Introduction
Acute lung injury (ALI) is a diffuse inflammatory
reaction in the lung caused by various internal and external
pathogenic factors, such as sepsis, pneumonia and trauma (1,2).
ALI, which is characterized by respiratory distress, refractory
hypoxemia and respiratory failure, is the primary cause of death in
critically ill patients with sepsis at present (3). Bacterial infection, shock, severe
trauma, sepsis and other factors induce the occurrence of ALI
(4). Although the rapid
development of medical technology has provided better treatment of
ALI, the specific mechanism underlying its pathogenesis has not
been fully elucidated and there is a lack of effective drug
treatments in clinical use (5,6).
Dipeptidyl peptidase-4 (DPP4), also known as CD26,
is a widely expressed serine membrane-anchored peptidase that
exists on the surface of various types of cell (7). Its expression level varies from cell
to cell (8,9). In different organs and tissues (such
as lung, muscle and heart), DPP4 activity is associated with its
presence in the microvasculature (10). Clinical and experimental research
over the past 30 years has demonstrated the involvement of DPP4 in
various physiological processes and diseases of immune system
(11,12). Recent experimental studies have
shown that DPP4 inhibition protects the lungs against severe injury
and relieves associated respiratory disease, including COVID-19
caused by SARS-CoV-2 and Middle East respiratory syndrome (MERS)
caused by MERS coronavirus (13-17),
suggesting that DPP4 inhibitors may be used to decrease LI. A
previous study proposed that the DPP4 inhibitor Saxagliptin
attenuates lipopolysaccharide (LPS)-induced oxidative stress,
inflammation and apoptosis (18).
Another DPP4 inhibitor, vildagliptin, has been demonstrated to
alleviate pulmonary fibrosis in LPS-induced LI by inhibiting
endothelial-to-mesenchymal transition in pulmonary microvascular
endothelial cells (PMVECs) (19).
Anagliptin, a novel selective inhibitor of DPP4, was
licensed for clinical treatment of type 2 diabetes mellitus in
2012(20). Anagliptin has been
shown to alleviate inflammation and endothelial cell injury. For
example, anagliptin prevents H2O2-induced
apoptosis of human umbilical vein endothelial cells (21). Anagliptin ameliorates high
glucose-induced endothelial dysfunction via suppression of NLR
family pyrin domain containing 3 inflammasome activation (22). Anagliptin inhibits neointimal
hyperplasia via endothelial cell-specific modulation of superoxide
dismutase-1/Ras homolog family member A/JNK signaling in the
arterial wall (23). Recent
comprehensive review articles indicated that DPP4 inhibitors
(anagliptin, vildagliptin and sitagliptin) developed and marketed
for their beneficial effects display multipotency in the management
of various types of pulmonary disease (14,24).
Seys et al (17)
demonstrated that anagliptin displays a stronger anti-inflammatory
action than sitagliptin. Another study suggested that anagliptin
improves LI in mice under chronic stress, potentially by mitigating
vascular inflammation (25).
However, whether anagliptin relieves LPS-induced human (H)PMVEC
injury remains to be elucidated.
In the present study, the effect of anagliptin on
viability, inflammation, apoptosis and endothelial dysfunction of
HPMVECs exposed to LPS, along with its underlying mechanism, were
investigated. The present study aimed to provide a basis for the
use of anagliptin in ALI treatment.
Materials and methods
Cell culture and treatment
HPMVECs were obtained from American Type Culture
Collection (cat. no. CRL-3244) and cultured in RPMI-1640 medium
supplemented with 10% fetal bovine serum (FBS; both Gibco; Thermo
Fisher Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml
streptomycin (Sigma-Aldrich; Merck KGaA) at 37˚C with 5%
CO2. For LPS stimulation, cells were exposed to 100
ng/ml LPS (Sigma-Aldrich; Merck KGaA) at 37˚C for 24 h. For
anagliptin treatment, cells were exposed to various concentrations
of anagliptin (1, 10, 50 or 100 µM; Sigma-Aldrich; Merck KGaA) at
37˚C for 24 h. For LPS and anagliptin co-treatment, cells were
sequentially exposed to 100 ng/ml LPS plus designated
concentrations of anagliptin (10, 50 or 100 µM) at 37˚C for 24
h.
Cell viability assessment
MTT assay (Beyotime Institute of Biotechnology) was
utilized to detect cell viability. Briefly, HPMVECs were seeded
into 96-well plates (5x104 cells/well) and incubated at
37˚C to 90% confluence. Subsequently, cells were exposed to various
concentrations of anagliptin (1, 10, 50 or 100 µM) or LPS ±
anagliptin for 24 h. Then, 50 µl MTT solution was added to each
well and maintained for 3 h at 37˚C. Cells were exposed to 150 µl
DMSO and shaken on an orbital shaker for 15 min, then absorbance of
each well was measured at 590 nm.
Measurement of NO production
The generation of NO in culture medium was measured
using an NO assay kit (cat. no. S0023; Beyotime Institute of
Biotechnology) in accordance with manufacturer's protocol. Briefly,
cultured cells were harvested and centrifuged at 8,000 x g for 15
min at room temperature. The culture supernatant of cells was added
to 96-well plates (50 µl/well). After samples were incubated with
50 µl Griess Reagent for 3 min at room temperature, the absorbance
was measured at 540 nm.
Western blot analysis
HPMVECs were lysed using RIPA buffer (Beyotime
Institute of Biotechnology) containing cocktail inhibitors (Thermo
Fisher Scientific, Inc.) and quantified using a Bicinchoninic Acid
Protein Assay kit (Abcam). Samples (40 µg per lane) were separated
by 12% SDS-PAGE and then transferred to 0.45 µM PVDF membranes
(MilliporeSigma). After being blocked with 5% non-fat milk for 1.5
h at room temperature, samples were probed with primary antibodies
overnight at 4˚C and horseradish peroxidase-conjugated secondary
antibody (1:2,000; cat. no. 7074P2; Cell Signaling Technology,
Inc.) at room temperature for 2 h. Bands were visualized using ECL
(Beyotime Institute of Biotechnology) and quantified with ImageJ
software (Version 6.0; National Institutes of Health). The
following antibodies from Abcam or Cell Signaling Technology, Inc.,
were used (all at a dilution of 1:1,000): Anti-DPP4 (cat. no.
ab215711); anti-p65 (cat. no. ab32536), anti-Lamin B (cat. no.
