Introduction
Vascular endothelial cells create a monolayer lining
on the surface of vascular intima. This is crucial in maintaining
the normal tension of blood vessels and the normal state of blood
vessels, and is a bridge between the blood and the vascular
muscular layer that participates in the development of blood
vessels. Endothelial cell apoptosis is the initiating factor of
vascular endothelial injury, which constitutes the pathological
foundation of various cardiovascular diseases, such as
atherosclerosis (1).
MicroRNAs (miRNAs/miRs) are a class of endogenous
non-protein coding, single-stranded small RNAs of 22-25 nucleotides
in length (2). In 1993, Lee et
al (3) discovered the first
miRNA that regulates temporal expression of cells in
Caenorhabditis elegans, known as Lin-4. Subsequently, novel
miRNAs have been identified in animals, plants and microorganisms,
which are involved in the regulation of coding proteins, and
regulate all pathological and physiological processes in
vivo (4).
Apoptosis is considered the initiating factor of
several cardiovascular diseases. Notably, miRNAs can mediate the
function of vascular endothelial cells by regulating their
apoptosis, and thus are involved in regulating the progression of
cardiovascular diseases (5).
Research on vascular endothelial cells have demonstrated the
significant change in the expression profile of miRNAs in human
umbilical vein endothelial cells (HUVECs) stimulated by oxidized
low density lipoprotein (OX-LDL) (6). It can inhibit Bcl-2 expression,
promote the production of reactive oxygen species induced by OX-LDL
via the mitochondrial apoptosis pathway, resulting in the apoptosis
of endothelial cells (7).
miR-216a-5p is a newly discovered miRNA (8-10).
Current literature focuses on the role of miR-216a-5p in tumors
(9,11). In addition, it has been reported
that miR-216a-5p can effectively improve the damage of bronchial
cells caused by H2O2 stimulation (12). However, the role of miR-216a-5p in
vascular endothelial injury and its molecular mechanism remain
unclear. In the present study, a model of vascular endothelial cell
injury induced by lipopolysaccharide (LPS) was established to
investigate the role of the knockdown and overexpression of
miR-216a-5p in vascular endothelial injury, and determine its
molecular mechanism.
Materials and methods
Cells and reagents
HUVECs were purchased from the American Type Culture
Collection. Cells were maintained with 5% CO2 at room
temperature in DMEM (Hyclone; Cytiva) supplemented with FBS
(Hangzhou Sijiqing Biological Engineering Materials Co., Ltd.). LPS
was obtained from Escherichia coli (Sigma-Aldrich; Merck
KGaA). Small interfering RNA (si)-miR-216a-5p, si-negative control
(NC), miR-NC and miR-216a-5p were purchased from Nanjing KeyGen
Biotech Co., Ltd. The PCR kit was purchased from Takara Bio, Inc.
Antibodies against Toll-like receptor 4 (TLR4; cat. no. ab13556;
1:500), MyD88 (cat. no. ab219413; 1:500) and phosphorylated
(p)-nuclear factor (NF)-κB(p65) (cat. no. ab76302; 1:500) were
purchased from Abcam, while the secondary antibody was purchased
from OriGene Technologies, Inc.
Cell transfection
Cell transfection was performed using
Lipofectamine® RNAiMAX reagent (Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
Subsequent experiments were performed 48 h post-transfection.
LPS treatment
Cells with routine culture and those transfected
with miR-216-5p for 48 h, after continued to culture for 24 h, the
cells were seeded into 6-well plates at a cell density of
1x105 cells/well or seeded into 96-well plates at the
cell density of 2x103 cells/well. Following incubation
for 24 h at room temperature, 1.0 mg/l LPS was added to each well
and cells were cultured for an additional 48 h.
Cell treatment protocols
Cells were classified into six treatment groups as
follows: No treatment (control), transfection with blank inhibitor
vector (si-NC), LPS treatment alone (LPS), transfection with
miR-216a-5p inhibitor (si-miR-216a-5p), transfection with miR-NC
(miR-NC) and transfection with miR-216a-5p+LPS treatment
(LPS+miR-216a-5p) groups. The sequences as following: si-NC:
5'-CAGUACUUUUGUGUAGUACAA-3'; si-miR-216a-5p,
5'-UCACAGUUGCCAGCUGAGAUUA; miR-NC, F: 5'-UUCUCCCAACGUGUCACGUTT-3';
R, 5'-ACG UGACACGUUCGGAGAATT-3'; miR-216a-5p F, 5'-UAAUCU
CAGCUGGCAACUGUGA-3'; miR-216a-5p R, 5'-ACAGUUG
CCAGCUGAGAUUAUU-3'.
EdU staining
Following treatment, cell proliferation was assessed
in each group via EdU staining (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Cells were observed
under a fluorescence microscope (x200).
Apoptosis analysis
Cell apoptosis was detected using the Annexin V-PI
Apoptosis Detection kit (Thermo Fisher Scientific, Inc.). Cells
were stained with Annexin V and PI, according to the manufacturer's
instructions. Cells were incubated for 30 min at room temperature
in the dark and detected via flow cytometric analysis by BD
FACSAria (BD Biosciences) and ModFit software version 3.2 (BD
Biosciences).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from cells using
TRIzol® (cat. no. 15596-026, Invitrogen; Thermo Fisher
Scientific, Inc.) reagent to detect the expression levels of
miR-216a-5p, TLR4, MyD88 and NF-κB(p65). The miScriptII RT reverse
transcription kit (Takara Bio, Inc.) was used to reverse transcribe
total RNA into cDNA. The extracted RNA was reverse transcribed into
cDNA using the RevertAid first strand cDNA synthesis kit to detect
the expression levels of TLR4, MyD88 and NF-κB(p65). The
synthesized cDNA was amplified and quantified using the Taqman
Real-time PCR Master Mixes SYBR-Green kit (Takara Bio, Inc.). The
quantitative system and reaction conditions of RT and amplification
were performed according to the manufacturer's instructions.
Relative expression levels were calculated using the
2-ΔΔCq method (13). U6
was the internal reference for miR-216a-5p, while GAPDH was the
internal reference for TLR4, MyD88 and NF-κB (p65). The primer
sequences used for qPCR are listed in Table I.
 | Table IPrimer sequences of different
genes. |
Table I
Primer sequences of different
genes.
| Gene name | Sequence
(5'-3') |
|---|
|
microRNA-216a-5p | F:
ATCCAGTGCGTGTCGTG |
| | R:
TGCTTAATCTCAGCTGGCA |
| U6 | F:
CTCGCTTCGGCAGCACA |
| | R:
ACGCTTCACGAATTTGCGT |
| Toll-like receptor
4 | F:
TGGATACGTTTCCTTATAAG |
| | R:
GAAATGGAGGCACCCCTTC |
| MyD88 | F:
ACCTGGCTGGTTTACACGTC |
| | R:
CTGCCAGAGACATTGCAGAA |
| NF-κB(p65) | F:
ATGCTTACTGGGTGCCAAAC |
| | R:
GGCAAGTCACTCAGCCTTTC |
| GAPDH | F:
AGGTCGGTGTGAACGGATTTG |
| | R:
5'-TGTAGACCATGTAGTTGAGG TCA-3' |
Western blotting
Cells in each group were treated with corresponding
treatments, and the protein samples were extracted by the whole
protein extraction kit (cat. no. KEP250; Nanjing KeyGen Biotech
Co., Ltd.) for electrophoresis. Protein concentration was measured
via the BCA methods. The collected lysate samples (20 µg/well) were
separated via SDS-PAGE on 12% gels, transferred onto nitrocellulose
membranes and blocked with 50 g/l skimmed milk for 2 h at room
temperature. The membranes were incubated with rabbit anti-mouse
TLR4 (cat. no. ab13556), MyD88 (cat. no. ab219413), p-NF-κB(p65)
(cat. no. ab76302) (all 1:500) and rabbit anti-mouse GAPDH (cat.
no. ab9485) (1:500) polyclonal antibodies for 12 h at 4˚C.
