal
MHC expression on mouse trophoblast affects uterine vascularization
and fetal growth. Proc Natl Acad Sci USA. 108:4012–4017.
2011.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Kieckbusch J, Gaynor LM, Moffett A and
Colucci F: MHC-dependent inhibition of uterine NK cells impedes
fetal growth and decidual vascular remodelling. Nat Commun.
5(3359)2014.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Skjaerven R, Vatten LJ, Wilcox AJ, Rønning
T, Irgens LM and Lie RT: Recurrence of pre-eclampsia across
generations: Exploring fetal and maternal genetic components in a
population based cohort. BMJ. 331(877)2005.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Hiby SE, Ashrafian-Bonab M, Farrell L,
Single RM, Balloux F, Carrington M, Moffett A and Ebrahimi Z:
Distribution of killer cell immunoglobulin-like receptors (KIR) and
their HLA-C ligands in two Iranian populations. Immunogenetics.
62:65–73. 2010.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Vukcevic D, Traherne JA, Næss S,
Ellinghaus E, Kamatani Y, Dilthey A, Lathrop M, Karlsen TH, Franke
A, Moffatt M, et al: Imputation of KIR types from SNP variation
data. Am J Hum Genet. 97:593–607. 2015.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Vento-Tormo R, Efremova M, Botting RA,
Turco MY, Vento-Tormo M, Meyer KB, Park JE, Stephenson E, Polański
K, Goncalves A, et al: Single-cell reconstruction of the early
maternal-fetal interface in humans. Nature. 563:347–353.
2018.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Moffett A, Chazara O, Colucci F and
Johnson MH: Variation of maternal KIR and fetal HLA-C genes in
reproductive failure: Too early for clinical intervention. Reprod
Biomed Online. 33:763–769. 2016.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Chaouat G: Reconsidering the Medawar
paradigm placental viviparity existed for eons, even in
vertebrates; without a ‘problem’: Why are Tregs important for
preeclampsia in great apes? J Reprod Immunol. 114:48–57.
2016.PubMed/NCBI View Article : Google Scholar
|














