1. Introduction
Cervical cancer is a common cause of morbidity and
mortality among women worldwide, with 604,000 new diagnosed cases
and ~342,000 mortalities reported in 2020 alone (1). Epidemiological data confirm that
persistent infections with high-risk (hr) genotypes of human
papillomavirus (HPV) are associated with cervical cancer
development. hrHPVs have the potential to induce carcinogenesis
driven by E6 and E7 viral oncoproteins (2). In sexually active women, acute hrHPV
infections are common and the majority resolve within 1-2 years
from onset without clinical consequences. A small percentage of
these infections persist and develops into cancer (3). However, the progression from
hrHPV-induced cervical lesion to cervical cancer is slow, taking
years or even decades and occurs through well-defined cytological
abnormalities. Thus, timely identification of such lesions is the
goal of cervical screening (3).
The long process of cervical cancer development allows periodic
screening tests to detect preneoplastic cellular changes
sufficiently early in order to prevent the onset of this
malignancy. Cervical cancer can therefore be reduced or even
prevented through screening programs that aim to diminish the
incidence and mortality of this type of cancer (4). Epigenetic modifications, as hallmarks
of carcinogenesis, can serve as a powerful tool for risk
stratification, prognosis and treatment of hrHPV-induced cervical
lesions in screening programs. For example, there are clinically
used panels that assess DNA hypermethylation or hypomethylation
associated with oncogene activation and genomic instability for
breast cancer. Such kits can be developed and used for cervical
cancer early detection, prognosis and treatment management,
particularly because such epigenetic modifications are reversible
(5).
2. Typical cervical screening and the
necessity of designing novel efficient assays
Traditional methods are available to screen women
for cervical precancer and cancer, including cytology-based
screening, visual examination of the cervix, colposcopy,
cervicography and HPV DNA test. Each screening test has its own
strengths and limitations and the selection of test will depend on
the context (lesion severity, socio-economic factors, sample
availability and invasiveness).
Cytological screening
In 1927, the Romanian pathologist Aurel Babes noted
the usefulness of exfoliated cells from the cervix in the detection
of cervical cancer (6).
Subsequently implemented by Papanicolaou and Traut under the name
of the Papanicolaou (Pap) test, this method has been used since
1960s in national cervical screening programs in Europe and USA
(7). In developed countries that
have introduced cytology-based screening programs, the incidence
and mortality associated with cervical cancer have been markedly
reduced and ≤2,000 new cases of cervical cancer are prevented per
month (8). At present, the
majority of cervical intraepithelial neoplasia (CIN) that may lead
to malignancies are detected with Pap cytology screening programs.
However, applying such a screening method and preserving a high
quality, particularly in developing countries, is markedly
difficult (9). The data published
over the last two decades reveal the high quality of Pap cytology,
but they also highlight that >30% of CIN2, CIN3 and invasive
carcinoma are not distinguished (10). Cytological testing has
disadvantages determined by the quality of the smear and the
experience of the cytopathologist, which contribute to the decrease
in sensitivity of this method. The diagnosis of cervical cancer by
Pap test is however limited due to its lack of sensitivity in
identifying precancerous lesions and its increased specificity,
which leads to over-diagnosis and over-treatment, mainly among
young women (11). Therefore,
alternative screening methods and algorithms to provide early,
sensitive and specific information regarding precursor lesions are
evaluated in the present review.
HPV testing
Based on the role of HPV infection in cervical
epithelium transformation and cervical cancer development, a new
approach for screening has been developed by using HPV oncogenic
genotypes testing, which has already been implemented in certain
developed countries, such as the USA. The increased sensitivity and
limited specificity of the HPV test do not allow discrimination
between women with transient, self-limiting infection and those at
risk of developing neoplastic lesions. HPV DNA detection does not
necessarily indicate a pre-cancerous lesion or cancer. However,
since the immune system fights against the majority of HPV
infections and prevents the development of cervical precursor
lesions or carcinomas, the main challenge is the management of
HPV-positive results in primary screening (12). Performing HPV testing as often as
Pap screening in women who do not present CIN2 or CIN3 would
produce more positive results than cytological screening (10). Furthermore, in a joint European
cohort study, an extended screening interval to 6 years was
recommended for women with a negative HPV test (13).
HPV testing has various limitations for screening,
including low specificity and reproducibility, as well as high
cost, considering the different types of tests used. Additionally,
HPV testing as a primary screening should be accompanied by a
highly specific second-line test, so that recommendations for
additional examinations (colposcopy) are reduced (11). Thus, in numerous countries,
screening strategies are based on co-testing (HPV and Pap testing).
For instance, the American Cancer Society (ACS) and the American
Society for Colposcopy and Cervical Pathology (ASCCP) recommend
either co-testing every 5 years or cytology screening every 3 years
for women aged 30-65 years. Since in young women (21-30 years) HPV
infection is more frequent, ACS and ASCCP recommend performing
cytology screening alone (14).
Currently, hrHPV testing is a more effective method of screening
precancerous lesions and carcinomas than cytology-based assay, as
the process relies on the detection of nucleic acids rather than
microscopic examination of the lesions (15).
To date, neither cytology nor HPV testing provide
sufficient specificity and sensitivity as a basis for screening.
Thus, there is still a need for developing a screening strategy
that is based on objective, easily reproducible and accurate
methods (16). In this regard,
viral proteins have received attention and several commercial
assays have been accordingly developed, which suggests that they
may have improved diagnostic capacities and may allow an improved
discrimination between productive and transforming infections. In
addition, these methods may be less invasive and more cost
effective than hrHPV testing (17).
New approaches in cancer
diagnosis
Classic diagnostic approaches for tumor subtyping
rely on tissue biopsies and histological analysis of tumor
specimens. These methods can provide an incomplete image of the
cellular composition of the tumor due to sampling a small
subpopulation of the cell types in a tumor and often miss the
aspect of heterogeneity. Neoteric epigenetic methods may use
samples obtained by using such techniques, or fresh frozen or
formalin-fixed paraffin-embedded (FFPE) tissue, thus providing a
more comprehensive analysis of the modifications occurring at a
cellular level (18). Novel
epigenetic methods can also be applied to liquid biopsies, thus
offering information on tumorigenesis, growth, immune-cancer
interactions and cell death by analyzing circulating tumor cells
and microvesicles (exosomes). Another advantage of this type of
assays is their ability to analyze circulating tumor DNA (ctDNA),
which can better illustrate the heterogeneity and clonality of the
tumor cell populations. Since it is likely that all cells
contribute to this type of cell-free DNA (cfDNA) (19). Analyzing the mutational and
epigenetic profiles without special handling of the samples while
using non-invasive or minimally invasive sampling techniques,
further illustrates the advantages of such novel approaches in
cancer diagnosis (20).
A previous study on the molecular mechanisms that
could contribute to the onset and progression of cervical cancer
have shown that the theory of genetic mutations alone cannot
explain the development of tumors (21). Research in the field of epigenetic
changes has shown that aberrant methylation of cellular DNA is a
common alteration in cancer and hypermethylation of specific DNA
regions during carcinogenesis could serve as a sensitive screening
tool (22), particularly because
distinct methylation patterns of tumor suppressor genes have been
found in HPV-induced tumors (23).
Epigenetic markers associated with molecular events occurring in
HPV-induced oncogenesis can be used to triage women at risk of
developing cervical cancer. Consequently, cervical screening is
undergoing major changes with the development of new screening
tests with improved diagnostic performances using the same clinical
specimens as those employed for cytology and HPV testing. The
specificity of methylation-based tests could be enhanced by
identifying the specific pattern of each type of lesion (24). On the other hand, the performance
of screening based on HPV detection could be increased through
co-testing of epigenetic changes. According to Cuzick (24), due to the lack of clear indicators
on whom to screen for cervical cancer, the necessity for a more
accurate initial triaging test is increased. Thus, combining HPV
genotyping, cytology screening and detection of p16 and methylation
levels through less invasive methods may contribute to the
development of more efficient approaches for cancer diagnosis
(24).
