Introduction
Chronic kidney disease (CKD) is a long-term
condition affecting ~10% of the global population. It is a key
cause of morbidity and mortality worldwide that accounts for a
large proportion of medical expenses (1,2).
Among the various factors that contribute to the progression of
CKD, proteinuria is relatively independent, and an important
pathophysiological condition for the generation of this symptom is
podocyte injury (3-5).
Podocytes are highly specialized terminally differentiated
epithelial cells that maintain the glomerular filtration barrier
and are vulnerable to multiple injuries (6-8).
One of the causes of podocyte loss and injury is apoptosis, thus it
is of clinical importance to identify the potential therapeutic
targets of podocyte apoptosis (9,10).
MicroRNAs (miRNAs) are small non-coding RNAs that
suppress gene expression by targeting the 3'-untranslated regions
(UTRs) of mRNAs, leading to mRNA degradation or transcriptional
blocking (11). Previous studies
reported that miRNAs are involved in numerous biological processes,
including cell apoptosis, proliferation, differentiation and
development, and in a variety of kidney diseases (12,13).
For example, miR-24 promotes renal ischemic injury by inducing the
apoptosis of endothelial and tubular epithelial cells (14). Furthermore, miR-34a promotes renal
fibrosis by inhibiting klotho expression in tubular epithelial
cells (15). The key role of
miR-29c in promoting the progression of diabetic nephropathy is
mainly achieved through the Spry1/Rho-kinase pathway (16). Recent studies demonstrated that
miR-199b-5p serves a critical role in numerous diseases (17). Lai et al (18) reported that miR-199b-5p is
downregulated in renal cell carcinoma and acts as a tumor
suppressor. However, no study has yet revealed the role of
miR-199b-5p in podocyte injury.
The present study used TargetScan and miRDB tools to
analyze the interactions between miR-199b-5p and different mRNAs,
and reported that miR-199b-5p had potential binding sites in the
3'-UTR of the regulator of G-protein signaling 10 (RGS10). As an
important member of the RGS protein superfamily, RGS10 is involved
in the regulation of certain physiological processes in various
types of disease (19,20); however, its role in renal diseases
remains unknown. The present study aimed therefore to investigate
the effects of miR-199b-5p on RGS10 expression, cell apoptosis and
injury in podocytes.
Materials and methods
Cell culture and transfection
The immortalized mouse podocytes (MPC5 cell line)
were gifted by the Nephrology Research Center of The Second
Affiliated Hospital of Nanjing Medical University. Podocytes were
cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin (Gibco; Thermo Fisher Scientific, Inc.),
100 µg/ml streptomycin and 10 U/ml IFN-γ (PeproTech, Inc.) and
placed at 33˚C in an incubator containing 5% CO2 for 2-3
generations. When proliferation reached 70-80% (~2x105
cells per well), medium was replaced with IFN-γ free medium for
differentiation and cells were placed in an incubator at 37˚C with
5% CO2 for 10-14 days. The miR-199b-5p mimic (sense,
5'-CCCAGU GUUUAGACUACCUGUUC-3'; antisense, 5'-ACAGGUAGU
CUAAACACUGGGUU-3'; GenePharma, Shanghai, China), miR-199b-5p
inhibitor (5'-GAACAGGUAGUCUAAACACU GGG-3'; Shanghai GenePharma Co.,
Ltd.), negative control mimic (sense, 5'-UUCUCCGAACGUGUCACGUTT-3';
antisense, 5'-ACGUGACACGUUCGGAGAATT-3'; Shanghai GenePharma Co.,
Ltd.), and negative control inhibitor (5'-CAG
UACUUUUGUGUAGUACAA-3'; Shanghai GenePharma Co., Ltd.) were
transfected for 6 h at a concentration of 100 nM using
Lipofectamine 2000 transfection reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
medium was changed again, and 1 µg/ml ADR (MedChemExpress) was
added for 24 h treatment (21). To
verify the effect of AKT/mTOR pathway on ADR-induced podocyte
apoptosis, the differentiated podocytes were treated with 1 mg/l
rapamycin (RAPA; MedChemExpress) after ADR intervention. In
addition, to verify the relationship between RGS10 and apoptosis,
rescue experiments were performed in which podocytes were
transfected with miR-199b-5p mimic and treated with ADR, then
transfected with 1x108 PFU/ml adenoviral RGS10 (AdRGS10;
Shanghai GenePharma Co., Ltd.) or adenoviral negative control
(AdNC; Shanghai GenePharma Co., Ltd.) for 48 h.
Bioinformatical analysis
Prediction of the miR-199b-5p-target gene was
performed using the TargetScan (http://www.targetscan.org/vert_71/) and miRDB
(http://mirdb.org/).
Western blotting
Cells were lysed using RIPA Lysis Buffer (Beyotime
Institute of Biotechnology) supplemented with 1% protease inhibitor
phenylmethylsulfonyl fluoride (Beyotime Institute of
Biotechnology). Protein concentration was determined using the BCA
method (Beyotime Institute of Biotechnology). Proteins (30 µg) were
separated by 8% or 12% SDS-PAGE and were transferred onto PVFD
membranes (MilliporeSigma; cat. no. HATF09025). Membranes were
blocked with 5% non-fat dry milk solution prepared in 0.01 M
Tris-buffered saline (pH 7.4, containing 0.01% Tween-20) at room
temperature for 2 h. Membranes were incubated with primary
antibodies against nephrin (1:1,000; mouse; cat. no. sc-376522;
Santa Cruz Biotechnology, Inc.), podocin (1:1,000; rabbit; cat. no.
ab181143; Abcam), RGS10 (1:1,000; rabbit; cat. no. DF4414; Affinity
Biosciences, Ltd.), Bax (1:1,000; rabbit; cat. no. AF0120; Affinity
Biosciences, Ltd.), Bcl-2 (1:1,000; rabbit; cat. no. AF6139;
Affinity Biosciences, Ltd.), mTOR (1:1,000; mouse; cat. no.
