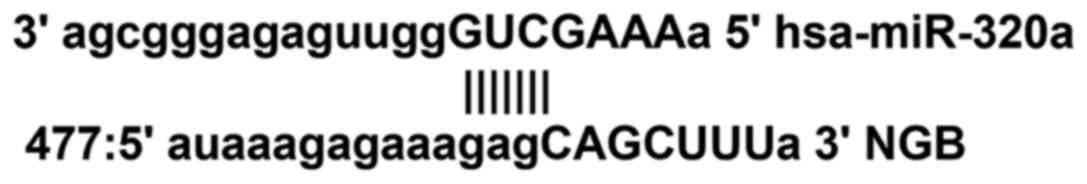Introduction
Post-operative cognitive dysfunction (POCD) has been
characterized by the progressive memory loss, personality changes
and/or post-operation cognitive function deterioration (1,2). The
incidence rate of POCD in patients receiving general anesthesia is
increased (3). The off-pump
coronary artery bypass grafting (OPCABG) completely avoids the
systemic inflammatory response caused by extracorporeal circulation
and reduces the blood transfusion rate, which, however, is still
arguable (4-6).
The intraoperative operation represents the key factor leading to
the instability of the hemodynamics. In some severe cases, the
sudden and violent fluctuations of the hemodynamics might lead to
POCD, together with the effects from anesthesia (7).
Neural damages might be one of the pathological
manifestations of POCD. Dexmedetomidine is a newly discovered
adrenergic receptor agonist, with high specificity to the
adrenergic α2 receptor. Dexmedetomidine is an auxiliary
anesthetic drug, which does not interfere with hemodynamics,
characterized by low respiratory inhibition effect and ease of
waking up after application. Dexmedetomidine would prevent the
propofol-induced hippocampal, thalamic and cortical damages
(8), significantly decreasing the
IL-6 and TNF-α levels, and increasing superoxide dismutase (SOD)
activity during general anesthesia (9). In addition, it has long-term effects
on the neurocognitive function, which could decrease the
inflammation and protect the neurological function, after traumatic
brain injuries (10), accompanied
by the high efficiency of pharmacokinetics, as well as the
anti-anxiety, anticonvulsant and anti-epilepsy effects (11).
The protective effects of neuroglobin (NGB) on brain
cells has already been confirmed. Under ischemia and hypoxia
stress, the survival ability of nerve cells would be significantly
increased by NGB overexpression, and the expression of associated
signal molecules involved in apoptosis response would also be
downregulated (12,13). In addition, NGB would eliminate
hypoxia-induced organelle aggregation (14), and improve the induction of actin
aggregation in mitochondrial transport (15). NGB is mainly distributed in the
anterior lobes and subthalamic nuclei in human beings and murine
models (16), which is specifically
expressed in neurons, while less expressed in glial cells (17).
Several studies have shown that multiple miRNAs play
important roles in the development of the nervous system, as well
as the functional and pathological processes, which has been shown
to be abnormally expressed during anesthesia (18-20),
including the miR-320a family (21). Moreover, the miR-320a family has
been predicted to be the upstream regulatory genes of NGB. The
miR-320a has been shown to be able to increase the neurite length,
while decreasing the total cell number, which has been considered
as a potential target for promoting the neuronal regeneration
following injuries (22). However,
the regulatory effect of miR-320a on NGB, and whether the gene
expression levels in the human body would be affected by
dexmedetomidine, have not been fully elucidated thus far.
Based on the aforementioned findings, the present
study was based on the hypothesis that the dexmedetomidine adjunct
anesthesia might protect the cognition function of patients
receiving OPCABG, involving the regulatory network of miR-320a and
NGB. Therefore, the present study investigated the role of
miR-320a/NGB in dexmedetomidine adjunct anesthesia, and the
expression levels of NGB and miR-320s in the blood samples of the
patients undergoing OPCABG with dexmedetomidine adjunct anesthesia
were detected and studied. Moreover, the association between the
NGB and miR320 was predicted and analyzed.
Materials and methods
Study subjects
In total, 40 patients, 12 males and 8 females, aged
33-60 years (with a median age of 40.8 years), who underwent the
OPCABG in the First Affiliated Hospital of Hebei North University,
between August 2014 and August 2018 and 20 normal subjects, 13
males and 7 females, aged 31-61 years (with a median age of 41.9
years), serving as a control, were included in the present study.
These patients were randomly divided into the following two groups
(n=20), based on a random number table: i) Dexmedetomidine group,
in which the patients were given dexmedetomidine before anesthesia;
and ii) control group, in which the patients were given saline
before anesthesia. The exclusion criteria were as follows: Those
with i) history of dementia and mental illness; ii) severe
cerebrovascular diseases; iii) multiple surgical history; iv)
history of severe infection in various systems during the
perioperative period; v) large blood loss and severe acid-base
balance and electrolyte imbalance in the perioperative period; vi)
diabetes; and vii) severely abnormal liver and/or kidney function.
Prior written and informed consent were obtained from every patient
and the study was approved by the ethics review board of the First
Affiliated Hospital of Hebei North University.
Anesthesia procedure
For both the dexmedetomidine and control groups, the
patients were routinely fasted before surgery. Particularly, the
off-pump surgery was performed, with 400 U/kg heparin and the ratio
of heparin to protamine of 1:1. The patients were subjected to the
venous access opening, oxygen mask, arterial blood pressure
monitoring, ECG, oxygen saturation monitoring and end-tidal carbon
dioxide monitoring. Patients from the dexmedetomidine group
underwent intravenous injection (by nurses, in a double-blinded
manner) of 0.6 µg/kg dexmedetomidine within 15 min prior to
anesthesia induction, and then pumped (0.5
µg·kg-1·h-1) until the end of surgery. On the
other hand, the patients from the control group were given with an
equal volume of 0.9% saline. For the intravenous induction, 2 mg
midazolam, 5 µg/kg sufentanil, 0.3 mg/kg etomidate and 0.3 mg/kg
cis-atracurium were given. Following the induction of anesthesia, a
small amount of propofol was continued via the intravenous
infusion, and anesthesia was maintained with the intermittent
intravenous injection of sufentanil and cis-atracurium. For the
anesthesia management, when the blood pressure fluctuated by
>20% of the baseline value, norepinephrine and nitroglycerin
would be supplemented. During surgery, intraoperative arterial
blood gas analysis was performed, and the electrolyte, blood
glucose, hematocrit, hemoglobin and lactic acid levels were
monitored. When the values were beyond the normal range, timely
intervention would be conducted to maintain the hemodynamic
stability, internal environment and electrolyte balance. When the
hemoglobin was <80 g/l, the infused red blood cells should be
administered. After the operation, the venous analgesia pump was
connected, and the patient was sent into the ICU.