3. Epigenetic alterations in human
tumors
DNA methylation
DNA methylation is one of the three different
epigenetic mechanisms that have been identified to date. Catalyzed
by DNA methyltransferase enzymes (DNMTs), DNA methylation often
occurs at CpG islands, when a methyl group is transferred from
S-adenyl methionine to the fifth carbon of a cytosine residue, thus
generating 5-methylcytosine (5-mC) (25). There are four members in the DNMT
family, including DNMT1, DNMT3A, DNMT3B and DNMT3L. DNMT1 encodes
for the maintenance methyltransferase, which is responsible for the
transfer of methylation patterns to newly synthesized DNA strands
following DNA replication, while DNMT3A and DNMT3B are involved in
de novo methylation and maintenance in addition to DNMT1
(26,27). Furthermore, the oxidation of 5-mC
leads to 5-hydroxymethylcytosine, 5-formylcytosine and
5-carboxylcytosine in the presence of Ten-eleven translocation
(TET) enzymes (TET1, TET2 and TET3) in a step-by-step reaction,
which indicates that methylation is a reversible process (28). DNA methylation that involves CpG
sites in the regulatory sequences leads to gene silencing, while
active genes have hypomethylated or unmethylated sites (29). In addition, in almost all types of
carcinoma, the genomes of tumor cells exhibit decreased levels of
5-mC, while regulatory sites display increased levels of 5-mC
(30). Additionally, the molecular
effectors and enzymes involved in DNA methylation process can
suffer cancer-specific alterations, such as those shown in Table I (31).
 | Table ISeveral DNA methylation genes altered
in various cancer types. |
Table I
Several DNA methylation genes altered
in various cancer types.
| Cancer type | DNA methylation
genes with modified expression |
|---|
| Breast cancer | DNMT3a, DNMT3b,
DNMT3L, TET |
| Cervical
cancer | DNMT3L |
| Colorectal
cancer | MBD1, MBD2, MDB3,
MBD4, Kaiso |
| Glioma | TET1, TET2,
TET3 |
| Lung cancer | MBD1, MBD2, MDB3,
Kaiso, TET |
| Ovarian cancer | DNMT1, DNMT3a |
| Pancreatic
cancer | TET |
| Prostate
cancer | MeCP2, MBD1, MBD2,
TET |
Methylated genes in cancer
Previous studies have highlighted the important role
served by DNA methylation in various types of cancer (32-39).
Genes that perform different functions (oncogenes or tumor
suppressors) are frequently targeted and, as a result, their
expression profile is altered, particularly in the transformation
process. A common phenomenon in cancer is the hypermethylation of
promoters of tumor suppressor genes, which leads to a decrease in
their expression pattern. To date, numerous methylated gene panels
have been identified and validated as biomarkers for various
pathologies (Table II).
| Table IISeveral histone modification genes
altered in various cancers. |
Table II
Several histone modification genes
altered in various cancers.
| Authors, year | Cancer type | Methylated
genes | Refs. |
|---|
| Reinert, 2012 | Bladder cancer | ZNF154, HOXA9,
POU4F2, TWIST1, VIM | (32) |
| de Groot et
al, 2014 | Breast cancer | SLC5A8, AKR1B1,
ALX1, GPX7, RASSGRF2, SFRP2, TM6SF1 and TMEFF2 | (33) |
| Yi, 2021 | Colorectal
cancer | TFPI2, FBN2, SEPT9,
SMAP8, MLH1, CDH1, TIMP3, O6-MGMT, SFRP1, SFRP2, p16, APC, HIC1,
CHFR | (34) |
| Etcheverry et
al, 2010; LeBlanc and Marra, 2016 | Glioblastoma | SLC5A8, MGMT,
SOX10, RUNX3, WIF1, CD133, HTATIP2, PDE4C, TES | (35,36) |
| Zhang et al,
2016 | Hepatocellular
carcinoma | RASSF1A, APC,
GSTP1, CDH1, p15, RUNX3, SOCS1, SFRP1, PRDM2, p14, RARβ and
p73 | (37) |
| Shen et al,
2019 | Lung cancer | SLC5A8, HOXA9,
KRTAP8-1, CCND1, TULP2 | (38) |
| Yang and Park,
2012 | Prostate
cancer | CAV1, CDKN2A,
CCND2, DAPK, HIC1, LPL, PITX2, PTGS2, RASSF1A, SLC5A8 | (39) |
Methylated genes specific to cervical
cancer
There have been a number findings on a series of
methylations in gene promoter sequences associated with the
pathogenesis and progression of cervical cancer. In addition,
epigenetic modifications have been reported to occur earlier than
genetic alterations in cervical cancer. In addition, abnormal DNA
methylation could occur as early as in low-grade intraepithelial
lesion (LSIL), indicating the potential application of abnormal DNA
methylation in the early diagnosis of cervical cancer alone or
combined with the existing monitoring methods. The detection of
altered DNA methylation at this stage serves an important role as
LSIL can either regress or progress to higher grade lesions
(40).
At present, the DNA methylation sites that have been
demonstrated to have a possible association with the early
pathogenesis of cervical cancer include cell adhesion molecule 1,
junctional adhesion molecule B, Ras association domain family
member 1 and fragile histidine triad and hypermethylation of some
of them has been detected in blood, urine and exfoliative cell
samples (40). Currently, the gene
methylation sites that have been validated to be possibly
associated with the prognosis of cervical cancer include
anaphase-promoting complex subunit 1, chromodomain helicase DNA
binding protein 1, voltage-dependent calcium channel subunit
α-2/δ-2, Dickkopf-3 and Cyclin Dependent Kinase Inhibitor 2A
(41). A previous study showed a
strong correlation between host and viral genome methylation and
the severity of cervical lesions (CIN2/CIN3) and cervical
carcinoma, respectively (42). The
most commonly reported host genes exhibiting promoter methylation
include erythrocyte membrane protein band 4.1 like 3 (EPB41L3),
myelin and lymphocyte protein, cell adhesion molecule (CADM),
family with sequence similarity 19 [chemokine (C-C motif)-like]
member A4 (FAM19A4) and microRNA (miRNA/miR) 124, as well as CpG
sites in the late regions of various HPV genomes (43). Among the genes that display higher
levels of methylation in higher grade lesions adenylate cyclase
activating polypeptide 1, achaete-scute family BHLH transcription
factor 1, CADM1, deleted in colorectal carcinoma, ATPase
phospholipid transporting 10A, deleted in breast cancer 1, heparan
sulfate-glucosamine 3-sulfotransferase 2, proto-oncogene
serine/threonine-protein kinase mos, SRY-box transcription factor
(SOX1), myogenic differentiation 1 (MYOD1), SOX17 and transmembrane
protein with EGF like and two follistatin like domains 2 are
included (44). In addition, LIM
homeobox 8 and ST6 N-acetylgalactosaminide α-2,6-sialyltransferase
5 exhibit increased DNA methylation in cervical cancer (45). Verlaat et al (46) identified novel potential biomarkers
including ankyrin repeat domain 18C pseudogene, chromosome 13 open
reading frame 18, junctional adhesion molecule 3 (JAM3), zinc
finger and SCAN domain containing 1, growth hormone secretagogue
receptor (GHSR), somatostatin (SST), zinc family member 1 (ZIC1),
phosphatase and actin regulator 3 (PHACTR3) and PR-domain
containing protein 14 (PRDM14)] that display increased levels of
methylation as lesions progress from precancerous to cancerous
driven by all hrHPV types. In an exploratory study, EPB41L3 and
JAM3 methylation had similar diagnostic accuracy in detecting
CIN2+ lesions as hrHPV testing (47).
Aberrant methylation levels are also found in the
promoter regions of p16INK4A, death-associated protein kinase,
O-6-methylguanine-DNA methyltransferase (MGMT), cadherin 1 and
retinoic acid receptor β of cervical cancer tissues, as well as
calcitonin related polypeptide α, human telomerase reverse
transcriptase, MYOD1, progesterone receptor and TIMP
metallopeptidase inhibitor 3 in serum samples (48).
Another mechanism involved in cervical
carcinogenesis is the methylation of the promoter of genes encoding
miRNAs. Varghese et al (49) demonstrate that hypermethylation of
the miR-434 gene's promoter is associated with the development of
cervical carcinoma. Botezatu et al (50) found a significant methylation
percentage in tumors vs. the healthy control group for miR-124a
(90.33% vs. 13.33%), miR-34b (74.19% vs. 6.67%) and miR-203 (87.09%
vs. 10.00%).
Regarding the classification of cervical cancer into
molecular subtypes based on the degree of methylation of certain
genes, a previous study has been conducted in silico, but
has not been validated in a patient cohort. Using The Cancer Genome
Atlas database, Li et al (41) identify in silico that 1,253
CpG islands were correlated with prognosis in cervical cancer. The
authors built a computational model based on novel biomarkers to
contribute to prognostic prediction and subtype classification at
the molecular level. This model could be of further use in the
clinic to guide the medical practitioner to a more personalized
approach (targeted therapy based on epigenetic subtypes).
DNA hypomethylation
In types of cancer, hypomethylation of transcription
regulatory regions is a phenomenon less encountered than
hypermethylation of gene promoters and is usually found in the
early stages of oncogenesis (51).
Yin et al (52) show that
DNA hypomethylation is caused by increased expression levels of the
serine/threonine kinase 31 gene, which is epigenetically
upregulated by E6 and E7 viral proteins. The promoter of collagen
type XVII α 1 chain was found hypomethylated in tumoral vs. normal
tissue and was able to predict invasiveness and patient outcome in
cervical cancer (53). Varghese
et al (49) show that
miR-200b and miR-34c are hypomethylated during cervical cancer
development. A methylation study performed in a cohort of women
with precancerous lesions and cervical carcinoma highlighted a
small number of hypomethylated genes: Non-SMC Condensin I Complex
Subunit G, presenilin, Histone cluster 1 H3 family member H and
ribonucleotide reductase regulatory subunit M2 with increased
expression, particularly in CIN3 lesions and cancer (54).