66888-1-Ig; ProteinTech Group, Inc.), phosphorylated (p-) mTOR
(1:1,000; rabbit; cat. no. 5536T; Cell Signaling Technology), AKT
(1:1,000; rabbit; cat. no. 10176-2-AP; ProteinTech Group, Inc.),
p-AKT (1:2,000; Ser473; mouse; cat. no. 66444-1-Ig; ProteinTech
Group, Inc.) and GAPDH (1:1,000; rabbit; cat. no. 5174; Cell
Signaling Technology, Inc.), overnight at 4˚C. Membranes were
washed with TBST and incubated with horseradish
peroxidase-conjugated secondary antibodies (goat anti-mouse,
1:2,000, cat. no. 91196S, Cell Signaling Technology, Inc.; or goat
anti-rabbit, 1:4,000, cat. no. S0001, Affinity Biosciences, Ltd.)
at room temperature for 2 h. Enhanced chemiluminescence reagent
(Biosharp) and a gel imaging analysis system were used to detect
the signal on the membrane. The data were analyzed via densitometry
using ImageJ software 1.8.0 (National Institute of Health) and
normalized to expression of the internal control (GAPDH).
Real-time quantitative PCR
(RT-q)PCR
Total RNA (including mRNA and microRNA) was
extracted using TRIzol® reagent (Life Technologies;
Agilent, Inc.). miRNA RT-qPCR was performed using the All-in-One
MicroRNA Assay Kit (iGeneBio, Inc.) and Applied Biosystem 7000
Quantitative PCR Instrument using U6 as an internal reference. mRNA
RT-qPCR was performed using the HiScript III RT SuperMix (Vazyme
Biotech Co., Ltd.) and GAPDH was used as an internal reference. The
sequences of the primers used (Ruijie Technology) are listed in
Table I. The thermocycling
conditions are presented in Tables
II and III. Three replicate
wells were set for each sample, and the experiment was repeated at
least three times. The relative expression levels were normalized
to endogenous controls and were expressed as 2-ΔΔCq
(22).
 | Table ISequences of the primer sequences
used for reverse transcription quantitative PCR. |
Table I
Sequences of the primer sequences
used for reverse transcription quantitative PCR.
| Genes | Forward primer | Reverse primer |
|---|
| miR-199b-5p |
5'-UUAUCCUAAUUGCUCCUACGGCU-3' |
5'-AUUCGGCAUCGCGCUAAACGUUA-3' |
| U6 |
5'-CTCGCTTCGGCAGCACATATACT-3' |
5'-ACGCTTCACGAATTTGCGTGTC-3' |
| GAPDH |
5'-TGGATTTGGACGCATTGGTC-3' |
5'-TTTGCACTGGTACGTTGAT-3' |
| Nephrin |
5'-ATGGGAGCTAAGGAAGCCACA-3' |
5'-GATGGAGGATTACGCTGGG-3' |
| Podocin |
5'-GCATCAAGCCCTCTGGATTAG-3' |
5'-AGACGGAGATCAACCTTGTGATA-3' |
| Bax |
5'-TGAAGACAGGGGCCTTTTTG-3' |
5'-AATTCGCCGGAGACACTCG-3' |
| Bcl-2 |
5'-GTCGCTACCGTCGTGACTTC-3' |
5'-CAGACATGCACCTACCCAGC-3' |
| RGS10 |
5'-TCCATGACGGAGATGGGAG-3' |
5'-AACAAGACATTCTCTTCGCTGAA-3' |
| Table IIThermocycling conditions used during
reverse transcription quantitative PCR for microRNA expression
level evaluation. |
Table II
Thermocycling conditions used during
reverse transcription quantitative PCR for microRNA expression
level evaluation.
| Stage | Step | Repetition | Temperature,
˚C | Duration |
|---|
| Stage 1 | Initial
denaturation | 1 | 95 | 10 min |
| Stage 2 | Amplification
cycles | 40 | 95 | 10 sec |
| | | | 60 | 20 sec |
| | | | 72 | 10 sec |
| Stage 3 | Dissociation
curve | 1 | 95 | 15 sec |
| Table IIIThermocycling conditions used during
reverse transcription quantitative PCR for mRNA expression level
evaluation. |
Table III
Thermocycling conditions used during
reverse transcription quantitative PCR for mRNA expression level
evaluation.
| Stage | Step | Repetition | Temperature,
˚C | Duration |
|---|
| Stage 1 | Initial
denaturation | 1 | 95 | 30 sec |
| Stage 2 | Amplification
cycles | 40 | 95 | 10 sec |
| | | | 60 | 30 sec |
| Stage 3 | Dissociation
curve | 1 | 95 | 15 sec |
| | | | 60 | 60 sec |
| | | | 95 | 15 sec |
Apoptosis detection (Annexin
V-APC/7-AAD)
Cells were stained with the Annexin V-APC/7-AAD
apoptosis detection kit (Nanjing KeyGen Biotech Co., Ltd.)
according to the manufacturer's instructions. The CytoFLEXLX Flow
cytometer (Beckman Coulter, Inc.) was used for data analysis. The
excitation wavelength was 633 nm. The FL4 channel was used for the
red fluorescence of Annexin V-APC. The FL2 channel was used for the
7-AAD red fluorescence. Annexin V-APC was used as the x-axis, and
7-AAD was used as the y-axis. The numerical axis was the
fluorescence signal intensity. A two-dimensional scatter diagram
was constructed and the apoptotic rate was evaluated. Data analysis
was conducted using FlowJo 7.6 (Treestar).
Statistical analysis
Data analyses were performed using GraphPad Prism
6.0 software (GraphPad Software, Inc.). Data are expressed as the
means ± standard deviation. Comparisons between two groups were
performed by t-test with Bonferroni correction. Comparisons between
more than two groups were performed by one-way ANOVA followed by a
Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-199b-5p is upregulated in
ADR-induced podocytes
To explore the role miR-199b-5p in podocyte injury,
ADR-treated podocytes were established as the in vitro
experimental model. Damage of podocytes stimulated by ADR was first
evaluated. To this end, podocytes were cultured and differentiated,
and then treated with ADR. Following treatment, podocyte apoptosis
was evaluated using Annexin V-APC/7-AAD double staining and flow
cytometry. The results demonstrated that the apoptosis rate in the
ADR group was significantly increased compared with the control
group (normal podocytes; Fig. 1A).