Specimen collection
One the days before and after surgery, respectively,
5 ml blood sample was taken from the elbow vein before the blood
test. The blood sample was placed in a test tube for 1 h. The upper
serum in the tube was collected and placed in another centrifuge
tube. After centrifugation at 400 x g at 4˚C for 10 min, 1.5 ml
serum sample was placed in an EP tube, and stored at -80˚C until
further analysis.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted by TRIzol®
(Thermo Fisher Scientific, Inc.). The cDNA template was obtained by
reverse transcription with the miRcute miRNA cDNA first-strand
synthesis kit (Tiangen Biotech Co., Ltd.) and the TIANScriptII cDNA
first-strand synthesis kit (KR107; Tiangen Biotech Co., Ltd.),
according to the manufacturer's instructions. Quantitative PCR was
performed with the miRcute miRNA kit (cat. no. FP401; Tiangen
Biotech Co., Ltd.) on the PCR-iQ5 qRT-PCR machine (Bio-Rad
Laboratories, Inc.). The primer sequences was as follows: miR-320a
upstream, 5'-GCGCAAAAGCTGGGTTGAGA-3'; miR-320a downstream,
5'-CAGTGCAGGGTCCGAGGT-3'; U6 upstream,
5'-CTCGCTTCGGCAGCACATATACT-3'; U6 downstream,
5'-ACGCTTCACGAATTTGCGTGTC-3'; NGB upstream,
5'-AAGGTGATGCTCGTGATTGATG-3'; NGB downstream,
5'-AGGCTGGCAAGGTACTCCT-3'; GAPDH upstream,
5'-AGGAGCGAGACCCCACTAACAT-3' GAPDH downstream,
5'-GTGATGGCATGGACTGTGGT-3'. The 20-ml system consisted of 10 ml
RT-qPCR-mix, 0.5 ml primer each, 2 ml cDNA and 7 ml
ddH2O. The PCR condition was set as follows: 95˚C for 5
min; 95˚C for 20 sec, 58˚C for 30 sec, 72˚C for 30 sec, for a total
of 40 cycles. The expression levels of target genes were calculated
with the 2-ΔΔCq method (23). U6 and GAPDH were used as internal
control.
Enzyme-linked immunosorbent assay
(ELISA)
Blood sample was centrifuged at 1,000 x g at 4˚C for
10 min. ELISA was performed according to the manufacturer's
instructions. The standard and sample wells were set on the human
NGB ELISA kit (cat. no. FK-QZ1868; Fanke). In total, 50 µl
standards at indicated concentrations were added into each standard
well, while 10 µl testing sample was added into the test well
(diluted with 40 µl diluting solution). Nothing was added to the
blank well. Except for the blank wells, 100 µl horseradish
peroxidase (HRP)-labeled detection antibody was added into each
standard well and sample well, which were sealed with membrane and
incubated for 1 h. After washing, substrates A and B (50 µl each)
were added into each well, which were incubated at 37˚C for 15 min.
Then, 50 µl stop solution was added into each well. The OD value at
450 nm was measured with a microplate reader within 15 min.
Western blot analysis
Total protein was extracted with the P0013B RIPA
lysis buffer, according to the manufacturer's instructions. The
protein concentration was determined with the BCA method (cat. no.
RTP7102; Real-times). In total, 20 mg protein sample was separated
by 10% SDS-PAGE, which was then electronically transferred onto a
PVDF membrane. After blocking with 5% non-fat milk at room
temperature for 1 h, the membrane was incubated with rabbit
anti-human anti-NGB primary antibody (1:1,500 dilution; cat. no.
sc-30144; Santa Cruz Biotechnology, Inc.), or rabbit anti-human
anti-β-actin primary antibody (1:3,000 dilution; cat. no. ab129348;
Abcam), at 4˚C overnight. Then, the membrane was incubated with the
goat anti-rabbit secondary antibody (1:3,000 dilution; cat. no.
ab6721; Abcam) at room temperature at 1 h. Color development was
performed with the ECL method (cat. no. ab65623; Abcam). Protein
bands were imaged and analyzed with the Image Lab 3.0 software
(Bio-Rad Laboratories, Inc.). β-actin was used as the internal
reference.
Bioinformatics prediction
Bioinformatics prediction is currently the basis and
main clue for the functional investigation of miRNA. Herein, the
miRanda target gene prediction software (http://www.microrna.org/microrna/home.do) was used to
search for the target genes that miR-320a might regulate.
Cell culture
The 293T cells and CATH.a cells were purchased from
The Cell Bank of Type Culture Collection of The Chinese Academy of
Sciences. These cells were cultured in RPMI-1640 medium (cat. no.
11875127; Gibco; Thermo Fisher Scientific, Inc.), containing 10%
FBS, in a 5% CO2, 37˚C incubator. The human serum for
cell culture was inactivated at 56˚C for 30 min, and filtered
through 0.22-µm micropore, which was used to prepare the RPMI-1640
medium containing 10% serum. For the cells induced by
dexmedetomidine (1 and 10 mM, respectively; Jiangsu Hengrui
Medicine Co. Ltd), the RPMI-1640 medium containing 10% FBS was used
for cell culture. At 24 h after induction, the protein indexes were
detected. For the co-incubation with NGB recombinant protein (cat.
no. ab63278; Abcam), the cells were stimulated with recombinant
protein (1 mg/l) for 48 h.
Cell transfection
Cells in the logarithmic growth phase were
inoculated onto 24-well plates, at the density of 3x105
cells/well. The cells were cultured with the F12/DMEM medium
(Thermo Fisher Scientific, Inc.), containing 10% FBS, without
antibiotics. When 70% confluence was reached, cell transfection was
performed. The transfection plasmid/siRNA/agomiR (100 nM), together
with 1 ml Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), was added to an EP tube containing 50 ml
OptiMemi medium (Gibco; Thermo Fisher Scientific, Inc.). After
placing at room temperature for 20 min, the mixture was added onto
the well to incubate with cells for 6 h. Then, the culture medium
was replaced with fresh F12/DMEM medium containing 10% FBS. After
48 h, the mRNA and protein samples were extracted and detected.
