Introduction
I/R injury is a pivotal etiological factor for
hepatic dysfunction, and graft failure during liver transplantation
and resection (1-3).
It has previously been reported that the mechanism of hepatic I/R
injury includes necrocytosis, metabolic disturbances and the immune
response of inflammation; however, the complete molecular pathology
of hepatic I/R injury is yet to be elucidated (4). Kupffer cells (KCs) are identified as
liver-settled macrophages and are responsible for identifying and
removing exogenous particles and immunoreactive materials that are
considered harmful to the human body (5). When activated by liver I/R treatment,
KCs adjust the outcome of the immune response in the liver via the
regulation of inflammatory signaling pathways (4,5).
The NLRP3 inflammasome, which consists of NLRP3 (a
transducer), ASC (an adapter) and caspase-1 (an effector), has been
identified as a multiprotein complex (6,7). The
stimulation of the NLRP3 inflammasome relies on a special ignition
system requiring two necessary processes: Priming and activation.
The priming step involves stimulating the synthesis of both NLRP3
and pro-interleukin (IL)-1β, which results in a decrease in the
threshold of NLRP3 inflammasome activation. The second step,
activation, is a poorly characterized event, during which the NLRP3
inflammasome is packaged so as to cleave pro-caspase-1 to mature
caspase-1. Once triggered, pro-IL-18 and pro-IL-1β are cleaved by
mature caspase-1 to produce functional inflammatory cytokines,
namely IL-18 and IL-1β (6,8). Uncontrollable NLRP3 inflammasome is
associated with several diseases, such as atherosclerosis (9), sepsis (10), Crohn's disease (11), acute myocardial infarction (12) and cancer (13). Furthermore, recent research has
revealed that the NLRP3 pathway is contained in the pathological
development of hepatic I/R injury (14-16).
CMPK2, a pyrimidine nucleoside monophosphate kinase,
was reported to be in connection with the generation of
mitochondrial DNA (mtDNA) (17).
CMPK2 is stimulated by poly(I:C) and LPS, indicating that CMPK2 has
a potent antiviral and antibacterial influence (18-20).
Certain other studies have reported the association between CMPK2
and several diseases, such as multiple sclerosis (21), human immunodeficiency virus
(22) and Porcine Reproductive and
Respiratory Syndrome Virus (23).
In addition, CMPK2 has been reported to be involved in the terminal
differentiation of monocytes/macrophages (24), macrophage activation and the
inflammatory response (17). The
mechanism by which CMPK2 plays a part in hepatic I/R injury is yet
to be fully elucidated. Therefore, the aim of the present study was
to investigate whether CMPK2 affects hepatic I/R injury and to
determine the underlying mechanism.
Materials and methods
Animals
Healthy C57BL/6J male mice (n=43; weight, 25-28 g;
age, 6-8 weeks) were purchased from Chongqing Medical University
Laboratory Animals Center. All animal experiments were approved by
the Animal Care and Use Committee of Chongqing Medical University.
Animal health and behavior was monitored twice a day before
operation and once hourly after the operation. All mice used in
this study were cared for in strict accordance with the Guide for
the Care and Use of Laboratory Animals (25). All mice were raised in a 12/12-h
light/dark cycle with 23±3˚C and 60-65% relative humidity and were
given a normal diet with free access to food and water.
Hepatic I/R model
A liver I/R model with 70% ischemia was constructed
as previously described (26).
Ischemia and reperfusion were performed for 1 and 6 h,
respectively. Pentobarbital sodium (intraperitoneal injection, 45
mg/kg) was used to anesthetize mice before the laparotomy was
performed. The artery and portal vein were occluded to produce the
70% hepatic ischemia for 1 h. Subsequently, the clamps were removed
to perform reperfusion for 6 h. Mice in the sham group underwent an
identical surgery but without liver vascular clamping. All mice
were euthanized after the operation according to the Guide for the
Care and Use of Laboratory Animals. The method of euthanasia used
was cervical dislocation. In this study, 40 mice were euthanized
after the experiment, 2 mice died from pneumothorax and 1 mouse
died following anesthesia. According to AVMA Guidelines for the
Euthanasia of Animals and the Guide for the Care and Use of
Laboratory Animals (25), mice
death was confirmed by recognizing the cessation of vital signs in
the mice being euthanized.
Cell culture
RAW 264.7 cells (a murine macrophage cell line) were
purchased from Cell Bank of the Cell Bank of Type Culture
Collection of the Chinese Academy of Sciences. All cells were
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.) mixed with
10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.).
Cells were cultured in the cell incubator (Thermo Fisher
Scientific, Inc.) at 37˚C with 95% air and 5% CO2.
Cell H/R model
A RAW 264.7 cell H/R model was constructed as
previously described (27). Hypoxia
was induced for 6 h by culturing cells in a 37˚C incubator (Thermo
Fisher Scientific, Inc.) with O2 (1%), CO2
(5%) and N2 (94%). The cells were then placed in a 37˚C
incubator with CO2 (5%) and air (95%) for varying
reoxygenation times (1, 6, 12 and 24 h).
Reverse transcription-quantitative PCR
(RT-qPCR) assay
Following the RNAiso Plus (cat. no. 9108; Takara
Bio, Inc.) protocol, total RNA was harvested from cells. Reverse
transcription was performed by PrimeScript™ RT reagent Kit (Takara
Bio, Inc.) according to the manufacturer's instructions. SYBR™
Green PCR kit (cat. no. 4309155; Thermo Fisher Scientific, Inc.)