17416S), anti-phosphorylated (p)-IκBα (cat. no. 2859T), anti-total
(t)-IκBα (cat. no. 4812S), anti-Bcl-2 (cat. no. 4223T), anti-Bax
(cat. no. 5023T), anti-apoptotic protease activating factor-1
(APAF-1; cat. no. 8969T), anti-cleaved-caspase3 (cat. no. 9664T),
anti-caspase3 (cat. no. 9662S), anti-AKT (cat. no. 4691T),
anti-p-AKT (cat. no. 4060T), anti-endothelial NO synthase (eNOS;
cat. no. 32027S), anti-inducible (i)NOS (cat. no. 20609S),
anti-high mobility group box 1 (HMGB1; cat. no. 6893S),
anti-receptor for advanced glycation end products (RAGE; cat. no.
6996S) and anti-GAPDH (cat. no. 5174T).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from cells using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. A total of 5 µg RNA
was reverse transcribed into cDNA using TaqMan One-Step RT kit
(Applied Biosystems; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. Amplification was performed using SYBR
Green PCR kit (Vazyme Biotech Co., Ltd) and ABI Prism 7500 sequence
detector (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
following thermocycling conditions were used for qPCR: Initial
denaturation for 10 min at 95˚C; followed by 40 cycles of
denaturation at 95˚C for 15 sec and annealing/extension at 55˚C for
45 sec. The primers were as follows: DPP4 forward,
5'-TCTGCTGAACAAAGGCAA TGA-3' and reverse,
5'-CTGTTCTCCAAGAAAACTGAGC-3'; tumor necrosis factor (TNF)-α
forward, 5'-CATCCAACCTT CCCAAACGC-3' and reverse,
5'-CGAAGTGGTGGTCTTGT TGC-3'; IL-1β forward, 5'-GAGCTCGCCAGTGAAATG
ATG-3' and reverse, 5'-TAGTGGTGGTCGGAGATTCG-3'; IL-6 forward,
5'-GTCCAGTTGCCTTCTCCCTG-3' and reverse, 5'-CTGAGATGCCGTCGAGGATG-3';
C-C motif chemokine ligand 2 (CCL2) forward, 5'-AGATCTGTGCTGAC
CCCAAG-3' and reverse, 5'-GGAGTTTGGGTTTGCTTG TCC-3'; and GAPDH,
forward, 5'-GCAACCGGGAAGGAAAT GAATG-3' and reverse,
5'-CCCAATACGACCAAATCAG AGA-3'. Results were normalized to GAPDH
expression and 2-ΔΔCq was used to calculate the relative
change in gene expression (26).
TUNEL staining
TUNEL assay was used to detect the apoptosis of
HPMVECs. Briefly, after cells were fixed with 4% paraformaldehyde
for 30 min at room temperature, apoptosis was detected using a
TUNEL assay kit (cat. no. QIA33; Sigma-Aldrich; Merck KGaA) in
accordance with the manufacturer's instructions; 50 µl TUNEL
reaction mixture was added for 1 h at 37˚C. The cells were treated
with DAPI (2 µg/ml) to stain the nucleus at 37˚C for 2-3 min. After