Dual-luciferase reporter assay
The wild-type and mutant seed regions for miR-320a
in the 3'-untranslated region (UTR) of the NGB gene were chemically
synthesized by Sangon Biotech Co., Ltd., with the Spe-1 and
HindIII restriction sites added to both ends. These two DNA
fragments were then cloned into the pMIR-REPORT luciferase reporter
plasmid (Ambion; Thermo Fisher Scientific, Inc.), and the mutant
3'-UTR seed region was used as the control. In total, 0.8 mg
plasmids, containing the wild-type and mutant 3'-UTR DNA sequences,
were transfected with Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) into the 293T cells, with the liposome method.
Then, the cells were treated with the agomiR-320a (100 nM) for 24
h. The cells were lysed, and the luciferase was detected with the
GloMax 20/20 luminometer (Promega Corporation). Renilla was
used as internal reference.
MTT assay
After transfection, the cells were seeded onto the
96-well plate, at a density of 2x103 cells/well. The
experiment was performed in triplicates. At 24, 48 and 72 h,
respectively, 20 µl MTT (5 g/l) was added to incubate with cells at
37˚C for 4 h. Then, 150 µl DMSO was added into each well. The
absorption at 490 nm was determined, and the cell proliferation
curve was plotted accordingly.
Statistical analysis
Data are expressed as mean ± SD. Statistical
analysis was performed with the SPSS 18.0 software (SPSS, Inc.).
One-way ANOVA was used for comparison of multiple sets of
measurement data, with the Dunnett's T3 test; except for data in
Figs. 1 and 4, which were analyzed by mixed two-way
analysis of variance, and then subjected to Bonferroni analysis.
The Student's t-test was used for the comparison between two groups
of measurement data. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of NGB in serum
Among the recruited subjects, in the dexmedetomidine
group, there were 12 males and 8 females, aged 33-60 years old,
with the median age of 40.8 years old. In the control group, there
were 13 males and 7 females, aged 31-61 years old, with the median
age of 41.9 years old (Table SI).
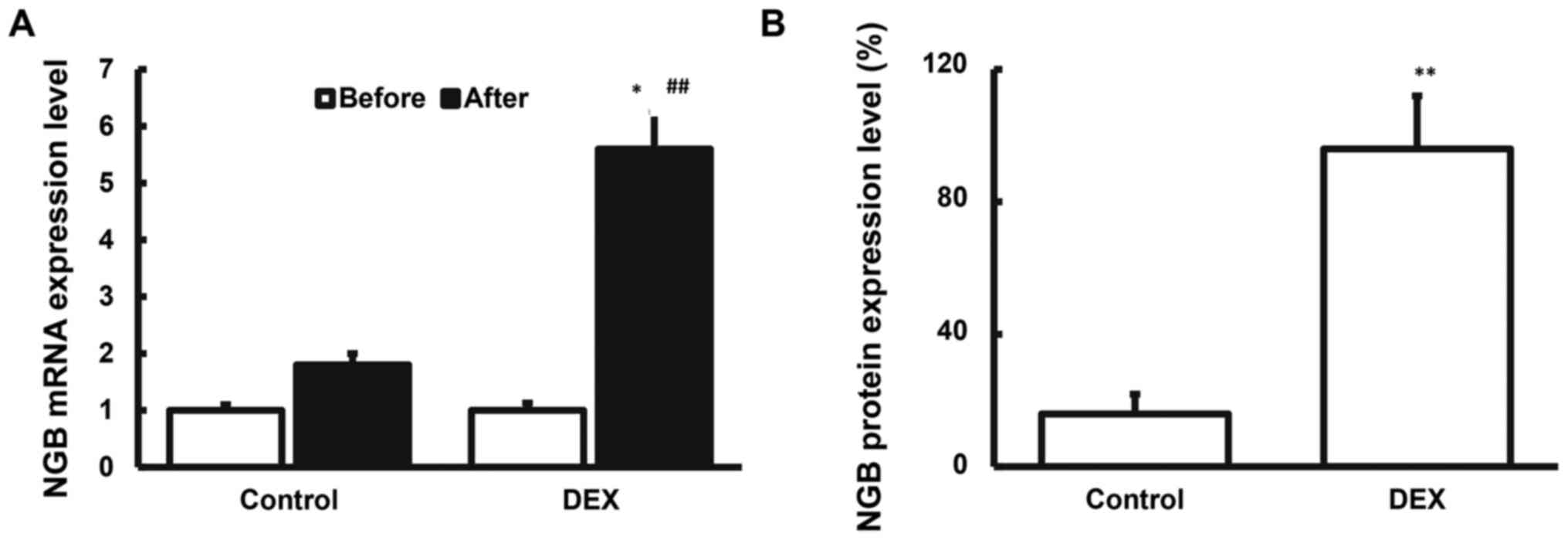
To investigate the expression levels of NGB in the serum from the
patients before and after surgery, RT-qPCR and ELISA were
performed. The results showed that both the mRNA and protein
expression levels of NGB were elevated in the patients after
surgery. Moreover, the elevated amplitude of NGB were greater in
the dexmedetomidine group (Fig. 1),
indicating that the addition of dexmedetomidine during anesthesia
may induce the release of NGB in the blood.