was used to detect the relative mRNA expression of CMPK2, NLRP3,
IL-1β, IL-18 and TNF-α. Thermocycling conditions were as follows:
Initial denaturation at 95˚C for 10 min; followed by 40 cycles of
95˚C for 15 sec and 60˚C for 60 sec. Fold changes were calculated
using the 2-ΔΔCq method (28). mRNA expression data were
standardized to β-actin. The primer sequences were as follows:
CMPK2 forward, 5'-GGCAATTATCTCGTGGCTTC-3' and reverse,
5'-GTAGCTATGGCGTAGGTGGC-3'; NLRP3 forward,
5'-GAGTTCTTCGCTGCTATGT-3' and reverse, 5'-GAGTTCTTCGCTGCTATGT-3';
IL-1β forward, 5'-AGTTGACGGACCCCAAAAGAT-3' and reverse,
5'-GTTGATGTGCTGCTGCGAGA-3'; IL-18 forward,
5'-TGGTTCCATGCTTTCTGGACTCCT-3' and reverse,
5'-TTCCTGGGCCAAGAGGAAGTGATT-3'; TNF-α forward,
5'-CAGGCGGTGCCTATGTCTC-3' and reverse,
5'-CGATCACCCCGAAGTTCAGTAG-3'; and β-actin forward,
5'-AGAGGGAAATCGTGCGTGAC-3' and reverse,
5'-CAATAGTGATGACCTGGCCGT-3'. All experiments were repeated in
triplicate.
Cell transfection
For the H/R + siCMPK2 group, CMPK2 siRNA was
transfected into RAW 264.7 cells using Lipofectamine®
3000 (Thermo Fisher Scientific, Inc.), and then cells were
transferred to complete medium 6 h after transfection at 37˚C, 24 h
before the H/R treatment, according to the manufacturer's protocol.
Similarly, for the H/R + scramble siRNA, H/R + siNLRP3, H/R +
siAIM2, H/R + NLRP3 siRNA + CMPK2 siRNA and H/R + AIM2 siRNA +
CMPK2 siRNA groups, scramble siRNA, NLRP3 siRNA, AIM2 siRNA, NLRP3
siRNA + CMPK2 siRNA and AIM2 siRNA + CMPK2 siRNA were transfected
into RAW 264.7 cells in the same way, respectively, and then cells
were transferred to complete medium 6 h after transfection at 37˚C,
24 h before the H/R treatment.
CMPK2, NLRP3, AIM2 and scramble siRNA were obtained
from Shanghai GenePharma Co., Ltd. The sequences were as follows:
CMPK2 sense, 5'-GGCAAUUAUCUCGUGGCUUTT-3' and antisense,
5'-AAGCCACGAGAUAAUUGCCTT-3'; NLRP3 sense,
5'-CAACAGGAGAGACCUUUAUTT-3' and antisense,
5'-AUAAAGGUCUCUCCUGUUGTT-3'; AIM2 sense,
5'-GCAGUGACAAUGACUUUAATT-3' and antisense,
5'-UUAAAGUCAUUGUCACUGCTT-3'; scramble siRNA sense,
5'-UUCUCCGAACGUGUCACGUTT-3' and antisense,
5'-ACGUGACACGUUCGGAGAATT-3'
siRNA injection into mice
Mice were randomly divided into four groups: Sham
group, I/R group, I/R + scramble siRNA group and I/R + CMPK2 siRNA
group; with 5 mice in each group. With 24 h before the operation,
CMPK2 siRNA and scramble siRNA were injected into mice of the CMPK2
siRNA group and the scramble siRNA group (2 mg/kg; dissolved in
normal saline; 100 µl injection volume), respectively, via the tail
vein, and the same volume of saline was injected into mice of the
sham group and I/R group via the tail vein.
Determination of hepatic enzyme
Alanine aminotransferase (ALT; cat. no. C009-2-1)
and aspartate aminotransferase (AST; cat. no. C010-2-1; both from
Nanjing Jiancheng Bioengineering Institute) serum expression levels
in mice were measured using special diagnostic kits according to
the manufacturer's protocol. A microplate system (BioTek
Instruments, Inc.) was applied to measure the sample absorbance at
510 nm.
Hematoxylin & eosin (H&E)
staining
Hepatic tissues were fixed with paraformaldehyde
(4%) for 24 h at room temperature and embedded in paraffin. Tissue
specimens were then cut into segments (4.5 µm thick). Then, H&E
staining was performed (staining with hematoxylin for 8 min
followed by staining with eosin for 4 min at room temperature). The
tissues were viewed through an optical microscope (Olympus
Corporation; magnification, x200).
ELISA
Cell supernatant was collected using a sterile tube,
centrifuged at 1,000 x g at 4˚C for 20 min, and the supernatant was
further collected. Blood was collected using a pyrogen- and
endotoxin-free test tube. The blood was coagulated naturally at
room temperature for 20 min, and then centrifuged at 1,000 x g at
4˚C for 20 min to collect the serum. The levels of IL-18 (cat no.
EMC011) and IL-1β (cat no. EMC001b) cytokines in cellular
supernatant and mice serum were detected using special kits from
Neobioscience Technology Co., Ltd. according to the manufacturer's
protocol, and subsequently quantified via a microplate system
(BioTek Instruments, Inc.) at 450 nm.
Western blot analysis
Total protein was harvested with RIPA Lysis Buffer
(Beyotime Institute of Biotechnology) and diluted with SDS-PAGE
Sample Loading Buffer (5X) (Beyotime Institute of Biotechnology).
The expression levels of the target proteins were detected using
the protein concentration assay kit (Beyotime Institute of
Biotechnology). The proteins (50 µg protein/lane) were separated
via SDS-PAGE (12% gel). Subsequently, the target protein was
transferred to the PVDF membranes. After 2 h blocking with skimmed
milk (5%) at 4˚C, the PVDF membranes, which the target proteins had
been transferred to, were washed three times with TBST (0.1%
Tween-20). Subsequently, the membranes were co-incubated overnight
at 4˚C with CMPK2 (cat. no. ab139720; 1:1,000; Abcam), NLRP3 (cat.
no. 15101; 1:1,000; Cell Signaling Technology, Inc.),
cleaved-caspase-1 (cat. no. sc-398715; 1:1,000; Santa Cruz
Biotechnology, Inc.), IL-1β (cat. no. A16288; 1:1,000; ABclonal
Biotech Co., Ltd.), IL-18 (cat. no. bs-0529R; 1:1,000; BIOSS), AIM2
(cat. no. sc-515642; 1:1,000, Santa Cruz Biotechnology, Inc.) and
β-actin (cat. no. 66009-1-lg; 1:5,000, ProteinTech Group, Inc.).
