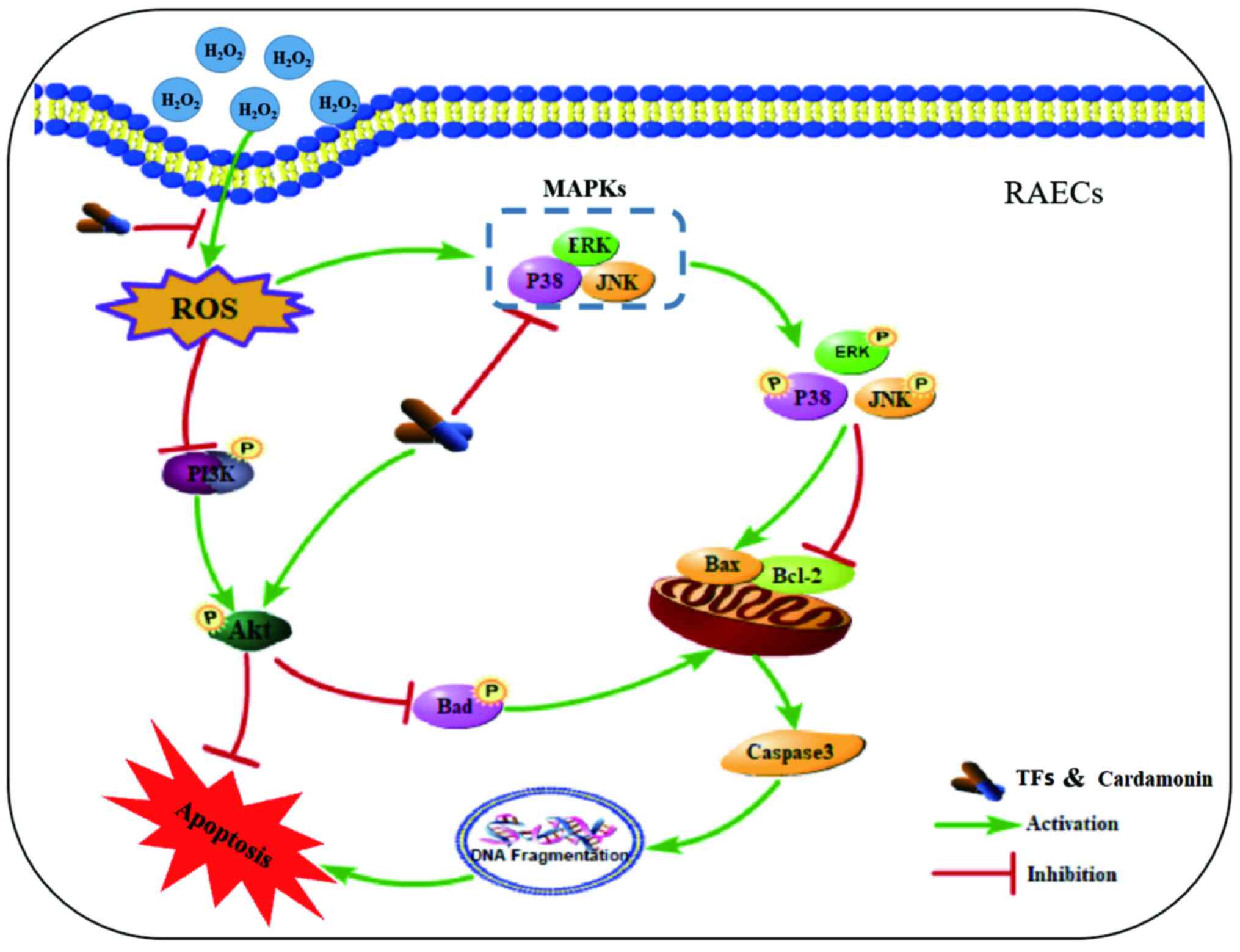
Introduction
Vascular pathologies rank amongst the most
life-threatening diseases in the world, and the cause of the
majority of these vascular pathologies is dysfunctional endothelial
cells (1). Endothelial cells cover
the inner surface of all blood vessels and play a notable role in
maintaining normal vascular function, regulating blood circulation
and exchanging substances in blood (2). Oxidative stress refers to the
cytopathological consequences when cells are exposed to a high
concentration of oxygen or chemical derivatives of oxygen (3), which are among the factors that induce
endothelial dysfunction (4).
Oxidative stress is associated with the occurrence and development
of cardiovascular diseases such as hypertension, atherosclerosis
and heart failure (5). Therefore,
protecting endothelial cells from oxidative injury and apoptosis
represents a beneficial strategy for the treatment of vascular
diseases. At present, the majority of antioxidant drugs are
chemical drugs. These chemicals have some limitations, including
varying degrees of toxicity, residual effects, deformities and
potential carcinogenicity. Therefore, it is of great clinical
significance to screen effective and safe natural oxygen free
radical scavengers as substitutes for synthetic antioxidants.
Natural products have been a rich source of
compounds for drug discovery and represent attractive alternatives
for disease prevention and treatment (6,7). To
date, numerous studies have been conducted to find effective drugs
to block oxidative damage (4,8). The
ability of various bioactive flavonoids from natural products and
traditional Chinese medicine to scavenge free radicals have been
reported (9,10). Wen et al (11) have demonstrated that flavonoids
extracted from Traditional Chinese medicine and natural plants are
broad-spectrum free radical scavengers that can effectively
eliminate free radicals in the body and play an antioxidant role
(11). Chen et al (12) identified 12 flavonoid components in
lotus plumule and demonstrated their significant antioxidant
activity. Wei et al (13)
isolated ten flavonoid derivatives from the fruits of Metaplexis
japonica, whose antibacterial and antioxidant activities were
also revealed in their study.
Carya cathayensis Sarg., a deciduous tree
belonging to the hickory family, has been widely used as a
conventional folk medicine for thousands of years. It was
commercially cultivated in the Zhejiang and Anhui provinces of
China (14). The effectiveness of
the anticancer, free radical scavenging, cardiotonic,
anti-inflammatory and antinociceptive actions of the husk and
kernel of Carya cathayensis Sarg. was reported (15); however, the value of the leaves of
Carya cathayensis Sarg. (LCCS) remains sparsely
investigated. In our previous study, total flavonoids (TFs) were
demonstrated to be abundant in the LCCS (16), and five flavonoid monomers,
cardamonin (Car), pinostrobin chalcone (PC), wogonin (Wo), chrysin
(Chr) and pinocembrin (Pin) (Fig.
1) were separated from the TFs (17). In addition, the pharmaceutical
properties of TFs, for example, anti-inflammatory activity
(12), antitumor activity (18), anti-early atherosclerosis lesion
formation in vivo (19) and
anti-human umbilical vein endothelial cell senescence (13) have been investigated. However, the
activities of flavonoids from LCCS against
H2O2-induced oxidative damage in vitro
remain poorly investigated.
The present study aimed to investigate the potential
antioxidant effects of TFs and flavonoid monomers from LCCS on
H2O2-induced oxidative damage in rat aortic
endothelial cells (RAECs). Fig. 2
describes the mechanisms by which the TFs and Car from LCCS protect
against H2O2-induced apoptosis.
Materials and methods
Materials and regents
LCCS was obtained in 2016 from Lin'an (Zhejiang,
China). The plants were authenticated by Professor Zhishan Ding
from Zhejiang Chinese Medical University, Hangzhou, Zhejiang,China.
The voucher specimens were deposited in the laboratory center of
the Medical Technology College at Zhejiang Chinese Medical
University (Hangzhou, China; voucher no. LCC-20160915-G). The TFs
were extracted from LCCS according to the method described in our
previous study (17). Analytical
grade H2O2 was purchased from Sinopharm
Chemical Reagent Co., Ltd. PC and Pin were further separated and
purified from TFs. Car, Wo and Chr were obtained from Shanghai
Yuanye Biotechnology, Co., Ltd. and Beijing Aoke Biological
Technology Co., Ltd. Malondialdehyde (MDA; cat. no. A003-4-1),
superoxide dismutase (SOD; cat. no. A001-3-2), lactate
dehydrogenase (LDH; cat. no. A020-2-2) and glutathione peroxidase
(GSH-Px; cat. no. A005-1-2) assay kits were all purchased from
Nanjing Jiancheng Bioengineering Institute. A FITC Annexin V
apoptosis detection kit was purchased from BD Biosciences (cat. no.