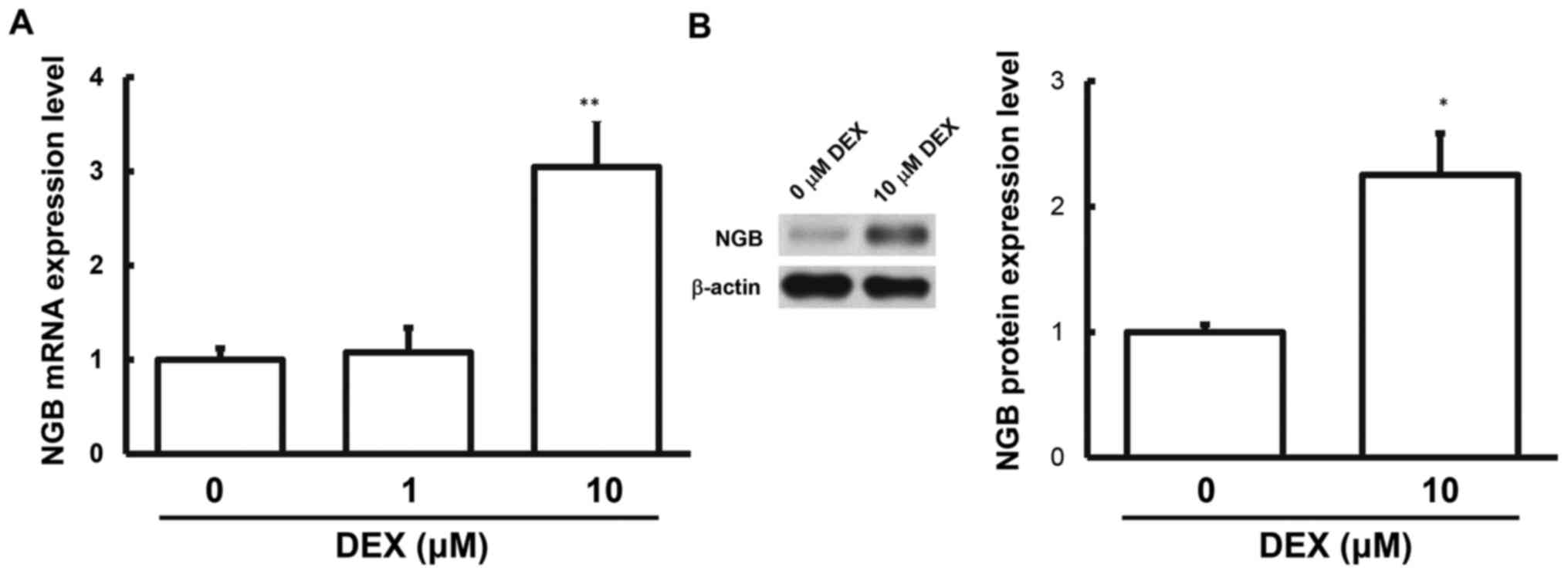
NGB expression after dexmedetomidine
induction
The human neuronal CATH.a cells were directly
induced with dexmedetomidine, and the mRNA and protein expression
levels of NGB were then detected. The results showed that, for the
induction with 1 µM dexmedetomidine, the mRNA expression levels of
NGB in the cells was not directly upregulated compared with the
control group. However, the induction with 10 µM dexmedetomidine
significantly elevated the expression levels of NGB in the CATH.a
cells. Moreover, the results showed that the protein expression
levels of NGB in the cells were significantly upregulated (Fig. 2). Together with the results from the
detection of human blood samples in clinic, the aforementioned
findings suggest that dexmedetomidine (especially at 10 µM) can
induce the NGB expression in the neuronal cells.
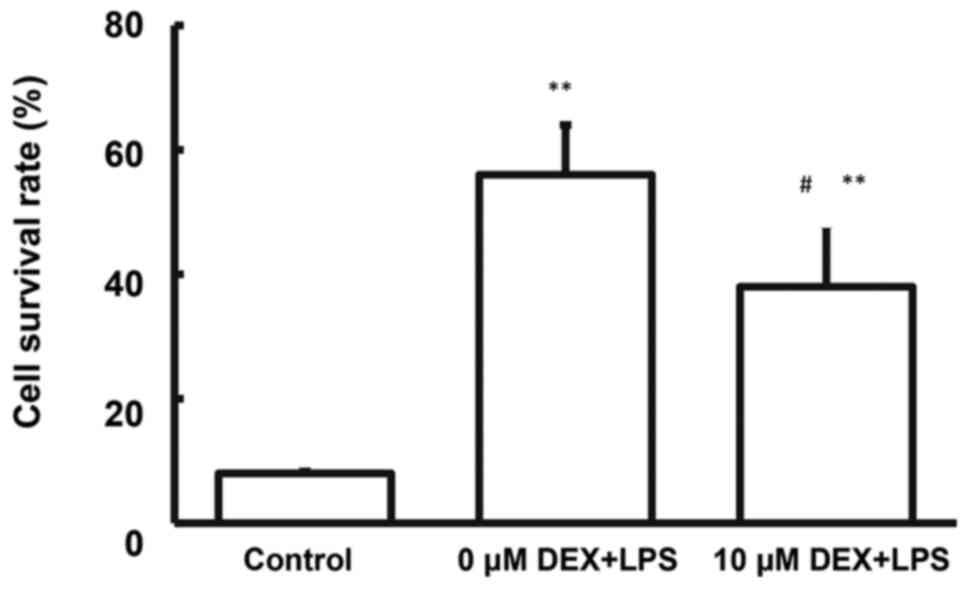
Survival of NGB-pre-treated cells with
LPS stimulation
The CATH.a cells was directly incubated with NGB
recombinant protein, which were then subjected to LPS stimulation
to simulate the survival stress. The results showed that
pretreatment with NGB significantly improved the survival of CATH.a
cells under LPS stress (Fig. 3).
These results suggest that NGB could exert neurologically
protective effects, especially for the external stress.
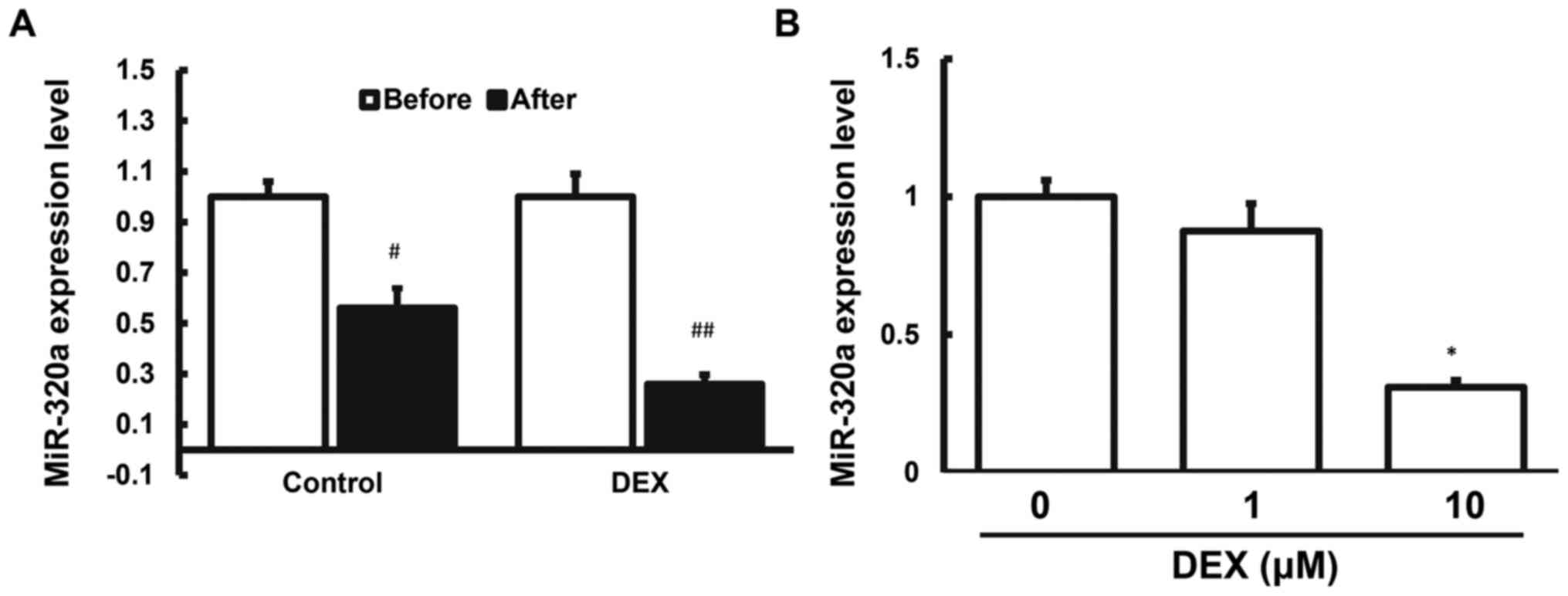
Expression levels of miR-320a in serum
samples and in cells induced by dexmedetomidine
The miR-320a has been a predicted potential upstream
regulating gene of NGB. Herein, the expression levels of miR-320a
in the serum samples from patients and in the CATH.a cells induced
by dexmedetomidine were detected with RT-qPCR. The results showed
that, in contrast to the elevated NGB expression, the expression
level of miR-320a was significantly decreased (Fig. 4). Overall, the results suggest that
these two factors (miR-320a and NGB) would have reverse regulatory
patterns.