washing twice with PBS, images were captured from three fields of
view using an inverted fluorescence microscope (Olympus
Corporation; x200 magnification).
Wound healing assay
HPMVECs were cultured in 6-well plates
(5x105 cells/well) to 70-80% confluence. The cell
surface was scratched with a 100-µl pipette tip to create an
artificial wound, and medium was replaced with serum-free RPMI-1640
(Gibco; Thermo Fisher Scientific, Inc.) containing LPS ± anagliptin
and cultured for 24 h at 37˚C. Images were captured at 0 and 24 h
using an inverted light microscope (magnification, x100; Olympus
Corporation). The cell migration rate was calculated as follows:
(Width at 0 h-width at 24 h)/width distance. The relative migration
rate was obtained by normalizing to the untreated group.
Statistical analysis
Data are expressed as the mean ± standard deviation
and were analyzed with SPSS 21.0 software (IBM Corp). All
experiments were performed in triplicate. An unpaired Student's
t-test was used for comparisons between two groups. One-way
analysis of variance followed by Tukey's post hoc test was used for
multiple comparisons. P<0.05 was considered to indicate a
statistically significant difference.
Results
DPP4 expression is increased following
LPS stimulation in HPMVECs
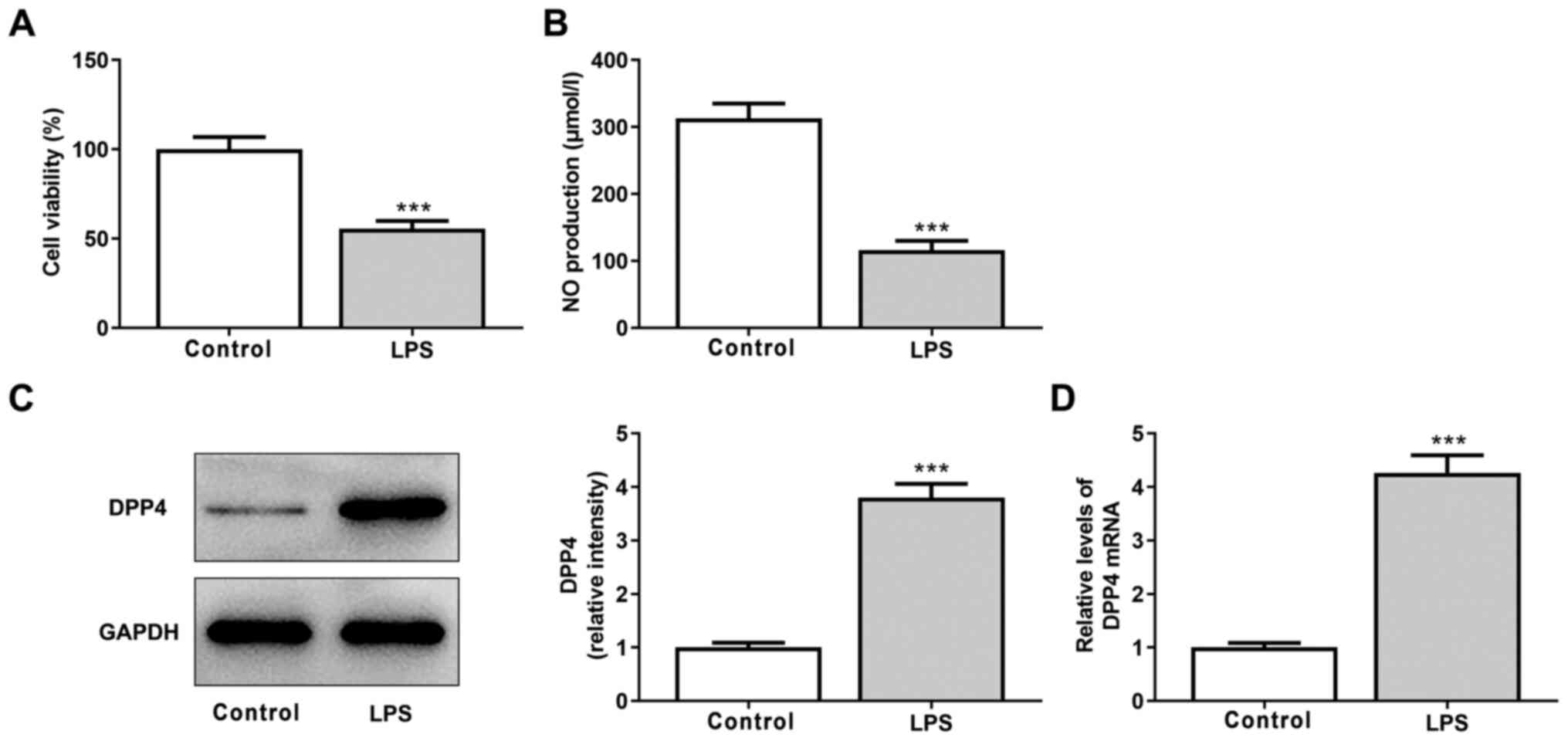
HPMVECs were exposed to 100 ng/ml LPS for 24 h to
simulate ALI in vitro. Viability and NO production of cells
significantly decreased following LPS stimulation, indicating that
LPS induced HPMVEC damage (Fig. 1A
and B). Furthermore, both protein
and mRNA expression levels of DDP4 were significantly increased
following LPS treatment of HPMVECs, suggesting that DDP4 expression
is increased in LPS-induced HPMVEC injury (Fig. 1C and D).
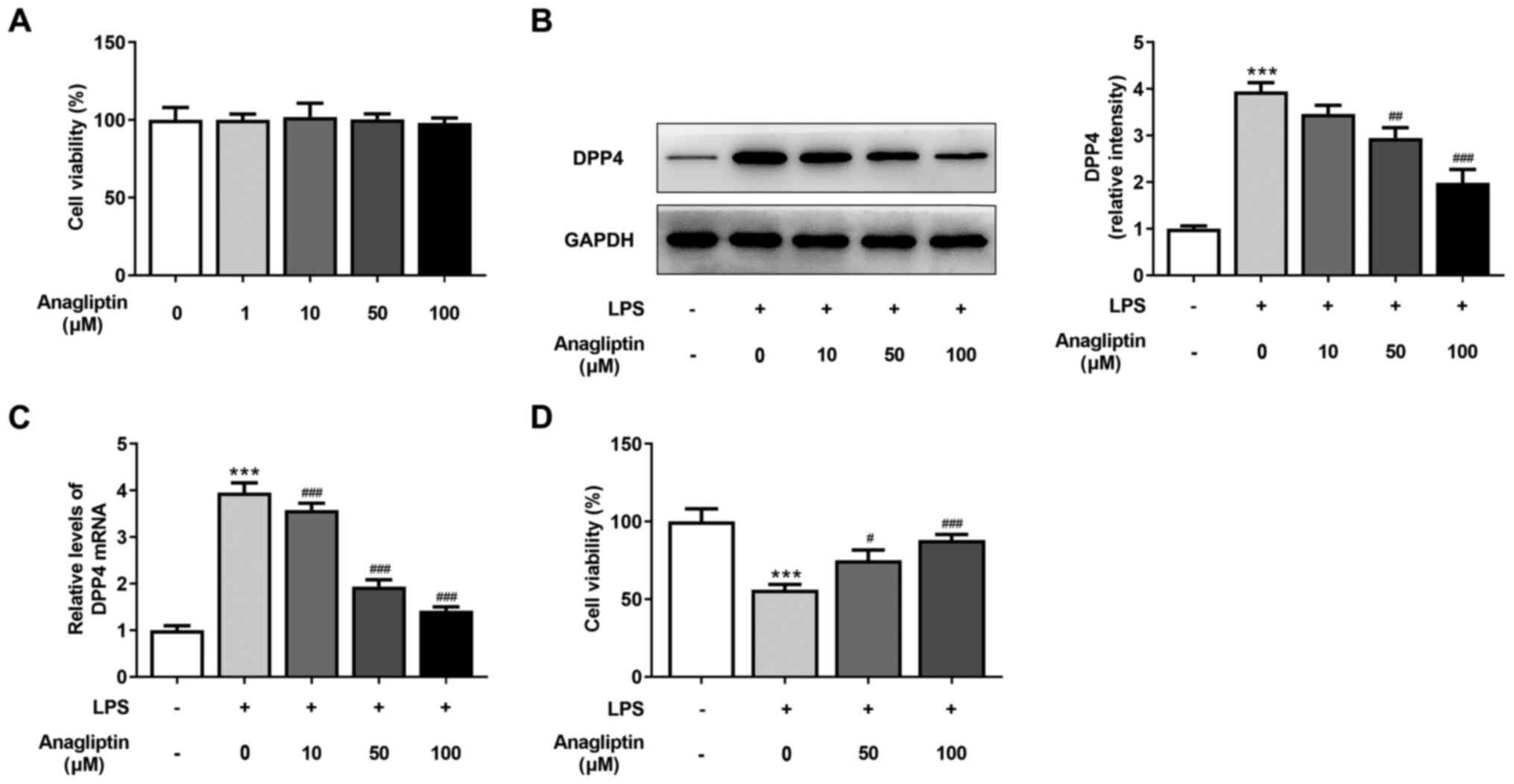
DPP4 inhibitor anagliptin inhibits
LPS-induced decrease in HPMVEC viability
Cells were stimulated with different concentrations
of anagliptin (0, 1, 10, 50 and 100 µM) for 24 h. Cell viability
was not altered following stimulation using different doses of
anagliptin (Fig. 2A). RT-qPCR and
western blot analysis were performed to detect DDP4 expression. The
results revealed that anagliptin (10 µM) decreased mRNA and protein
expression levels of DPP4, but this was not significantly different
compared with the LPS-alone group (Fig. 2B and C). Additionally, 50 or 100 µM anagliptin
significantly decreased DPP4 mRNA and protein levels (Fig. 2B and C) when compared to the LPS-alone group.