The PVDF membranes were washed and co-incubated with the matching
secondary antibodies (Beyotime Institute of Biotechnology) for 1 h
at 4˚C. BeyoECL Moon (Beyotime Institute of Biotechnology) was used
to observe (Bio-Rad Biotechnology) the target protein.
Statistical analysis
All statistical analyses of data were performed
using SPSS software (version 20.0; IBM Corp.) and are presented as
the mean ± standard deviation. The experiments in vitro were
performed with 3 independent repeats and the experiments in
vivo were performed with 5 independent repeats (3 samples in
each group). Unpaired Student's t-test was used to calculate the
differences between two groups, while one-way ANOVA followed by
Bonferroni's post hoc test was used to analyze multiple groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
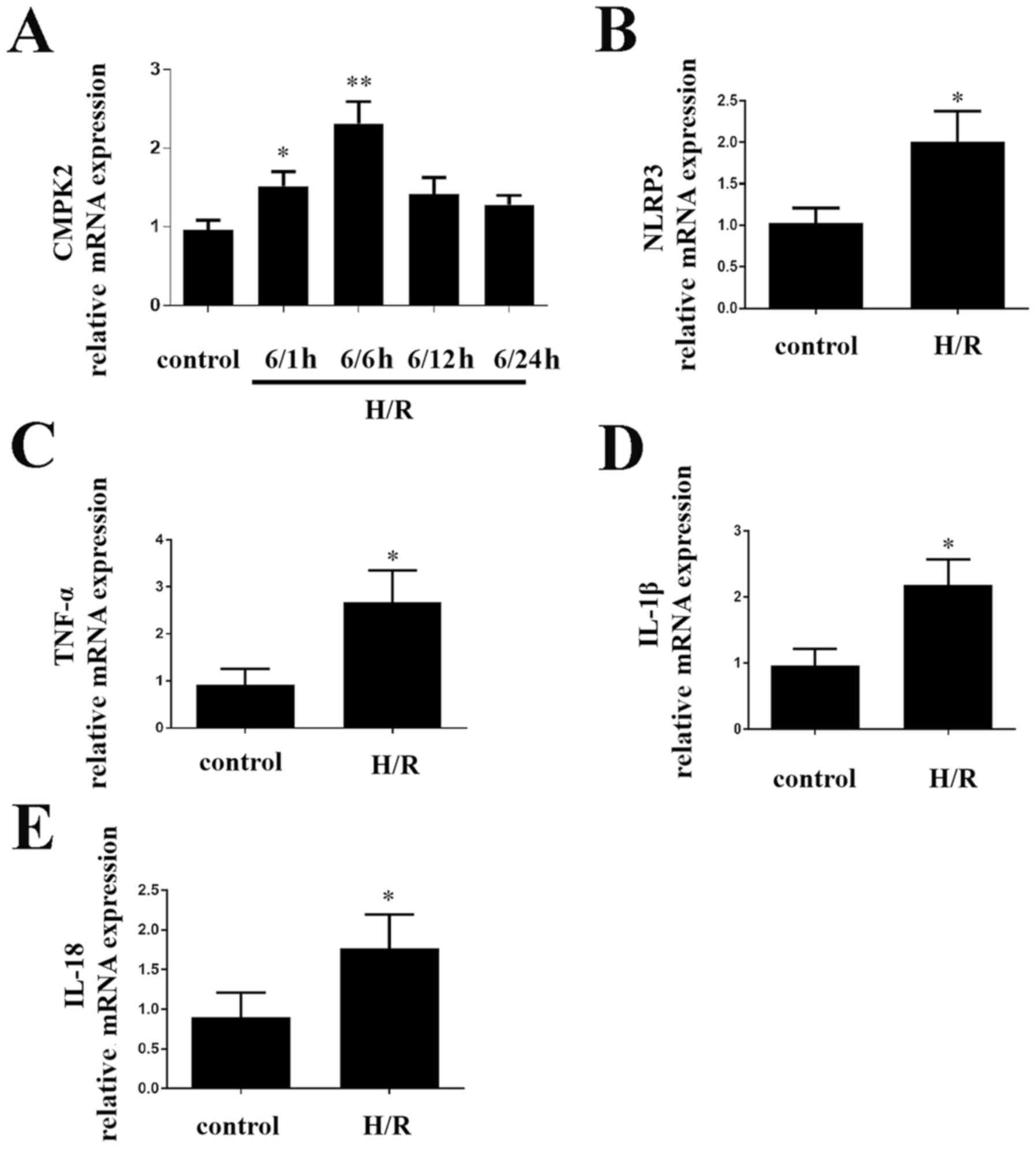
H/R treatment alters the expression
levels of CMPK2 and NLRP3 in cells
In order to determine the impact of CMPK2 on
H/R-induced inflammatory response in cells, the present study first
detected the CMPK2 levels. CMPK2 (Fig.
1A), NLRP3 (Fig. 1B) and TNF-α,
IL-1β and IL-18 (Fig. 1C-E,
respectively) mRNA expression levels were increased in RAW 264.7
cells compared with the control group. After reoxygenation for 6 h,
CMPK2 mRNA levels were significantly higher (Fig. 1A). Therefore, 6/6 h was selected as
an appropriate H/R time node (Fig.
1A). The results demonstrated that increased CMPK2 is likely to
be associated with liver I/R.
Knockdown of CMPK2 decreases
NLRP3-associated inflammation factor
The present study further investigated the molecular
mechanisms underlying CMPK2 in H/R-induced inflammatory model. In
order to determine the effect of CMPK2 on H/R-associated
inflammation, cells were transfected with CMPK2 siRNA and scramble
siRNA. The efficacy of transfection was then evaluated by
performing a western blot assay. It was revealed that CMPK2
expression level was decreased following the transfection of CMPK2
siRNA compared with the scramble siRNA (Fig. 2A).