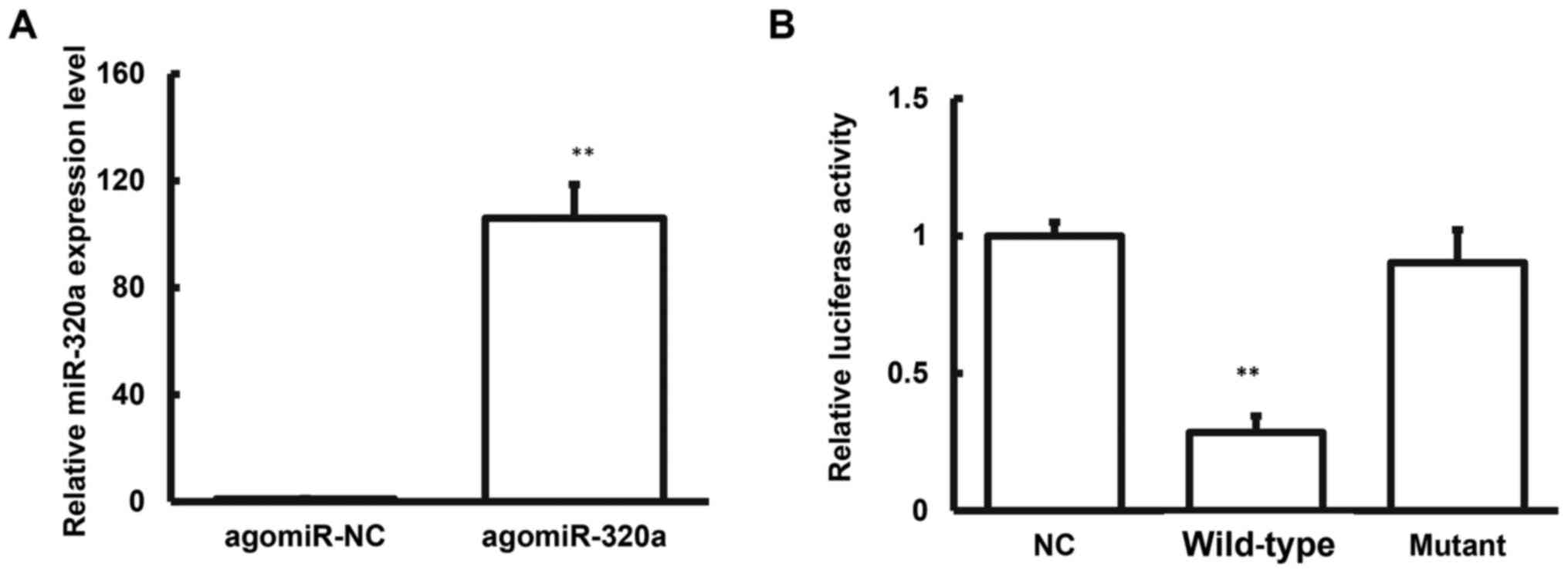
Dual-luciferase reporter assay
Based on the bioinformatics analysis with the
miRanda software, the NGB was predicted as a potential target genes
for miR-320a (Fig. 5). The results
from the dual luciferase reporter assay showed that compared with
the control group, the luciferase activity was significantly
decreased after co-transfection with the agomiR-320a and
pMIR-REPORT luciferase reporter plasmids (P<0.05), while no
significant difference was observed between the control and mutant
groups (P>0.05) (Fig. 6). The
results suggest that miR-320a could bind to the 3'-UTR of NGB to
regulate the gene expression.