Following the primary incubation, membranes were incubated with
secondary antibody Goat Anti-Rabbit IgG (cat. no. ab150077;
1:1,000) at room temperature for 1 h. Protein bands were then
visualized using an ECL reagent (Thermo Fisher Scientific, Inc.),
using Tanon 5200 detection system (Tanon Science & Technology
Co., Ltd.) for imaging. Protein band densitometry was quantified
using ImageJ software for Windows V 1.52v (National Institutes of
Health) and normalized to GAPDH.
Cell immunofluorescence assay
Following treatment and once HUVECs reached
confluence (70-80%), cells were fixed with 4% paraformaldehyde for
30 min at room temperature and blocked with 10% normal goat serum
(Sigma-Aldrich; Merck KGaA) for 30 min at room temperature. Cells
were incubated with p-NF-κB(p65)1 antibody (1:200) in the wet box
overnight at 4˚C. Subsequently, cells were incubated with goat
anti-rabbit IgG at 37˚C for 30 min. Nuclei were stained with 5
µg/ml DAPI for 10 min at room temperature and subsequently sealed
with glycerin-buffered saline (Sigma-Aldrich; Merck KGaA). Cells
were observed under a fluorescence inverted phase contrast
microscope (magnification, x200).
Dual-luciferase reporter assay
The sequences of wild-type (WT) and mutant (MUT)
TLR4 mRNA in 3'-untranslated regions (UTRs) were synthesized and
cloned into fluorescent reporter plasmids (cat. no. KGAF040;
Nanjing KeyGen Biotech Co., Ltd.), named WT and MUT. The
fluorescent reporter plasmids was subsequently transfected using
Lipofectamine® 3000 (Thermo Fisher Scientific, Inc.)
into HUVECs, with miR-216a-5p mimic sequence or miR-NC,
respectively for 24 h at room temperature. Transfected cells were
seeded into 96-well plates at the density of 1x104
cells/well and cultured for 48 h. Luciferase activities were
measured using the Promega dual-luciferase reporter assay system
(Promega Corporation). Firefly luciferase activity was normalized
to Renilla luciferase activity.
Statistical analysis
Statistical analysis was performed using the SPSS
17.0 software (SPSS, Inc.). All experiment repeated three times.
Data are presented as the mean ± SD. Tukey's following one-way
ANOVA were used for pairwise comparison between multiple groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effect of miR-216a-5p knockdown and
LPS intervention on endothelial cell proliferation
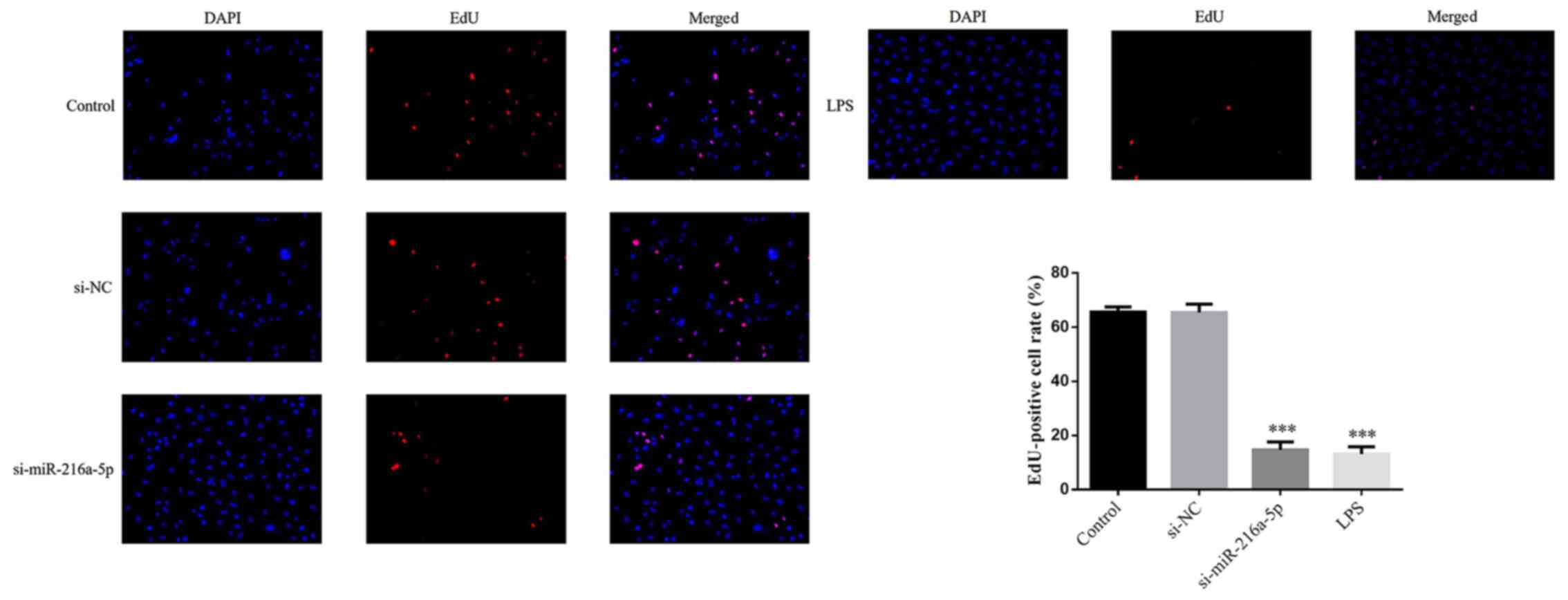
No significant difference in the number of
EdU-positive cells was observed in the si-NC group compared with
the control group (P>0.05; Fig.
1). The results demonstrated that transfection with si-NC
caused no injury to HUVECs. Notably, the number of EdU-positive
cells significantly decreased in the si-miR-216a-5p and LPS groups
(P<0.001; Fig. 1). Taken
together, the results suggest that miR-216a-5p knockdown or LPS
stimulation can decrease the proliferation of HUVECs.
Effect of miR-216a-5p knockdown and
LPS intervention on endothelial cell apoptosis
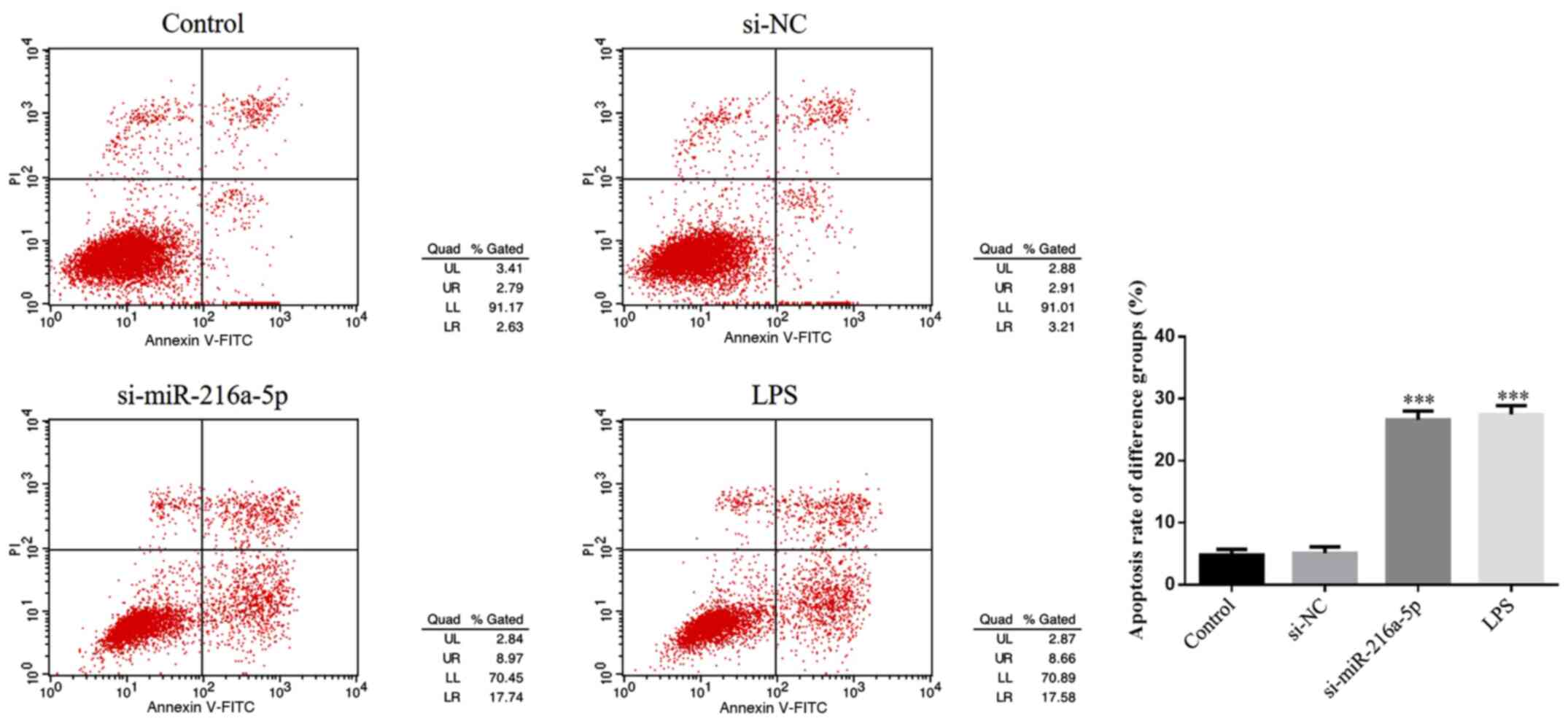
No significance difference in cell apoptosis was
observed between the control and si-NC groups (P>0.05; Fig. 2), suggesting that transfection with
si-NC caused no injury to HUVECs. However, cell apoptosis was
notably increased in the si-miR-216a-5p and LPS groups (P<0.001;
Fig. 2). Collectively, the results
suggest that miR-216a-5p knockdown or LPS intervention promote the
apoptosis of HUVECs.