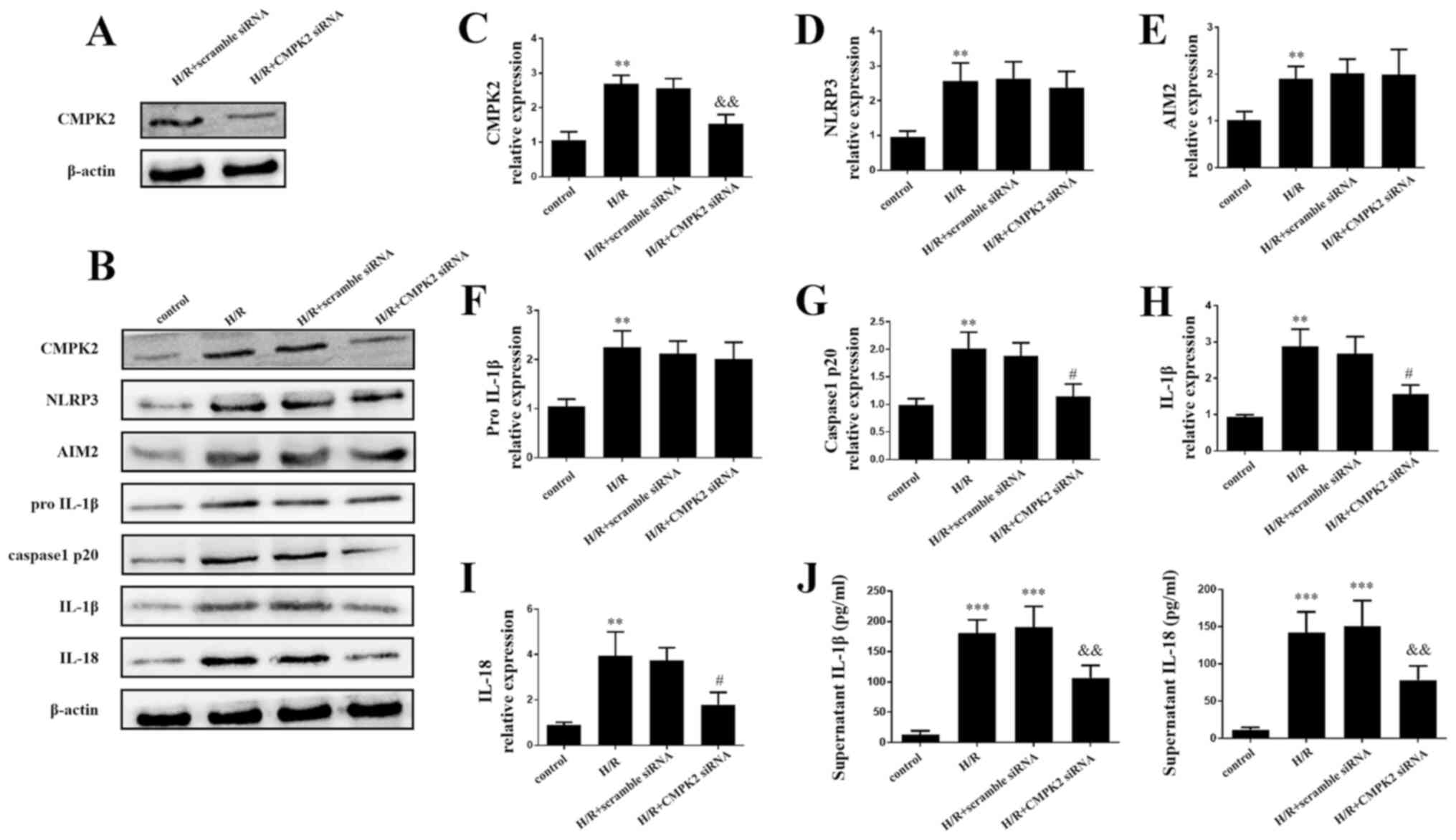 | Figure 2Knockdown of CMPK2 decreased the
NLRP3-associated inflammation factor. (A) Protein detection of
CMPK2 after H/R treatment in scramble siRNA or CMPK2
siRNA-transfected RAW 264.7 cells. (B) Western blotting and
subsequent quantification of (C) CMPK2, (D) NLRP3, (E) AIM2, (F)
pro-IL-1β, (G) cleaved-caspase-1 (H) IL-1β and (I) IL-18 protein
levels. (J) IL-18 and IL-1β levels in the supernatant were detected
by ELISA. **P<0.01, ***P<0.001 vs.
control group, #P<0.05,
&&P<0.01 vs. H/R group. CMPK2, cytidine
monophosphate kinase 2; NLRP3, NLR family pyrin domain containing
3; H/R, hypoxia/reoxygenation; siRNA, small interfering RNA; AIM2,
absent in melanoma 2. |
The results of the western blot assay and ELISAs
demonstrated that the protein levels of CMPK2, NLRP3, AIM2,
cleaved-caspase-1, IL-18, pro-IL-1β and IL-1β in both H/R groups
and H/R + scramble siRNA groups were increased compared with the
control group (Fig. 2B-J). Notably,
the H/R + CMPK2 siRNA group exhibited decreased expression of
CMPK2, cleaved-caspase-1, IL-18 and IL-1β compared with the H/R
group; however, there was no change in the protein levels of AIM2,
NLRP3 and pro-IL-1β, compared with the H/R groups (Fig. 2B-H). The results of the present
study indicate that CMPK2 may increase H/R-induced inflammation by
activating (but not priming) the NLRP3 inflammasome.