The LPS-induced impaired cell viability was rescued by co-treatment
with 50 and 100 µM anagliptin (Fig.
2D). These results reveal that the DPP4 inhibitor anagliptin
suppressed the LPS-induced decrease in HPMVEC viability.
DPP4 inhibitor anagliptin inhibits
LPS-induced inflammation and NF-κB activation in HPMVECs
LPS resulted in a significant increase in the
expression levels of pro-inflammatory cytokines, including TNF-α,
IL-1β, IL-6 and CCL2, but 50 and 100 µM anagliptin significantly
decreased the expression of these cytokines (Fig. 3A-D). Activation of NF-κB signaling
induces the inflammatory response (27). Following LSP treatment, NF-κB
signaling was activated, as demonstrated by the significant
increase in nuclear p65 expression and p/t-IκBα and the decrease in
cytoplasmic p65 in HPMVECs (Fig.
3E). However, following stimulation with 50 or 100 µM
anagliptin, nuclear p65 expression and p/t-IκBα expression were
effectively suppressed, while the expression of cytoplasmic p65 was
upregulated (Fig. 3E).
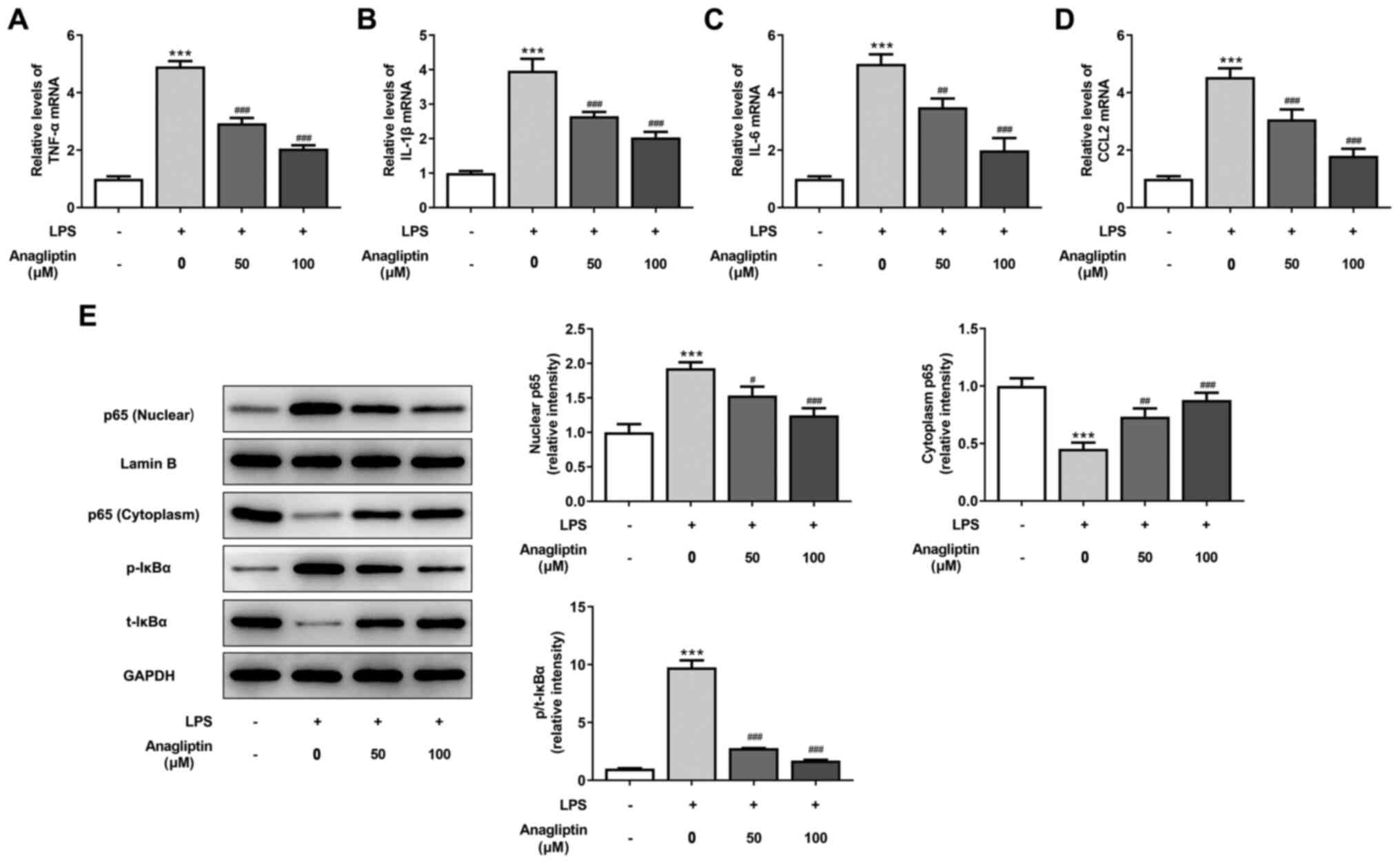 | Figure 3Anagliptin inhibits LPS-induced
inflammation and NF-κB p65 activation in HPMVECs. HPMVECs were
co-treated with 100 ng/ml LPS in the presence or absence of
different concentrations of anagliptin for 24 h, then mRNA levels
of (A) TNF-α, IL-(B) 1β, (C) IL-6 and (D) CCL2 were measured by
reverse transcription-quantitative PCR. (E) Protein expression of
nuclear and cytoplasmic p65 and p/t-IκBα was measured by western
blotting. ***P<0.001 vs. untreated cells;
#P<0.05, ##P<0.01 and
###P<0.001 vs. LPS alone. HPMVEC, human pulmonary
microvascular endothelial cell; LPS, lipopolysaccharide; TNF-α,