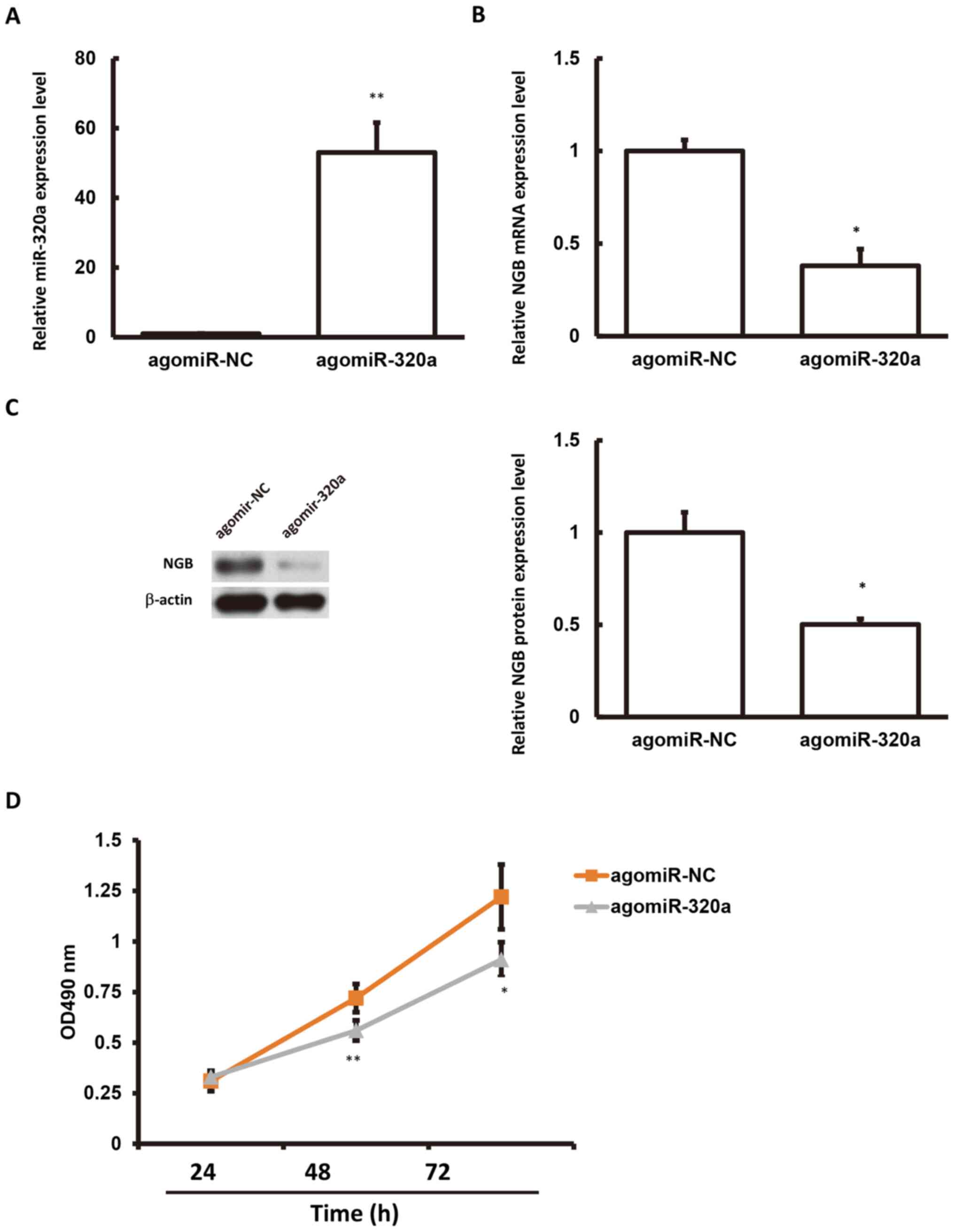
miR-320a regulates NGB expression and
affects neuronal cell proliferation
Next, the agomiR-320a and agomiR-NC were transfected
into CATH.a cells, respectively. The results showed that the
miR-320a expression levels were significantly elevated in these
cells. Moreover, the mRNA and protein expression levels of NGB in
the cells were significantly decreased. Meanwhile, the
proliferative activity of cells was detected by MTT assay. The
proliferative activity of neurons overexpressing miR-320a was
significantly decreased (Fig. 7).
The aforementioned results suggest that agomiR-320a may affect the
neuronal cell proliferation through NGB.
Discussion
POCD is a type of injury of the central nervous
system, for which the age has been found as a major risk factor for
POCD (24). Moreover, the surgery
type has also been shown to represent an important factor in the
development of POCD in elderly patients, especially for those
undergoing major operation (25).
In addition, POCD may also be associated with the surgery type,
hypoxia, intraoperative hypotension, stress, alcohol consumption,
medication history, cognitive function and other factors (26). Until now, the specific pathogenesis
and etiology of POCD have not yet been elucidated. The OPCABG
completely avoids the systemic inflammatory responses caused by
extracorporeal circulation, which however, needs high level of
surgical performance; otherwise it may lead to extremely unstable
hemodynamics. Therefore, clinical anesthesia needs to protect the
central nervous system and inhibit the inflammatory responses, so
as to decrease the incidence of POCD. In the present study, the
results showed that pre-administration of dexmedetomidine could
decrease the expression levels of miR-320 in the blood samples of
patients undergoing OPCABG, which also stimulated the high
expression of cerebral hemoglobin and increased the proliferation
activity of neuronal cells. The negative association between the
miR-320 and NGB was also preliminarily discussed.
Dexmedetomidine is one of the most widely studied
anesthesia adjuvants in recent years (27). Dexmedetomidine has mainly been used
to regulate the neurotransmitters through acting on locus nucleus
in the brain stem of patients. The application of dexmedetomidine
in the cardiac surgery would lead to safe and effective outcomes,
thus improving the patient's condition. Dexmedetomidine can
selectively excite the α2 receptor of the presynaptic
membrane, to stimulate the negative feedback effects of sympathetic
nerves, thus achieving sedation and inhibiting anxiety and finally
decreasing the anesthesia medication (11). It has been shown that
dexmedetomidine has a satisfactory effect on decreasing the
inflammatory mediators after surgery, and exerts brain-protecting
effects in the craniotomy of acute craniocerebral injury (10). Therefore, it was speculated that
POCD might have protective and therapeutic effects against
inflammatory responses. Moreover, the myocardial protective effects
of dexmedetomidine on patients undergoing the OPCABG have been
confirmed (28). Currently, it has
been shown that dexmedetomidine would alleviate the postoperative
cognitive dysfunction in elderly patients. However, the impacts of
dexmedetomidine on patients undergoing OPCABG have not yet been
reported.
NGB is expressed in the nervous cells of the
mammalian nervous systems, as well as in the retina of most mammals
(29-32).
Overexpressed NGB can lower the neuronal damages under hypoxic
conditions and local cerebral ischemia (33,34).
The infarct size and oxidative stress have been shown to be
significantly decreased in the tissues after stroke in the
transgenic mice overexpressing NGB (35). In recent years, NGB has been
reported to have a certain role in scavenging ROS. It has been well
established that the anti-apoptotic effects of NGB are mainly
through the endogenous pathways (36,37).
NGB could restore the CytoC, which triggers the apoptotic cascade
only in the oxidative state, thereby exerting protective effects.