556547). Antibodies against caspase-3 (cat. no. YM3431), Bax (cat.
no. YT0455), Bcl-2 (cat. no. YT0470), β-actin (cat. no. YM3028),
Akt (cat. no. YT6111), phosphorylated (p)-Akt (cat. no. YP0590),
ERK (cat. no. YT1624), p-ERK (cat. no. YP0101), JNK (cat. no.
YT2441), p-JNK (Thr183/Tyr185; cat. no. YP0157), p38 (cat. no.
YT3514) and p-p38 MAPK (Thr180/Tyr182; cat. no. YP0338) were
supplied by ImmunoWay Biotechnology Company. Horseradish
peroxidase-conjugated goat anti-rabbit IgG (cat. no. abs20002) and
goat anti-mouse IgG (cat. no. abs20003) were purchased from Aibixin
(Shanghai) Biotechnology Co., Ltd. An MTS assay kit (cat. no.
G3580) with
3-(4,5-dimethylthiazoll-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium
reagent and an inner salt was purchased from Promega Corporation. A
First Strand complementary (c)DNA Synthesis kit was used (Takara
Bio, Inc.) and SYBR® Premix Ex Taq™ II were purchased
from Takara Bio, Inc.
Cell culture
The RAECs were purchased from Yingwan Biotechnology
Co., Ltd. (cat. no. C3048) The RAECs were cultured in DMEM
supplemented with 10% heat-inactivated FBS (cat. no. 70220-8611;
Zhejiang Tianhang Biotechnology Co., Ltd.), 100 mg/ml streptomycin
and 100 U/ml penicillin at 37˚C in a humidified 5% CO2
incubator.
Determination of TFs and the
concentrations of five flavonoids using MTS assay
The effects of TFs and five flavonoids (Car, PC,
Won, Chr and Pin) on the proliferation of RAEC cell lines were
evaluated using the MTS assay. Briefly, cells were seeded in
96-well plates at a density of 1.5x105 cells/well in
phenol-free red DMEM/F12 (1:1) medium with 10% CS-FBS purchased
from Biosharp Biotechnology Co., Ltd (cat. no. BL305A). After
incubation at 37˚C for 24 h, the cells were treated with TFs at 0,
5, 10, 20, 40 and 80 µg/ml, or the five flavonoids (Car, PC, Won,
Chr and Pin) at 0, 5, 10, 20, 40 and 80 µM for 24 h. Subsequently,
20 µl of MTS solution was added to each well and incubated for an
additional 2 h at 37˚C in a CO2 incubator. The
absorbance values were detected by a microplate reader (Dynatech
Nevada, Inc.) at 490 nm. The average cell activity was calculated
according to the following formula: Percentage of cell viability =
[optical density (OD) of TFs and five flavonoid cells/OD of control
cells] x100. The oxidative stress injury model induced by
H2O2 was established using the MTS assay.
Briefly, RAECs were cultured in a 96-well microtiter plate at a
density of 1.5x105 cells/well at 37˚C for 24 h. After
that, RAECs were exposed to H2O2 (0, 50, 100,
150, 200 or 400 µM) for 24 h. Subsequently, RAECs were treated with
appropriate H2O2 concentrations for 0, 2, 4,
6 and 8 h. Cell viability was measured by MTS assay as
aforementioned.
Cell viability assay
RAECs were cultured in a 96-well microtiter plate at
a density of 1.5x105 cells/well for 24 h, and the cells
were divided into the following five groups: i) Control group; ii)
model group; iii) quercetin (Que) group (20 µM); iv) TF group (5
µg/ml, 10 µg/ml, 20 µg/ml); v) five flavonoids group (5, 10 and 20
µM). Subsequently, the RAECs were treated with TFs and five
flavonoids at 37˚C for 24 h, followed by 100 µM
H2O2 treatment at 37˚C for 2 h to establish
an injury model. Control group cells were cultured with 200 µl of
complete medium. Quercetin was used as a positive control (20). Briefly, 20 µl of MTS solution was
added to each well and incubated for an additional 2 h at 37˚C in a
CO2 incubator. The absorbance values were detected by a
microplate reader (Dynatech Nevada, Inc.) at 490 nm. The morphology
of the cells was observed using a Nikon ECLIPSE Ti fluorescence
inverted microscope (magnification, x20; Nikon Instruments Inc.),
and images were captured.
Determination of intracellular MDA,
LDH, SOD, and GSH-Px
To assess the effects of TFs and Car on oxidative
injury in RAECs, RAECs were plated in a six-well plate at a density
of 1.5x105 cells/ml (2.5 ml/well) and cultured as
described above. After pretreatment with TFs or Car for 24 h, cells
were exposed to H2O2 for 2 h at 37˚C. The
supernatant of each group was collected to evaluate the LDH and MDA
levels. The cells in each group were digested and collected with
0.25% trypsin, and the cell homogenate was obtained by ultrasonic
crushing. The adherent cells were lysed and then centrifuged at 4˚C
and 12,000 x g for 15 min to collect the supernatant. The
activities of SOD and GSH-Px were measured according to the
manufacturer's instructions for the assay kits.
Apoptosis analysis by flow
cytometry
An Annexin V-FITC detection kit was used to
determine the effects of TFs and Car on apoptosis induced by
H2O2. Briefly, cells were seeded into a
six-well plate and treated as described above. After harvesting by
trypsin, the cells were washed twice with precooled PBS,
centrifuged at 4˚C and 500 x g/min for 5 min to remove the cell
debris, and then re-suspended in 1X binding buffer. A 100 µl cell
suspension was placed in a 5 ml flow tube, and the cells were
stained with FITC Annexin V and propidium iodide (PI) for 15 min at
room temperature. Cells were quantified using flow cytometry (BD
Biosciences; Accuri C6 ) within 1 h. The percentage distributions
of normal (viable), early apoptotic, late apoptotic and necrotic
cells were calculated using Summit software (FCS Express 5; version
3.0).
Reverse transcription-quantitative PCR
(RT-qPCR)
RAECs were treated with 5, 10 and 20 µg/ml of TFs,
and 5, 10 and 20 µM of Car for 24 h, followed by 100 µM
H2O2 treatment for 2 h to establish an injury
model. Total cell RNA was extracted using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. The instructions of the First Strand
cDNA Synthesis kit were followed to synthesize cDNA by RT-PCR. SYBR
Green real-time fluorescent quantitative PCR was used to detect the
mRNA expression levels of caspase-3, Bax and Bcl-2 in each group of
cells, and the reactions were carried out using Step One Plus
Real-Time PCR (Applied Biosystems; Thermo Fisher Scientific, Inc.).
The reaction system was SYBR Premix EX Taq™ II 10 µl, upstream
primer 0.5 µl, downstream primer 0.5 µl, cDNA 0.5 µl and
ddH2O 8.5 µl. The reaction condition were 95˚C for 5 min
and then 40 cycles of 95˚C for 20 sec, 55˚C for 20 sec and 72˚C for
15 sec. The primers used are listed in Table I and were synthesized by Shanghai
Shenggong Biology Engineering Technology Service, Ltd. All
experimental procedures were in accordance with the manufacturer's
protocols. The expression of mRNA was analyzed by the
2-ΔΔCq method (21).
 | Table IList of primer sequences used in the
present study. |
Table I
List of primer sequences used in the
present study.
| Primer name | Sequence,
(5'-3') | Length, bp | Ta, ˚C |
|---|
| Rat GAPDH
forward | CCC ACG GCA AGT TCA
ACG GCA | 21 | 63.9 |
| Rat GAPDH
reverse | TGG CAG GTT TCT CCA
GGC GGC | 21 | 65.8 |
| Rat Caspase-3
forward | CTG GAC TGC GGT ATT
GAG | 18 | 57.3 |
| Rat Caspase-3
reverse | GGG TGC GGT AGA GTA
AGC | 18 | 59.6 |
| Rat Bax
forward | GCA AAC TGG TGC TCA
AGG | 18 | 57.3 |
| Rat Bax
reverse | TCC CGA AGT AGG AAA
GGA G | 19 | 57.6 |
| Rat Bcl-2
forward | TCT AAC ATC CCA GCT
TCA T | 19 | 53.2 |
| Rat Bcl-2
reverse | GCA ATC CGA CTC ACC
AAT A | 19 | 55.4 |
Western blotting
Total cytosolic protein of RAECs cultured with
various concentrations of TFs and Car for 24 h and injured by
H2O2 for 2 h as aforementioned. Cells were
washed twice with cold PBS and lysed by cold lysis buffer
(RIPA:PMSF=100:1) for 40 min on ice. Cell lysates were centrifuged
at 12,000 x g for 15 min at 4˚C. The protein concentration was
determined using a BCA protein assay kit according to the
manufacturer's instructions. Harvested proteins were denatured at
100˚C for 10 min, 50 µg of protein/per lane from each sample was
electrophoresed by SDS-PAGE and then transferred onto PVDF
membranes. Membranes were blocked with 5% non-fat milk in TBS with
0.5% Tween-20 at room temperature for 2 h, followed by overnight
incubation at 4˚C with primary antibodies (caspase-3, Bax, Bcl-2,
β-actin, Akt, ERK, JNK, p38 MAPK, p-ERK, p-JNK and p-p38 MAPK) at a
dilution of 1:1,000. Subsequently, the blots were incubated with
horseradish peroxidase-conjugated goat anti-rabbit IgG (1:5,000)
for 2 h at room temperature and detected using an enhanced
chemiluminescence system. The bands were quantified by densitometry
analysis with ImageJ software (National Institutes of Health;
version, 1.8.0.112).