tumor necrosis factor-α; CCL2, C-C motif chemokine ligand 2; p-,
phosphorylated; t-, total. |
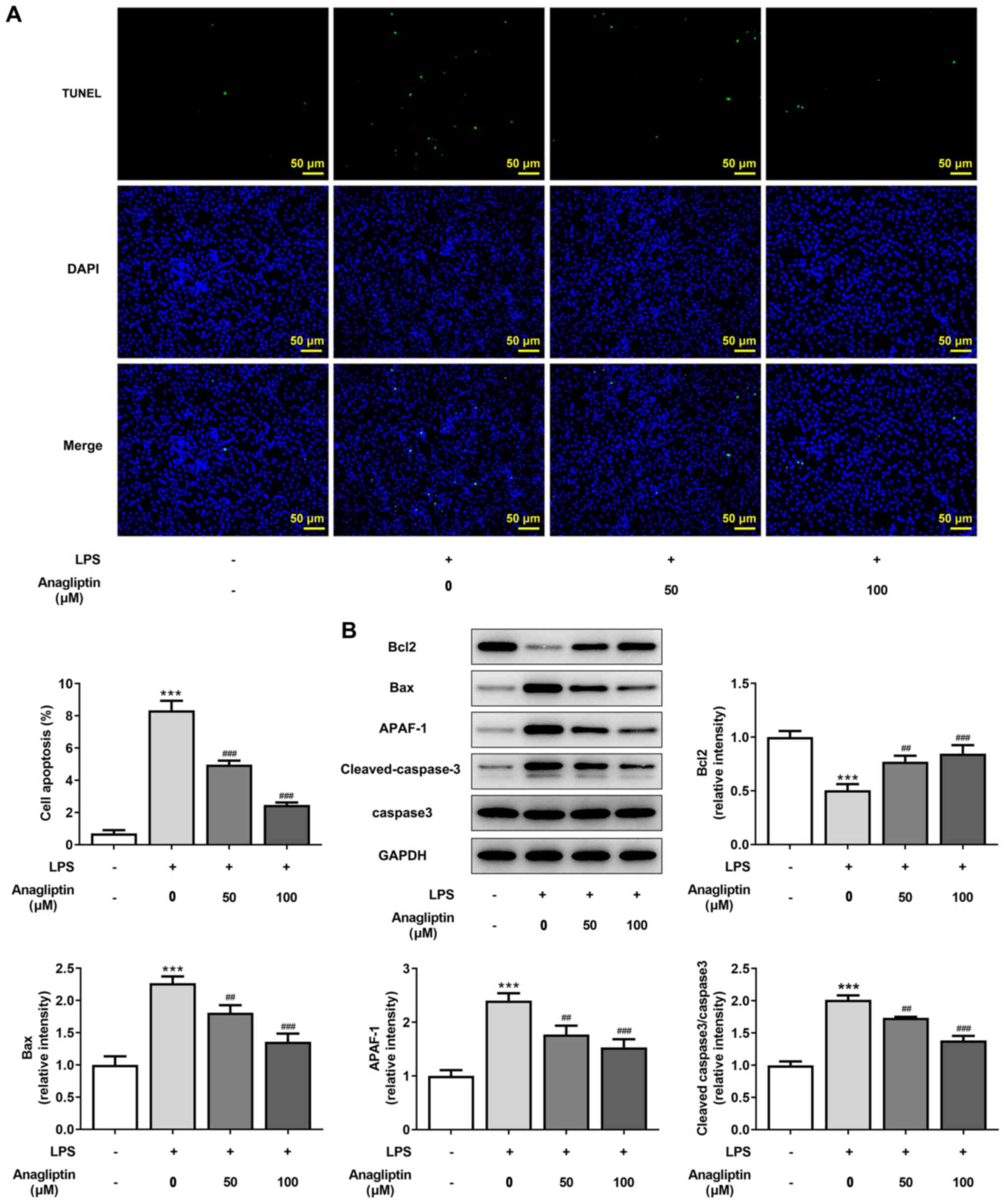
DPP4 inhibitor anagliptin inhibits
LPS-induced apoptosis in HPMVECs
TUNEL staining was used to assess cell apoptosis.
The number of apoptotic cells was significantly increased following
LPS treatment, whereas 50 and 100 µM anagliptin significantly
decreased this (Fig. 4A).
Consistently, LPS resulted in lower Bcl2 and higher Bax, APAF-1 and
cleaved-caspase-3/caspase-3 expression compared with the untreated
cells (Fig. 4B). Furthermore, 50
and 100 µM anagliptin enhanced Bcl2 but decreased Bax, APAF-1 and
cleaved-caspase-3/caspase-3 expression levels (Fig. 4B). These data suggest that the DPP4
inhibitor anagliptin inhibited LPS-induced apoptosis in
HPMVECs.
DPP4 inhibitor anagliptin rescues
migration and endothelial function and decreases HMGB1/RAGE
expression in LPS-treated HPMVECs
Impaired endothelial cell migration and NO
production are primary causes of endothelial dysfunction (28). LPS led to a significant decrease in
cell migration and NO production, which were effectively rescued by
treatment with 50 and 100 µM anagliptin (Fig. 5A-C). Moreover, 50 and 100 µM
anagliptin significantly upregulated the decreased expression
levels of p-AKT/AKT and eNOS and downregulated the increased
expression levels of iNOS induced by LPS (Fig. 5D). The enhanced protein and mRNA
expression levels of HMGB1 and RAGE induced by LPS exposure were
also significantly downregulated by 50 and 100 µM anagliptin,
indicating the inhibitory effect of anagliptin on HMGB1/RAGE
expression levels (Fig. 5E and
F).