NGB works as a possible endogenous neuroprotective agent in the
nervous system. At present, however, there is no relevant study on
the effect of dexmedetomidine on the expression of NGB. The present
study found that the expression level of NGB in the blood of
patients given dexmedetomidine adjunct anesthesia was significantly
elevated, suggesting that the elevated NGB expression may be caused
by the drug in the brain cells of the patient during OPCABG, which
might be one of the underlying protective mechanisms. When the
neuronal cells were directly induced by dexmedetomidine, the
expression levels of NGB in the cells were also significantly
elevated. Pre-incubation of CATH.a cells with NGB antibody
significantly increased the survival rate of the cells stimulated
with LPS, further confirming the protective effects of NGB on brain
cells. However, the specific regulatory mechanism of
dexmedetomidine on NGB has not been fully clarified.
The miRNAs usually play biological roles by cleaving
mRNAs. The bioinformatics prediction showed that NGB might be a
potential target gene for miR-320a. Several studies have reported
the role of NGB in the apoptotic pathway. It has been shown that
miR-320a is downregulated in various tumor diseases, such as the
prostate cancer, glioma, colon cancer and leukemia (38-40).
The downregulated expression levels of miR-320a in the liver cancer
can promote the invasion of tumor cells and lead to poor prognosis
(41). These findings suggest that
miR-320a is closely associated with cell proliferation. Therefore,
it was speculated that dexmedetomidine might affect the NGB
expression levels by regulating miR-320a. The results showed that
the expression levels of miR-320a in the blood samples of patients
given dexmedetomidine adjunct anesthesia was significantly lower
compared with the control group, while the dexmedetomidine
induction downregulated the expression levels of miR-320a in the
CATH.a cells. The present study results from the dual luciferase
reporter assay showed the direct binding between miR-320a and NGB.
The cell proliferation activity assay also confirmed that the
miR-320a overexpression can lower the proliferation activity of the
CATH.a cell line. The aforementioned results suggest that the
expression of miR-320a is decreased by dexmedetomidine, which is
key to increase the NGB expression levels. However, the limited
subjects might be one of the limitations of the present study, and
further in-depth studies are still needed with enlarged subject
sizes. Moreover, the lack of neuropsychological assessment and
analysis of direct interactions between miR-320/NGB and POCD in the
cohort might be another limitation of the present study.
In conclusion, the present results showed that
pre-administration of dexmedetomidine could increase the expression
of NGB in the blood of patients undergoing OPCABG, protecting the
brain cells. The possible mechanism might be that dexmedetomidine
decreases the miR-320a expression and thus, increases the
proliferative activity of neuronal cells.
Supplementary Material
Basic patients' information.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by the Medical Science Research
Project of the Health Committee of Hebei Province in 2019 (grant
no. 20190890).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YG designed the study, performed the experiments and
was a major contributor in writing the manuscript. XZ performed the
experiments and drafted the manuscript. HY provided the materials
and collected the data. WW and YW analyzed and interpreted the
data. FL provided the materials, interpreted the data and revised
the manuscript. JT designed the study and provided administrative
support. YG and JT confirm the authenticity of all the raw data.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the ethics review board of
the First Affiliated Hospital of Hebei North University. All
patients provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Borozdina A, Qeva E, Cinicola M and
Bilotta F: Perioperative cognitive evaluation. Curr Opin
Anaesthesiol. 31:756–761. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Evered LA and Silbert BS: Postoperative
cognitive dysfunction and noncardiac surgery. Anesth Analg.
127:496–505. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Needham MJ, Webb CE and Bryden DC:
Postoperative cognitive dysfunction and dementia: What we need to
know and do. Br J Anaesth. 119 (Suppl 1):i115–i125. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Niu X, Zhang Q, Xiao D and Zhang Y: A
retrospective study of hemodynamic changes in patients after
off-pump coronary artery bypass graft surgery using impedance
cardiography. Med Sci Monit. 25:3454–3462. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Glumac S, Kardum G, Sodic L, Supe-Domic D
and Karanovic N: Effects of dexamethasone on early cognitive
decline after cardiac surgery: A randomised controlled trial. Eur J
Anaesthesiol. 34:776–784. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Glumac S, Kardum G and Karanovic N:
Postoperative cognitive decline after cardiac surgery: A narrative
review of current knowledge in 2019. Med Sci Monit. 25:3262–3270.
2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Liu YH, Wang DX, Li LH, Wu XM, Shan GJ, Su
Y, Li J, Yu QJ, Shi CX, Huang YN and Sun W: The effects of
cardiopulmonary bypass on the number of cerebral microemboli and
the incidence of cognitive dysfunction after coronary artery bypass
graft surgery. Anesth Analg. 109:1013–1022. 2009.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Sanders RD, Xu J, Shu Y, Januszewski A,
Halder S, Fidalgo A, Sun P, Hossain M, Ma D and Maze M:
Dexmedetomidine attenuates isoflurane-induced neurocognitive
impairment in neonatal rats. Anesthesiology. 110:1077–1085.
2009.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Han C, Fu R and Lei W: Beneficial effects
of dexmedetomidine on early postoperative cognitive dysfunction in
pediatric patients with tonsillectomy. Exp Ther Med. 16:420–426.
2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Ren X, Ma H and Zuo Z: Dexmedetomidine
postconditioning reduces brain injury after brain hypoxia-ischemia
in neonatal rats. J Neuroimmune Pharmacol. 11:238–247.
2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Barends CR, Absalom A, van Minnen B,
Vissink A and Visser A: Dexmedetomidine versus midazolam in
procedural sedation. A systematic review of efficacy and safety.
PLoS One. 12(e0169525)2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Chan AS, Saraswathy S, Rehak M, Ueki M and
Rao NA: Neuroglobin protection in retinal ischemia. Invest
Ophthalmol Vis Sci. 53:704–711. 2012.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Ye SQ, Zhou XY, Lai XJ, Zheng L and Chen
XQ: Silencing neuroglobin enhances neuronal vulnerability to
oxidative injury by down-regulating 14-3-3gamma. Acta Pharmacol
Sin. 30:913–918. 2009.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Liu X, Gao Y, Yao H, Zhou L, Sun D and
Wang J: Neuroglobin involvement in the course of arsenic toxicity
in rat cerebellar granule neurons. Biol Trace Elem Res.
155:439–446. 2013.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Khan AA, Mao XO, Banwait S,
DerMardirossian CM, Bokoch GM, Jin K and Greenberg DA: Regulation
of hypoxic neuronal death signaling by neuroglobin. FASEB.