Statistical analysis
All studies were performed with three independent
experiments. All data were analyzed using GraphPad Prism 8.0
software (GraphPad Software, Inc.). The data are expressed as the
means ± standard deviation for continuous variables. Comparisons
between groups were performed using one-way ANOVA, and normally
distributed data were analyzed using Tukey's multiple comparisons
post hoc test. Non-normally distributed data were analyzed using
Dunn's test for intergroup comparisons. P<0.05 was considered to
indicate a statistically significant difference.
Results
TFs and five monomeric flavonoids
inhibit H2O2-induced injury in RAECs
As presented in Fig.
3A, treatment with 0-400 µM H2O2 for 24 h
significantly decreased the survival rate of RAECs in a
concentration-dependent manner (P<0.01). The cell viability
decreased to 54.73±4.91% when the concentration of
H2O2 increased to 100 µM. As presented in
Fig. 3B, the cell viability
decreased to 61.83±2.73% after 2 h of treatment, which was
significantly different compared with that of the untreated group
(P<0.01). Based on these results, 100 µM
H2O2 for 2 h was selected as the treatment
condition for the following experiments.
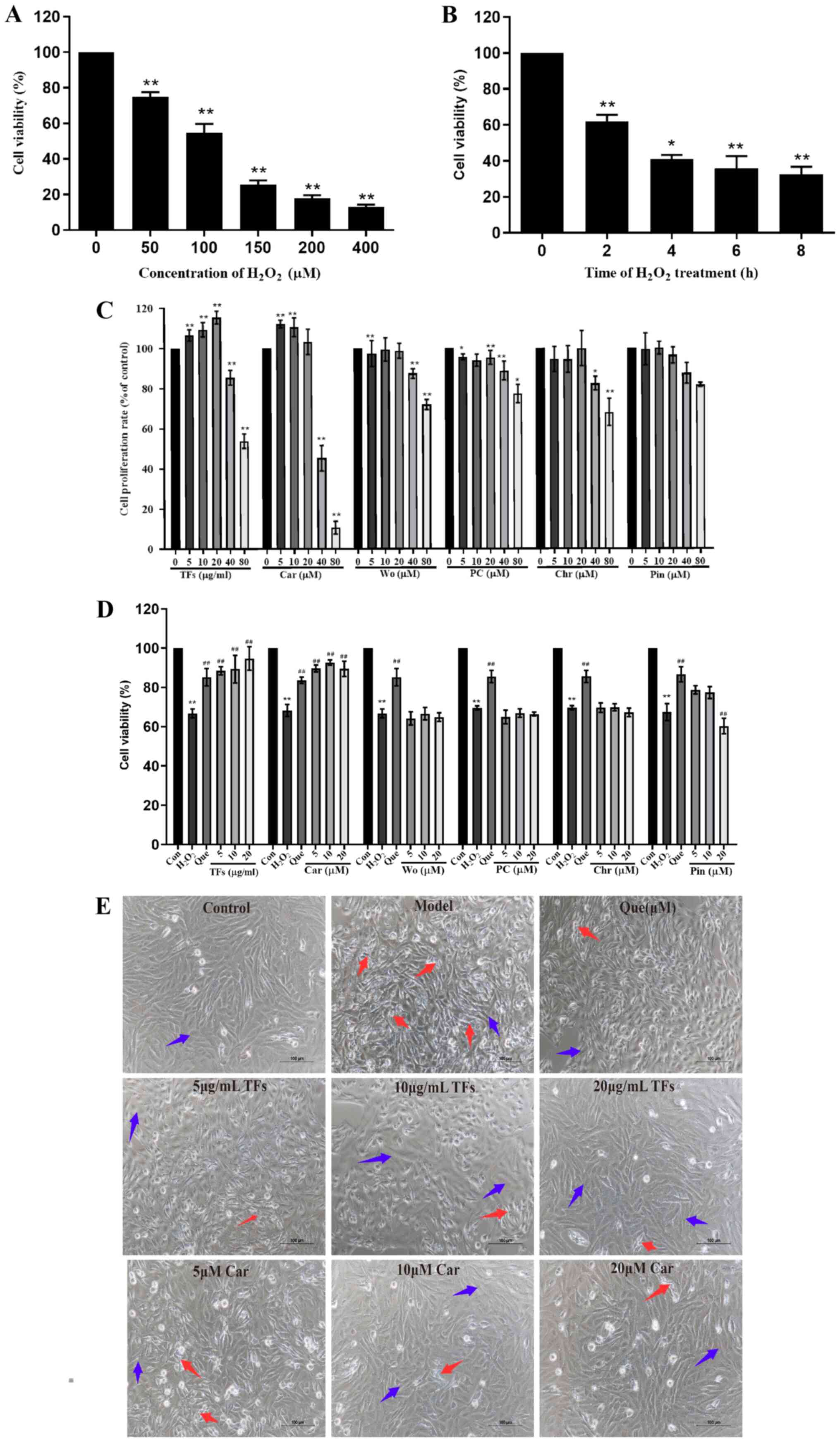 | Figure 3TFs and five monomeric flavonoids
inhibit H2O2-induced injury in RAECs. (A)
RAECs were treated with increasing concentrations of
H2O2 (0-400 µM) for 2 h and cell viability
was assessed. (B) RAECs were treated with 100 µM
H2O2 for 0-8 h. (C) Effects of increasing
concentrations of TFs and five monomeric flavonoids on the
proliferation rate of RAECs. (D) Protective effects of increasing
concentrations of TFs and five monomeric flavonoids on
H2O2-induced RAEC damage. (E) Effects of TFs
and Car on the morphology of RAECs after
H2O2-induced injury (magnification, x200;
scale bar, 100 µm) n=6. *P<0.05,
**P<0.01 vs. control group. ##P<0.01
vs. H2O2 model group. RAECs, rat aortic
endothelial cells; TFs, total flavonoids; Car, cardamonin; Wo,
wogonin; PC, pinostrobin chalcone; Chr, chrysin; Pin, pinocembrin;
Que, quercetin; Con, control. |
The results demonstrate that treatments with TFs and
five monomeric flavonoids (Car, PC, Wo, Chr and Pin) at
concentrations >40 µg/ml and 40 µM, respectively, inhibited cell
proliferation compared with the control groups (P<0.01).
Therefore, the optimal treatment concentrations for TFs (5, 10 and
20 µg/ml) and five monomeric flavonoids (5, 10 and 20 µM) were
selected for subsequent experiments (Fig. 3C).
To assess the protective effects of TFs and five
monomeric flavonoids on H2O2-induced injury,
RAECs were pretreated with TFs (5, 10 or 20 µg/ml) and five
monomeric flavonoids (5, 10 or 20 µM) for 24 h before exposure to
100 µM H2O2 for 2 h. As presented in Fig. 3D, the rate of cell viability was
significantly decreased in the model group compared with the
control group (P<0.01), and the cell viability after
pretreatment with TFs and Car was significantly higher compared
with that of the model group. TFs and Car displayed a protective
effect that was close to that of quercetin (the positive control).
However, a protective effect was not observed in cells treated with
Wo, PC, Chr and Pin. Therefore, TFs and Car were selected for
further antioxidant and antiapoptotic studies.
The morphologies of the cells were spindle shaped
with clear and bright boundaries in the control group. In the
H2O2 group, large numbers of suspended cells
and cell fragments were observed. In the cells pretreated with TFs
and Car, the shrinkage of cells decreased and few suspended cells
and cell fragments were observed. The protective effect was
dose-dependent, and its effect was similar to that of quercetin
(Fig. 3E).