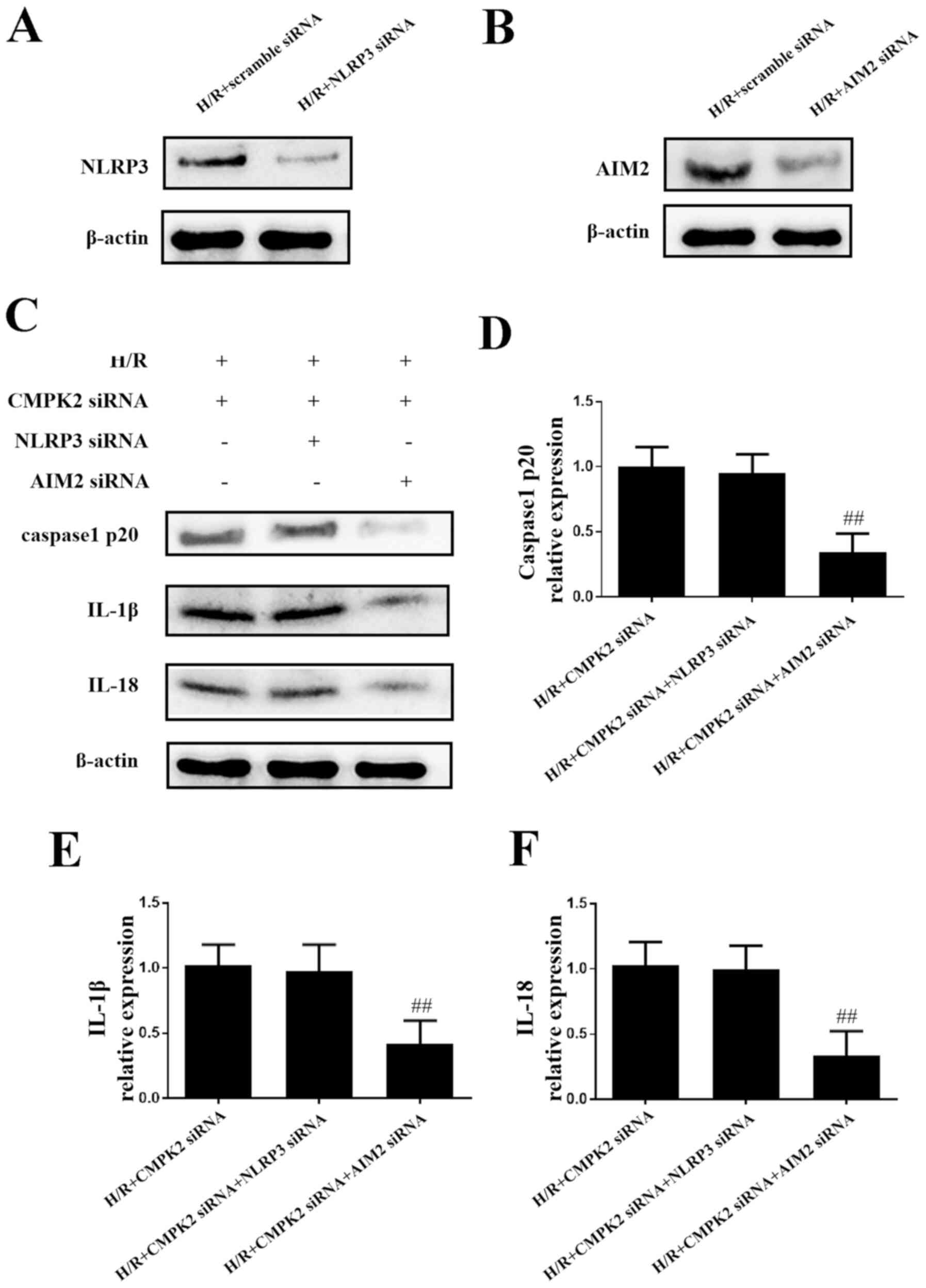
Knockdown of CMPK2 decreases the
expression levels of IL-18 and IL-1β by inhibiting the NLRP3
inflammasome, and not by inhibiting AIM2
In order to eliminate the possibility that CMPK2 has
an effect on AIM2 expression levels and to investigate the role of
CMPK2 in the NLRP3 pathway in H/R-induced inflammation, NLRP3 siRNA
and AIM2 siRNA were transfected into cells to knock down the
expression of NLRP3 and AIM2, respectively (Fig. 3A and B). As presented in Fig. 3C-F, the protein levels of
cleaved-caspase-1, IL-18 and IL-1β in the H/R + CMPK2 siRNA + NLRP3
siRNA groups remained unchanged, compared with the H/R + CMPK2
siRNA groups. Knockdown of CMPK2 + AIM2, but not CMPK2 + NLRP3,
suppressed the synthesis of cleaved-caspase-1, IL-18 and IL-1β
(Fig. 3C-F). In summary, knockdown
of NLRP3, but not AIM2, had no effect on cleaved-caspase-1, IL-18
and IL-1β expression levels following knockdown of CMPK2. This
indicates that the knockdown of CMPK2 decreased H/R-induced IL-18
and IL-1β synthesis by targeting the NLRP3 pathway and not
AIM2.
Knockdown of CMPK2 alleviates
I/R-induced deterioration of hepatic injury
In order to assess the impact of CMPK2 on hepatic
I/R induced liver dysfunction, mice were injected with CMPK2 siRNA
or scramble siRNA via the tail vein, followed by hepatic I/R
(Fig. 4A). The secretion of IL-18
and IL-1β in mouse serum was measured using an ELISA. It was
revealed that treatment with CMPK2 siRNA significantly decreased
the secretion of serum IL-18 and IL-1β, while treatment with
scramble siRNA resulted in an unchanged secretion of serum IL-18
and IL-1β, compared with the I/R group (Fig. 4B and C). In addition, the present study detected
the contents of serum ALT and AST in each group. Compared with the
scramble siRNA group, the levels of ALT and AST, which are released
following I/R, were significantly inhibited in the CMPK2 siRNA
group (Fig. 4D and E). Furthermore, H&E staining was used
to assess the damage following hepatic I/R injury.