Alteration in histone modification
genes
Another epigenetic mechanism, histone modifications
rely on post-translational modifications (PTMs) that include
methylation, phosphorylation, acetylation, ubiquitylation and
SUMOylating. These PTMs lead to conformational change in
nucleosomes, thus facilitating a closed or open chromatin
structure. Histone methylation involves the transfer of methyl
groups from S-adenosyl-L-methionine to lysine or arginine residues
of histone proteins by histone methyltransferases (HMTs) (55). In the cell nucleus, HMTs are often
part of regulatory complexes that control and modulate DNA
methylation Histone methylation affects gene expression via
chromatin-dependent transcriptional repression or activation
(56). Histone methyltransferases
are specific for the lysine or arginine residue that they modify.
SET Domain Containing 1A (SET1), SET7/9, ASH1 Like Histone Lysine
Methyltransferase, acute lymphoblastic leukemia, methyltransferase
mixed lineage leukemia, augmenter of liver regeneration,
thioredoxin and SET and MYND domain containing 3 are histone
methyltransferases that catalyze the methylation of histone H3 at
lysine 4 (H3-K4) in mammalian cells, which is associated with an
active transcription status (57).
The methylation of both H3-K9 and H3-K27 is associated with
heterochromatin formation and leads to silenced gene expression.
This is achieved by the activity of the following HMTs:
ERG-associated protein with SET domain, Euchromatic Histone Lysine
Methyltransferase 2 (EHMT2), suppressor of variegation 3-9 homolog
(SUV39-h) 1, SUV39-h2, SET domain bifurcated histone lysine
methyltransferase 1, Dim-5, enhancer of zeste homolog 2 (EZH2) and
EHMT2(57). Increased global
H3-K27 methylation is also found to be involved in certain
pathological processes such as cancer progression. The histone
methylation process is reversible and methyl groups are removed by
histone demethylases that belong to two families: Amino oxidase
homolog lysine demethylase (KDM)1 and JmjC domain-containing
histone demethylases (58).
Histone acetyl transferases (HATs) promote gene
transcription, while histone deacetylases (HDACs) are responsible
for removing the acetyl group from lysine, silencing gene
expression. Histone modification genes exhibit alterations in
tumors and each cancer type exhibits a specific molecular pattern,
as summarized in Table III
(31). The identification of key
factors in cancer histone modification could help to develop novel
epigenetic tests (59).
| Table IIISeveral histone modification genes
that are altered in various cancer types. |
Table III
Several histone modification genes
that are altered in various cancer types.
| Cancer type | Histone
modification genes with abnormal expression |
|---|
| Bladder cancer | HBO1 |
| Breast cancer | DAC4, HDAC6, SIRT3,
SIRT7, p300, GCN5, HBO1, SRC1, NCOA3, ATF2, ELP3, EHMT2, EZH2,
SUZ12, BMI1, NSD3, SYMD2, CARM1, Suv4-20h, LSD1, BHC110, JHDM2A,
JMJD1A, TSGA, JARID1A-D |
| Cervical
cancer | HDAC1 |
| Colon cancer | HDAC1, HDAC2,
HDAC3, HDAC4, HDAC5, HDAC7, HDAC8, SIRT1, EHMT2, SYMD2 |
| Glioblastoma | JHDM1a, FBXL10,
FBXL11 |
| Lung cancer | SETDB1, LSD1,
BHC110, JMJD2A, JHDM3A, UTX, JMJD3 |
| Ovarian cancer | GCN5, EZH2, SUZ12,
BMI1 |
| Prostate
cancer | Tip60, GCN5, SRC1
EZH2, SUZ12, BMI1, CARM1, Suv4-20h, JHDM1a, FBXL10, FBXL11,
JARID1A-D |
Histone modifications in cervical
cancer
In cervical cancer, chromatin modifications induced
by hrHPV infection have not been used thus far in screening
strategies, but they have been documented thoroughly in order to
further understand the evolution and behavior of HPV infection.
A study by Groves et al (59) shows that the distribution of
interactions between the viral genome and host chromosomes are
uniform between all viral genes, with a greater percentage of
interactions deriving from the hrHPV E7 gene. In cervical cancer,
silencing of tumor suppressor genes occurs through histone
modifications as well as other epigenetic mechanisms. For example,
the HPV E7 protein can block the interactions of HDACs with
hypoxia-inducible factor-1α, which leads to an increase in
pro-angiogenic factors (60).
HPV infection mediates modifications of cellular
chromatin, leading to a differential expression profile of host
genes. These changes often occur in CpG islands of gene promoters
under the influence of DNMTs (40). These enzymes have been found to be
considerably upregulated in hrHPV infections and these
modifications can be used as biomarkers with prognostic value
(40).
Among the HPV-induced epigenetic changes, cellular
histone methylation status has an important role (61). E2 protein associates with a H3-K4
demethylase, lysine-specific demethylase 5C, to demethylate the
HPV18 upstream regulatory region in order to downregulate early
promoter activity, leading to a decreased E6/E7 expression. Another
mark of HPV-mediated control is shown by a decrease in the global
levels of H3-K27me3 under the regulation of KDM6A and KDM6B, which
target H3-K27 specifically and their expression is increased by E7
activity, leading to a transcriptionally repressed chromatin
(61). The increase in histone
acetylation induced by E7 may be required to activate E2 factor
(E2F)-responsive genes that favor the cell cycle re-entry (61).
Shadeo et al (62) highlight a modified expression level
pattern of dihydrofolate reductase, mortality factor 4-like protein
2, Morf4 family associated protein 1, nuclear receptor corepressor
1 and SWI/SNF related, matrix associated and actin dependent
regulator of chromatin subfamily c member 1; genes associated with
chromatin remodeling in early cervical lesions (including CIN2),
that is maintained throughout their progression, albeit at a
reduced frequency in CIN3. These findings have revealed novel genes
and events in the early stages of cervical dysplasia that can serve
as markers in cervical screening and provide information regarding
lesion progression.
Non-coding RNAs (ncRNAs)
ncRNAs include a number of categories and the best
known type of ncRNAs are miRNAs, which are single-stranded RNAs
with ~20 nucleotides that serve a major role in regulating gene
expression. miRNAs downregulate gene expression by binding
complementarily to the 3'untranslated region of mRNA. The majority
of miRNAs are located within CpG islands, thus allowing miRNAs to
undergo DNA methylation and/or other epigenetic modifications
(63). Another category of
interest in ncRNAs include long ncRNAs (lncRNAs), a family of
transcripts with >200 nucleotides in length, although previous
studies have shown them capable of encoding functional peptides
with short open reading frames, making their mechanisms of action
more elaborated than previously considered (64). Additionally, miRNAs and lncRNAs
exhibit epigenetic modifications themselves and their biological
functions are also affected by the methylation status (Table IV) (30,65,66).
| Table IVSeveral non-coding RNAs with altered
expression in various cancers. |
Table IV
Several non-coding RNAs with altered
expression in various cancers.
| Cancer type | Non-coding RNAs
with modified expression |
|---|
| Bladder cancer | miR-127, miR-205¸
MALAT1, Linc00346, NEAT |
| Breast cancer | miR-21, miR-372,
miR-373, miR-155, miR-146, miR-520, miR-10b, miR-9, miR-125,
miR-34a, miR-200c, miR-141, miR-429, miR-126, miR-218, miR-145,
miR-101, miR-9-1, miR-205, miR-335, miR-342, LINC00922, ATB, BCAR4,
PNUTS, ANRIL, miR-199a |
| Cervical
cancer | miR-21, miR-127,
miR-199a, miR-143, CCHE1, HOTAIR |
| Colorectal
cancer | miR-124, miR-34b,
miR-34c, miR-21, miR-155, miR-342, miR-145, CCAT2, PCAT-1 |
| Glioblastoma | miR-21,
miR-137 |
| Lung cancer | miR-17, miR-92,
miR-21, miR-155, miR-29, miR-200c, miR-141, miR-429, miR-126,
miR-9, miR-218, miR-145, miR-25, miR32, miR-142, miR-124, miR183,
miR-181, miR-101, LINC00922, MALAT1, HOTAIR, CCAT2, AK126698,
PANDA, lncRNA-RoR, loc28519, TUG-1 |
| Ovarian cancer | miR-9, miR-9-1,
miR199, miR-342, DUXAP10 |
| Pancreatic
cancer | miR-146, miR-190,
miR-196, lncRNA miR31HG |
| Prostate
cancer | miR-21, miR-146,
miR-34a, miR-146a, miR-146b, miR-145, miR-218, miR-101, miR-205,
CCAT2, PCAT-1 |
ncRNAs in cervical cancer
miRNAs expression profile allows discrimination
between neoplastic and normal cells. Previous studies found that
miR-16, miR-25, miR-92a and miR-378 are upregulated, while miR-22,
miR-27a, miR-29a and miR-100 are downregulated in cervical tissue
specimens (67-69).
These results correlate with cervical cancer progression. miR-21
targets programmed cell death 4 and overexpression of miR-21 can
promote proliferation in HeLa cells. miR-29 has been shown to
inhibit cell cycle progression (70).