Effect of TFs and Car on LDH, MDA,
GSH-Px and SOD levels
To determine the effects of TFs and Car on oxidative
stress in RAECs, the levels of LDH, MDA, GSH-Px and SOD were
measured. As presented in Fig. 4A
and B, the levels of LDH and MDA in
the supernatant of the model group were significantly increased
compared with those in the control group (P<0.01). Compared with
the model group, treatment with TFs and Car significantly reduced
the production of LDH and MDA in a dose-dependent manner
(P<0.05). In the model group, the contents of GSH-Px and SOD
were significantly reduced compared with those in the control group
(P<0.01). After pretreatment with TFs and Car, the activity of
GSH-Px and SOD was markedly increased compared with that of the
model group (Fig. 4C and D).
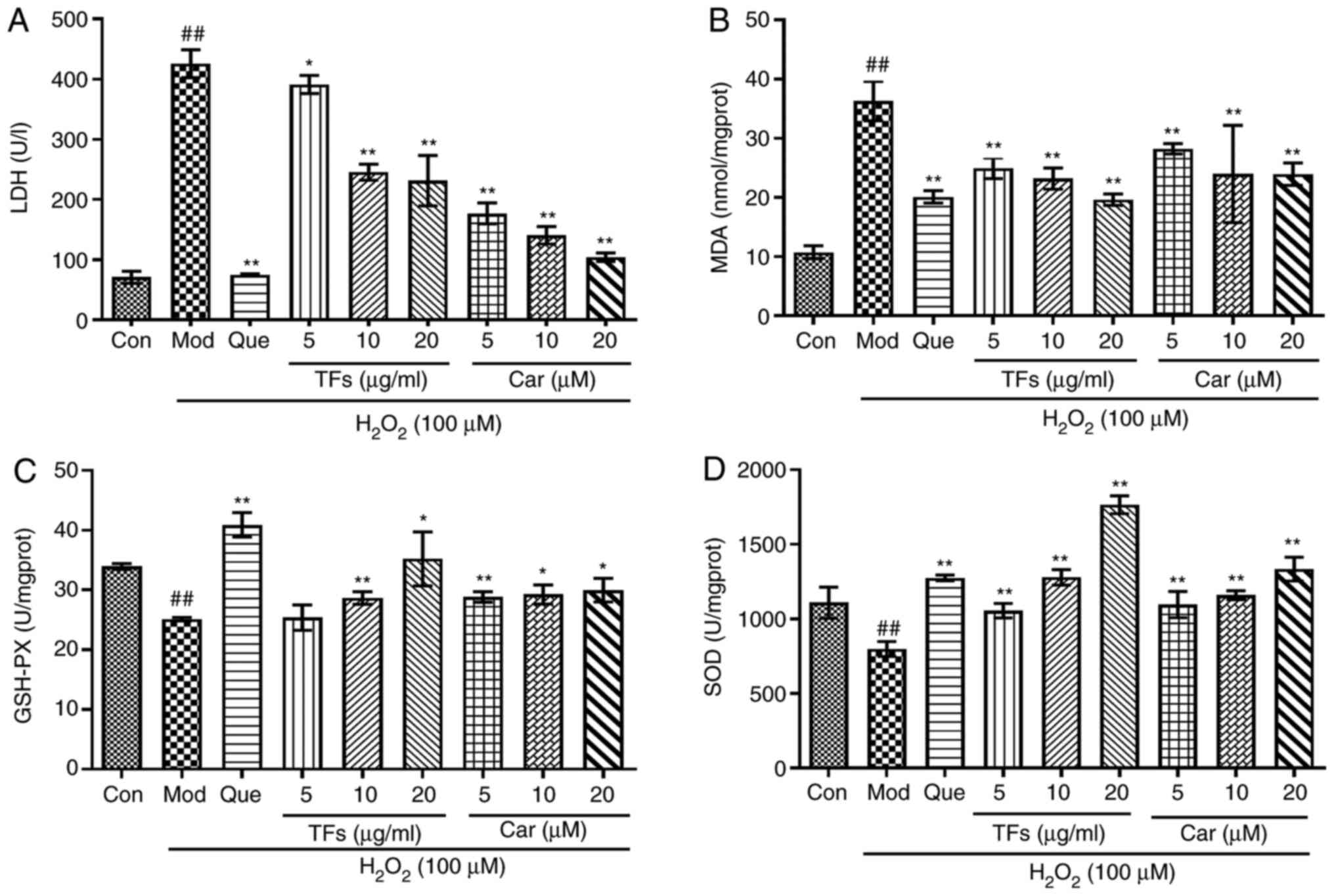 | Figure 4Protective effects of TFs and
flavonoids by modulating the activities of enzymes. (A) Level of
LDH released in RAECs. (B) Content of intracellular MDA. (C)
Activities of GSH-PX and (D) SOD. n=3. ##P<0.01 vs.
control group; *P<0.05, **P<0.01 vs.
H2O2 model group. TFs, total flavonoids; LDH,
lactate dehydrogenase; RAECs, rat aortic endothelial cells; MDA,
malondialdehyde; GSH-PX, glutathione peroxidase; SOD, super oxide
dismutase; Con, control; Mod, model; Que, quercetin; Car,
cardamonin. |
TFs and Car suppresses
H2O2-induced intracellular ROS
production
Intracellular ROS play an important role in
oxidative stress-induced cell damage. In the model group,
intracellular ROS generation was significantly increased by
>3.8-fold compared with that in the control group (P<0.01;
Fig. 5). In contrast, the relative
fluorescence intensities were reduced by 1.04±0.05, 1.36±0.05 and
1.60±0.06 in the 5, 10 and 20 µg/ml TF-pretreated groups, and by
1.26±0.04, 1.34±0.02 and 1.55±0.06 in the 5, 10 and 20 µM
CAR-pretreated groups, respectively. Therefore, pretreatment with
TFs and Car prevented the elevation of ROS levels in a
dose-dependent manner (P<0.01; Fig.
5).
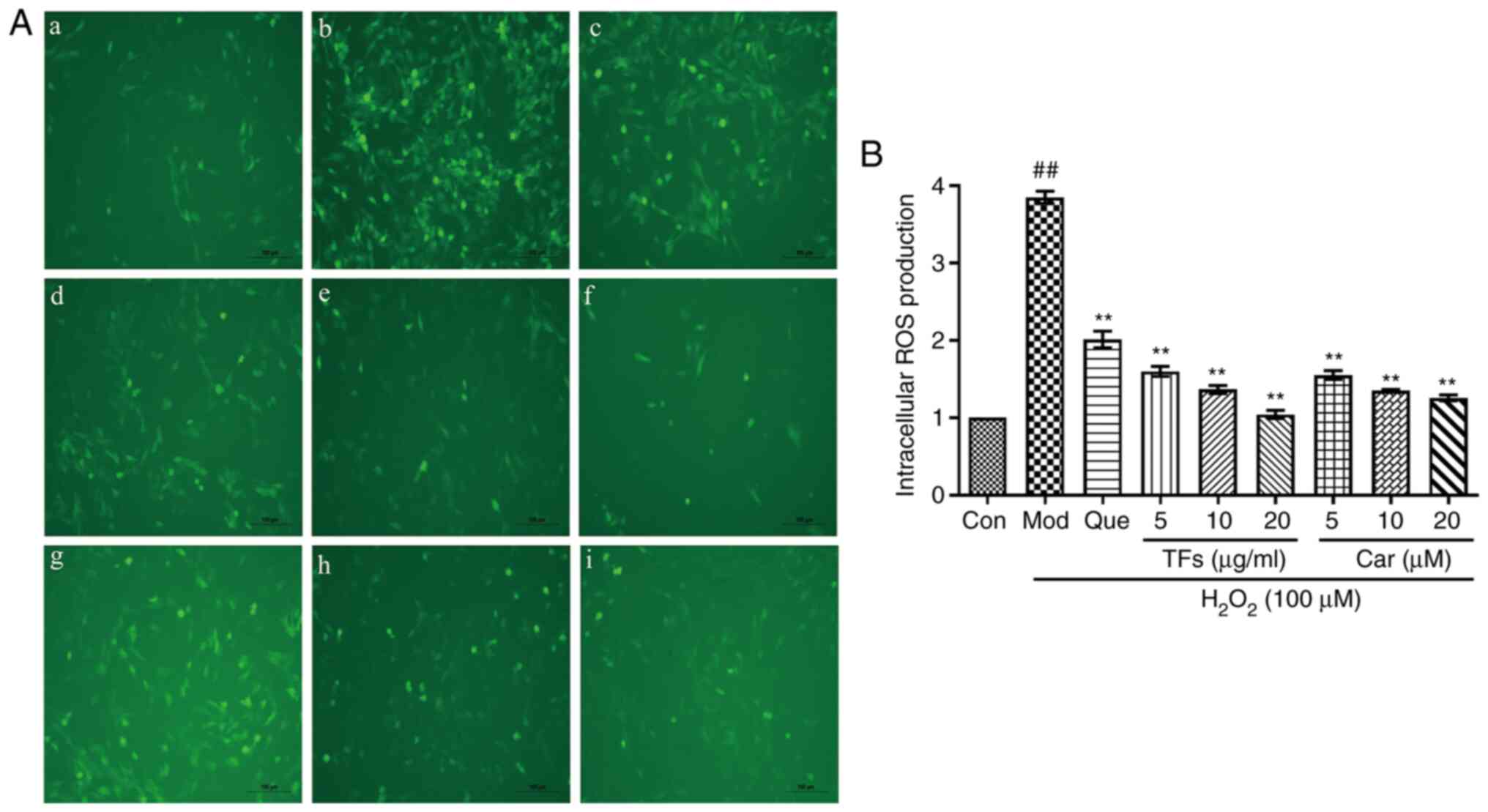 | Figure 5Protective effects of TFs and Car by
reducing intracellular ROS induced by H2O2.