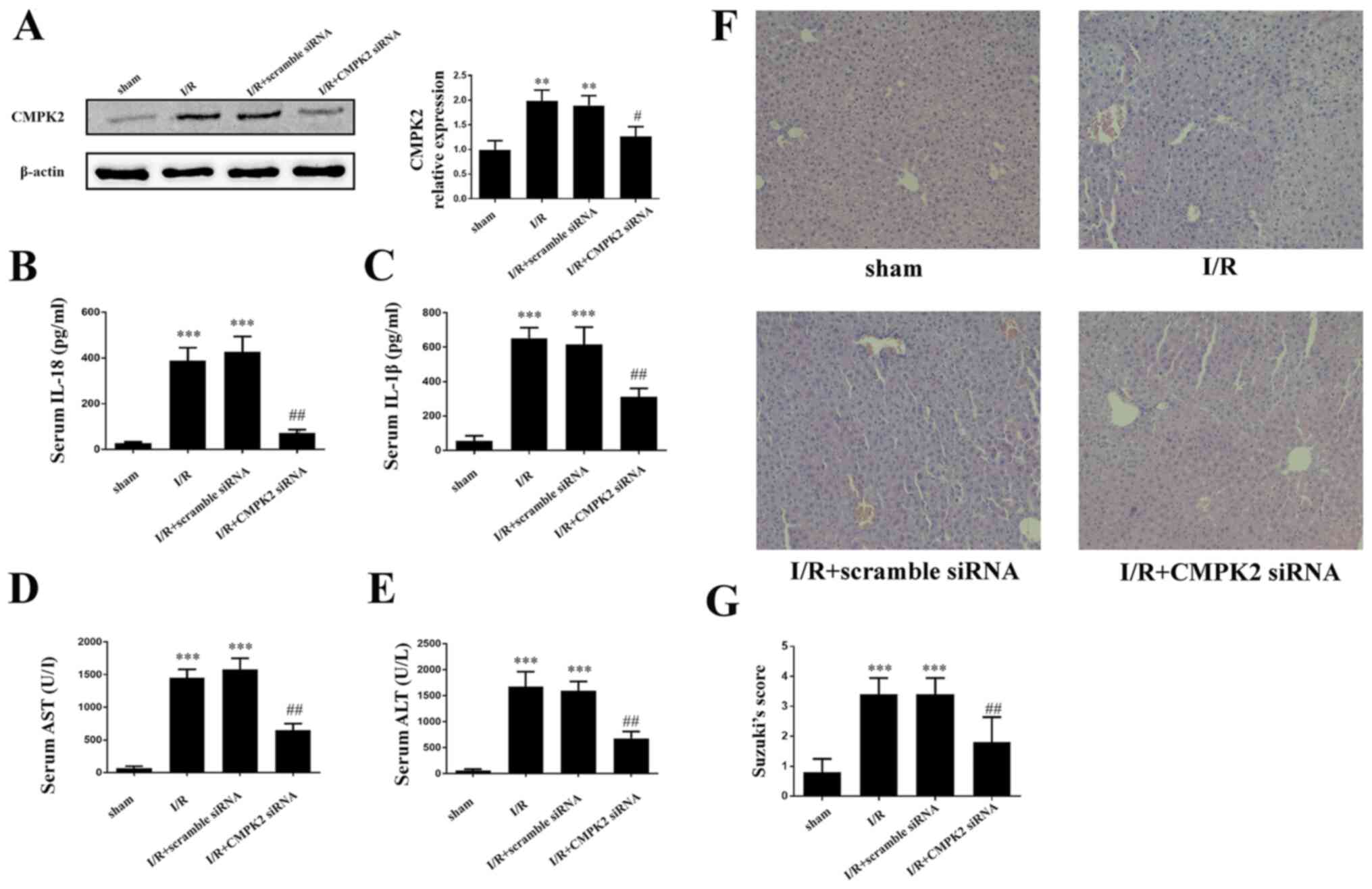 | Figure 4Suppression of CMPK2 alleviates
I/R-induced deterioration of hepatic injury. (A) Protein detection
of CMPK2 in liver tissue. Secretion of (B) IL-18 and (C) IL-1β was
measured by ELISA. Hepatocellular function of experimental mice was
evaluated by detecting the release of (D) AST and (E) ALT. (F and
G) Histological changes of the liver in the sham, I/R, I/R +
scramble siRNA and I/R + CMPK2 siRNA groups. Original
magnification, x200. **P<0.01,
***P<0.001 vs. sham group; #P<0.05,
##P<0.01 vs. I/R group. CMPK2, cytidine monophosphate
kinase 2; NLRP3, NLR family pyrin domain containing 3; I/R,
ischemia/reperfusion; siRNA, small interfering RNA; ALT, alanine
aminotransferase; AST, aspartate aminotransferase. |
Compared with the scramble siRNA group, the
amelioration of cellular swelling, bubble degeneration and
inflammatory cell infiltration were detected in the CMPK2 knockdown
group following hepatic I/R (Fig.
4F and G).
Discussion
As a severe complication of liver transplantation,
hepatic I/R injury increases the possibility of postoperative
infection, transplant failure and mortality rate (29). Previously, several studies have
indicated that hepatic I/R injury triggered necrocytosis, metabolic
disturbances and the inflammatory immune response (4,29).
Among these, the inflammatory immune response has been demonstrated
to be a crucial contributor to I/R injury (4,29).
NLRP3 inflammasome has been previously identified as
a pivotal regulator in the inflammatory immune response (7). AIM2 inflammasome has also been
demonstrated to participate in IL-18 and IL-1β synthesis (30). Although the NLRP3 pathway serves an
essential role in several diseases (7), the mechanism that regulates its
activation is not yet fully understood. Likewise, the role of the
NLRP3 inflammasome in hepatic I/R injury has not yet been fully
elucidated. In the present study it was demonstrated that H/R
increased the expression level of NLRP3. Furthermore, the
activation of NLRP3 inflammasome in both cerebral and cardiac I/R
injury has been demonstrated in previous studies (31,32),
the results of which are consistent with those of the present
study.