Subsequently, expression levels of Bax and Bcl-2, two known
indicators of apoptosis (23),
were evaluated. The results from western blotting (Fig. 1B) and RT-qPCR (Fig. 1C) demonstrated that, compared with
the control group, ADR treatment increased the expression level of
Bax, whereas it decreased the level of Bcl-2 in ADR-treated
podocytes, resulting in their apoptosis. Furthermore, the
expression levels of nephrin and podocin, which are markers of
podocyte function, were also evaluated in the control and
ADR-treated groups. The results from western blotting (Fig. 1B) and RT-qPCR (Fig. 1C) showed significant decrease in
nephrin and podocin expression levels in ADR-treated podocytes
compared with the control group. In addition, a significant
increase in miR-199b-5p expression in ADR-treated podocytes was
demonstrated compared with that in normal podocytes (Fig. 1D). These findings suggested that
the elevated miR-199b-5p expression may contribute to ADR-mediated
podocyte apoptosis.
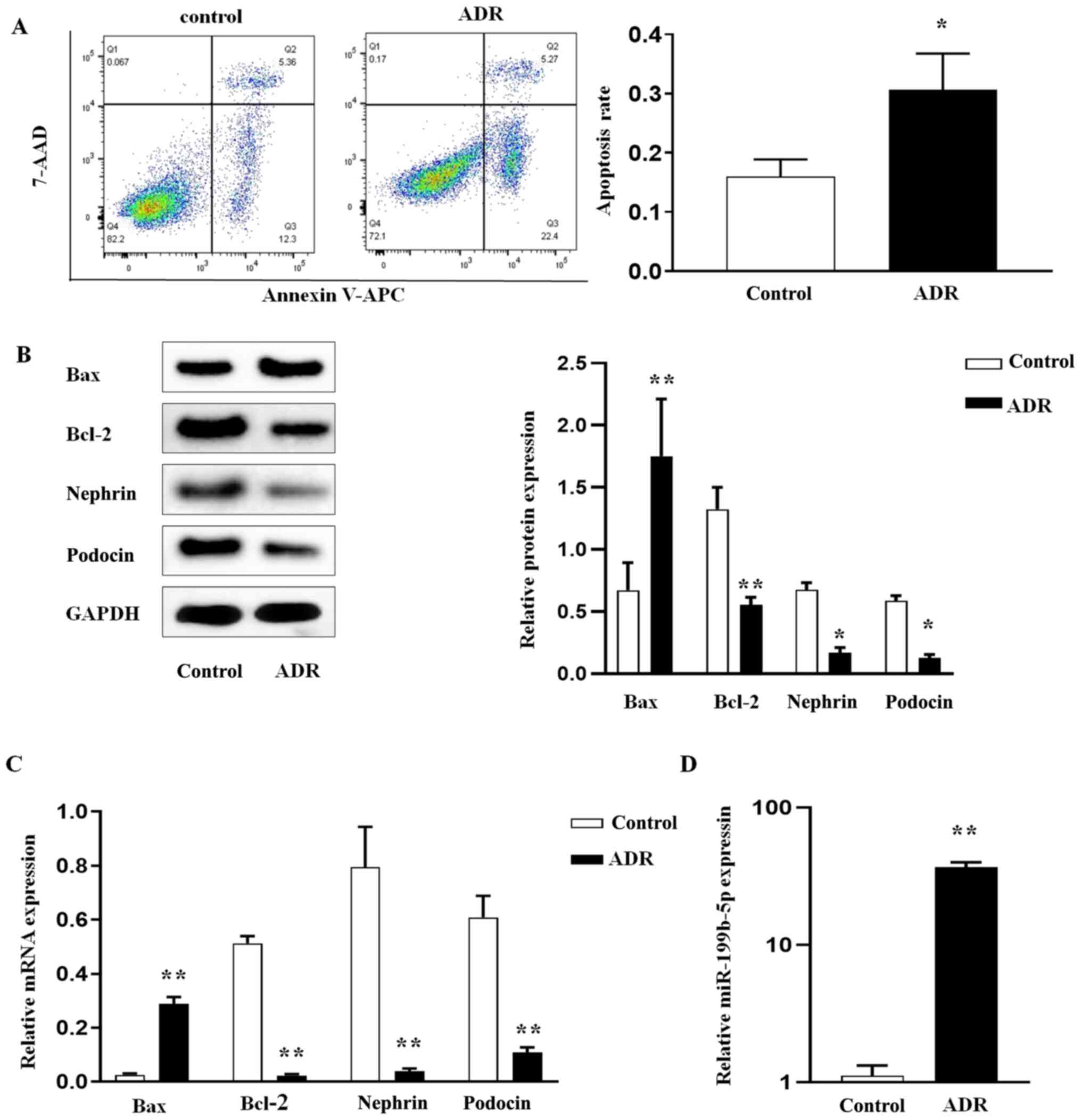 | Figure 1ADR induces podocyte apoptosis and the
regulation of miR-199b-5p. After podocytes were treated with ADR,
(A) the apoptosis rate was determined by flow cytometry. (B) Bax,
Bcl-2, nephrin, and podocin protein expression was determined by
western blotting. (C) RT-qPCR analysis of Bax, Bcl-2, nephrin, and
podocin in podocytes. (D) RT-qPCR analysis of miR-199b-5p in
podocytes. Control indicates normal podocytes. n=3 in each group.