(A) ROS generation was measured. Representative images of RAECs
loaded with DCFH-DA (10 µM) were captured using a fluorescence
microscope (magnification, x200). (a) Control group, (b) Model
group, (c) quercetin group, (d) 5 µg/ml TFs group, (e) 10 µg/ml TFs
group, (f) 20 µg/ml TFs group, (g) 5 µM Car group, (h) 10 µM Car
group and (i) 20 µM Car group. (B) Quantitative analysis of DCF
fluorescence intensity. n=3. ##P<0.01 vs. control
group; **P<0.01 vs. H2O2 model
group. TFs, total flavonoids; RAECs, rat aortic endothelial cells;
Con, control; Mod, model; Que, quercetin; Car, cardamonin. |
Effect of TFs and Car on apoptosis in
H2O2-induced RAECs
As presented in Fig.
6A, the apoptosis scatter plot can be divided into four
quadrants: Upper left quadrant R1 (Annexin
V-FITC)-/PI+, which refers to mechanically
injured cells or a few late apoptotic cells; upper right quadrant
R2 (Annexin V-FITC)+/PI+, for late apoptotic
cells; lower left quadrant R3 (Annexin
V-FITC)-/PI-, normal living cells; and the
lower right quadrant R4 (Annexin
V-FITC)+/PI-, which presents early apoptotic
cells. The total apoptotic cells were the sum of early apoptotic
cells and late apoptotic cells. As presented in Fig. 6B, in the normal control group, only
a few cells were apoptotic and the apoptosis rate was 9.31±1.84%.
Compared with the normal control group, large numbers of apoptotic
cells and fragments were revealed in the H2O2
model group, and the apoptosis rate was 28.71±4.41% (P<0.01).
Compared with the model group, the apoptotic rate of cells
pretreated with TFs and Car resulted in a significant decrease
(P<0.01). These results indicated that TFs and Car could inhibit
the cell damage and apoptosis induced by
H2O2.
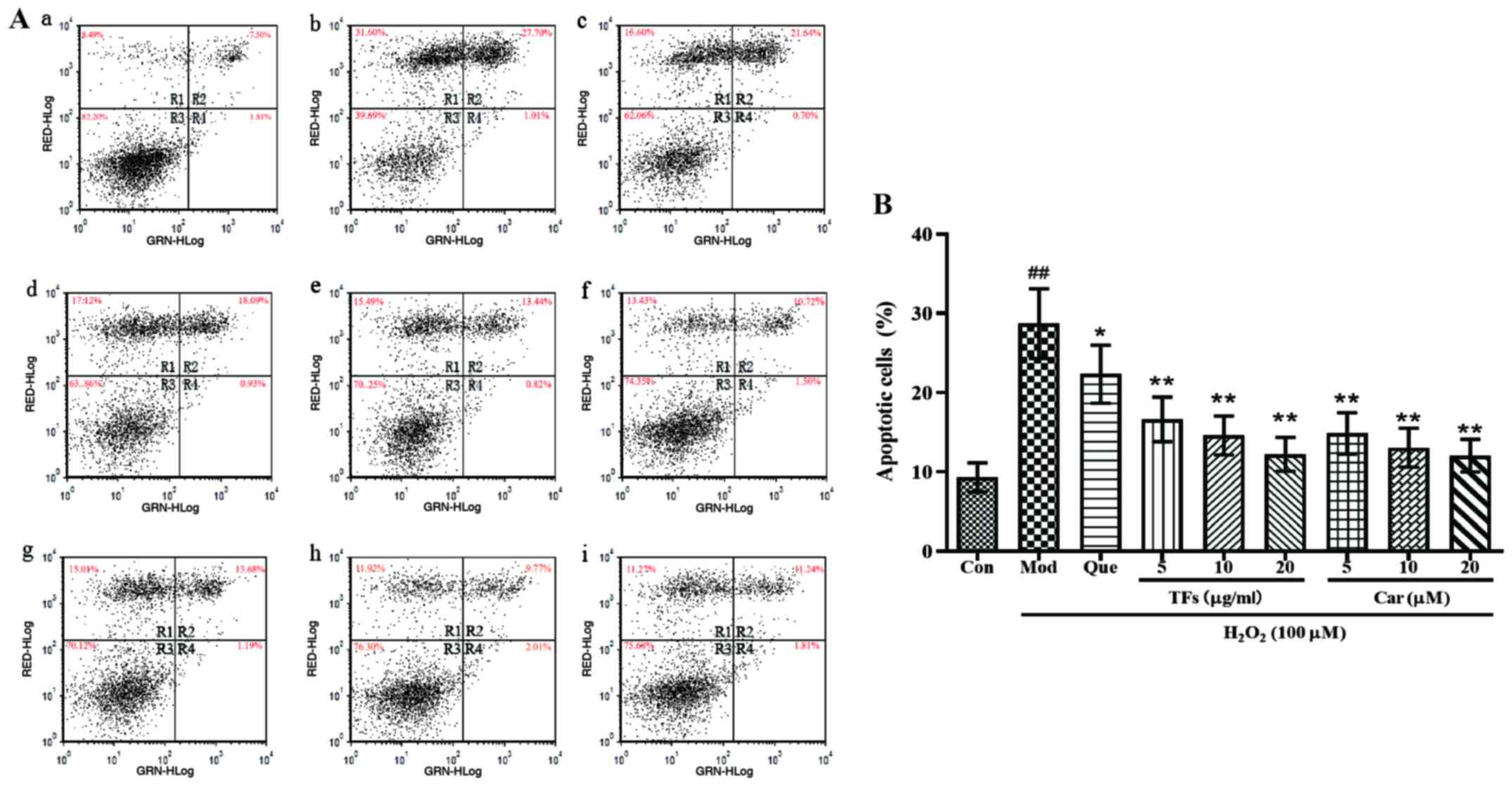 | Figure 6Effects of TFs and Car on
H2O2-induced RAEC apoptosis. (A) Apoptosis
was examined using Annexin V/PI staining followed by flow cytometry.
R1, PI-positive cells (necrotic); R2, Annexin V-FITC-positive and
PI-positive cells (late apoptotic/necrotic); R3, Annexin
V-FITC-negative and PI-negative cells (normal viable cells); R4,
Annexin V-FITC-positive and PI-negative cells (early apoptosis);
total apoptotic cells are the sum of early apoptotic cells and late
apoptotic cells. (a) Control group, (b) Model group, (c) quercetin
group, (d) 5 µg/ml TFs group, (e) 10 µg/ml TFs group, (f) 20 µg/ml
TFs group, (g) 5 µM Car group, (h) 10 µM Car group, (i) 20 µM Car
group. (B) Percentages of apoptotic cells were calculated. n=3.