A study by Gibb et al (71) found 123, 105 and 76 differentially
expressed lncRNAs in mild (CIN1), moderate (CIN2) and severe (CIN3)
dysplasia. Of these, 13 lncRNAs were common for all dysplasia
stages. However, there is limited research on the role of aberrant
lncRNA expression in cancer, although studies are currently
underway to elucidate the complex mechanisms involved. In this
regard, HOX Transcript Antisense RNA (HOTAIR), H19, X-inactive
specific transcript, cervical carcinoma high-expressed 1,
EZH2-binding lncRNA in cervical cancer, metastasis associated lung
adenocarcinoma transcript 1, antisense ncRNA in the INK4 locus,
lethal, nuclear paraspeckle assembly transcript 1, bladder cancer
associated transcript 1, ubiquitin-fold modifier-conjugating enzyme
1, small nucleolar RNA host gene (SNHG16) and SNHG20 have been
shown to serve major roles in various processes and greatly
influence the mechanisms of oncogenesis (72).
Interplay between E6 and E7 viral
proteins and the epigenetic machinery
The HPV E6 and E7 oncoproteins interact with host
cell factors to alter numerous cellular pathways and maintain the
infected cell in a proliferative state. Viral oncoproteins act at
all epigenetic levels to hijack the normal cell function to
facilitate the replication of the viral genome. It has been shown
that HPV16 E6 and E7 proteins increase DNMT1 activity and HPV16 E7
protein binds directly to DNMT1 through the zinc-finger conserved
region 3 (CR3; Fig. 1) (73). Previous studies showed that E7
oncoproteins upregulate DNMT3A and DNMT3B at the protein level
(74,75). Using this strategy, the virus
ensures its viral replication by regulation of the methylation
level of both the viral and host genomes (hypermethylation of tumor
suppressor genes), thus promoting the oncogenesis process (75). Epigenetic modulation is
accomplished not only by interaction of viral oncoproteins with
different enzymes and epigenetic factors, but also by their ability
to affect the transcription of these modulators (76).
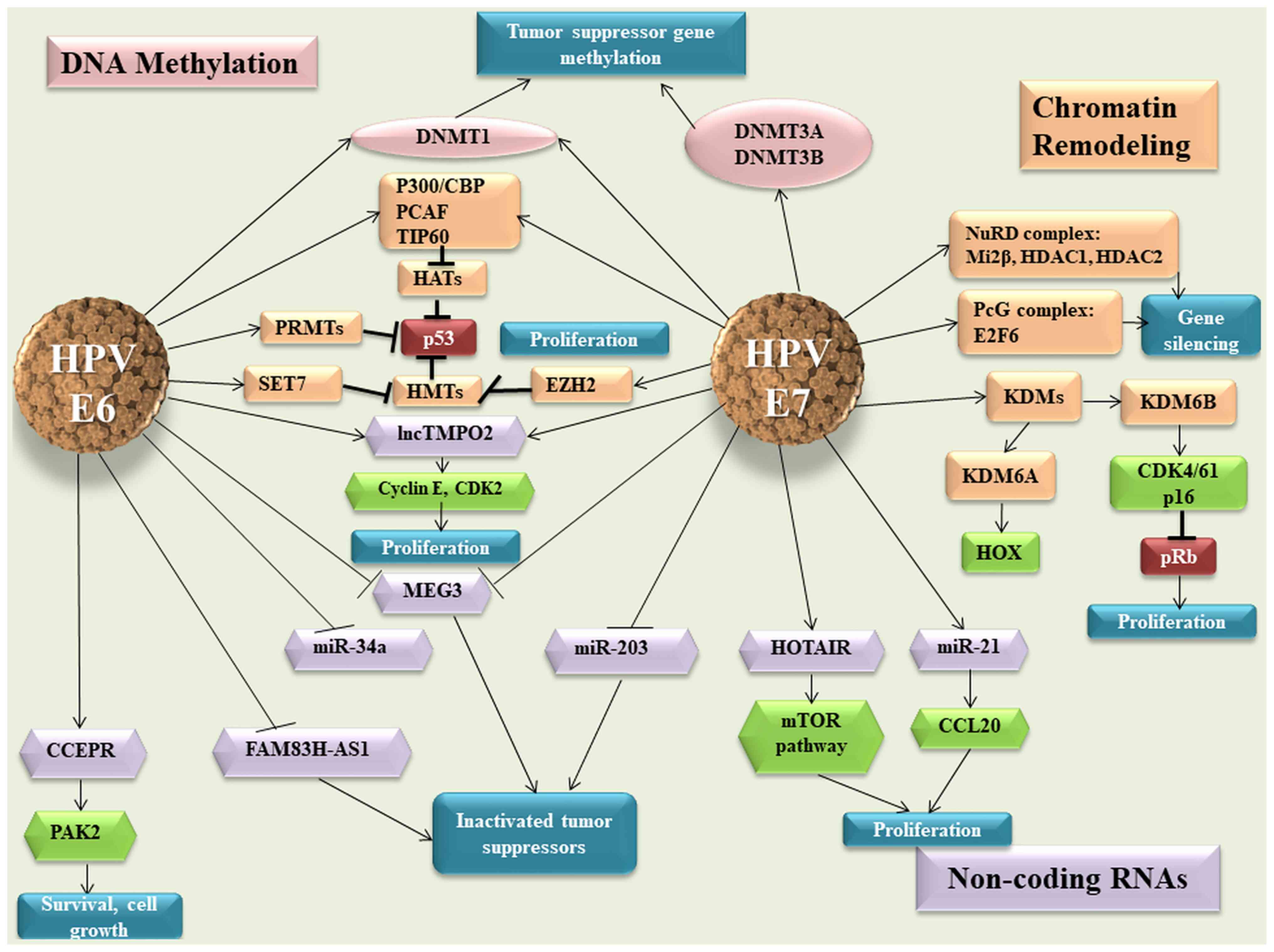 | Figure 1E6 and E7 viral oncoproteins and
their effect on the host epigenetic machinery. HPV, human
papillomavirus: DNMT, DNA methyltransferase enzymes; HATs, histone
acetyl transferases; TIP60, Tat interactive protein 60; PRMTs,
protein arginine methyltransferases; SET, SET Domain Containing 1A;
EZH2, enhancer of zeste homolog 2; lnc, long non-coding RNA; MEG3,
maternally expressed 3; miR, microRNA; CCEPR, cervical carcinoma
expressed PCNA regulatory lncRNA; PAK2, P21 (RAC1) activated kinase
2; FAM83H-AS1, family with sequence similarity 83 member H
anti-sense 1; HOTAIR, HOX Transcript Antisense RNA; PcG, polycomb
group; NuRD, nucleosome remodelling and deacetylase; HDACs, histone
deacetylases; KDM, lysine-specific demethylase; Transcription
factor E2F6; CCL20, C-C motif chemokine ligand 20; CDK, Cyclin
Dependent Kinase Inhibitor; pRB, retinoblastoma protein. |
The E7 viral protein interacts with HDACs and E2F6
transcription factor, leading to the inactivation of retinoblastoma
protein (pRB) in order to maintain the proliferative status of the
cell. The two viral oncogenes inhibit p53 p300-CREB-binding
protein-mediated acetylation, thus blocking p53 activity (77,78).
Other HATs affected by viral oncoproteins are Tat interactive
protein 60 (TIP60) and p300/CBP-associated factor, which promote an
inhibitory effect on the function of p53 and NF-κB, thus allowing
the infected cells to escape the immune response (79-81).
HPVs not only modify the host genes, but also the expression of
viral oncogenes (82). TIP60
targets E6 for degradation, which derepresses the HPV early
promoter, thus representing an E2-independent repression mechanism
(83). The E7 oncoprotein
interacts not only with HATs, but also with HDAC1 and HDAC2 through
Chromodomain Helicase DNA Binding Protein 4, which forms part of
the nucleosome remodelling and deacetylase (NuRD) chromatin
remodelling complex (84).
This interaction modulates the transcription of
several genes that are necessary for the differentiation-dependent
phase of the virus life cycle, but also represses IFN-β gene
transcription, which leads to a suppressed cellular immune response
to HPV viral infection (Fig. 1)
(85,86).
Coactivator associated arginine methyltransferase 1,
protein arginine methyltransferase 1 and SET7 methyltransferases
are modulated by E6 protein, thus affecting p53 function (76). It has been shown that HPV E7
oncoprotein induces the expression of KDM6A and KDM6B, thereby
inducing pRB degradation and the reactivation of transcription of
genes that are normally repressed, such as HOX genes and p16INK4A
(87).
Regarding ncRNAs, cervical carcinoma expressed PCNA
regulatory lncRNA is upregulated in response to HPV16 E6
expression, enhancing P21 (RAC1) activated kinase 2 expression
through miR-922 sponging (88).
In silico analysis predicts that HPV16 E7 may bind HOTAIR
and this was validated by E7 immunoprecipitation followed by
reverse transcription-quantitative (RT-q) PCR analysis (89). This interaction may impede the
ability of HOTAIR to interact with polycomb repressive complex 2
and/or KDM1A, thus causing the de-repression of polycomb-regulated
genes (87). In cervical cancer,
maternally expressed 3 (MEG3) presents a decreased expression as a
direct consequence of HPV E6/E7 expression (90). In vitro ectopic MEG3
expression inhibits cell proliferation, increases apoptosis and
reduces tumorigenicity in xenograft models (90).