Effect of miR-216a-5p knockdown and
LPS intervention on gene expression
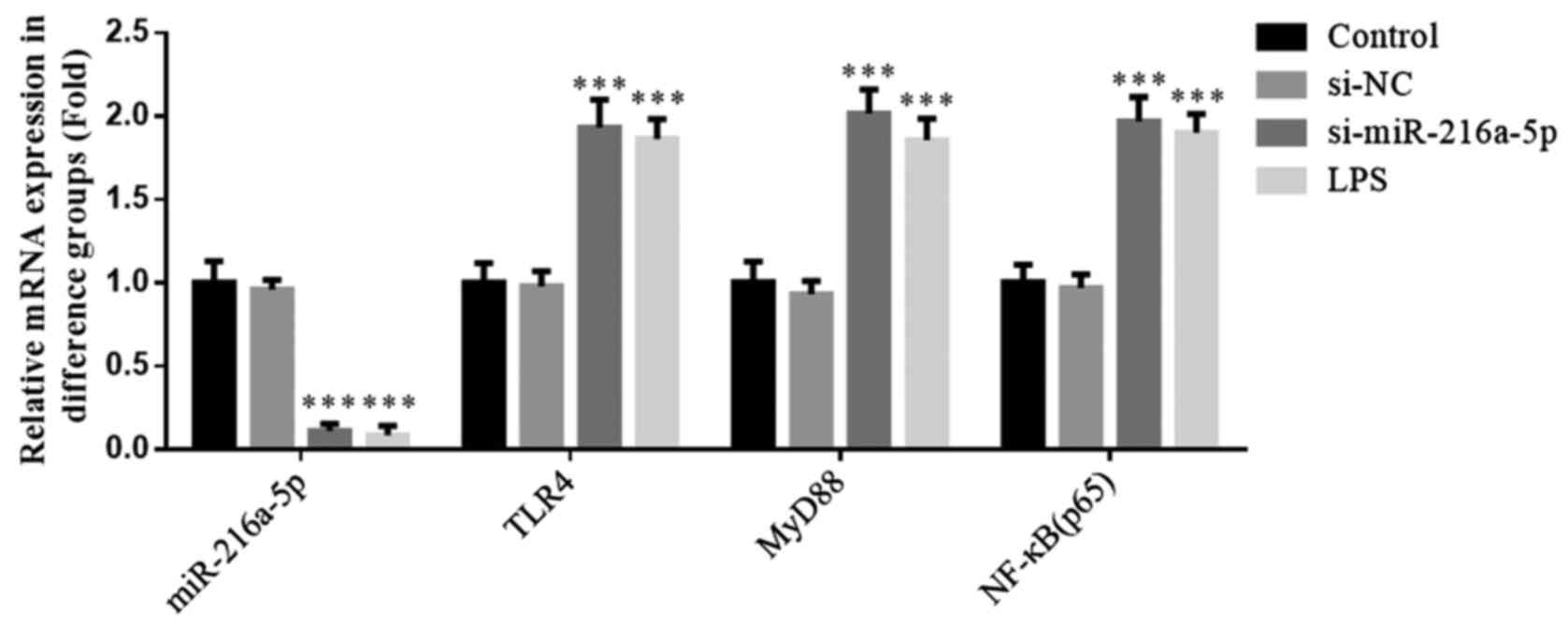
No significant differences were observed in the
expression levels of miR-216a-5p, TLR4, MyD88 and NF-κB(p65)
between the control and si-NC groups (P>0.05; Fig. 3). Moreover, the results revealed no
influence of si-NC transfection in HUVECs on miR-216a-5p, TLR4,
MyD88 and NF-κB(p65) mRNA expression levels. Furthermore,
miR-216a-5p expression was notably decreased in the si-miR-216a-5p
and LPS groups, while the mRNA expression levels of TLR4, MyD88 and
NF-κB(p65) were significantly increased (P<0.001; Fig. 3). Taken together, the results
suggest that miR-216a-5p knockdown or LPS intervention affect the
expression levels of miR-216a-5p, TLR4, MyD88 and NF-κB(p65).
Effect of miR-216a-5p knockdown and
LPS intervention on protein expression
No significant differences in the protein expression
levels of TLR4, MyD88 and NF-κB(p65) were observed between the
control and si-NC groups (P>0.05; Fig. 4). The protein expression levels of
TLR4, MyD88 and NF-κB(p65) were significantly increased in the
si-miR-216a-5p and LPS groups (P<0.001; Fig. 4). Collectively, these results
suggest that miR-216a-5p knockdown or LPS intervention affect the
protein expression levels of miR-216a-5p, TLR4, MyD88 and
NF-κB(p65).