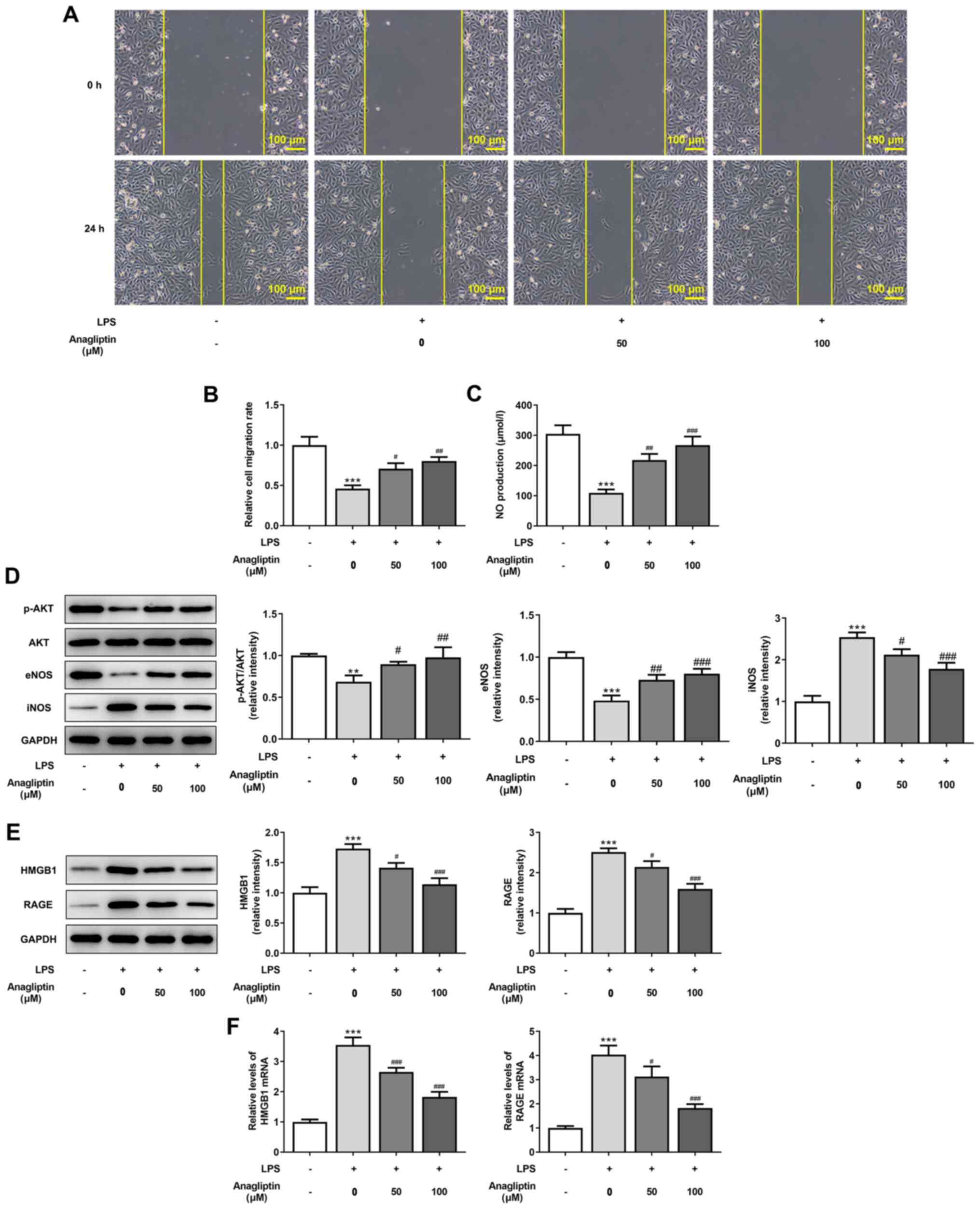 | Figure 5Anagliptin rescues cell migration and
inhibits HMGB1/RAGE expression in LPS-induced HPMVECs. HPMVECs were
co-treated with 100 ng/ml LPS in the presence or absence of
different concentrations of anagliptin for 24 h. (A and B) Wound
healing assay was utilized to assess cell migration. (C) NO
production was tested by NO assay kit. (D) Protein expression
levels of p-AKT/AKT, eNOS and iNOS and (E) HMGB1 and RAGE were
detected by western blotting. (F) mRNA levels of HMGB1 and RAGE
were measured by reverse transcription-quantitative PCR.
**P<0.01 and ***P<0.001 vs. untreated
cells; #P<0.05, ##P<0.01 and
###P<0.001 vs. LPS alone. HPMVEC, human pulmonary
microvascular endothelial cell; LPS, lipopolysaccharide; HMGB1,
high mobility group box 1; RAGE, receptor for advanced glycation
end products; NO, nitric oxide; p-, phosphorylated; eNOS,
endothelial NO synthase; i, inducible. |
Discussion
The alveolar capillary unit formed by PMVECs is the
basic structure that maintains ventilation-perfusion balance, and
is susceptible to harmful external stimuli, such as LPS (29). LPS is one of the pathogenic factors
leading to abnormal microcirculation in ALI (30,31).
In the present study, HPMVECs were exposed to LPS. The results
showed that LPS impaired cell viability, NO production and cell
migration and induced inflammation, NF-κB signaling activation and
apoptosis. However, anagliptin effectively protected HPMVECs
against LPS-induced injury, indicating that anagliptin may be used
in the treatment of ALI.
Uncontrolled inflammation is the primary
pathophysiological basis of ALI (32). The activation of NF-κB signaling
induces inflammatory response (27). The present study verified that LPS
induced expression of pro-inflammatory cytokines and activation of
NF-κB signaling, suggesting that inflammatory responses resulted
from LPS in HPMVECs. Additionally, the mechanism of ALI is
associated with increased vascular endothelial apoptosis and
recovery of endothelial cell viability is reported to improve ALI
(33). The present study
demonstrated an increase in apoptosis ratio and Bax, APAF-1 and
cleaved-caspase-3 expression levels, as well as a decrease in Bcl2
expression in LPS-induced cells. As a key molecule in the intrinsic
or mitochondrial pathway of apoptosis, APAF-1 leads to caspase-3
cleavage (34). Therefore, the
impaired cell viability caused by LPS may induce inflammation and
cell apoptosis. LPS decreased NO production and cell migration
along with p-AKT and eNOS expression, but increased iNOS
expression. Decreased NO, which is produced by eNOS, and impaired
endothelial cell migration are linked to endothelial dysfunction,
which results in an imbalance in vascular homeostasis, leading to a
prothrombotic and proinflammatory condition (35,36).
The AKT/eNOS pathway serves a key role in endothelial mobilization
and migration (37). The present
results showed that LPS caused endothelial dysfunction of HPMVECs
in vitro.
Anagliptin is a novel selective inhibitor of
DPP4(38). DPP4 inhibition has
been reported to prevent systemic inflammation, vascular
dysfunction and end-organ damage in mice with endotoxemia (39). In the study of ALI, DPP4 inhibitor
saxagliptin has been reported to decrease LPS-induced oxidative
stress, inflammation and apoptosis (18). Another DPP4 inhibitor,
vildagliptin, ameliorates pulmonary fibrosis in LPS-induced LI by
inhibiting endothelial-to-mesenchymal transition in PMVECs
(19). Whether anagliptin inhibits
LI is still unknown. The present study demonstrated that anagliptin
recovered DPP4 expression, rescued cell viability, inhibited NF-κB
activation-mediated inflammation and
Bcl2/Bax/APAF-1/caspase3-meditaed apoptosis in LPS-treated HPMVECs.