*P<0.05 and **P<0.01. ADR, adriamycin;
miR, microRNA; RT-qPCR, reverse transcription quantitative PCR. |
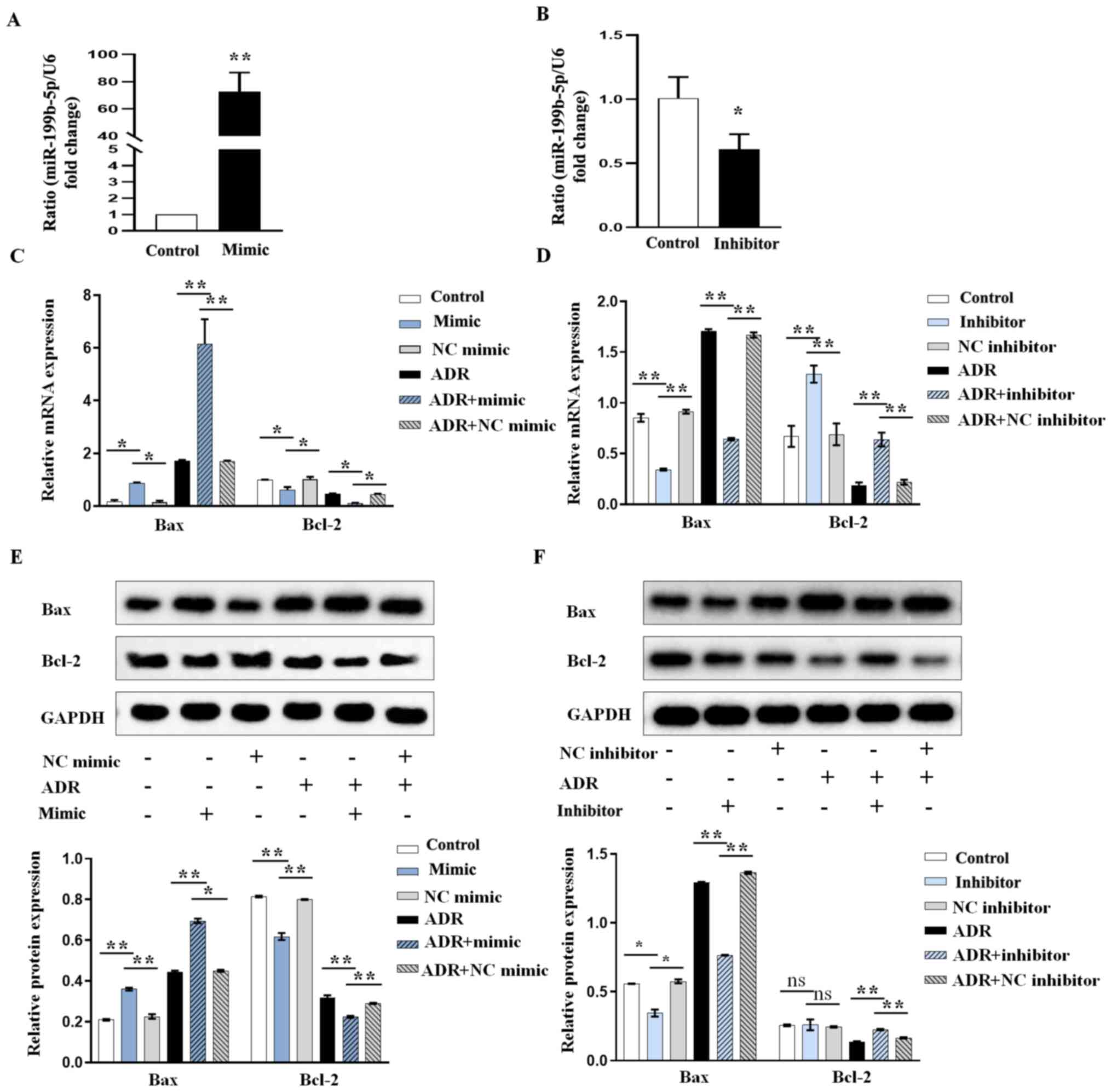
miR-199b-5p downregulation inhibits
ADR-induced podocyte apoptosis and miR-199b-5p overexpression
promotes apoptosis
We further investigated the role of miR-199b-5p in
the induction of apoptosis using miR-199b-5p overexpression or
knockdown. The results from RT-qPCR confirmed that the miR-199b-5p
mimic (abbreviated as mimic in all figures) could significantly
increase miR-199b-5p expression (Fig.
2A) while miR-199b-5p inhibitor (abbreviated as inhibitor in
all figures) significantly decreased miR-199b-5p expression in
podocytes (Fig. 2B). Furthermore,
according to the results from RT-qPCR and western blotting, the
increase in Bax and decrease in Bcl-2 expression levels in
ADR-treated podocytes were significantly aggravated by miR-199b-5p
mimic (Fig. 2C and E), whereas these were significantly
neutralized by miR-199b-5p inhibitor (Fig. 2D and F). These results indicated that
miR-199b-5p may be considered as an important regulator in
ADR-induced podocyte injury.
miR-199b-5p regulates the expression
of RGS10 in podocytes
To elucidate the molecular mechanisms by which
miR-199b-5p exerts its function, miR-199b-5p targets were explored
using TargetScan and miRDB bioinformatics algorithms. RGS10 was
predicted to be a potential target of miR-199b-5p (Fig. 3A). Thus, we further examined the
effect of miR-199b-5p on the expression of RGS10. The results from
RT-qPCR confirmed that miR-199b-5p overexpression significantly
decreased the mRNA expression of RGS10 (Fig. 3B). Conversely, silencing
miR-199b-5p significantly increased RGS10 expression level in
podocytes (Fig. 3C). The results
from western blotting confirmed that overexpression of miR-199b-5p
also inhibited the protein expression of RGS10 (Fig. 3D), while knocking down miR-199b-5p
had the opposite effect (Fig. 3E).
These results suggested that miR-199b-5p may negatively regulate
RGS10 expression at the post-transcriptional level in podocytes.
Subsequently, we determined whether RGS10 overexpression could
rescue the pro-apoptotic effect of miR-199b-5p on ADR-induced
podocytes. Podocytes were transfected with AdRGS10 to increase
RGS10 expression and AdNC was used as the negative control
(Fig. 3F). The results from
western blotting demonstrated that podocyte co-transfection with
miR-199b-5p mimic and AdRGS10 significantly reversed miR-199b-5p
mimic-induced cell apoptosis in ADR-induced podocytes, as evidenced
by the decreased expression of Bax and increased expression of
Bcl-2 (Fig. 3G). These results
further confirmed that miR-199b-5p may induce apoptosis via
regulating RGS10 expression in podocytes.