##P<0.01 vs. control group; *P<0.05,
**P<0.01 vs. H2O2 model group.
TFs, total flavonoids; Car, cardamonin; RAECs, rat aortic
endothelial cells; PI, propidium iodide; Con, control; Mod, model;
Que, quercetin. |
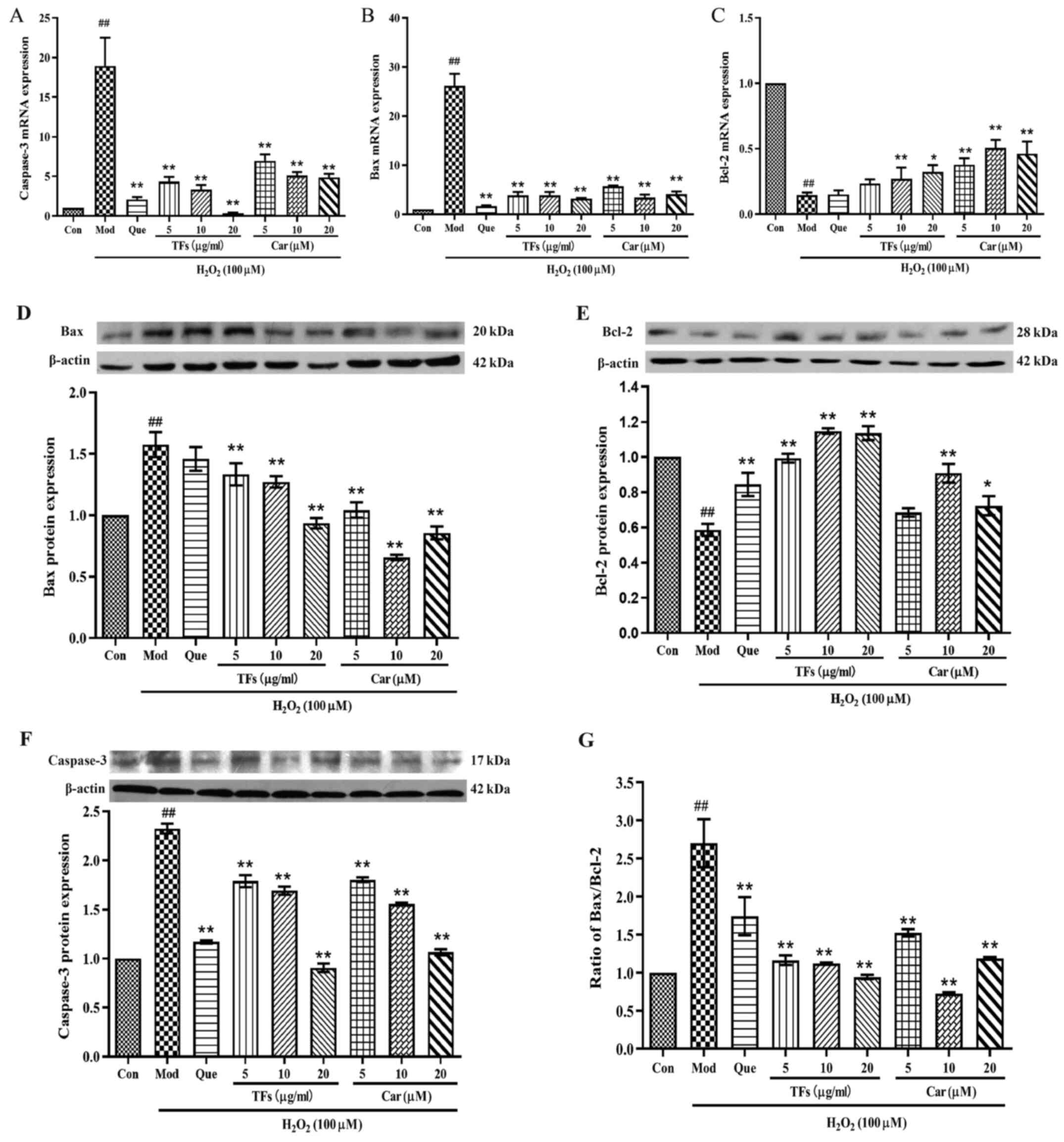
TFs and Car downregulates
apoptosis-related gene and protein expression levels in RAECs
To elucidate the molecular mechanism of the
protective effect of TFs and Car against
H2O2-induced oxidative damage in RAECs, the
mRNA expression of apoptosis-related genes, including caspase-3,
Bax and Bcl-2, were investigated. Compared with the control group,
the relative expression levels of caspase-3 and Bax mRNA in the
model group were significantly increased (P<0.01; Fig. 7A and B). Pretreatment with TFs and Car for 24 h
significantly inhibited the mRNA expression levels of caspase-3 and
Bax compared with the model group in a dose-dependent manner
(P<0.01); furthermore, its effect was similar to that of the
positive control drug, quercetin. In addition, the relative
expression of Bcl-2 mRNA in the model group was significantly lower
compared with that in the control group (P<0.01; Fig. 7C). Compared with the model group,
cells pretreated with TFs and Car for 24 h significantly reduced
the H2O2-induced inhibition of Bcl-2 mRNA
expression (P<0.01), and both TFs and Car exhibited superior
effects relative to quercetin (Fig.
7C).
Next, the expression of apoptosis-related proteins
were examined, including caspase-3, Bcl-2 and Bax, using western
blotting. Compared with the control group,
H2O2 treatment caused significantly increased
protein expression levels of caspase-3 (Fig. 7F) and Bax (Fig. 7D), and decreased protein expression
of Bcl-2 (Fig. 7E) in the model
group. By contrast, treatment with TFs significantly inhibited the
elevation of caspase-3 and Bax protein levels while upregulating
the expression of Bcl-2 protein compared with the control group
(P<0.01). Similar effects were observed when cells were
pretreated with Car. The expression levels of caspase-3 and Bax
were significantly downregulated, and the expression of Bcl-2 was
significantly upregulated. In addition, TFs and Car pretreatments
attenuated the increase in the Bax/Bcl-2 ratio (Fig. 7G).
TFs and CAR regulate the expression
levels of MAPKs and Akt phosphorylation
The Akt and MAPK pathways are important cell
signaling pathways. To further demonstrate the antiapoptotic
effects of TFs and Car, the roles of the Akt and MAPK pathways were
investigated. The results indicated that the expression levels of
p-ERK (Fig. 8A), p-JNK (Fig. 8B) and p-p38 MAPK (Fig. 8C) activation were significantly
increased in the model group compared with the control group
(P<0.01). Pretreatment with TFs and Car significantly attenuated
the activation of all three MAPKs compared with the model group
(all P<0.05). By contrast, H2O2 greatly
decreased the phosphorylation of Akt compared with the control
group, while treatment with TFs and Car significantly stimulated
phosphorylation compared with the model group (P<0.01; Fig. 8D).
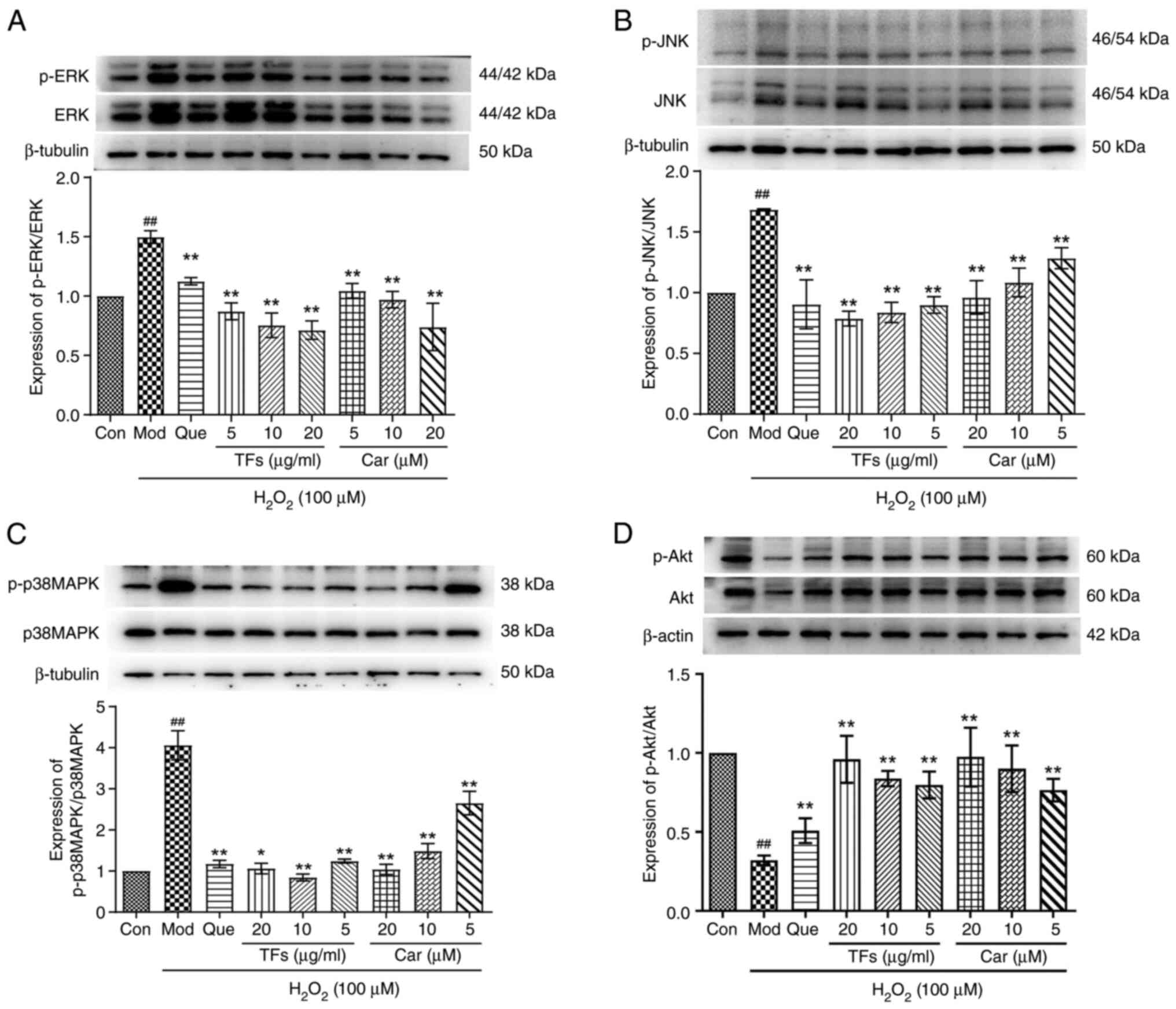 | Figure 8Effects of TFs and Car on the AKT and
MAPK signal transduction pathways in
H2O2-induced RAECs. Expression levels of (A)
p-ERK, (B) p-JNK, (C) p-p38 MAPK and (D) p-Akt proteins in RAECs
treated with TFs and Car were quantified using western blotting.