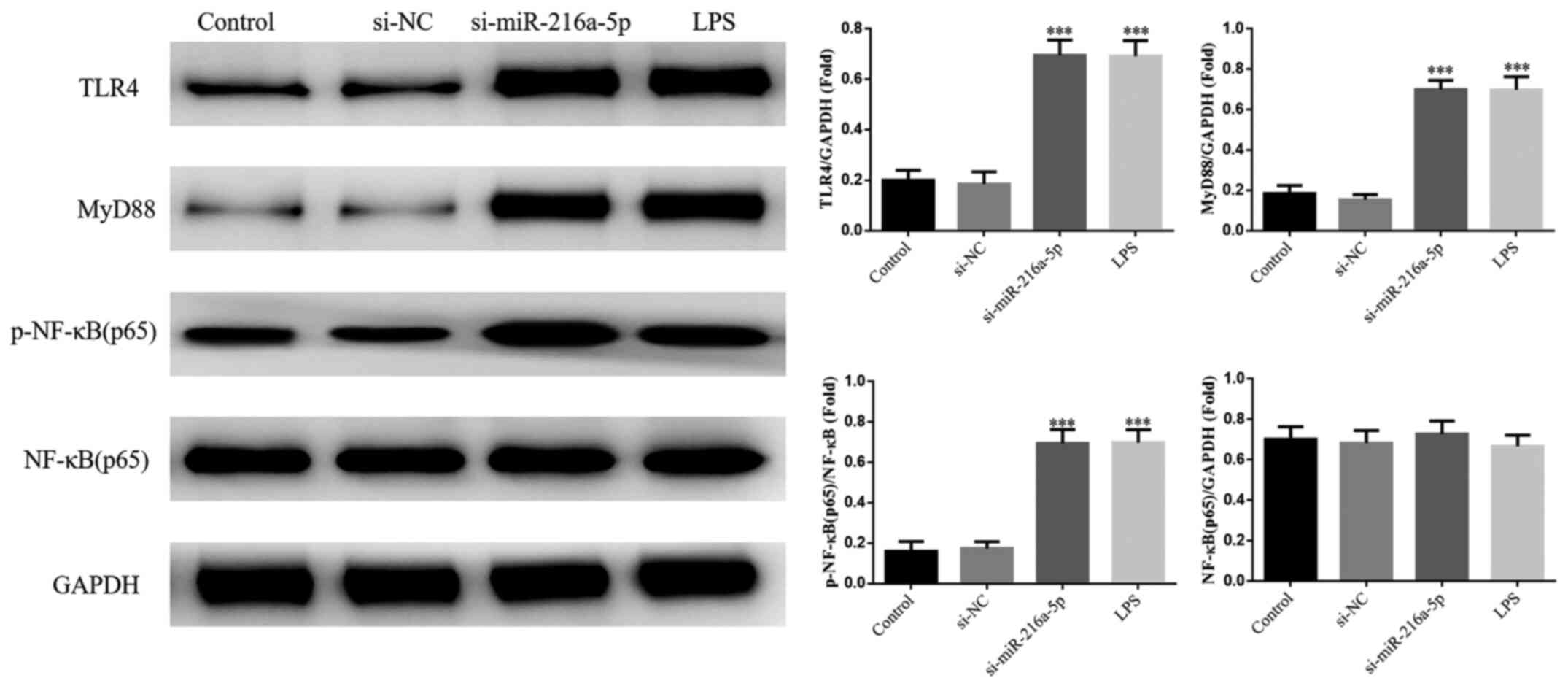 | Figure 4Effect of miR-216a-5p knockdown and
LPS intervention on protein expression. Control, cells were
cultured under normal conditions; si-NC, cells were transfected
with si-NC; si-miR-216a-5p, cells were transfected with
si-miR-216a-5p; LPS, cells were treated with LPS (1.0 mg/l).
***P<0.001 vs. control. miR, microRNA; LPS,
lipopolysaccharide; si, small interfering RNA; NC, negative
control; TLR4, Toll-like receptor 4; p-, phosphorylated; NF-κB,
nuclear factor-κB. |
Effect of miR-216a-5p knockdown and
LPS intervention on the protein transportation of p-NF-κB(p65) to
the nucleus
No significant differences in the protein
transportation of p-NF-κB(p65) to the nucleus were observed between
the control and si-NC groups (P>0.05; Fig. 5). Notably, the protein
transportation of p-NF-κB(p65) to the nucleus was significantly
increased in the si-miR-216a-5p and LPS groups (P<0.001;
Fig. 5). Taken together, the
results suggest that miR-216a-5p knockdown or LPS intervention
affect protein transportation of p-NF-κB(p65) to the nucleus.
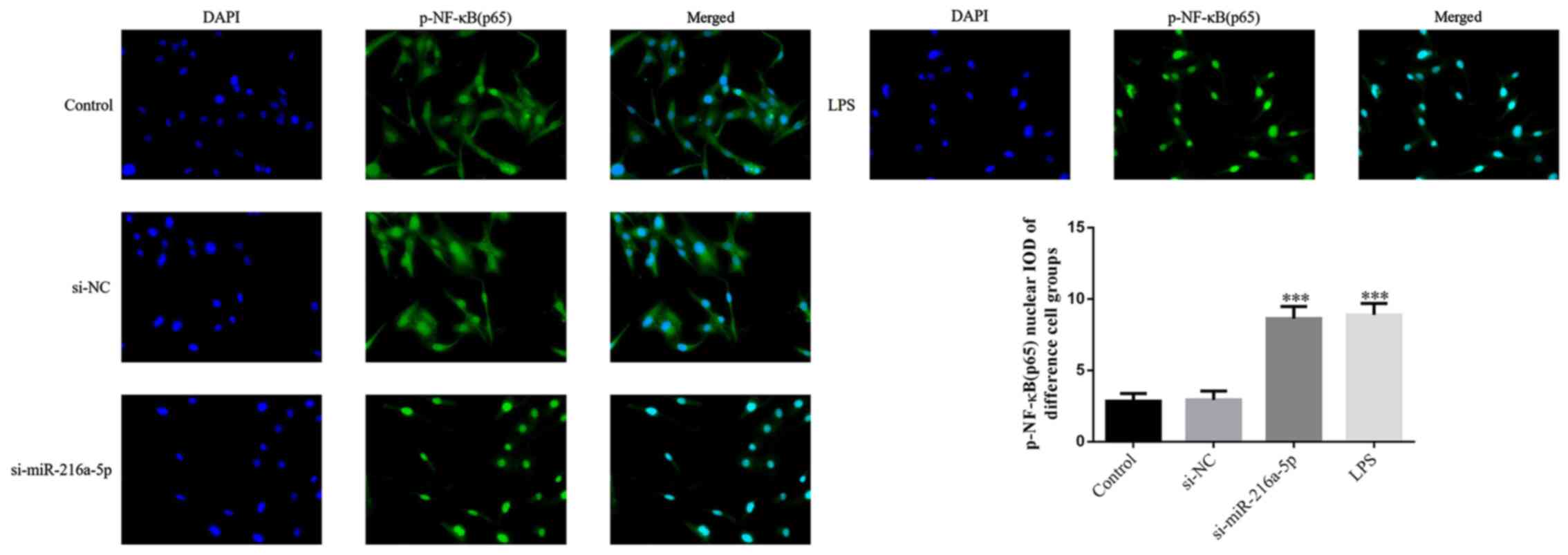 | Figure 5Effect of miR-216a-5p knockdown and
LPS intervention on the protein transportation of p-NF-κB(p65) IOD
to the nucleus. Control, cells were cultured under normal
conditions; si-NC, cells were transfected with si-NC;
si-miR-216a-5p, cells were transfected with si-miR-216a-5p; LPS,
cells were treated with LPS (1.0 mg/l). Magnification, x200.
***P<0.001 vs. control. miR, microRNA; LPS,
lipopolysaccharide; si, small interfering RNA; NC, negative
control; p-NF-κB, phosphorylated nuclear factor-κB. |
miR-216a-5p reverses the effect of LPS
in inhibiting HUVEC proliferation
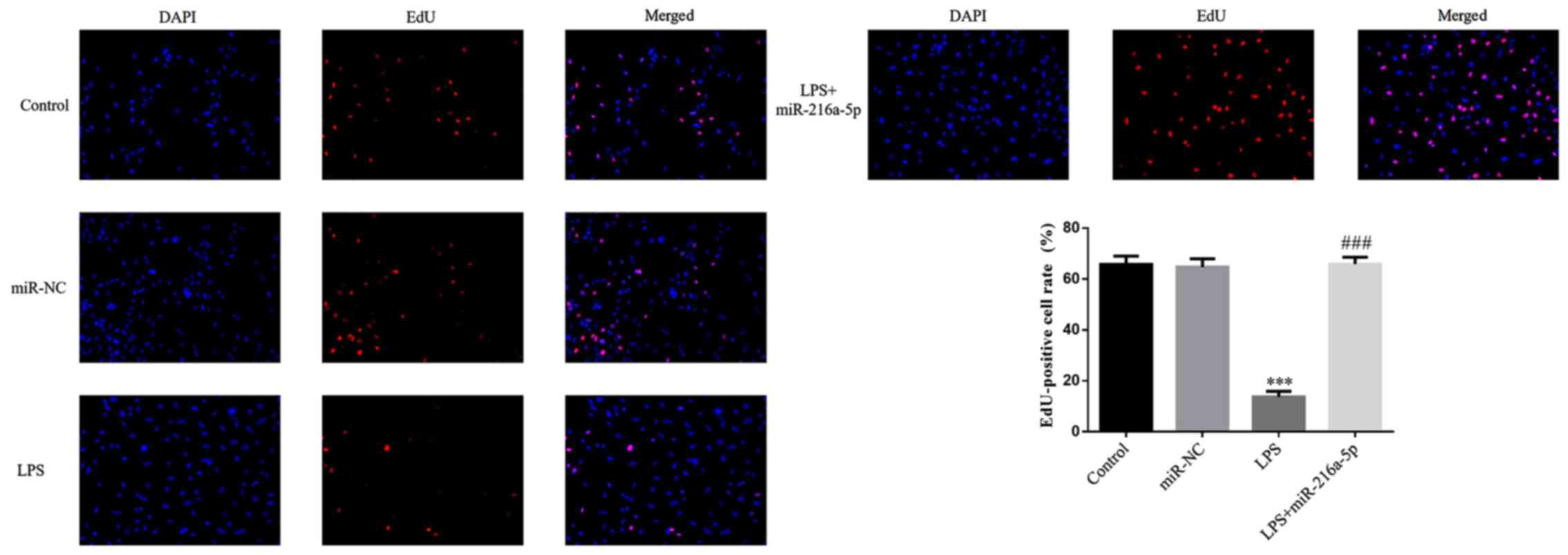
In the present study, plasmids were used for
transfection, and the results (Fig.