Furthermore, NO production and AKT/eNOS pathway-mediated cell
migration, which were impaired by LPS, were all markedly rescued by
anagliptin. These data indicated that anagliptin protected HPMVECs
against LPS-induced injury.
HMGB1 is a typical damage-associated molecular
pattern protein that exerts its biological activity primarily by
binding to RAGE. Anagliptin has been verified to suppress HMGB1
expression (40,41). Notably, LPS binds to HMGB1 to serve
a key role in endothelial dysfunction (42). The present results showed that LPS
significantly upregulated HMGB1 and RAGE expression levels, but
anagliptin effectively inhibited this effect. As a result, it was
speculated that anagliptin may exert its beneficial role in
LPS-induced HPMVEC injury via inhibiting LPS-mediated HMGB1/RAGE
upregulation.
To the best of our knowledge, the present study is
the first to investigate the effects of anagliptin on LPS-induced
ALI. Anagliptin alleviated LPS-induced HPMVEC injury by the
decreasing inflammation, apoptosis and endothelial dysfunction via
inhibiting LPS-mediated HMGB1/RAGE upregulation. However, the
specific mechanisms involved in the action of anagliptin need to be
clarified in subsequent experiments. In addition, safety evaluation
and pharmacokinetic studies of anagliptin should be performed in
future. These are limitations of the present study and
comprehensive and in-depth analysis will be conducted in future to
provide further evidence for the treatment of ALI using
anagliptin.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Author's contributions
JZ and LL contributed to study conception and design
and acquisition, analysis and interpretation of data. JZ drafted
the manuscript. LL revised the manuscript critically for important
intellectual content. Both authors have read and approved the final
manuscript. JZ and LL confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dushianthan A, Grocott MP, Postle AD and
Cusack R: Acute respiratory distress syndrome and acute lung
injury. Postgrad Med J. 87:612–622. 2011.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Ali H, Khan A, Ali J, Ullah H, Khan A, Ali
H, Irshad N and Khan S: Attenuation of LPS-induced acute lung
injury by continentalic acid in rodents through inhibition of
inflammatory mediators correlates with increased Nrf2 protein
expression. BMC Pharmacol Toxicol. 21(81)2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Butt Y, Kurdowska A and Allen TC: Acute
Lung Injury: A Clinical and Molecular Review. Arch Pathol Lab Med.
140:345–350. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Hughes KT and Beasley MB: Pulmonary
Manifestations of Acute Lung Injury: More Than Just Diffuse
Alveolar Damage. Arch Pathol Lab Med. 141:916–922. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Hu Q, Wang Q, Han C and Yang Y: Sufentanil
attenuates inflammation and oxidative stress in sepsis-induced
acute lung injury by downregulating KNG1 expression. Mol Med Rep.
22:4298–4306. 2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Schmidt GA: Managing Acute Lung Injury.
Clin Chest Med. 37:647–658. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Piao L, Li Y, Narisawa M, Shen X and Cheng
XW: Role of Dipeptidyl Peptidase-4 in Atherosclerotic
Cardiovascular Disease in Humans and Animals with Chronic Stress.
Int Heart J. 62:470–478. 2021.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Yip HK, Lee MS, Li YC, Shao PL, Chiang JY,
Sung PH, Yang CH and Chen KH: Dipeptidyl Peptidase-4 deficiency
effectively protects the brain and neurological function in rodent
after acute Hemorrhagic Stroke. Int J Biol Sci. 16:3116–3132.
2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lee M, Shin E, Bae J, Cho Y, Lee JY, Lee
YH, Lee BW, Kang ES and Cha BS: Dipeptidyl peptidase-4 inhibitor
protects against non-alcoholic steatohepatitis in mice by targeting
TRAIL receptor-mediated lipoapoptosis via modulating hepatic
dipeptidyl peptidase-4 expression. Sci Rep.
10(19429)2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Klemann C, Wagner L, Stephan M and von
Hörsten S: Cut to the chase: A review of CD26/dipeptidyl
peptidase-4's (DPP4) entanglement in the immune system. Clin Exp
Immunol. 185:1–21. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zhao Y: CD26 in autoimmune diseases: The
other side of ‘moonlight protein’. Int Immunopharmacol.
75(105757)2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wang X, Ke J, Zhu YJ, Cao B, Yin RL, Wang
Y, Wei LL, Zhang LJ, Yang LY and Zhao D: Dipeptidyl peptidase-4
(DPP4) inhibitor sitagliptin alleviates liver inflammation of
diabetic mice by acting as a ROS scavenger and inhibiting the NFκB
pathway. Cell Death Discov. 7(236)2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kawasaki T, Chen W, Htwe YM, Tatsumi K and
Dudek SM: DPP4 inhibition by sitagliptin attenuates LPS-induced
lung injury in mice. Am J Physiol Lung Cell Mol Physiol.
315:L834–L845. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Solerte SB, Di Sabatino A, Galli M and
Fiorina P: Dipeptidyl peptidase-4 (DPP4) inhibition in COVID-19.
Acta Diabetol. 57:779–783. 2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Strollo R and Pozzilli P: DPP4 inhibition:
Preventing SARS-CoV-2 infection and/or progression of COVID-19?
Diabetes Metab Res Rev. 36(e3330)2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zou H, Zhu N and Li S: The emerging role
of dipeptidyl-peptidase-4 as a therapeutic target in lung disease.
Expert Opin Ther Targets. 24:147–153. 2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Seys LJM, Widagdo W, Verhamme FM, Kleinjan
A, Janssens W, Joos GF, Bracke KR, Haagmans BL and Brusselle GG:
DPP4, the Middle East Respiratory Syndrome Coronavirus Receptor, is
Upregulated in Lungs of Smokers and Chronic Obstructive Pulmonary
Disease Patients. Clin Infect Dis. 66:45–53. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Guo K and Jin F: Dipeptidyl peptidase-4
(DPP-4) inhibitor saxagliptin alleviates lipopolysaccharide-induced
acute lung injury via regulating the Nrf-2/HO-1 and NF-κB pathways.