Total protein was used as a loading control. n=3.
##P<0.01 vs. Con group; *P<0.05,
**P<0.01 vs. H2O2 model group.
TFs, total flavonoids; Car, cardamonin; RAECs, rat aortic
endothelial cells; p-, phosphorylated; Con, control; Mod, model;
Que, quercetin. |
Discussion
The present study established an oxidative stress
damage model by treating RAECs with H2O2
in vitro and investigated the protective effects of
flavonoids against oxidative stress. The results indicated that TFs
and Car could significantly inhibit injury in
H2O2-induced RAECs, alleviate changes in
nuclear morphology and cause cell-protective effects. Endothelial
cells are important in maintaining the normal functions of blood
vessels by producing and secreting multiple compounds (22). Abnormal endothelial structure and
cellular malfunction are the main causes of numerous types of
diseases, such as vasculitis, atherosclerosis and thrombosis
(23).
ROS play an important role in the pathogenesis of
endothelial dysfunction and cardiovascular diseases (24). The imbalance between the production
of ROS and antioxidant defense can result in oxidative stress,
which may give rise to metabolic impairment and cell death
(25). The present study indicated
that TFs and Car could partially suppress oxidative stress by
decreasing the generation of ROS and demonstrated their potential
to protect RAECs against damage by H2O2.
The degree of oxidative damage could be determined
by the production of markers of oxidative damage products, such as
MDA and LDH. MDA is one of the indexes used to measure the degree
of oxidative stress, lipid peroxidation and cell viability. It
indirectly reflects the degree of damage due to oxygen free
radicals (26). LDH exists in cells
and participates in glycolysis, and the activity of LDH can be used
as an indicator of the degree of cell membrane damage, which is
closely associated with apoptosis. In the present study, the
results indicated that H2O2 dramatically
increased MDA levels and LDH release in RAECs, while these effects
were alleviated by treatment with TFs and Car. Therefore, these
results suggested that TFs and Car could protect RAECs from
oxidative damage.
Mammalian cells have developed an antioxidant
defense system to protect against oxidative stress, and the defense
system includes some essential antioxidant enzymes, such as SOD and
GSH-Px (27,28). SOD activity indirectly reflects the
ability to scavenge oxygen free radicals, which plays an important
role in regulating the balance between oxidation and antioxidation
(3). It can reduce the oxidative
damage induced by superoxide anions. GSH-Px has the ability to
reduce the toxic by-products of the synthetic pathway by catalyzing
peroxidative intermediates to the intermediates used in metabolism
(29). The main biological function
of GSH-Px is to remove lipid peroxides and
H2O2 (30).
Therefore, the present study analyzed the activities of SOD and
GSH-Px. The results demonstrated that TFs and Car enhanced the
antioxidant defense system by increasing the activity of SOD and
GSH-Px,which is consistent with the results of another study
(31).
Oxidative damage induced by excessive ROS generation
is known to be a potential inducer of apoptosis. Apoptosis is
associated with two family members (Bcl-2 and Bax) and caspase
family members (caspase-3, caspase-8 and caspase-9) (32). Bcl-2 family proteins, which usually
regulate the mitochondrial pathway of apoptosis, can be divided
into two groups: Anti- and pro-apoptotic proteins. Bcl-2 and Bax
proteins are the two main members of the Bcl-2 multigene family.
Bcl-2 inhibits apoptosis, whereas Bax exerts a proapoptotic effect
(33). In the present study,
pretreatment with TFs and Car decreased Bax and caspase-3 protein
expression and increased Bcl-2 protein expression. The imbalance of
the Bax/Bcl-2 ratio switched on the apoptosis process (34). The increased Bax/Bcl-2 ratio
transmitted apoptotic signals to caspase-9. Following activation,
caspase-9 activates caspase-3, which is a key executor of apoptosis
(35). The aforementioned process
is known as the mitochondrial apoptotic pathway (36). As expected, the Bax/Bcl-2 ratio was
significantly increased in the TF and Car groups compared with the
model group of the present study. These results revealed that TFs
and Car could regulate the expression of apoptosis-related proteins
and inhibit the initiation of apoptosis.
A number of studies have demonstrated that the
mechanisms of endothelial cell damage and apoptosis induced by
H2O2 are associated with the MAPK pathway
(36,37). When MAPKs are activated, they can
phosphorylate their specific cascade proteins, thus controlling
numerous cell activities, including cell proliferation,
differentiation and cell death (30,34).
The MAPK family, which comprises serine/threonine protein kinases,
is involved in the ERK, JNK and p38-MAPK signaling pathways
(38). Moreover, p38 and JNK can
initiate the mitochondria-dependent intrinsic cell death pathway
via the direct phosphorylation of the Bcl-2 family apoptotic
protein Bax (39). The present
study demonstrated that the phosphorylation levels of JNK, ERK and
p38 MAPK were all markedly increased in the model group, while
pretreatment with TFs and Car significantly attenuated this
elevation. These results suggested that TFs and Car inhibited the
activation of the JNK, ERK and p38 MAPK pathways.
Akt, a serine/threonine-protein kinase, plays a key
role in multiple cellular processes, such as apoptosis, cell
proliferation and migration (40).
However, further experiments are needed to verify the results, and
the mechanism can provide direction for preventing radiotherapy
damage via the ROS pathway.
In conclusion, TFs and Car from LCC had a strong
protective effect against H2O2-induced
oxidative damage and apoptosis of RAECs in vitro.
Anti-apoptotic activity may mediate mitochondrial Bcl-2 and Bax and
inhibit activation of the survival signal-related factor caspase-3.
The TFs extracted from LCC could be developed as effective and
potential candidate drugs to prevent oxidative stress in the
future, and they could also provide a novel direction for
preventing antioxidant activity through the ROS pathway caused by
radiation damage.
Acknowledgements
Not applicable.
Funding
This work was supported by Zhejiang Chinese Medical Science and
Technology Program (grant no. 2019ZB120).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JJF conceived the study. FMZ and JJH performed the
experiments, analyzed the data, and wrote the manuscript. XJH and
JW contributed to the methodology and data analysis. BQZ, SH and
ZSD analyzed the data, and wrote and revised the manuscript. JJF
acquired funding, contributed to resources, and supervised the
study. FMZ and ZSD confirm the authenticity of all the raw data.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Eelen G, de Zeeuw P, Simons M and
Carmeliet P: Endothelial cell metabolism in normal and diseased
vasculature. Circ Res. 116:1231–1244. 2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Figarola JL, Singhal J, Rahbar S, Awasthi
S and Singhal SS: LR-90 prevents methylglyoxal-induced oxidative
stress and apoptosis in human endothelial cells. Apoptosis.
19:776–788. 2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Li H, Horke S and Förstermann U: Oxidative
stress in vascular disease and its pharmacological prevention.