S1) showed that the plasmid transfection rate of miR-NC
(78.82±2.67%) and miR-216a-5p (79.17±2.82%) were higher compared
with the control group. RT-qPCR demonstrated that the miR-216a-5p
mRNA expression level of the miR-216a-5p group was significantly
upregulated compared with the control group, (P<0.001, Fig. S2). No significant difference in the
number of EdU-positive cells was observed between the miR-NC and
control groups (P>0.05; Fig. 6).
The number of EdU-positive cells was significantly decreased
following treatment with LPS (P<0.001; Fig. 6), suggesting that LPS intervention
decreases the proliferation of HUVECs. The number of EdU-positive
cells was significantly increased in the LPS + miR-216a-5p group
compared with the LPS group (P<0.001; Fig. 6).
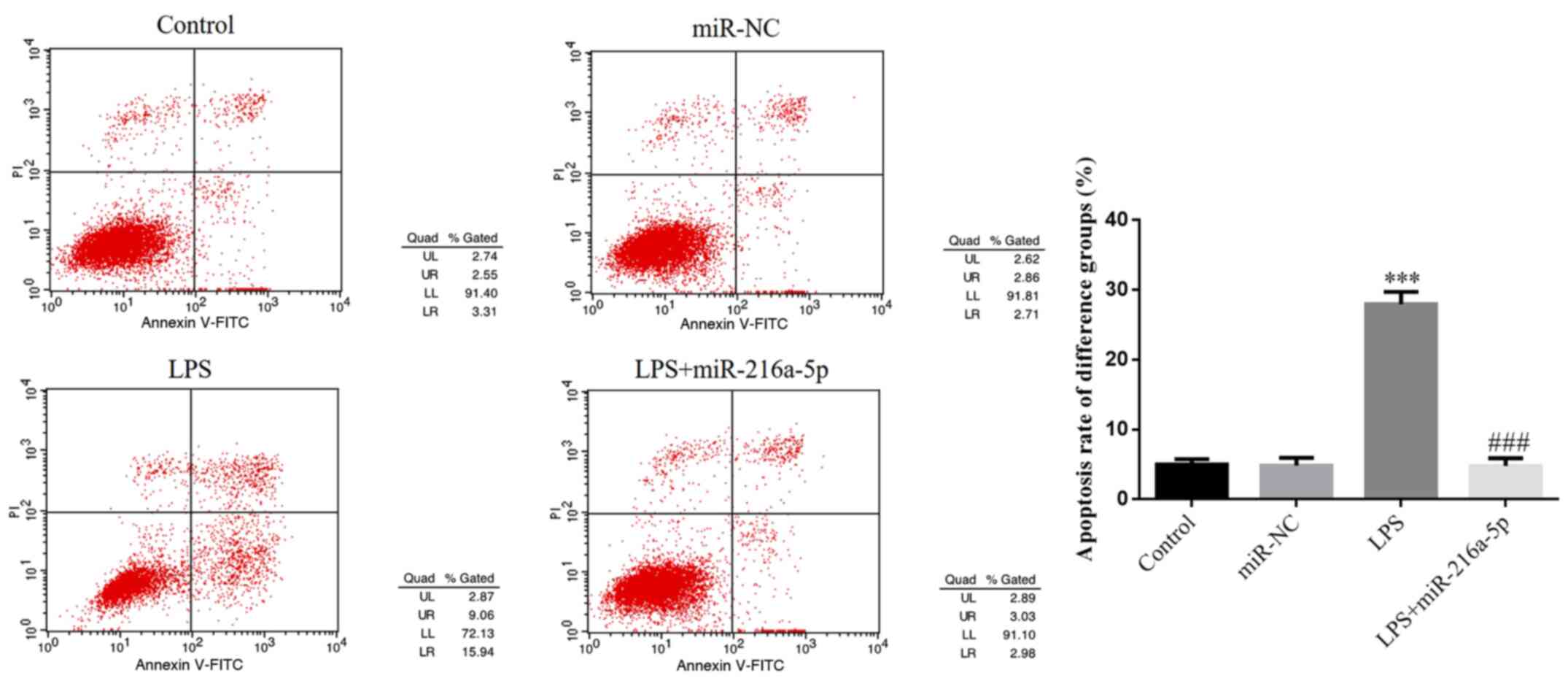
miR-216a-5p reverses the effect of LPS
in inducing HUVEC apoptosis
There was no significant difference in cell
apoptosis between the control and miR-NC groups (P>0.05;
Fig. 7), suggesting that
transfection with miR-NC causes no injury to HUVECs. Notably, cell
apoptosis was decreased in the LPS group (P<0.001; Fig. 7), suggesting that LPS intervention
promotes the apoptosis of HUVECs. Cell apoptosis was significantly
decreased in the LPS + miR-216a-5p group compared with the LPS
group (P<0.001; Fig. 7).
Effect of miR-216a-5p on gene
expression induced by LPS
No significant differences in the expression levels
of miR-216a-5p, TLR4, MyD88 and NF-κB(p65) were observed between
the control and miR-NC groups (P>0.05; Fig. 8). miR-216a-5p expression was
decreased, while the expression levels of TLR4, MyD88 and
NF-κB(p65) were significantly increased in the LPS group
(P<0.001; Fig. 8). The
aforementioned results suggest that LPS intervention affects the
expression levels of miR-216a-5p, TLR4, MyD88 and NF-κB(p65).
miR-216a-5p expression was significantly increased, while the
expression levels of TLR4, MyD88 and NF-κB(p65) were significantly
decreased following transfection of HUVECs with miR-216a-5p
compared with the LPS group (P<0.001; Fig. 8).