In the present study, it was revealed that that H/R
treatment enhanced the expression levels of CMPK2, IL-18 and IL-1β;
the latter two proinflammatory factors were reported to be the
final products of the NLRP3 signaling pathway (7). The increased expression of CMPK2
following H/R treatment indicated that CMPK2 may have a potential
function associated with the H/R-induced inflammatory process. In
order to confirm the effect of CMPK2 on inflammation, the present
study transfected cells with CMPK2 siRNA to decrease the protein
levels of CMPK2. Although CMPK2 knockdown did not alter the
expression levels of NLRP3 and pro-IL-1β, it did decrease the
protein levels of cleaved-caspase-1, IL-18 and IL-1β following H/R
treatment. In addition, it was revealed that NLRP3 knockdown, but
not AIM2 deficiency, had no additional impact on cleaved-caspase-1,
IL-18 and IL-1β synthesis in the CMPK2 siRNA group. These results
indicated that CMPK2 affected the NLRP3 pathway rather than the
AIM2 pathway during hepatic I/R injury, by targeting the NLRP3
pathway. Consistent with the results of the present study, a
previous inflammatory model demonstrated that CMPK2 had similar
effects on the NLRP3 pathway (8).
The present study only focused on the effect of CMPK2 knockdown,
and not on CMPK2 overexpression, in hepatic I/R. Future studies
will continue to investigate CMPK2 in a comprehensive and in-depth
manner.
Previous studies have demonstrated that CMPK2
promoted mtDNA synthesis by providing dCTP (17). Several other studies have reported
that mtDNA deficiency inhibited cleaved-caspase-1 and IL-1β
production caused by NLRP3 inflammasome activation (8,33).
Based on these consequences, it was speculated that the approach of
CMPK2-mediated NLRP3 inflammasome activation may be connected with
CMPK2-dependent mtDNA synthesis.
To the best of the authors knowledge, the present
study reported for the first time that CMPK2 accelerates hepatic
I/R injury by activating the NLRP3 inflammasome. CMPK2
siRNA-mediated knockdown not only decreased the NLRP3
inflammasome-mediated inflammatory response, but also relieved
hepatic I/R injury. Furthermore, the results of the present study
identified a potentially promising therapeutic strategy to
alleviate hepatic I/R injury by targeting CMPK2-mediated NLRP3
inflammasome activation.
Acknowledgements
Not applicable.
Funding
The present study was funded by the National Natural Science
Foundation of China (grant no. 81703063), the National Natural
Science Foundation of China (grant no. 81873592) and the graduate
tutor team construction project of Chongqing Municipal Education
Commission Foundation, China (grant no. dstd201801).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL and ZW contributed to the experimental design,
data acquisition and preparation of the manuscript. JP, WC and YZ
assessed the liver function and performed the statistical analysis.
TM, DZ and ZH contributed to the analysis and interpretation of the
data and revised the present article critically for important
intellectual content. YL and ZW confirm the authenticity of all the
raw data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experiments were approved by the Animal Care and
Use Committee of Chongqing Medical University (Chongqing,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Peralta C, Jiménez-Castro MB and
Gracia-Sancho J: Hepatic ischemia and reperfusion injury: Effects
on the liver sinusoidal milieu. J Hepatol. 59:1094–1106.
2013.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Eltzschig HK and Eckle T: Ischemia and
reperfusion-from mechanism to translation. Nat Med. 17:1391–1401.
2011.PubMed/NCBI View
Article : Google Scholar
|
|
3
|
Ni D, Wei H, Chen W, Bao Q, Rosenkrans ZT,
Barnhart TE, Ferreira CA, Wang Y, Yao H, Sun T, et al: Ceria
nanoparticles meet hepatic ischemia-reperfusion injury: The perfect
imperfection. Adv Mater. 31(e1902956)2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Zhai Y, Petrowsky H, Hong JC, Busuttil RW
and Kupiec-Weglinski JW: Ischaemia-reperfusion injury in liver
transplantation--from bench to bedside. Nat Rev Gastroenterol
Hepatol. 10:79–89. 2013.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Li P, He K, Li J, Liu Z and Gong J: The
role of Kupffer cells in hepatic diseases. Mol Immunol. 85:222–229.
2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Gross O, Thomas CJ, Guarda G and Tschopp
J: The inflammasome: An integrated view. Immunol Rev. 243:136–151.
2011.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Swanson KV, Deng M and Ting JP: The NLRP3
inflammasome: Molecular activation and regulation to therapeutics.
Nat Rev Immunol. 19:477–489. 2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zhong Z, Liang S, Sanchez-Lopez E, He F,
Shalapour S, Lin XJ, Wong J, Ding S, Seki E, Schnabl B, et al: New
mitochondrial DNA synthesis enables NLRP3 inflammasome activation.
Nature. 560:198–203. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zhuang T, Liu J, Chen X, Zhang L, Pi J,
Sun H, Li L, Bauer R, Wang H, Yu Z, et al: Endothelial Foxp1
suppresses atherosclerosis via modulation of Nlrp3 inflammasome
activation. Circ Res. 125:590–605. 2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Martínez-García JJ, Martínez-Banaclocha H,
Angosto-Bazarra D, de Torre-Minguela C, Baroja-Mazo A, Alarcón-Vila
C, Martínez-Alarcón L, Amores-Iniesta J, Martín-Sánchez F, Ercole
GA, et al: P2X7 receptor induces mitochondrial failure in monocytes
and compromises NLRP3 inflammasome activation during sepsis. Nat
Commun. 10(2711)2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Mehto S, Jena KK, Nath P and Chauhan S,
Kolapalli SP, Das SK, Sahoo PK, Jain A, Taylor GA and Chauhan S:
The crohn's disease risk factor IRGM limits NLRP3 inflammasome
activation by impeding its assembly and by mediating its selective
autophagy. Mol Cell. 73:429–445.e7. 2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Mauro AG, Bonaventura A, Mezzaroma E,
Quader M and Toldo S: NLRP3 inflammasome in acute myocardial
infarction. J Cardiovasc Pharmacol. 74:175–187. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Ghiringhelli F, Apetoh L, Tesniere A,
Aymeric L, Ma Y, Ortiz C, Vermaelen K, Panaretakis T, Mignot G,
Ullrich E, et al: Activation of the NLRP3 inflammasome in dendritic
cells induces IL-1beta-dependent adaptive immunity against tumors.