E6 and E7 hrHPV are found to regulate the expression
of lncRNA thymopoietin pseudogene 2 (TMPOP2) in a feedback-positive
manner, leading to an increased expression of this lncRNA (91). The molecular mechanism involves the
tumor suppressor p53, which represses the transcription of lncRNA
TMPOP2 through direct interaction (91). The same study shows that TMPOP2 has
a role in cell proliferation, which suggests that this lncRNA could
be a potential diagnostic marker and therapeutic target of cervical
cancer (Fig. 1) (91).
It has been demonstrated that certain miRNAs with a
tumor suppressor role present a downregulated expression in
cervical cancer, which is associated with the expression of viral
oncogenes. Decreased miR-34a expression is associated with the
early productive phase and is correlated with E6 expression
(92). Melar-New and Laimins
(93) show that miR-203 exhibits
an inhibitory effect upon HPV amplification and HPV proteins act to
suppress the expression of this miRNA to allow viral replication in
differentiating cells (Fig. 1).
miR-21 is another miRNA that is associated with HPV infection and
is upregulated in cervical cancer (94). The same study suggests that
chemokine ligand 20 expression is negatively correlated with miR-21
expression and promotes cell proliferation (94).
Viral genes modifications
The HPV genome is subjected to epigenetic
regulation, besides DNA methylation, at the level of PTMs of
histones, including acetylation, phosphorylation and methylation.
These processes control the viral gene expression, serving a main
part in the life cycle pf the virus (61).
In addition to host genes methylation, the
methylation of viral genes exhibits high levels, particularly in
capsid genes and late open reading frames. This has been associated
with the progression of lesions from CIN2/3 to invasive cancer
(95,96). In a study on HPV-infected Asian
women with different grade of lesions, the methylation status of
CpG islands in HPV16, 18, 52 and 58 was investigated. A correlation
was observed between the methylation status of the Papillomavirus
Major Capsid Protein 1 (L1) gene and the disease severity and
progression (96). The methylation
status of the L1 gene of HPV16 and HPV18 can discriminate between
normal tissue and CIN1 lesions. In HPV52 infection, it can
differentiate between <CIN2 and ≥CIN2 lesions and between
<CIN3 and ≥CIN3 lesions. Furthermore, improved cervical lesion
diagnosis was achieved by combining host and HPV genome methylation
analysis. Thus, a positive correlation between HPV16 and HPV18
methylation status and paired box 1 (PAX1) and SOX1 was observed.
On the other hand, a negative correlation was observed between
HPV52 methylation and the same host genes (96). This led to the development of a
biomarker panel called S5, using the DNA methylation levels of
HPV16 L1, HPV16 Minor Capsid Protein (L2), HPV18 L2, HPV31 L1,
HPV33 L2 and EPB41L3 tumor suppressor gene. The S5 methylation
assay provided a 90% sensitivity and a 49% specificity for
detecting high-grade cervical lesions in a cohort of women from
United Kingdom (97). In a Mexican
study, the S5 classifier was able to detect all cancer cases,
reducing the number of colposcopies required by 30-50% as opposed
to triage by cytology and HPV16/18 genotyping (42) Considering that only a small
percentage of lesions progress to cervical cancer and the process
is rather slow, it is important to identify accordingly the women
who are at risk of developing a malignancy and avoid unnecessary
expenses and treatment for the women who are most likely to
overcome naturally the infection (42,98).
4. DNA methylation for cervical
screening
Methylation markers can be used in cervical cancer
screening programs, with studies demonstrating their increased
specificity compared with that of testing for HPV and
immunohistochemistry (p16/Ki-67) (99). Previous studies that have shown a
correlation between the expression of HPV E6 and E7 oncogenes and
the activity of DNMTs support the validation of DNA
hypermethylation as a common event during cervical carcinogenesis
(100-103).
Certain studies have demonstrated that methylation assays are a
useful tool for triaging women with advanced HPV infections,
although their performance is rather poor in on-going hrHPV
infections that have not progressed yet to CIN3 or cervical cancer
(97,104-106).
In addition, with advancements in technology, HPV DNA testing could
be replaced by DNA methylation screening (95). At present, methylation testing is
based on RT-qPCR to estimate the percentage of methylated DNA
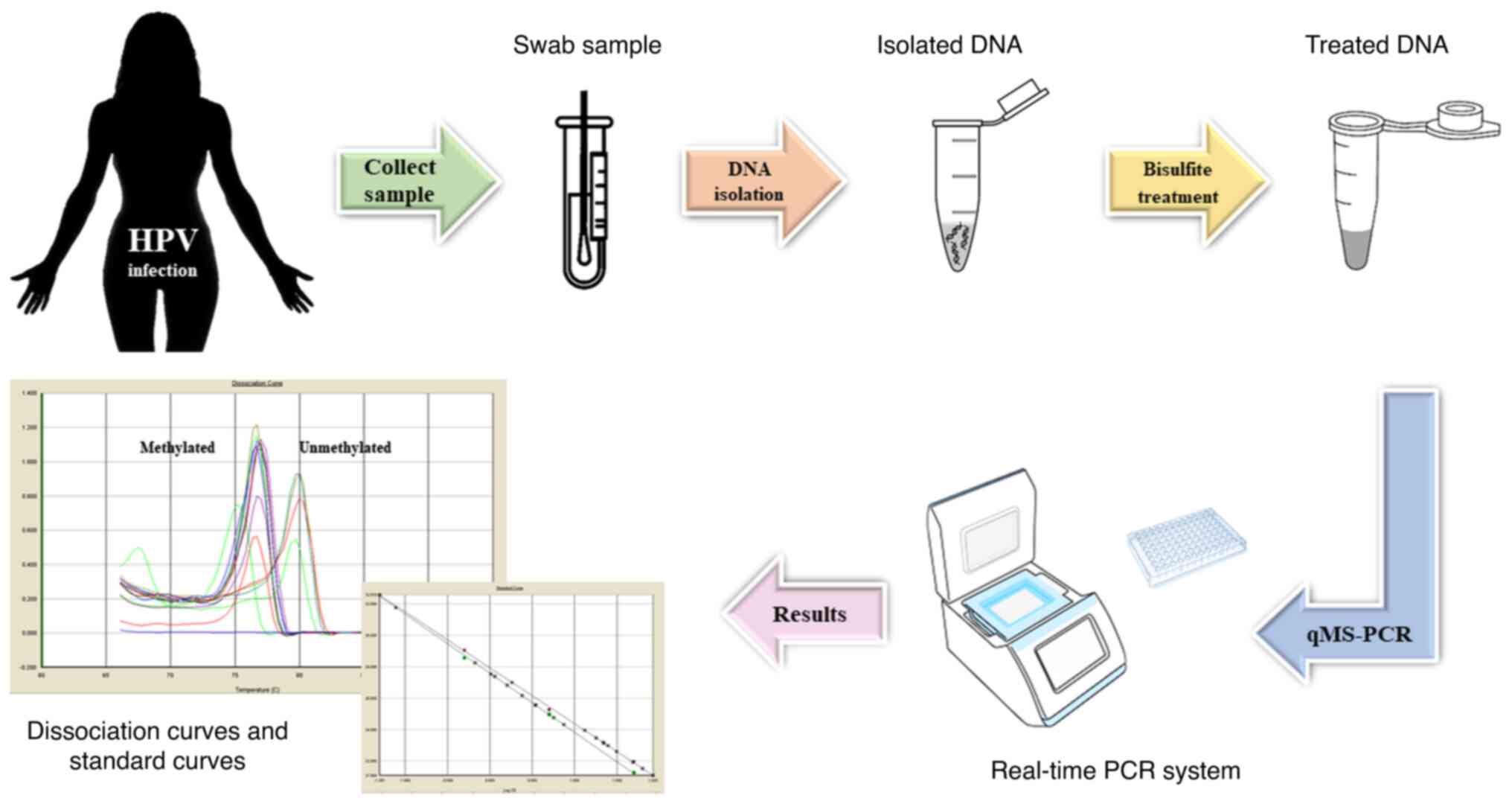
molecules in the sample for the targeted region (Fig. 2). Currently, the most important
application of novel biomarkers in cervical carcinoma is the triage
of women with positive cytology and/or HPV tests.
Commercial screening kits based on DNA
methylation
The QIAsure Methylation Test (Qiagen GmbH) is a
multiplex quantitative methylation-specific PCR (qMSP)-based assay
that amplifies the methylated promoter regions of the tumor
suppressor genes family with sequence similarity 19 [chemokine (C-C
motif)-like) member A4] and Homo sapiens-miR-124-2, as well
as a methylation-unspecific fragment of a reference gene, actin β,
that acts as a quality control marker. The QIAsure Methylation Test
has a sensitivity of 20.5% for identifying ≤CIN1 infections, 38.1%
for CIN2 infections, 67% for CIN3 infections, 90% for advanced
transforming CIN and 100% for cervical cancer in hrHPV-positive
samples (107).