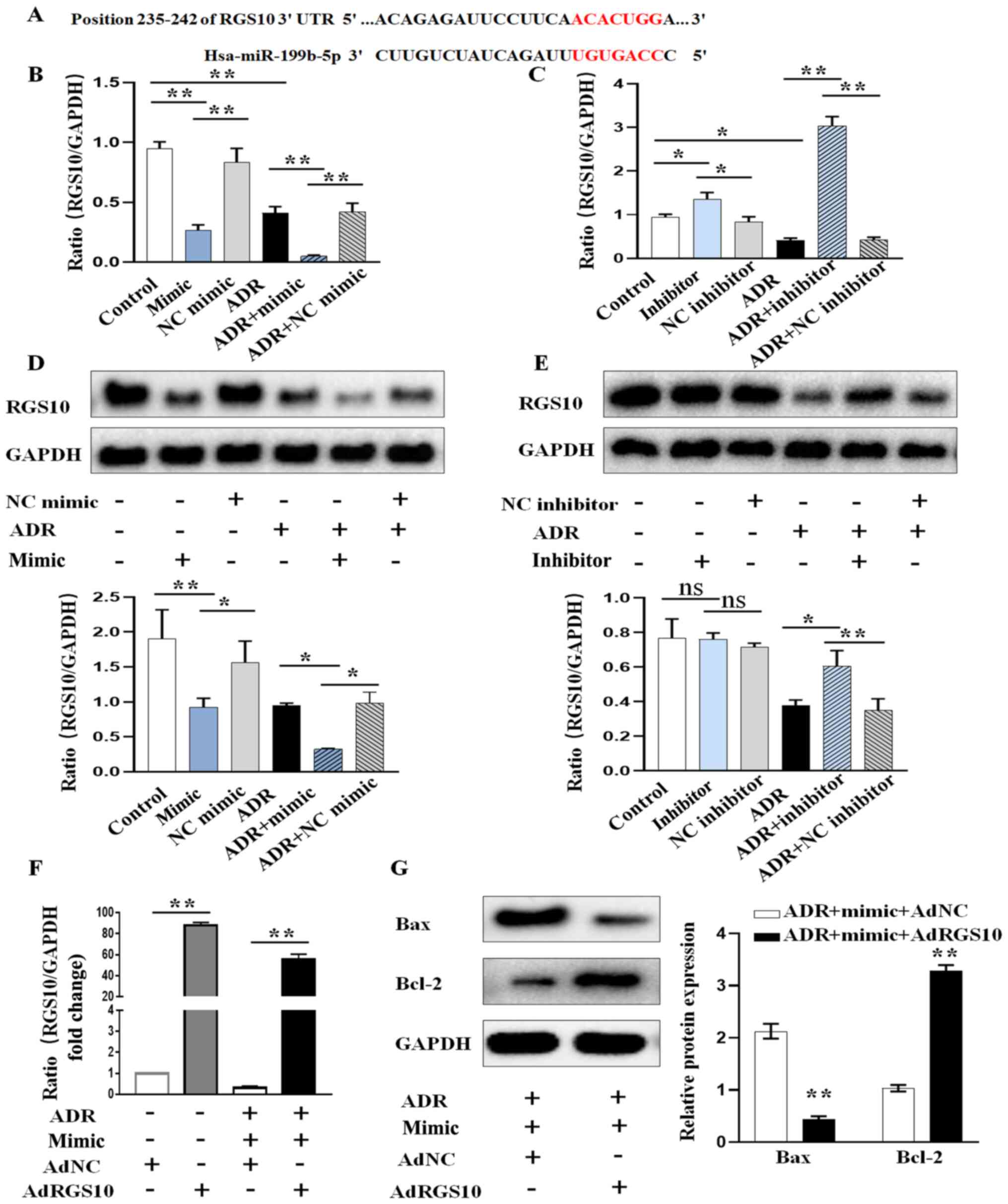 | Figure 3miR-199b-5p regulates the expression
of RGS10. (A) Predicted binding sites of miR-199b-5p on the 3'UTR
of RGS10. (B) RGS10 expression level after transient transfection
and treatment of podocytes with miR-199b-5p mimic, NC mimic, ADR,
ADR+miR-199b-5p mimic or ADR+NC mimic. (C) RGS10 expression level
after the transient transfection and treatment of podocytes with
miR-199b-5p inhibitor, NC inhibitor, ADR, ADR+miR-199b-5p inhibitor
or ADR+NC inhibitor. (D) RGS10 protein expression in podocytes
treated as in (B). (E) RGS10 protein expression in podocytes
treated as in (C). (F) Expression level of RGS10 was determined by
RT-qPCR in ADR-treated podocytes transfected with miR-199b-5p mimic
or miR-199b-5p mimic+AdRGS10. (G) Expression of Bax and Bcl-2 was
determined by western blotting in ADR-treated podocytes transfected
as in (F). Control indicates normal podocytes. n=3 in each group.
*P<0.05 and **P<0.01. ADR, adriamycin;
miR, microRNA; RT-qPCR, reverse transcription quantitative PCR; NC,
negative control; RGS10, regulator of G-protein signaling 10. |
miR-199b-5p promotes the apoptosis of
ADR-stimulated podocytes by inhibiting RGS10 and activating the
AKT/mTOR signaling pathway
To evaluate the effects of miR-199b-5p on the
downstream signaling of RGS10, we determined the expression of
p-AKT and p-mTOR in ADR-induced podocytes. Podocytes were
transfected with miR-199b-5p mimic, inhibitor or vector. Western
blotting analysis demonstrated that miR-199b-5p mimic could promote
the ADR-induced phosphorylation of AKT and mTOR in podocytes
(Fig. 4A). Conversely, miR-199b-5p
knockdown significantly inhibited the increase in p-AKT and p-mTOR
induced by ADR (Fig. 4B). These
findings indicated that the high expression of p-AKT and p-mTOR
ADR-treated podocytes overexpressing miR-199b-5p was reversed by
RGS10 overexpression (Fig. 4C). In
addition, we confirmed that inhibition of AKT/mTOR by RAPA
neutralized the pro-apoptotic effect of miR-199b-5p mimic treatment
in ADR-triggered podocytes (Fig.
4D). Taken together, these results indicated that miR-199b-5p
may regulate podocyte apoptosis via the RGS10/AKT/mTOR axis.