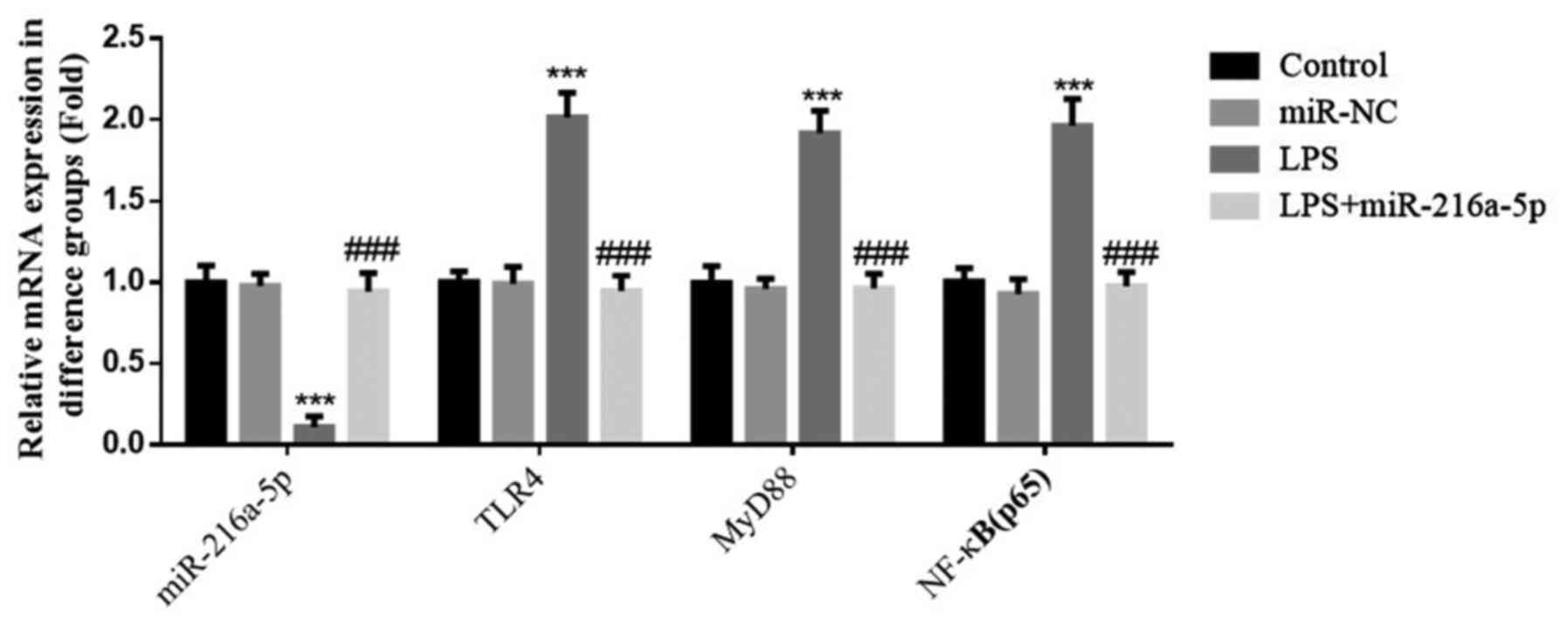 | Figure 8Effect of miR-216a-5p on gene
expression induced by LPS. Control, cells were cultured under
normal conditions; miR-NC: Cells were transfected with miR-NC; LPS,
cells were treated with 1.0 mg/l LPS; LPS+miR-216a-5p, cells
transfected with miR-216a-5p were treated with 1.0 mg/l LPS.
***P<0.001 vs. control; ###P<0.001 vs.
the LPS group. miR, microRNA; LPS, lipopolysaccharide; HUVECs,
human umbilical vein endothelial cells; NC, negative control; TLR4,
Toll-like receptor 4; p-, phosphorylated; NF-κB, nuclear
factor-κB. |
Effect of miR-216a-5p on protein
expression induced by LPS
No significant differences were observed in the
protein expression levels of TLR4, MyD88 and p-NF-κB(p65) between
the miR-NC and control groups (P>0.05; Fig. 9). Furthermore, the protein
expression levels of TLR4, MyD88 and p-NF-κB(p65) were
significantly increased in the LPS group (P<0.001; Fig. 9), suggesting that LPS intervention
affects the protein expression levels of TLR4, MyD88 and
NF-κB(p65). Notably, the protein expression levels of TLR4, MyD88
and p-NF-κB(p65) were significantly decreased in the LPS +
miR-216a-5p compared with the LPS group (P<0.001; Fig. 9). In addition, no significant
difference in the protein expression of NF-κB(p65) was observed
between the groups.
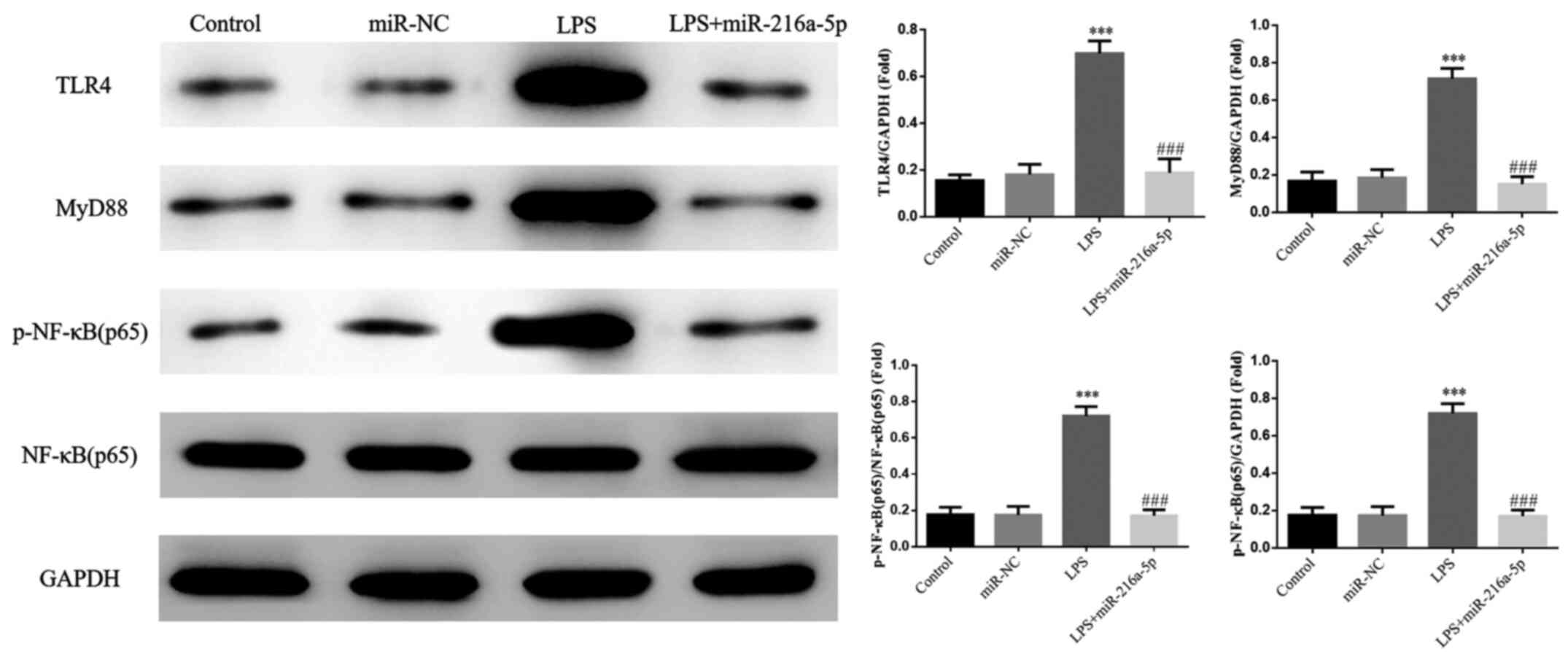 | Figure 9Effect of miR-216a-5p on protein
expression induced by LPS. Control, cells were cultured under
normal conditions; miR-NC, cells were transfected with miR-NC; LPS,
cells were treated with 1.0 mg/l LPS; LPS+miR-216a-5p, cells
transfected with miR-216a-5p were treated with 1.0 mg/l LPS.