GynTect® (Oncgnostics GmbH) is another
MSP-based assay that distinguishes between cervical lesion types by
investigating the methylation status of the promoter regions of six
genes (astrotactin1, distal-less homeobox 1, integrin subunit α4,
relaxin family peptide receptor 3, SOX17 and zinc finger protein
671) and uses two quality control markers (iduronate 2-sulfatase-M
and acetylcholinesterase). Data from clinical trials show that
GynTect® markers are positive prior to any histological
finding of CIN1, CIN2 or CIN3 in a number of patients (108).
The QIAsure Methylation Test records slightly higher
detection rates than GynTect® among CIN samples, but no
significant differences have been found. Instead, a significant
difference is observed in the detection rates of negative for
intraepithelial lesion or malignancy samples, irrespective of the
hrHPV status. Also, a notable difference was noticed in the
specificity of CIN2+ and CIN3+ cases. The
tests scored positive predictive values of 64.3% for
GynTect® and 51.4% for the QIAsure Methylation Test and
their negative predictive values were 85.5 and 88.1%, respectively.
The two kits showed a markedly high sensitivity for the detection
of cancer cases, recognizing all cancer and carcinoma in
situ samples. Another study found similar results for the
detection rates of different stages of CIN, ranging from 13.3-20%
for CIN1, 33.3-44.4% for CIN2 and 60-61.2% for CIN3 for the
GynTect® assay (109).
For the QIAsure Methylation Test, the detection rates were 26.7%
for CIN1, 27.8% for CIN2 and 74.3% for CIN3. With the exception of
CIN2 cases, these positivity rates are in agreement with those from
previous studies (CIN1, 27.7%; CIN2, 44.3%; and CIN3, 75.8%)
(24,108,109). The differences between the
capabilities of these two tests are illustrated in Table V.
| Table VThe differences between two
methylation assays. |
Table V
The differences between two
methylation assays.
| QIAsure Methylation
Test | Sensitivity, % | Specificity, % |
|---|
|
CIN 2+ | 67.9 | 66.2 |
|
CIN 3+ | 73.3 | 65.9 |
|
Cancer | 100.0 | - |
| GynTect | - | - |
|
CIN 2+ | 60.4 | 88.2 |
|
CIN 3+ | 66.7 | 86.9 |
|
Cancer | 100.0 | - |
The CONFIDENCE™ assay developed by Neumann
Diagnostics Kft. detects HPV DNA (CONFIDENCE HPV™) via a
TaqManVR-based L1 region-specific multiplex qPCR assay and it also
contains a human epigenetic biomarker test (CONFIDENCE Marker™)
that measures the methylation level of CpG sites in the promoter
region of POU class 4 homeobox 3 by qMSP. As an internal reference
for normalizing the methylation level, a quantitative measurement
of type II collagen is used. The CONFIDENCE Marker™ test was
compared with cytology-based methods for triaging hrHPV-positive
cases and it was found to be more sensitive and to have a
comparable specificity, scoring for CIN2+ and
CIN3+ a relative sensitivity of 1.67 and 1.74,
respectively, with a relative specificity of 1.01 for
CIN2+ and 0.98 for CIN3+ (110).
PAX1 DNA Detection kit and ZNF582 DNA Detection kit
(manufactured by iStat Biomedical Co. Ltd.) are based on qPCR
technology and target the PAX1 and zinc finger protein 582 (ZNF582)
genes. PAX1 exhibits biomarker potential in a study on 443 patient
samples. The study revealed that CIN3+ detection
sensitivity was 92%, while the specificity was 83% (111). Another study that included 449
patients reveals that the presence of HPV16/18 and the methylation
status of PAX1 and ZNF582 genes is correlated with CIN3+
dysplasia (110). HPV 16/18
detection combined with PAX1 displayed a sensitivity of 89.2% and a
specificity of 76.0%, while HPV 16/18 detection combined with
ZNF582 displayed a sensitivity and specificity of 85.4 and 80.1%,
respectively (111,112).
Methylation-based assays have been developed and
used successfully for other types of cancer, demonstrating that
this type of triaging/diagnostic approach is feasible and less
invasive than traditional methods. Each test includes
cancer-specific genes and they aim at recurrence, survival and
early detection.
For bladder cancer, the necessity of early detection
and the monitoring of recurrence led to the development and
approval of different types of DNA methylation based-assays. The
following assays are all based on DNA extracted from patients'
urine samples. Bladder EpiCheck® (Nucleix), a
non-muscle-invasive bladder cancer (NMIBC) diagnosis assay, is
based on 15 methylation biomarkers and the results are compiled as
an EpiScore ranging from 0 to 100, where scores >60 indicate a
positive result for recurrence (113-115).
A similar assay is Bladder CARE™ (Pangaea Laboratories, Ltd.),
which is used in NMIBC recurrence and is based on a three-gene
panel: SOX1, interleukin 1 receptor associated kinase 3 and long
interspersed nuclear element 1(116). By contrast, AssureMDx™ for
Bladder Cancer (MDxHealth) is an assay based on a negative result
that excludes bladder cancer by targeting the orthodenticle
homeobox 1, one cut homeobox 2 and Twist family BHLH transcription
factor 1 genes in via MSP (117).
Similarly, UroMark (University College London) has the capacity to
detect the methylation status of 150 loci across the genome. The
two assays are promising options in patients presenting hematuria
(118).
For breast cancer detection, the current DNA
methylation-based assays are limited and the only available
predictive test is therascreen® PITX2 RQG test developed
by Qiagen GmbH/Therawis Pharma GmbH. The assay is based on a qPCR
assay that establishes the ratio between methylated and
unmethylated DNA in tumoral samples. The ratio is also an indicator
of survival when standard therapy is associated with anthracyclines
(119).
For the diagnosis of primary colorectal cancer
(CRC), two DNA methylation assays with Food and Drug Administration
approval are available: Epi proColon® (Epigenomics AG)
and Cologuard® (Exact Sciences Corporation). The former
is a blood-based assay that targets the methylation changes of the
septin 9 (SEPT9) gene promoter in cfDNA. Similar,
Cologuard® is a multi-target stool DNA-based test used
in screening that targets the methylated promoter of the bone
morphogenetic protein 3 and N-myc downstream-regulated gene 4 genes
and seven mutations in the KRAS gene. The two assays are available
for CRC screening of patients who are ≥50 years of age, with
re-testing recommended every 3 years (120).
The current detection method based on DNA
methylation in patients with cirrhosis is the HCCBloodTest
(Epigenomics AG). This assay is designed to quantify the
methylation levels of the SEPT9 gene in blood samples using RT-qPCR
technology (121). Another test,
OncoguardTM Liver (Exact Sciences Corp.), for detection
of early hepatocellular carcinoma (HCC) was developed in
association with Mayo Clinic Healthcare. The panel test is blood
based and targets four genes (disabled homolog 2-interacting
protein (DAB2IP), empty spiracles homeobox 1 (EMX1), homeobox A1
(HOXA1) and testis-specific Y-encoded-like protein 5 (TSPYL5)) and
two protein markers [α fetoprotein (AFP) and lectin-bound AFP]
using immunochemical methods and QuARTS™ technology (122). Another promising assay is IvyGene
Dx Liver Cancer Test (Laboratory for Advanced Medicine Inc.), which
detects the methylation levels of genes in cfDNA isolated from
plasma. The test validates the presence of HCC by using
next-generation sequencing technology with 95% sensitivity and
97.5% specificity in detection (123).
For glioblastoma diagnosis, MGMT methylation
detection status represents the target for different assays. Of
these assays, four include CE-marked (Therascreen® MGMT
Pyro® kit from Qiagen GmbH, Human MGMT Gene Methylation
Detection kit from Xiamen Spacegen Co. Ltd., MGMT Promoter
Methylation Detection kit from EntroGen Inc. and EntroGen MGMT
Promoter Methylation Detection kit from EntroGen Inc.). These tests
use different molecular methods (pyrosequencing technology,
amplification refractive mutation system and
pyrophosphorolysis-activated polymerization reaction and
semi-quantitative RT-PCR with fluorescent hydrolysis probes
(124). Another assay is
PredictMDx (MdxHealth), which is a laboratory developed test (LDT)
based on qMSP technology for detecting MGMT methylation in DNA
extracted from FFPE biopsies (125).
At present, the only assay available for early
detection in prostate cancer is an LDT. ConfirmMDx (MDxHealth) is a
biopsy tissue-based assay design to target glutathione
S-transferase π 1, Ras association domain family member 1 and
adenomatous polyposis coli genes that uses DNA qMSP technology
(126).
Currently, Epi proLung® (Epigenomics AG)
is the only CE-marked test available for lung cancer diagnosis in
patients at increased risk of the disease. The assay is based on a
triplex RT-qPCR with fluorescent hydrolysis probe that targets two
genes, short stature homeobox 2 and prostaglandin E receptor 4,
which are detected in cfDNA from plasma. The test also uses a third
gene, β-actin, as a housekeeping gene (127).