J Invest Surg. 34:695–702. 2021.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Suzuki T, Tada Y, Gladson S, Nishimura R,
Shimomura I, Karasawa S, Tatsumi K and West J: Vildagliptin
ameliorates pulmonary fibrosis in lipopolysaccharide-induced lung
injury by inhibiting endothelial-to-mesenchymal transition. Respir
Res. 18(177)2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Roussel R, Duran-García S, Zhang Y, Shah
S, Darmiento C, Shankar RR, Golm GT, Lam RLH, O'Neill EA, Gantz I,
et al: Double-blind, randomized clinical trial comparing the
efficacy and safety of continuing or discontinuing the dipeptidyl
peptidase-4 inhibitor sitagliptin when initiating insulin glargine
therapy in patients with type 2 diabetes: The CompoSIT-I Study.
Diabetes Obes Metab. 21:781–790. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhao X, Sun J, Chen Y, Su W, Shan H, Li Y,
Wang Y, Zheng N, Shan H and Liang H: lncRNA PFAR Promotes Lung
Fibroblast Activation and Fibrosis by Targeting miR-138 to Regulate
the YAP1-Twist Axis. Mol Ther. 26:2206–2217. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Jiang T, Jiang D, Zhang L, Ding M and Zhou
H: Anagliptin ameliorates high glucose- induced endothelial
dysfunction via suppression of NLRP3 inflammasome activation
mediated by SIRT1. Mol Immunol. 107:54–60. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Li Q, Zhang M, Xuan L, Liu Y and Chen C:
Anagliptin inhibits neointimal hyperplasia after balloon injury via
endothelial cell-specific modulation of SOD-1/RhoA/JNK signaling in
the arterial wall. Free Radic Biol Med. 121:105–116.
2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Pantanetti P, Cangelosi G and Ambrosio G:
Potential role of incretins in diabetes and COVID-19 infection: A
hypothesis worth exploring. Intern Emerg Med. 15:779–782.
2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Zhang S, Li P, Xin M, Jin X, Zhao L, Nan Y
and Cheng XW: Dipeptidyl peptidase-4 inhibition prevents lung
injury in mice under chronic stress via the modulation of oxidative
stress and inflammation. Exp Anim. 21-0067:2021.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Hoesel B and Schmid JA: The complexity of
NF-κB signaling in inflammation and cancer. Mol Cancer.
12(86)2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Marcelo KL, Goldie LC and Hirschi KK:
Regulation of endothelial cell differentiation and specification.
Circ Res. 112:1272–1287. 2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Pan X, Xu S, Zhou Z, Wang F, Mao L, Li H,
Wu C, Wang J, Huang Y, Li D, et al: Fibroblast growth factor-2
alleviates the capillary leakage and inflammation in sepsis. Mol
Med. 26(108)2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Wang T, Yegambaram M, Gross C, Sun X, Lu
Q, Wang H, Wu X, Kangath A, Tang H, Aggarwal S, et al: RAC1
nitration at Y32 IS involved in the endothelial barrier
disruption associated with lipopolysaccharide-mediated acute lung
injury. Redox Biol. 38(101794)2021.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Liu X, Wang D, Zhang X, Lv M, Liu G, Gu C,
Yang F and Wang Y: Effect and mechanism of phospholipid scramblase
4 (PLSCR4) on lipopolysaccharide (LPS)-induced injury to human
pulmonary microvascular endothelial cells. Ann Transl Med.
9(159)2021.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Lin J, Lin Z and Lin L: miR-490 alleviates
sepsis-induced acute lung injury by targeting MRP4 in new-born
mice. Acta Biochim Pol. 68:151–158. 2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wang Y, Chen H, Li H, Zhang J and Gao Y:
Effect of angiopoietin-like protein 4 on rat pulmonary
microvascular endothelial cells exposed to LPS. Int J Mol Med.
32:568–576. 2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Shakeri R, Kheirollahi A and Davoodi J:
Apaf-1: Regulation and function in cell death. Biochimie.
135:111–125. 2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Cyr AR, Huckaby LV, Shiva SS and
Zuckerbraun BS: Nitric Oxide and Endothelial Dysfunction. Crit Care
Clin. 36:307–321. 2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Zheng X, Zhang W and Wang Z: Simvastatin
preparations promote PDGF-BB secretion to repair LPS-induced
endothelial injury through the PDGFRβ/PI3K/Akt/IQGAP1 signalling
pathway. J Cell Mol Med. 23:8314–8327. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Everaert BR, Van Craenenbroeck EM, Hoymans
VY, Haine SE, Van Nassauw L, Conraads VM, Timmermans JP and Vrints
CJ: Current perspective of pathophysiological and interventional
effects on endothelial progenitor cell biology: Focus on
PI3K/AKT/eNOS pathway. Int J Cardiol. 144:350–366. 2010.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Kang SM, Jung HS, Kwon MJ, Lee SH and Park
JH: Effects of anagliptin on the stress induced accelerated
senescence of human umbilical vein endothelial cells. Ann Transl
Med. 9(750)2021.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Steven S, Jurk K, Kopp M, Kröller-Schön S,
Mikhed Y, Schwierczek K, Roohani S, Kashani F, Oelze M, Klein T, et
al: Glucagon-like peptide-1 receptor signalling reduces
microvascular thrombosis, nitro-oxidative stress and platelet
activation in endotoxaemic mice. Br J Pharmacol. 174:1620–1632.
2017.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Ma Y, Wang J, Wang C, Zhang Q, Xu Y, Liu
H, Xiang X and Ma J: DPP-4 inhibitor anagliptin protects against
hypoxia-induced cytotoxicity in cardiac H9C2 cells. Artif Cells
Nanomed Biotechnol. 47:3823–3831. 2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Sato A, Suzuki S, Watanabe S, Shimizu T,
Nakamura Y, Misaka T, Yokokawa T, Shishido T, Saitoh SI, Ishida T,
et al: DPP4 Inhibition Ameliorates Cardiac Function by Blocking the
Cleavage of HMGB1 in Diabetic Mice After Myocardial Infarction. Int
Heart J. 58:778–786. 2017.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Liu Z, Wang J, Xing W, Peng Y, Quan J and
Fan X: LPS binding to HMGB1 promotes angiogenic behavior of
endothelial cells through inhibition of p120 and CD31 via
ERK/P38/Src signaling. Eur J Cell Biol. 96:695–704. 2017.PubMed/NCBI View Article : Google Scholar
|