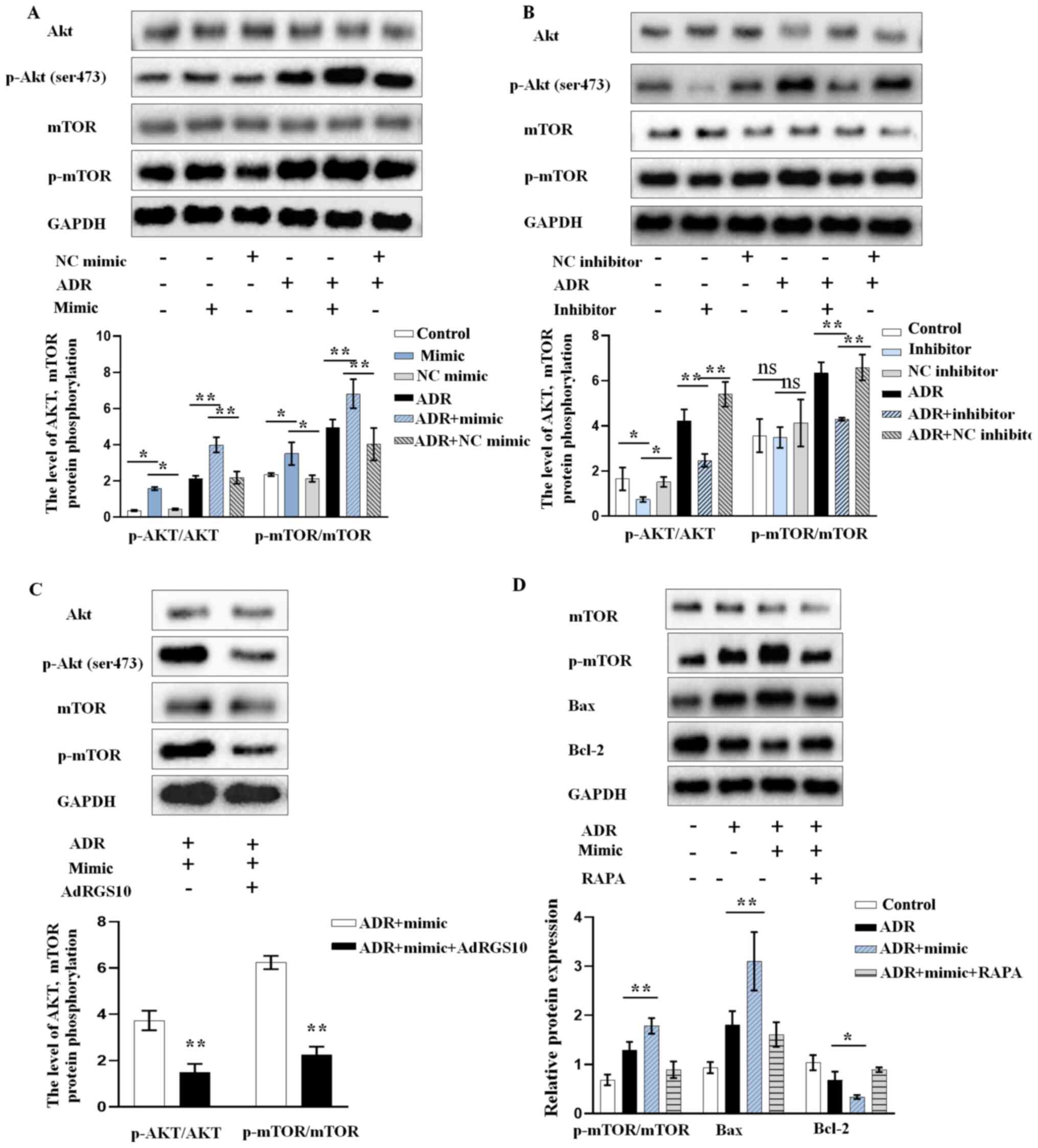 | Figure 4miR-199b-5p activates mTOR/AKT
signaling via regulating RGS10 expression. AKT, p-AKT, mTOR, and
p-mTOR protein expression level after transient transfection and
treatment of podocytes with (A) miR-199b-5p mimic, NC mimic, ADR,
ADR+miR-199b-5p mimic or ADR+NC mimic. and (B) miR-199b-5p
inhibitor, NC inhibitor, ADR, ADR+miR-199b-5p inhibitor or ADR+NC
inhibitor. (C) AKT, p-AKT, mTOR, and p-mTOR protein expression
level in ADR-treated podocytes transfected with miR-199b-5p mimic
or miR-199b-5p mimic+AdRGS10. (D) Expression of p-mTOR, mTOR and
apoptosis-related proteins in podocytes treated with ADR,
ADR+miR-199b-5p mimic or ADR+miR-199b-5p mimic+RAPA evaluated by
western blotting. Control indicates normal podocytes. n=3 in each
group. *P<0.05 and **P<0.01. ADR,
adriamycin; RAPA, rapamycin; ns, non-significant; Ad, adenovirus;
RGS10, regulator of G-protein signaling 10; mTOR, mechanistic
target of rapamycin; NC, negative control; p, phosphorylated. |
Discussion
Podocyte apoptosis is critical in the development of
CKD. To the best of our knowledge, the present study demonstrated
for the first time that miR-199b-5p may serve a critical role in
the pathogenesis of podocyte apoptosis. This study suggested that
miR-199b-5p was upregulated in ADR-induced podocytes of mice.
Furthermore, overexpression of miR-199b-5p in ADR-stimulated
podocytes exacerbated podocyte apoptosis, whereas miR-199b-5p
downregulation ameliorated podocyte lesions. It was also found that
miR-199b-5p could negatively regulate RGS10 and activate the
AKT/mTOR signaling pathway in podocytes. These findings indicated
that miR-199b-5p may be considered as be a novel therapeutic target
for CKD.
There is increasing evidence suggesting that
miR-199b-5p can regulate apoptosis in a variety of diseases. A
previous study demonstrated that miR-199b-5p inhibits cell
apoptosis and promotes cell proliferation and metastasis in
cervical cancer cells (24).
Furthermore, in the metastasis of papillary thyroid carcinoma,
miR-199b-5p was found to be upregulated and involved in the process
of cancer cell apoptosis (25). In
addition, Wang et al (26)
reported that miR-199b-5p overexpression enhances cell apoptosis in
oral cancer. So far, few studies have evaluated the role of
miR-199b-5p in kidney disease; however, Kang et al (27) reported that miR-199b-5p aggravates
the renal tubular injury in diabetic nephropathy by promoting
apoptosis in renal tubular cells via targeting klotho. However, the
role of miR-199b-5p in podocyte apoptosis remains unclear and was
therefore investigated in the present study. The results from cell
functional experiments indicated that miR-199b-5p overexpression
promoted podocyte apoptosis, whereas miR-199b-5p knockdown had the
opposite effect. Furthermore, silencing miR-199b-5p silencing
rescued the aggravated impact of podocyte apoptosis induced by
ADR.