***P<0.001 vs. control; ###P<0.001 vs.
the LPS group. miR, microRNA; LPS, lipopolysaccharide; NC, negative
control; TLR4, Toll-like receptor 4; p-, phosphorylated; NF-κB,
nuclear factor-κB. |
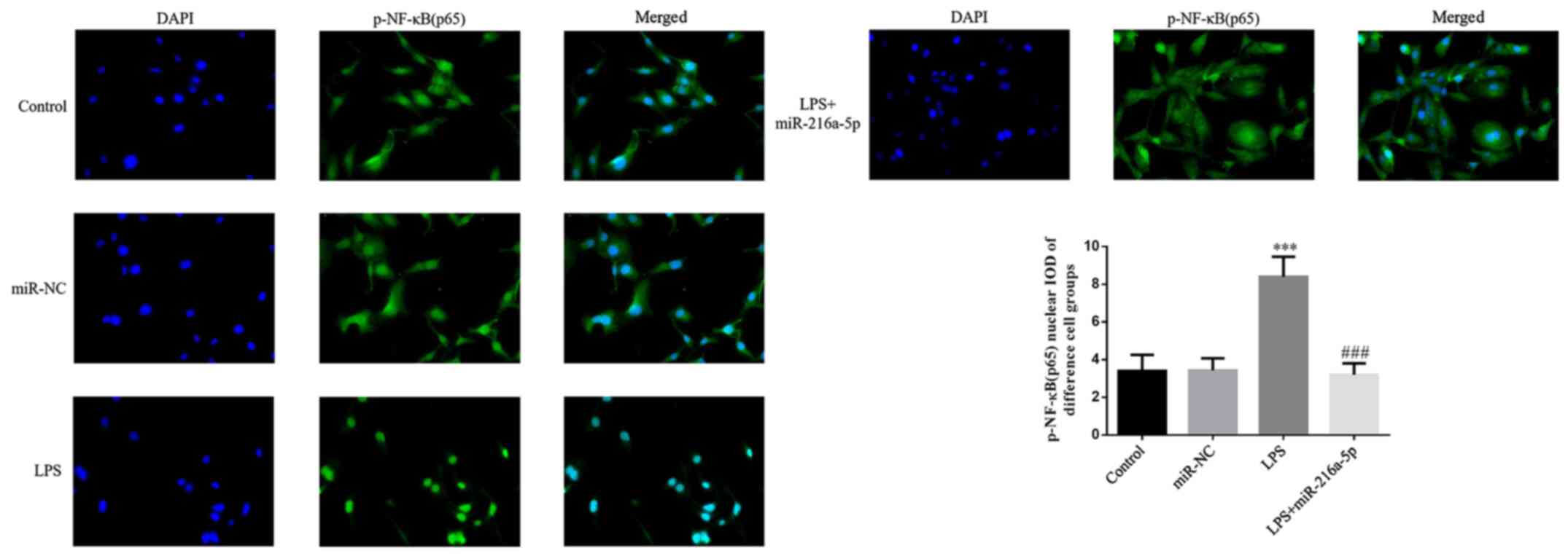
Effect of miR-216a-5p on the protein
transportation of p-NF-κB(p65) to the nucleus induced by LPS
No significant difference in protein transportation
of p-NF-κB(p65) to the nucleus was observed between the miR-NC and
control groups (P>0.05; Fig.
10). Treatment with LPS significantly increased the protein
transportation of p-NF-κB(p65) to the nucleus (P<0.001; Fig. 10), suggesting that LPS affects the
protein transportation of p-NF-κB(p65) to the nucleus. In addition,
protein transportation of p-NF-κB(p65) to the nucleus significantly
decreased in the LPS + miR-216a-5p group compared with the LPS
group (P<0.001; Fig. 10).
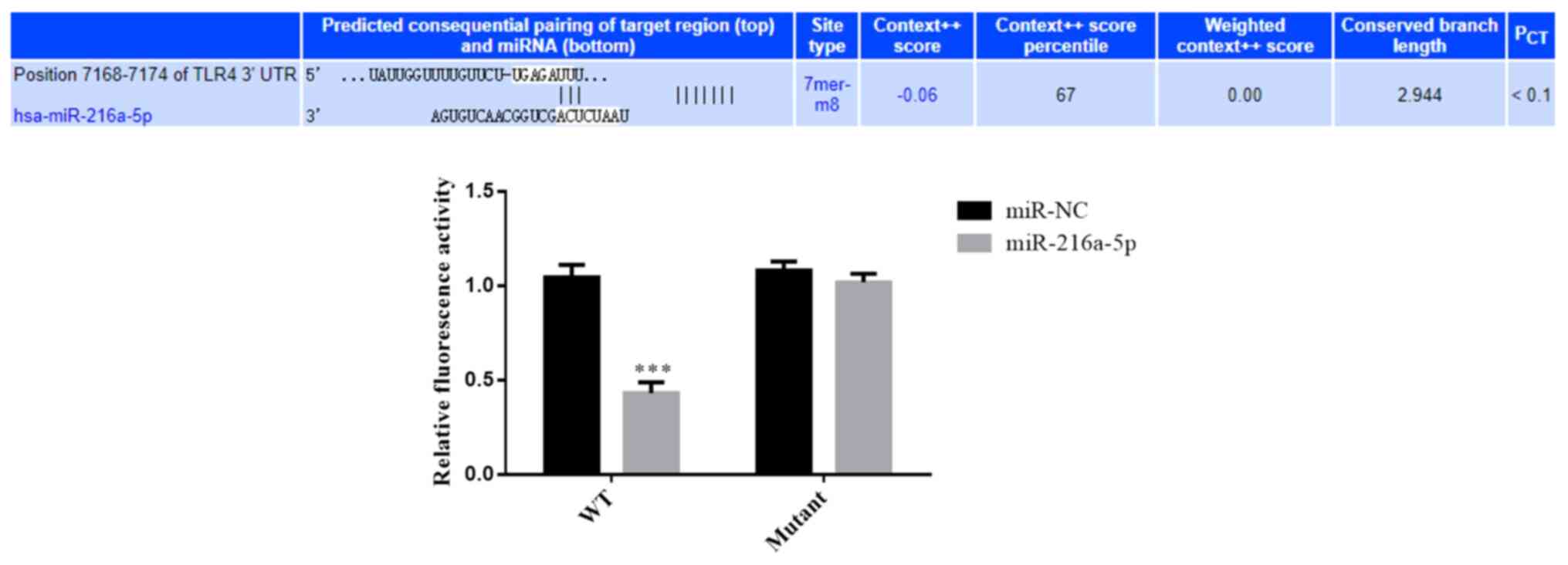
Association between miR-216a-5p and
TLR4
According to the results of the dual-luciferase
reporter assay, the relative fluorescence value was lower in cells
transfected with miR-216a-5p and WT reporter plasmids, and the
difference was statistically significant compared with the other
groups (P<0.001; Fig. 11). The
aforementioned results suggest that miR-216a-5p can bind to the
3'-UTR of TLR4 and target this gene.
Discussion
The results of the present study demonstrated that
miR-216a-5p significantly promoted the apoptosis of HUVECs.
Furthermore, miR-216a-5p knockdown notably inhibited the
proliferation, and significantly promoted the apoptosis of HUVECs,
similar to the results following treatment with LPS. Taken
together, the results suggest that miR-216a-5p may play a role in
LPS-induced injury of HUVECs. It has been reported that miRNAs may
have crucial effect on LPS-induced cell injury (14). According to the present study,
transfection of HUVECs with miR-216a-5p overexpression vector
reversed the induced apoptosis and inhibited proliferation induced
by LPS. The expression levels of the corresponding genes and
proteins were detected, and TLR4/NF-κB(p65) was identified as
playing a key role in this process.
TLRs are key components of the innate immune system,
which play a key role in the pathogenesis of inflammation. It has
been reported that TLR-mediated signaling pathways exhibit a close
association with the development of diabetes mellitus, diseases
belonging to the cardiovascular and nervous systems, and disorders
of the liver and kidney (15-17).
Simultaneously, TLRs are involved in the process of multiple organ
injury caused by inflammation (18,19).