For the detection of multiple tumors, two types of
assay have been developed. The first, IvyGene® Cancer
Blood Test (Laboratory for Advanced Medicine Inc.) is designed to
detect the presence of breast, colon, liver and lung tumors from
blood samples by evaluating the methylation levels of cfDNA by
using a panel of 46 markers (128). The second, EPICUP™ (Merete GmbH)
is a tissue-based assay that can use either freshly frozen or FFPE
biopsies. To classify the type of Cancer of Unknown Primary, the
test uses bead array technology from Illumina, Inc. (129).
5. Conclusions and perspectives
HPV diagnostic and triaging assays need to advance
in terms of capacities and affordability, particularly in
low-to-middle income countries where the prevalence of this type of
infection remains markedly high. One area that shows promise is
associated with DNA methylation classifiers, which can be improved
by identifying improved gene combinations and by automating the
process.
A study by Snoek et al (130) found a correlation coefficient
varying from 0.691 [PR/SET domain 14 (PRDM14)] to 0.9 (SST) for six
DNA methylation markers (FAM19A4, GHSR, PHACTR3, PRDM14, SST and
ZIC1) in samples obtained from urine sediments and native urine. In
the same study, a correlation coefficient varying from 0.508
(PRDM14) to 0.717 (PHACTR3) was observed between urine sediments
and cervical scrapes, although the DNA methylation levels in
cervical scrapes were considerably higher than those in urine. A
significant correlation was found in DNA methylation markers in
patients with cervical cancer compared with those in controls. The
study showed that DNA methylation marker testing is feasible for
detecting cervical cancer and all markers have a high
discriminatory power in urine sediments.
Another study by Clarke et al (18) shows that HPV type-specific DNA
methylation performs well for risk stratification and has an
improved performance compared with that of HPV 16/18 genotyping and
Pap cytology. The association between increased methylation with
CIN3 or adenocarcinoma in situ has been evaluated across 12
carcinogenic HPV types and is shown to have a strong correlation,
allowing for an improved observation of the transition from HPV
lesions to cancer (42).
DNA methylation modifications and chromatin changes
have been demonstrated to be hallmarks of epigenetic reprogramming
in malignant disease and should be investigated in order to further
understand the precise role of chromatin remodeling and protein-DNA
interactions in HPV-induced carcinogenesis (131). The findings could provide novel,
more suitable markers for triaging HPV-infected women.
DNA methylation tests present numerous advantages.
They can be used not only in tissue samples, but in any body fluid
(liquid biopsy) (18). In cervical
cancer, this type of assay may be combined with traditional
screening for an improved triage and therapy management, adding
valuable information regarding epigenetic profiles (24).
Such novel approaches would provide a clearer
primary diagnosis, whereas a number of classic tests would be
inconclusive and would help triaging via a minimally invasive
screening method. With further research, the epigenetic profiles
and subtypes of the tumors could be elaborated, which would aid in
therapy selection by opening avenues in personalized precision
medicine. Response to therapy could also be evaluated through such
methods and the accessibility of liquid biopsies would allow a
constant monitoring of the patient's status without invasive
sampling techniques.
In addition, tissue biopsies sample only a
subpopulation of all the cell types found within a tumor and could
potentially miss the aspect of heterogeneity and clonality of the
entire tumor, thus providing a skewed image of the cellular
phenotype (18).
According to previous studies, ctDNA may offer a
more accurate picture regarding the molecular composition of the
tumor and precursor lesions in HPV infection since all cell types
are likely to contribute (18,132,133). Other sources of information
regarding the composition of the tumor include cfDNA and nucleic
acids from exosomes. Exosomes are found in both physiological and
pathological conditions in the majority of body fluids, including
blood, urine, cerebrospinal fluid, saliva, serum, amniotic fluid,
breast milk and cervical lavage (132). An important feature of exosomes
is that they represent with high fidelity a ‘snapshot’ of the cell
they originate from and their molecular cargo can be transferred to
other cells. It has been noticed that, in the oncogenic process,
the number of exosomes increases significantly and tumors cells
release a high number of exosomes that contain different markers
that could promote cancer progression and metastasis (133).
Furthermore, there is increasing evidence that
supports the involvement of ncRNA species, such as lncRNAs, in
cancer pathogenesis. The data show a different lncRNA profile of
expression in cervical lesions and cancer and a large number of the
identified molecules have the potential to be used as markers for
diagnostic and prognosis (134).
Therefore, exosomal lncRNAs from cervical lavage and urine samples
could be used as cancer biomarkers and could serve to develop a
novel, non-invasive array for early detection/triage of precursor
lesions and cervical cancer (135).
Acknowledgements
Not applicable.
Funding
The present study was funded by UEFISCDI (grant no.
PN-III-P1-1.1-TE 39/2020).
Availability of data and materials
Not applicable.
Authors' contributions
AA, AF, AB and GA conceived and designed the
review. AA, AF, AB, IVI, AP and GA performed the literature
research. AA, AF, AB, IVI, AP and GA were involved in the
interpretation of the findings in the literature. AA, AF, AB and GA
were involved in the writing of the manuscript. Data authentication
if not applicable. All authors have read and approved the final
manuscript for publication.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global Cancer Statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249.
2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Durzynska J, Lesniewicz K and Poreba E:
Human papillomaviruses in epigenetic regulations. Mutat Res Rev
Mutat Res. 772:36–50. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Sawaya GF, Brown AD, Washington AE and
Garber AM: Current approaches to cervical-cancer screening. N Engl
J Med. 344:1603–1607. 2001.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Sawaya GF, Smith-McCune K and Kuppermann
M: Cervical cancer screening: More choices in 2019. JAMA.
321:2018–2019. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Botezatu A, Iancu IV, Plesa A, Manda D,
Popa O, Bostan M, Mihaila M, Albulescu A, Fudulu A, Vladoiu SV, et
al: Methylation of tumour suppressor genes associated with thyroid
cancer. Cancer Biomark. 25:53–65. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Tasca L, Ostör AG and Babeş V: XII. Aurel
Babeş. Int J Gynecol Pathol. 21:198–202. 2002.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Tan SY and Tatsumura Y: George
Papanicolaou (1883-1962): Discoverer of the Pap Smear. Singapore
Med J. 56:586–587. 2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Petry KU, Woermann B and Schneider A:
Benefits and risks of cervical cancer screening. Oncol Res Treat.
37 (Suppl 3):S48–S57. 2014.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lazcano-Ponce E, Lorincz AT, Cruz-Valdez
A, Salmeron J, Uribe P, Velasco-Mondragón E, Nevarez PH, Acosta RD
and Hernandez-Avila M: Self-collection of vaginal specimens for
human papillomavirus testing in cervical cancer prevention (MARCH):
A community-based randomised controlled trial. Lancet.
378:1868–1873. 2011.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Koliopoulos G, Nyaga VN, Santesso N,
Bryant P, Martin-Hirsch P, Mustafa RA, Schünemann H, Paraskevaidis
E and Arbyn M: Cytology vs. HPV testing for cervical cancer
screening in the general population. Cochrane Database Syst Rev.
8(CD008587)2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Schmitz M, Eichelkraut K, Schmidt D,
Zeiser I, Hilal Z, Tettenborn Z, Hansel A and Ikenberg H:
Performance of a DNA methylation marker panel using liquid-based
cervical scrapes to detect cervical cancer and its precancerous
stages. BMC Cancer. 18(1197)2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wentzensen N, Schiffman M, Palmer T and
Arbyn M: Triage of HPV positive women in cervical cancer screening.
J Clin Virol. 76 (Suppl 1):S49–S55. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Dillner J, Rebolj M, Birembaut P, Petry
KU, Szarewski A, Munk C, de Sanjose S, Naucler P, Lloveras B, Kjaer
S, et al: Long term predictive values of cytology and human
papillomavirus testing in cervical cancer screening: Joint European
cohort study. BMJ. 337(a1754)2008.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Saslow D, Solomon D, Lawson HW, Killackey
M, Kulasingum SL, Cain J, Garcia FAR, Moriarty AT, Waxman AG,
Wilbur DC, et al: American Cancer Society, American Society for
Colposcopy and Cervical Pathology and American Society for Clinical
Pathology screening guidelines for the prevention and early
detection of cervical cancer. CA Cancer J Clin. 62:147–172.
2012.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Bian M, Cheng J, Ma L, Cong X, Liu J, Chen
Y and Chen X: Evaluation of the detection of 14 high-risk human
papillomaviruses with HPV 16 and HPV 18 genotyping for cervical
cancer screening. Exp Ther Med. 6:1332–1336. 2013.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Tian Y, Wu NYY, Liou YL, Yeh CT, Cao L,
Kang YN, Wang HJ, Li Y, Chu TY, Li W, et al: Utility of gene
methylation analysis, cytological examination and HPV-16/18
genotyping in triage of high-risk human papilloma virus-positive
women. Oncotarget. 8:62274–62285. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Fudulu A, Albulescu A and Anton G: Human
papillomaviruses' proteins with clinical utility. J Immunoassay
Immunochem. 40:81–90. 2013.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Clarke MA, Gradissimo A, Schiffman M, Lam
J, Sollecito CC, Fetterman B, Lorey T, Poitras N, Raine-Bennett TR,
Castle PE, et al: Human papillomavirus DNA methylation as a
biomarker for cervical precancer: Consistency across 12 genotypes
and potential impact on management of HPV-positive women. Clin
Cancer Res. 24:2194–2202. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Gai W and Sun K: Epigenetic biomarkers in
cell-free DNA and applications in liquid biopsy. Genes (Basel).