Another major finding from the present study was the
identification of RGS10 as a novel target gene of miR-199b-5p.
RGS10 is one of the smallest members of the RGS family and it
exerts various biological effects in multiple organs (19,28).
It has been reported that ovarian cancer cell apoptosis is
decreased following RGS10 knockdown (29). Furthermore, a previous study
demonstrated that premature apoptosis occurs in osteoclasts after
RGS10 downregulation (30).
However, the role and underlying mechanism of RGS10 in podocyte
apoptosis remain to be elucidated. The results from the present
study demonstrated that RGS10 was significantly downregulated in
ADR-treated podocytes. In addition, RGS10 expression was inhibited
by miR-199b-5p mimic and elevated by miR-199b-5p inhibitor. These
results indicated that RGS10 expression may be negatively
correlated with miR-199b-5p expression. The effects of RGS10 on
podocyte apoptosis in podocytes were further verified by performing
cell functional experiments. Overexpression of RGS10 could reverse
the promoting effects of miR-199b-5p on podocyte apoptosis. The
results suggested that miR-199b-5p could stimulate podocyte
apoptosis by inhibiting RGS10.
The AKT/mTOR signaling pathway is involved in cell
apoptosis in a variety of kidney diseases (31,32).
There is evidence that miR-199b-5p induces cell apoptosis via
directly targeting mTOR in endometrial endometrioid adenocarcinoma
(33) and negatively regulates
cell proliferation via the AKT signaling pathway in glioma
(34). These results show that
miR-199b-5p might play an important role in the activation of the
AKT/mTOR signaling pathway. A previous study reported that RGS10
silencing results in mTOR pathway activation in ovarian cancer
cells (35) and that RGS10
overexpression inhibits AKT phosphorylation and regulates cell
apoptosis in ovarian cancer cells (36). These findings indicate that RGS10
could exert biological functions by activating the AKT/mTOR
signaling pathway. Subsequently, both miR-199b-5p and RGS10 may be
involved in the activation of the AKT/mTOR signaling pathway. In
the present study, we demonstrated that miR-199b-5p overexpression
and RGS10 downregulation positively regulated the AKT/mTOR pathway,
and that treating cells with RAPA, which is an AKT/mTOR inhibitor,
could abolish the aggravated effects of the miR-199b-5p mimic on
ADR-induced apoptosis. Thus, these findings confirmed that
miR-199b-5p may activate the AKT/mTOR signaling pathway by
inhibiting RGS10 expression. However, it remains to be established
whether the present results could be also observed in animal
experiments and other cell lines.
In summary, the present study demonstrated that
miR-199b-5p may facilitate podocyte apoptosis via AKT/mTOR
signaling by targeting RGS10. miR-199b-5p may therefore be
considered as a potential target in the treatment of CKD in the
future.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural Science
Foundation of China (grant no. 81970664), the Natural Science
Foundation of Jiangsu Province (grant no. BK20191083), the 789
Outstanding Talent Program of SAHNMU (grant nos. 789ZYRC202080119
and 789ZYRC202090251) and the Science and Technology Development
Foundation of Nanjing Medical University (grant no.
NMUB2020052).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
AZ and WG designed and managed the study. GQ, TH and
AD performed the experiments. GQ, TH, AD, YZ, SL, HS and DG
contributed to the data collection and analysis. GQ, TH, AZ, AD and
WG drafted and finalized the manuscript. DG, SL, HS and AD
contributed to revision of the manuscript. All authors have read
and approved the final manuscript. GQ, TH and AZ confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kota SK and Kota SB: Noncoding RNA and
epigenetic gene regulation in renal diseases. Drug Discov Today.
22:1112–1122. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Chen L, Yang T, Lu D-W, Zhao H, Feng Y-L,
Chen H, Chen DQ, Vaziri ND and Zhao YY: Central role of
dysregulation of TGF-β/Smad in CKD progression and potential
targets of its treatment. Biomed Pharmacother. 101:670–681.
2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Webster AC, Nagler EV, Morton RL and
Masson P: Chronic Kidney Disease. Lancet. 389:1238–1252.
2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Regeniter A, Freidank H, Dickenmann M,
Boesken WH and Siede WH: Evaluation of proteinuria and GFR to
diagnose and classify kidney disease: Systematic review and proof
of concept. Eur J Intern Med. 20:556–561. 2009.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Zhou L, Chen X, Lu M, Wu Q, Yuan Q, Hu C,
Miao J, Zhang Y, Li H, Hou FF, et al: Wnt/β-catenin links oxidative
stress to podocyte injury and proteinuria. Kidney Int. 95:830–845.
2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Nagata M: Podocyte injury and its
consequences. Kidney Int. 89:1221–1230. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Liu M, Liang K, Zhen J, Zhou M, Wang X,
Wang Z, Wei X, Zhang Y, Sun Y, Zhou Z, et al: Sirt6 deficiency
exacerbates podocyte injury and proteinuria through targeting Notch
signaling. Nat Commun. 8(413)2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Assady S, Wanner N, Skorecki KL and Huber
TB: New Insights into Podocyte Biology in Glomerular Health and
Disease. J Am Soc Nephrol. 28:1707–1715. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Wang T, Gao Y, Wang X, Shi Y, Xu J, Wu B,
He J and Li Y: Calpain-10 drives podocyte apoptosis and renal
injury in diabetic nephropathy. Diabetes Metab Syndr Obes.
12:1811–1820. 2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Schiffer M, Bitzer M, Roberts IS, Kopp JB,
ten Dijke P, Mundel P and Böttinger EP: Apoptosis in podocytes
induced by TGF-β and Smad7. J Clin Invest. 108:807–816.
2001.PubMed/NCBI View
Article : Google Scholar
|
|
11
|
Hannon GJ: RNA interference. Nature.
418:244–251. 2002.PubMed/NCBI View
Article : Google Scholar
|
|
12
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Jaswani P, Prakash S, Dhar A, Sharma RK,
Prasad N and Agrawal S: MicroRNAs involvement in renal
pathophysiology: A bird's eye view. Indian J Nephrol. 27:337–341.