For example, TLRs can interact with their associated signaling
molecules to activate the expression of cytokines, thus,
participating in renal injury caused by immune response (20). Overactivation of TLRs may induce
injury of the organism (21). In
addition, TLRs participate in the MyD88-dependent signaling pathway
and activate NF-κB(p65), resulting in the release of inflammatory
factors (22). Organisms under high
oxidative stress and strong inflammatory reaction may experience
the generation and release of several oxygen free radicals and
inflammatory factors, further stimulating protein transportation of
p-NF-κB(p65) to the nucleus, and thus inducing serious organ damage
(23,24). TLR4 can activate NF-κB(p65) via
activation of the MyD88-dependent signaling pathway, which triggers
the activation of inflammatory cytokines and results in apoptosis
(25). In the present study,
treatment with LPS increased the expression levels of TLR4, MyD88
and NF-κB(p65), resulting in aggravated cell injury. Following
simultaneous transfection with miR-216a-5p under the same
condition, the protein expression levels of MyD88 and p-NF-κB(p65)
decreased, which in turn decreased cell apoptosis and increased
cell proliferation. It was suggested that the decreased
proliferation and increased apoptosis of HUVECs induced by LPS
intervention can be attributed to the downregulated miR-216a-5p
expression. Simultaneous transfection with miR-216a-5p notably
recovered HUVECs proliferation and decreased cell apoptosis.
In conclusion, overexpression of miR-216a-5p may
exert a positive role in alleviating LPS-induced vascular
endothelial injury by regulating the TLR4/MyD88/NF-κB(p65)
signaling pathway.
Supplementary Material
Transfection efficiency. Cells were
transfected with control, miR-NC or miR-216a-5p mimics. miR,
microRNA; NC, negative control.
miR-216a-5p expression in different
cell groups. Cells were transfected with control, miR-NC or
miR-216a-5p mimics. ***P<0.001 vs. control. miR,
microRNA; NC, negative control.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural Science
Foundation of China (grant no. 81760339), the Ningxia Natural
Science Foundation of China (grant no. 2020AAC03331) and the Fourth
Batch of Ningxia Youth Talents Supporting Program (grant no.
TJGC2019087).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
WL and QY conceived the study and established the
initial design of the study. WL, WX, YL, KH, XZ and YW performed
the experiments and analyzed the data. WL prepared the manuscript.
WL and QY confirmed the authenticity of all the raw data. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patients consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Li FP, Lin DQ and Gao LY: LncRNA TUG1
promotes proliferation of vascular smooth muscle cell and
atherosclerosis through regulating miRNA-21/PTEN axis. Eur Rev Med
Pharmacol Sci. 22:7439–7447. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Bartel DP: Metazoan MicroRNAs. Cell.
173:20–51. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Lee CH, Cheng CL, Yang YH, Chao TH, Chen
JY, Liu PY, Lin CC, Chan SH, Tsai LM, Chen JH, et al: Trends in the
incidence and management of acute myocardial infarction from 1999
to 2008: Get with the guidelines performance measures in Taiwan. J
Am Heart Assoc. 3(e001066)2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Jing R, Zhong QQ, Long TY, Pan W and Qian
ZX: Downregulated miRNA-26a-5p induces the apoptosis of endothelial
cells in coronary heart disease by inhibiting PI3K/AKT pathway. Eur
Rev Med Pharmacol Sci. 23:4940–4947. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Qin B, Xiao B, Liang D, Xia J, Li Y and
Yang H: MicroRNAs expression in ox-LDL treated HUVECs: MiR-365
modulates apoptosis and Bcl-2 expression. Biochem Biophys Res
Commun. 410:127–133. 2011.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zhang Y, Wang L, Xu J, Kong X and Zou L:
Up-regulated miR-106b inhibits ox-LDL-induced endothelial cell
apoptosis in atherosclerosis. Braz J Med Biol Res.
53(e8960)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zeng X, Liu Y, Zhu H, Chen D and Hu W:
Downregulation of miR-216a-5p by long noncoding RNA PVT1 suppresses
colorectal cancer progression via modulation of YBX1 expression.
Cancer Manag Res. 11:6981–6993. 2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Chen P, Quan J, Jin L, Lin C, Xu W, Xu J,
Guan X, Chen Z, Ni L, Yang S, et al: miR-216a-5p acts as an
oncogene in renal cell carcinoma. Exp Ther Med. 15:4039–4046.
2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhang Y, Lin P, Zou J-Y, Zou G, Wang WZ,
Liu YL, Zhao HW and Fang AP: MiR-216a-5p act as a tumor suppressor,
regulating the cell proliferation and metastasis by targeting PAK2
in breast cancer. Eur Rev Med Pharmacol Sci. 23:2469–2475.
2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zhang J, Gao S, Zhang Y, Yi H, Xu M, Xu J,
Liu H, Ding Z, He H, Wang H, et al: MiR-216a-5p inhibits
tumorigenesis in pancreatic cancer by targeting TPT1/mTORC1 and is
mediated by LINC01133. Int J Biol Sci. 16:2612–2627.
2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Chaoyang Y, Qingfeng B and Jinxing F:
MiR-216a-5p protects 16HBE cells from H2O2-induced oxidative stress
through targeting HMGB1/NF-kB pathway. Biochem Biophys Res Commun.
508:416–420. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Moschos SA, Williams AE, Perry MM, Birrell
MA, Belvisi MG and Lindsay MA: Expression profiling in vivo
demonstrates rapid changes in lung microRNA levels following
lipopolysaccharide-induced inflammation but not in the
anti-inflammatory action of glucocorticoids. BMC Genomics.
8(240)2007.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Sharma S, Garg I and Ashraf MZ: TLR
signalling and association of TLR polymorphism with cardiovascular
diseases. Vascul Pharmacol. 87:30–37. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Leitner GR, Wenzel TJ, Marshall N, Gates
EJ and Klegeris A: Targeting toll-like receptor 4 to modulate
neuroinflammation in central nervous system disorders. Expert Opin
Ther Targets. 23:865–882. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Volarevic V, Markovic BS, Jankovic MG,
Djokovic B, Jovicic N, Harrell CR, Fellabaum C, Djonov V,
Arsenijevic N and Lukic ML: Galectin 3 protects from
cisplatin-induced acute kidney injury by promoting TLR-2-dependent
activation of IDO1/Kynurenine pathway in renal DCs. Theranostics.
9:5976–6001. 2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kasimsetty SG and McKay DB: Ischemia as a
factor affecting innate immune responses in kidney transplantation.
Curr Opin Nephrol Hypertens. 25:3–11. 2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Dominguez-Villar M, Gautron AS, de Marcken
M, Keller MJ and Hafler DA: TLR7 induces anergy in human CD4(+) T
cells. Nat Immunol. 16:118–128. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
20
|
Mancini F, Rossi O, Necchi F and Micoli F:
OMV vaccines and the role of TLR agonists in immune response. Int J
Mol Sci. 21(4416)2020.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Mohamed FE, Al-Jehani RM, Minogue SS,
Andreola F, Winstanley A, Olde Damink SW, Habtesion A, Malagó M,
Davies N, Luong TV, et al: Effect of toll-like receptor 7 and 9
targeted therapy to prevent the development of hepatocellular
carcinoma. Liver Int. 35:1063–1076. 2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Su Q, Li L, Sun Y, Yang H, Ye Z and Zhao
J: Effects of the TLR4/Myd88/NF-κB signaling pathway on NLRP3
Inflammasome in coronary microembolization-induced myocardial
injury. Cell Physiol Biochem. 47:1497–1508. 2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Shen X, Hu B, Xu G, Chen F, Ma R, Zhang N,
Liu J, Ma X, Zhu J, Wu Y, et al: Activation of Nrf2/HO-1 pathway by
glycogen synthase kinase-3β inhibition attenuates renal
ischemia/reperfusion injury in diabetic rats. Kidney Blood Press
Res. 42:369–378. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Neri M, Fineschi V, Di Paolo M, Pomara C,
Riezzo I, Turillazzi E and Cerretani D: Cardiac oxidative stress
and inflammatory cytokines response after myocardial infarction.
Curr Vasc Pharmacol. 13:26–36. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Zhou ZX and Sun L: Immune effects of R848:
Evidences that suggest an essential role of TLR7/8-induced, Myd88-
and NF-κB-dependent signaling in the antiviral immunity of Japanese
flounder (Paralichthys olivaceus). Dev Comp Immunol.
49:113–120. 2015.PubMed/NCBI View Article : Google Scholar
|