Trends Pharmacol Sci. 34:313–319. 2013.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Toghueo K, Marie R and Boyom FF:
Endophytes from ethno-pharmacological plants: Sources of novel
antioxidants - A systematic review. Biocatal Agric Biotechnol: Nov
14, 2019 (Epub ahead of print).
|
|
5
|
Mahdi A, Kövamees O and Pernow J:
Improvement in endothelial function in cardiovascular disease - Is
arginase the target? Int J Cardiol. 301:207–214. 2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Harvey AL, Edrada-Ebel R and Quinn RJ: The
re-emergence of natural products for drug discovery in the genomics
era. Nat Rev Drug Discov. 14:111–129. 2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Rao T, Tan Z, Peng J, Guo Y, Chen Y, Zhou
H and Ouyang D: The pharmacogenetics of natural products: A
pharmacokinetic and pharmacodynamic perspective. Pharmacol Res.
146(104283)2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Beretta G and Facino RM: Recent advances
in the assessment of the antioxidant capacity of pharmaceutical
drugs: From in vitro to in vivo evidence. Anal Bioanal Chem.
398:67–75. 2010.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Olszowy M: What is responsible for
antioxidant properties of polyphenolic compounds from plants? Plant
Physiol Biochem. 144:135–143. 2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Brainina K, Tarasov A, Khamzina E, Stozhko
N and Vidrevich M: Contact hybrid potentiometric method for on-site
and in situ estimation of the antioxidant activity of fruits and
vegetables. Food Chem. 309(125703)2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wen L, Jiang Y, Yang J, Zhao Y, Tian M and
Yang B: Structure, bioactivity, and synthesis of methylated
flavonoids. Ann N Y Acad Sci. 1398:120–129. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Chen GL, Fan MX, Wu JL, Li N and Guo MQ:
Antioxidant and anti-inflammatory properties of flavonoids from
lotus plumule. Food Chem. 277:706–712. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wei L, Yang M, Huang L and Lin Li J:
Antibacterial and antioxidant flavonoid derivatives from the fruits
of Metaplexis japonica. Food Chem. 289:308–312.
2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Feng S, Wang L, Belwal T, Li L and Luo Z:
Phytosterols extraction from hickory (Carya cathayensis
Sarg.) husk with a green direct citric acid hydrolysis extraction
method. Food Chem. 315(126217)2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Xiang L, Wang Y, Yi X, Wang X and He X:
Chemical constituent and antioxidant activity of the husk of
Chinese Hickory. J Funct Foods. 23:378–388. 2016.
|
|
16
|
Zhu X, Li W, Yu Y, Jiang F and Ding Z:
Total flavonoids preparation of the Carya cathayensis Sarg.
leaves. Zhonghua Zhongyiyao Xuekan. 31:147–149. 2013.
|
|
17
|
Shen Y, Liu NN, Min XU, Zhang K, Jiang FS,
Chen JZ, Tian SS and Ding ZS: HPLC determination of the content of
the five flavonoid aglycones from the leaves of Carya
cathayensis Sarg. Yaowu Fenxi Zazhi. 33:804–807. 2013.
|
|
18
|
Cao XD, Ding ZS, Jiang FS, Ding XH, Chen
JZ, Chen SH and Lv GY: Antitumor constituents from the leaves of
Carya cathayensis. Nat Prod Res. 26:2089–2094.
2012.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Tian SS, Jiang FS, Zhang K, Zhu XX, Jin B,
Lu JJ and Ding ZS: Flavonoids from the leaves of Carya
cathayensis Sarg. inhibit vascular endothelial growth
factor-induced angiogenesis. Fitoterapia. 92:34–40. 2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Abu Bakar MF, Mohamed M, Rahmat A, Burr SA
and Fry JR: Cellular assessment of the extract of bambangan
(Mangifera pajang) as a potential cytoprotective agent for
the human hepatocellular HepG2 cell line. Food Chem. 136:18–25.
2013.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Livak KJ and Schmittgen TDL: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Chen S, Tang Y, Qian Y, Chen R, Zhang L,
Wo L and Chai H: Allicin prevents
H2O2-induced apoptosis of Huvecs by
inhibiting an oxidative stress pathway. BMC Complement Altern Med.
14(321)2014.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Jansen F, Li Q, Pfeifer A and Werner N:
Endothelial- and immune cell-derived extracellular vesicles in the
regulation of cardiovascular health and disease. JACC Basic Transl
Sci. 2:790–807. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Sugamura K and Keaney JF Jr: Reactive
oxygen species in cardiovascular disease. Free Radic Biol Med.
51:978–992. 2011.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Srivastava KK and Kumar R: Stress,
oxidative injury and disease. Indian J Clin Biochem. 30:3–10.
2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Ma X, Zhang K, Li H, Han S, Ma Z and Tu P:
Extracts from Astragalus membranaceus limit myocardial cell
death and improve cardiac function in a rat model of myocardial
ischemia. J Ethnopharmacol. 149:720–728. 2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Ghaffari H, Ghassam BJ and Prakash HS:
Hepatoprotective and cytoprotective properties of Hyptis suaveolens
against oxidative stress-induced damage by CCl(4) and H(2)O(2).
Asian Pac J Trop Med. 5:868–874. 2012.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Kwok HH, Ng WY, Yang MS, Mak NK, Wong RN
and Yue PY: The ginsenoside protopanaxatriol protects endothelial
cells from hydrogen peroxide-induced cell injury and cell death by
modulating intracellular redox status. Free Radic Biol Med.
48:437–445. 2010.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Cohen G and Hochstein P: Glutathione
peroxidase: The primary agent for the elimination of hydrogen
peroxide in erythrocytes. Biochemistry. 2:1420–1428.
1963.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Wu P, Ma G, Li N, Deng Q, Yin Y and Huang
R: Investigation of in vitro and in vivo antioxidant activities of
flavonoids rich extract from the berries of Rhodomyrtus
tomentosa(Ait.) Hassk. Food Chem. 173:194–202. 2015.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Wu Y, Wang Y and Nabi X: Protective effect
of Ziziphora clinopodioides flavonoids against
H2O2-induced oxidative stress in HUVEC cells.
Biomed Pharmacother. 117(109156)2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Xu F, Ren L, Song M, Shao B, Han Y, Cao Z
and Li Y: Fas- and mitochondria-mediated signaling pathway involved
in osteoblast apoptosis induced by AlCl3. Biol Trace Elem Res.
184:173–185. 2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Miura M, Chen XD, Allen MR, Bi Y, Gronthos
S, Seo BM, Lakhani S, Flavell RA, Feng XH, Robey PG, et al: A
crucial role of caspase-3 in osteogenic differentiation of bone
marrow stromal stem cells. J Clin Invest. 114:1704–1713.
2004.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Lindqvist LM, Heinlein M, Huang DC and
Vaux DL: Prosurvival Bcl-2 family members affect autophagy only
indirectly, by inhibiting Bax and Bak. Proc Natl Acad Sci USA.
111:8512–8517. 2014.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Chen J, Gu Y, Shao Z, Luo J and Tan Z:
Propofol protects against hydrogen peroxide-induced oxidative
stress and cell dysfunction in human umbilical vein endothelial
cells. Mol Cell Biochem. 339:43–54. 2010.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Alamdary SZ, Khodagholi F, Shaerzadeh F,
Ansari N, Sonboli A and Tusi SK: S. choloroleuca, S.
mirzayanii and S. santolinifolia protect PC12 cells from
H(2)O(2)-induced apoptosis by blocking the intrinsic pathway.
Cytotechnology. 64:403–419. 2012.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Wang B, Luo T, Chen D and Ansley DM:
Propofol reduces apoptosis and up-regulates endothelial nitric
oxide synthase protein expression in hydrogen peroxide-stimulated
human umbilical vein endothelial cells. Anesth Analg.
105:1027–1033. 2007.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Al-Azayzih A, Gao F, Goc A and Somanath
PR: TGFβ1 induces apoptosis in invasive prostate cancer and bladder
cancer cells via Akt-independent, p38 MAPK and JNK/SAPK-mediated
activation of caspases. Biochem Biophys Res Commun. 427:165–170.
2012.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Shi L, Yu X, Yang H and Wu X: Advanced
glycation end products induce human corneal epithelial cells
apoptosis through generation of reactive oxygen species and
activation of JNK and p38 MAPK pathways. PLoS One.
8(e66781)2013.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Abbas A, Abdelsamea MM and Gaber MM:
Classification of Covid-19 in chest X-Ray images using detrac deep
convolutional neural network. Appl Intell. 51:854–864. 2020.
|