2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lorenzen JM, Kaucsar T, Schauerte C,
Schmitt R, Rong S, Hübner A, Scherf K, Fiedler J, Martino F,
Kumarswamy R, et al: MicroRNA-24 antagonism prevents renal ischemia
reperfusion injury. J Am Soc Nephrol. 25:2717–2729. 2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Liu Y, Bi X, Xiong J, Han W, Xiao T, Xu X,
Yang K, Liu C, Jiang W, He T, et al: MicroRNA-34a promotes renal
fibrosis by downregulation of Klotho in tubular epithelial cells.
Mol Ther. 27:1051–1065. 2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Long J, Wang Y, Wang W, Chang BH and
Danesh FR: MicroRNA-29c is a signature microRNA under high glucose
conditions that targets Sprouty homolog 1, and its in vivo
knockdown prevents progression of diabetic nephropathy. J Biol
Chem. 286:11837–11848. 2011.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Pang Q, Wang Y, Bi D and Lu H: LRRC75A-AS1
targets miR-199b-5p/PDCD4 axis to repress multiple myeloma. Cancer
Biol Ther. 21:1051–1059. 2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Lai Y, Quan J, Lin C, Li H, Hu J, Chen P,
Xu J, Guan X, Xu W, Lai Y, et al: miR-199b-5p serves as a tumor
suppressor in renal cell carcinoma. Exp Ther Med. 16:436–444.
2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Miao R, Lu Y, Xing X, Li Y, Huang Z, Zhong
H, Huang Y, Chen AF, Tang X, Li H, et al: Regulator of G-Protein
signaling 10 negatively regulates cardiac remodeling by blocking
mitogen-activated protein kinase–extracellular signal-regulated
protein kinase 1/2 signaling. Hypertension. 67:86–98.
2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Almutairi F, Lee J-K and Rada B: Regulator
of G protein signaling 10: Structure, expression and functions in
cellular physiology and diseases. Cell Signal.
75(109765)2020.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Li S, Liu Y, He X, Luo X, Shi H, Qu G, Wen
X, Gan W, Wang J and Zhang A: tRNA-Derived Fragments in Podocytes
with Adriamycin-Induced Injury Reveal the Potential Mechanism of
Idiopathic Nephrotic Syndrome. BioMed Res Int.
2020(7826763)2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Peng B, Hu Q, Liu X, Wang L, Chang Q, Li
J, Tang J, Wang N and Wang Y: Duchesnea phenolic fraction
inhibits in vitro and in vivo growth of cervical cancer through
induction of apoptosis and cell cycle arrest. Exp Biol Med
(Maywood). 234:74–83. 2009.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Xu LJ, Duan Y, Wang P and Yin HQ:
miR-199b-5p promotes tumor growth and metastasis in cervical cancer
by down-regulating KLK10. Biochem Biophys Res Commun. 503:556–563.
2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Ren L, Xu Y, Qin G, Liu C, Yan Y and Zhang
H: miR-199b-5p-Stonin 2 axis regulates metastases and
epithelial-to-mesenchymal transition of papillary thyroid
carcinoma. IUBMB Life. 71:28–40. 2019.PubMed/NCBI View
Article : Google Scholar
|
|
26
|
Wang H, Guo Y, Mi N and Zhou L: miR-101-3p
and miR-199b-5p promote cell apoptosis in oral cancer by targeting
BICC1. Mol Cell Probes. 52(101567)2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Kang WL and Xu GS: Atrasentan increased
the expression of klotho by mediating miR-199b-5p and prevented
renal tubular injury in diabetic nephropathy. Sci Rep.
6(19979)2016.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Alqinyah M, Almutairi F, Wendimu MY and
Hooks SB: RGS10 regulates the expression of cyclooxygenase-2 and
tumor necrosis factor alpha through a G protein-independent
mechanism. Mol Pharmacol. 94:1103–1113. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Cacan E, Ali MW, Boyd NH, Hooks SB and
Greer SF: Inhibition of HDAC1 and DNMT1 Modulate RGS10 Expression
and Decrease Ovarian Cancer Chemoresistance. PLoS One.
9(e87455)2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Yang S, Chen W, Stashenko P and Li Y-P:
Specificity of RGS10A as a key component in the RANKL signaling
mechanism for osteoclast differentiation. J Cell Sci.
120:3362–3371. 2007.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Xu J, Deng Y, Wang Y, Sun X, Chen S and Fu
G: SPAG5-AS1 inhibited autophagy and aggravated apoptosis of
podocytes via SPAG5/AKT/mTOR pathway. Cell Prolif.
53(e12738)2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Yingjie K, Haihong Y, Lingwei C, Sen Z,
Yuanting D, Shasha C, Liutong P, Ying W and Min Z: Apoptosis
repressor with caspase recruitment domain deficiency accelerates
ischemia/reperfusion (I/R)-induced acute kidney injury by
suppressing inflammation and apoptosis: The role of AKT/mTOR
signaling. Biomed Pharmacother. 112(108681)2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Cai J, Zhang Y, Huang S, Yan M, Li J, Jin
T and Bao S: miR-100-5p, miR-199a-3p and miR-199b-5p induce
autophagic death of endometrial carcinoma cell through targeting
mTOR. Int J Clin Exp Pathol. 10:9262–9272. 2017.PubMed/NCBI
|
|
34
|
Yang R, Yi L, Dong Z, Ouyang Q, Zhou J,
Pang Y, Wu Y, Xu L and Cui H: Tigecycline inhibits glioma growth by
regulating miRNA-199b-5p-HES1-AKT pathway. Mol Cancer Ther.
15:421–429. 2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Altman MK, Alshamrani AA, Jia W, Nguyen
HT, Fambrough JM, Tran SK, Patel MB, Hoseinzadeh P, Beedle AM and
Murph MM: Suppression of the GTPase-activating protein RGS10
increases Rheb-GTP and mTOR signaling in ovarian cancer cells.
Cancer Lett. 369:175–183. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Shelley B: Hooks, Phillip Callihan, Molly
K Altman, Jillian H Hurst, Mourad W Ali, Mandi M Murph. Regulators
of G-Protein signaling RGS10 and RGS17 regulate chemoresistance in
ovarian cancer cells. Mol Cancer. 9(289)2010.PubMed/NCBI View Article : Google Scholar
|