22:1737–1747. 2008.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Uchida Y, Ohshima T, Sasaki Y, Suzuki H,
Yanai S, Yamashita N, Nakamura F, Takei K, Ihara Y, Mikoshiba K, et
al: Semaphorin3A signalling is mediated via sequential Cdk5 and
GSK3beta phosphorylation of CRMP2: Implication of common
phosphorylating mechanism underlying axon guidance and Alzheimer's
disease. Genes Cells. 10:165–179. 2005.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhang C, Wang C, Deng M, Li L, Wang H, Fan
M, Xu W, Meng F, Qian L and He F: Full-length cDNA cloning of human
neuroglobin and tissue expression of rat neuroglobin. Biochem
Biophys Res Commun. 290:1411–1419. 2002.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Bukauskas T, Kairyte M, Mickus R,
Puleikyte L and Macas A: Values of circulating molecular biomarkers
(microRNAs) for the evaluation of renal failure during urgent
abdominal sepsis anaesthesia. Acta Med Litu. 26:17–24.
2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Bell JD, Cho JE and Giffard RG: MicroRNA
changes in preconditioning-induced neuroprotection. Transl Stroke
Res. 8:585–596. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Barik M, Kumar A, Mishra PR and Kapoor PM:
Role of MicroRNA in cardiac anesthesia: An innovative consequences
and new possibility. Ann Card Anaesth. 20:274–275. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Tadano T, Kakuta Y, Hamada S, Shimodaira
Y, Kuroha M, Kawakami Y, Kimura T, Shiga H, Endo K, Masamune A, et
al: MicroRNA-320 family is downregulated in colorectal adenoma and
affects tumor proliferation by targeting CDK6. World J Gastrointest
Oncol. 8:532–542. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
White RE and Giffard RG: MicroRNA-320
induces neurite outgrowth by targeting ARPP-19. Neuroreport.
23:590–595. 2012.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Berger M, Nadler JW, Browndyke J, Terrando
N, Ponnusamy V, Cohen HJ, Whitson HE and Mathew JP: Postoperative
cognitive dysfunction: Minding the gaps in our knowledge of a
common postoperative complication in the elderly. Anesthesiol Clin.
33:517–550. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Feinkohl I, Winterer G, Spies CD and
Pischon T: Cognitive reserve and the risk of postoperative
cognitive dysfunction. Dtsch Arztebl Int. 114:110–117.
2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Safavynia SA and Goldstein PA: The role of
neuroinflammation in postoperative cognitive dysfunction: Moving
from hypothesis to treatment. Front Psychiatry.
9(752)2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Sottas CE and Anderson BJ:
Dexmedetomidine: The new all-in-one drug in paediatric anaesthesia?
Curr Opin Anaesthesiol. 30:441–451. 2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chi X, Liao M, Chen X, Zhao Y, Yang L, Luo
A and Yang H: Dexmedetomidine attenuates myocardial injury in
off-pump coronary artery bypass graft surgery. J Cardiothorac Vasc
Anesth. 30:44–50. 2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Wystub S, Laufs T, Schmidt M, Burmester T,
Maas U, Saaler-Reinhardt S, Hankeln T and Reuss S: Localization of
neuroglobin protein in the mouse brain. Neurosci Lett. 346:114–116.
2003.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Reuss S, Saaler-Reinhardt S, Weich B,
Wystub S, Reuss MH, Burmester T and Hankeln T: Expression analysis
of NGB mRNA in rodent tissues. Neuroscience. 115:645–656.
2002.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Schmidt M, Giessl A, Laufs T, Hankeln T,
Wolfrum U and Burmester T: How does the eye breathe? Evidence for
neuroglobin-mediated oxygen supply in the mammalian retina. J Biol
Chem. 278:1932–1935. 2003.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Sun Y, Jin K, Mao XO, Zhu Y and Greenberg
DA: NGB is up-regulated by and protects neurons from
hypoxic-ischemic injury. Proc Natl Acad Sci USA. 98:15306–15311.
2001.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Khan AA, Sun Y, Jin K, Mao XO, Chen S,
Ellerby LM and Greenberg DA: A NGB-overexpressing transgenic mouse.
Gene. 398:172–176. 2007.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Khan AA, Wang Y, Sun Y, Mao XO, Xie L,
Miles E, Graboski J, Chen S, Ellerby LM, Jin K and Greenberg DA:
Neuroglobin-overexpressing transgenic mice are resistant to
cerebral and myocardial ischemia. Proc Natl Acad Sci USA.
103:17944–17948. 2006.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Wang X, Liu J, Zhu H, Tejima E, Tsuji K,
Murata Y, Atochin DN, Huang PL, Zhang C and Lo EH: Effects of
neuroglobin overexpression on acute brain injury and long-term
outcomes after focal cerebral ischemia. Stroke. 39:1869–1874.
2008.PubMed/NCBI View Article : Google Scholar
|
|
36
|
De Marinis E, Acaz-Fonseca E, Arevalo MA,
Ascenzi P, Fiocchetti M, Marino M and Garcia-Segura LM:
17β-Oestradiol anti-inflammatory effects in primary astrocytes
require oestrogen receptor β-mediated neuroglobin up-regulation. J
Neuroendocrinol. 25:260–270. 2013.PubMed/NCBI View Article : Google Scholar
|
|
37
|
De Marinis E, Marino M and Ascenzi P:
Neuroglobin, estrogens, and neuroprotection. IUBMB Life.
63:140–145. 2011.PubMed/NCBI View
Article : Google Scholar
|
|
38
|
Sun JY, Huang Y, Li JP, Zhang X, Wang L,
Meng YL, Yan B, Bian YQ, Zhao J, Wang WZ, et al: MicroRNA-320a
suppresses human colon cancer cell proliferation by directly
targeting beta-catenin. Biochem Biophys Res Commun. 420:787–792.
2012.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhang Y, He X, Liu Y, Ye Y, Zhang H, He P,
Zhang Q, Dong L, Liu Y and Dong J: microRNA-320a inhibits tumor
invasion by targeting neuropilin 1 and is associated with liver
metastasis in colorectal cancer. Oncol Rep. 27:685–694.
2012.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Gerber JM, Qin L, Kowalski J, Smith BD,
Griffin CA, Vala MS, Collector MI, Perkins B, Zahurak M, Matsui W,
et al: Characterization of chronic myeloid leukemia stem cells. Am
J Hematol. 86:31–37. 2011.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Lu C, Liao Z, Cai M and Zhang G:
MicroRNA-320a downregulation mediates human liver cancer cell
proliferation through the Wnt/β-catenin signaling pathway. Oncol
Lett. 13:573–578. 2017.PubMed/NCBI View Article : Google Scholar
|