Nat Med. 15:1170–1178. 2009.PubMed/NCBI View
Article : Google Scholar
|
|
14
|
Li C, Jin Y, Wei S, Sun Y, Jiang L, Zhu Q,
Farmer DG, Busuttil RW, Kupiec-Weglinski JW and Ke B: Hippo
signaling controls NLR family pyrin domain containing 3 activation
and governs immunoregulation of mesenchymal stem cells in mouse
liver injury. Hepatology. 70:1714–1731. 2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Miyauchi T, Uchida Y, Kadono K, Hirao H,
Kawasoe J, Watanabe T, Ueda S, Okajima H, Terajima H and Uemoto S:
Up-regulation of FOXO1 and reduced inflammation by β-hydroxybutyric
acid are essential diet restriction benefits against liver injury.
Proc Natl Acad Sci USA. 116:13533–13542. 2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Yue S, Zhu J, Zhang M, Li C, Zhou X, Zhou
M, Ke M, Busuttil RW, Ying QL, Kupiec-Weglinski JW, et al: The
myeloid heat shock transcription factor 1/β-catenin axis regulates
NLR family, pyrin domain-containing 3 inflammasome activation in
mouse liver ischemia/reperfusion injury. Hepatology. 64:1683–1698.
2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Xu Y, Johansson M and Karlsson A: Human
UMP-CMP kinase 2, a novel nucleoside monophosphate kinase localized
in mitochondria. J Biol Chem. 283:1563–1571. 2008.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhang F, Qi Y, Harrison TJ, Luo B, Zhou Y,
Li X, Song A, Huang W and Wang Y: Hepatitis E genotype 4 virus from
feces of monkeys infected experimentally can be cultured in
PLC/PRF/5 cells and upregulate host interferon-inducible genes. J
Med Virol. 86:1736–1744. 2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Liu W, Chen B, Chen L, Yao J, Liu J, Kuang
M, Wang F, Wang Y, Elkady G, Lu Y, et al: Identification of fish
CMPK2 as an interferon stimulated gene against SVCV infection. Fish
Shellfish Immunol. 92:125–132. 2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ueta M, Kawai T, Yokoi N, Akira S and
Kinoshita S: Contribution of IPS-1 to polyI: C-induced cytokine
production in conjunctival epithelial cells. Biochem Biophys Res
Commun. 404:419–423. 2011.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Goertsches RH, Hecker M, Koczan D,
Serrano-Fernandez P, Moeller S, Thiesen HJ and Zettl UK: Long-term
genome-wide blood RNA expression profiles yield novel molecular
response candidates for IFN-beta-1b treatment in relapsing
remitting MS. Pharmacogenomics. 11:147–61. 2010.PubMed/NCBI View Article : Google Scholar
|
|
22
|
El-Diwany R, Soliman M, Sugawara S,
Breitwieser F, Skaist A, Coggiano C, Sangal N, Chattergoon M,
Bailey JR, Siliciano RF, et al: CMPK2 and BCL-G are associated with
type 1 interferon-induced HIV restriction in humans. Sci Adv.
4(eaat0843)2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Kommadath A, Bao H, Choi I, Reecy JM,
Koltes JE, Fritz-Waters E, Eisley CJ, Grant JR, Rowland RR, Tuggle
CK, et al: Genetic architecture of gene expression underlying
variation in host response to porcine reproductive and respiratory
syndrome virus infection. Sci Rep. 7(46203)2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Chen YL, Lin DW and Chang ZF:
Identification of a putative human mitochondrial thymidine
monophosphate kinase associated with monocytic/macrophage terminal
differentiation. Genes Cells. 13:679–689. 2008.PubMed/NCBI View Article : Google Scholar
|
|
25
|
National Research Council Committee for
the Update of the Guide for the C and Use of Laboratory A: The
National Academies Collection: Reports funded by National
Institutes of Health. In: Guide for the Care and Use of Laboratory
Animals. National Academies Press (US) Copyright © 2011. National
Academy of Sciences, Washington, DC, 2011.
|
|
26
|
He D, Guo Z, Pu JL, Zheng DF, Wei XF, Liu
R, Tang CY and Wu ZJ: Resveratrol preconditioning protects
hepatocytes against hepatic ischemia reperfusion injury via
Toll-like receptor 4/nuclear factor-κB signaling pathway in vitro
and in vivo. Int Immunopharmacol. 35:201–209. 2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Wang X, Mao W, Fang C, Tian S, Zhu X, Yang
L, Huang Z and Li H: Dusp14 protects against hepatic
ischaemia-reperfusion injury via Tak1 suppression. J Hepatol.
S0168-8278:32275–32284. 2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zhai Y, Busuttil RW and Kupiec-Weglinski
JW: Liver ischemia and reperfusion injury: New insights into
mechanisms of innate-adaptive immune-mediated tissue inflammation.
Am J Transplant. 11:1563–1569. 2011.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Lugrin J and Martinon F: The AIM2
inflammasome: Sensor of pathogens and cellular perturbations.
Immunol Rev. 281:99–114. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
He Q, Li Z, Meng C, Wu J, Zhao Y and Zhao
J: Parkin-dependent mitophagy is required for the inhibition of
ATF4 on NLRP3 inflammasome activation in cerebral
ischemia-reperfusion injury in rats. Cells. 8(897)2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Darwesh AM, Keshavarz-Bahaghighat H,
Jamieson KL and Seubert JM: Genetic deletion or pharmacological
inhibition of soluble epoxide hydrolase ameliorates cardiac
ischemia/reperfusion injury by attenuating NLRP3 inflammasome
activation. Int J Mol Sci. 20(3502)2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Zhou R, Yazdi AS, Menu P and Tschopp J: A
role for mitochondria in NLRP3 inflammasome activation. Nature.
469:221–225. 2011.PubMed/NCBI View Article : Google Scholar
|