10(32)2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Dagogo-Jack I and Shaw AT: Tumour
heterogeneity and resistance to cancer therapies. Nat Rev Clin
Oncol. 15:81–94. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Baylin SB and Jones PA: Epigenetic
determinants of cancer. Cold Spring Harb Perspect Biol.
8(a019505)2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Lleras RA, Smith RV, Adrien LR, Schlecht
NF, Burk RD, Harris TM, Childs G, Prystowsky MB and Belbin TJ:
Unique DNA methylation loci distinguish anatomic site and HPV
status in head and neck squamous cell carcinoma. Clin Cancer Res.
19:5444–5455. 2013.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wentzensen N, Sherman ME, Schiffman M and
Wang SS: Utility of methylation markers in cervical cancer early
detection: Appraisal of the state-of-the-science. Gynecol Oncol.
112:293–299. 2009.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lorincz AT: Virtues and weaknesses of DNA
methylation as a test for cervical cancer prevention. Acta Cytol.
60:501–512. 2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Plesa A, Iancu IV, Botezatu A, Huica I,
Stoian M and Anton G: The involvement of epigenetic mechanisms in
HPV-induced cervical cancer, Rajamanickam Rajkumar, human
papillomavirus-research in a global perspective, IntechOpen,
London, 191-239, 2016. https://www.intechopen.com/chapters/50425. Accessed
June, 16, 2021.
|
|
26
|
Du J, Johnson LM, Jacobsen SE and Patel
DJ: DNA methylation pathways and their crosstalk with histone
methylation. Nat Rev Mol Cell Biol. 16:519–532. 2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Jin B and Robertson KD: DNA
methyltransferases, DNA damage repair and cancer. Adv Exp Med Biol.
754:3–29. 2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Williams K, Christensen J and Helin K: DNA
methylation: TET proteins-guardians of CpG islands? EMBO Rep.
13:28–35. 2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Schübeler D: Function and information
content of DNA methylation. Nature. 517:321–326. 2015.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Sina AA, Carrascosa LG, Liang Z, Grewal
SY, Wardiana A, Shiddiky MJA, Gardiner RA, Samaratunga H, Gandhi
MK, Scott RJ, et al: Epigenetically reprogrammed methylation
landscape drives the DNA self-assembly and serves as a universal
cancer biomarker. Nat Commun. 9(4915)2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kanwal R, Gupta K and Gupta S: Cancer
epigenetics: An introduction. Methods Mol Biol. 1238:3–25.
2015.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Reinert T: Methylation markers for
urine-based detection of bladder cancer: The next generation of
urinary markers for diagnosis and surveillance of bladder cancer.
Adv Urol. 2012(503271)2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
de Groot JS, Pan X, Meeldijk J, van der
Wall E, van Diest PJ and Moelans CB: Validation of DNA promoter
hypermethylation biomarkers in breast cancer-a short report. Cell
Oncol (Dordr). 37:297–303. 2014.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Yi JM: DNA Methylation change profiling of
colorectal disease: Screening towards clinical use. Life (Basel).
11(412)2021.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Etcheverry A, Aubry M, de Tayrac M,
Vauleon E, Boniface R, Guenot F, Saikali S, Hamlat A, Riffaud L,
Menei P, et al: DNA methylation in glioblastoma: Impact on gene
expression and clinical outcome. BMC Genomics.
11(701)2010.PubMed/NCBI View Article : Google Scholar
|
|
36
|
LeBlanc VG and Marra MA: DNA methylation
in adult diffuse gliomas. Brief Funct Genomics. 15:491–500.
2016.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zhang C, Li J, Huang T, Duan S, Dai D,
Jiang D, Sui X, Li D, Chen Y, Ding F, et al: Meta-analysis of DNA
methylation biomarkers in hepatocellular carcinoma. Oncotarget.
7:81255–81267. 2016.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Shen N, Du J, Zhou H, Chen N, Pan Y,
Hoheisel JD, Jiang Z, Xiao L, Tao Y and Mo X: A diagnostic panel of
DNA methylation biomarkers for lung adenocarcinoma. Front Oncol.
9(1281)2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Yang M and Park JY: DNA methylation in
promoter region as biomarkers in prostate cancer. Methods Mol Biol.
863:67–109. 2012.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Fang J, Zhang H and Jin S: Epigenetics and
cervical cancer: From pathogenesis to therapy. Tumour Biol.
35:5083–5093. 2014.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Li C, Ke J, Liu J and Su J: DNA
methylation data-based molecular subtype classification related to
the prognosis of patients with cervical cancer. J Cell Biochem.
121:2713–2724. 2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Hernández-López R, Lorincz AT,
Torres-Ibarra L, Reuter C, Scibior-Bentkowska D, Warman R, Nedjai
B, Mendiola-Pastrana I, León-Maldonado L, Rivera-Paredez B, et al:
Methylation estimates the risk of precancer in HPV-infected women
with discrepant results between cytology and HPV16/18 genotyping.
Clin Epigenetics. 11(140)2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Lorincz AT: Cancer diagnostic classifiers
based on quantitative DNA methylation. Expert Rev Mol Diagn.
14:293–305. 2016.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Clarke MA, Luhn P, Gage JC, Bodelon C,
Dunn ST, Walker J, Zuna R, Hewitt S, Killian JK, Yan L, et al:
Discovery and validation of candidate host DNA methylation markers
for detection of cervical precancer and cancer. Int J Cancer.
141:701–710. 2017.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Kremer WW, Van Zummeren M, Novianti PW,
Richter KL, Verlaat W, Snijders PJF, Heideman DAM, Steenbergen RDM,
Dreyer G and Meijer CJ: Detection of hypermethylated genes as
markers for cervical screening in women living with HIV. J Int AIDS
Soc. 21(e25165)2018.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Verlaat W, Van Leeuwen RW, Novianti PW,
Schuuring E, Meijer CJLM, Van Der Zee AGJ, Snijders PJF, Heideman
DAM, Steenbergen RDM and Wisman GBA: Host-cell DNA methylation
patterns during high-risk HPV-induced carcinogenesis reveal a
heterogeneous nature of cervical pre-cancer. Epigenetics.
13:769–778. 2018.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Kong L, Wang L, Wang Z, Xiao X, Xu T, Wu
H, Wu M, Liu P and Li L: DNA methylation for cervical cancer
screening: A training set in China. Clin Epigenetics.
12(91)2020.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Shivapurkar N and Gazdar AF: DNA
methylation-based biomarkers in non-invasive cancer screening. Curr
Mol Med. 10:123–132. 2010.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Varghese VK, Shukla V, Kabekkodu SP,
Pandey D and Satyamoorthy K: DNA methylation regulated microRNAs in
human cervical cancer. Mol Carcinog. 57:370–382. 2018.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Botezatu A, Goia-Rusanu CD, Iancu IV,
Huica I, Plesa A, Socolov D, Ungureanu C and Anton G: Quantitative
analysis of the relationship between microRNA-124a,-34b and-203
gene methylation and cervical oncogenesis. Mol Med Rep. 4:121–128.
2011.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Ehrlich M: DNA hypomethylation in cancer
cells. Epigenomics. 1:239–259. 2009.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Yin FF, Wang N, Bi XN, Yu X, Xu XH, Wang
YL, Zhao CQ, Luo B and Wang YK: Serine/threonine kinases 31(STK31)
may be a novel cellular target gene for the HPV16 oncogene E7 with
potential as a DNA hypomethylation biomarker in cervical cancer.
Virol J. 13(60)2016.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Thangavelu PU, Krenács T, Dray E and Duijf
PHG: In epithelial cancers, aberrant COL17A1 promoter methylation
predicts its misexpression and increased invasion. Clin Epigenet.
8(120)2016.PubMed/NCBI View Article : Google Scholar
|
|
54
|
García AD, Abba MC, Briceño I, Aristizabal
FA and Arregui AC: DNA methylation pattern in high-grade cervical
intraepithelial neoplasia and cancer revealed by genomewide
methylation analysis of cervical DNA. Integr Mol Med. 4:1–13.
2017.doi: 10.15761/IMM.1000309.
|
|
55
|
Miller JL and Grant PA: The role of DNA
methylation and histone modifications in transcriptional regulation
in humans. Subcell Biochem. 61:289–317. 2013.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Greer EL and Shi Y: Histone methylation: A
dynamic mark in health, disease and inheritance. Nat Rev Genet.
13:343–357. 2012.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Bannister AJ and Kouzarides T: Regulation
of chromatin by histone modifications. Cell Res. 21:381–395.
2011.PubMed/NCBI View Article : Google Scholar
|
|
58
|
D'Oto A, Tian QW, Davidoff AM and Yang J:
Histone demethylases and their roles in cancer epigenetics. J Med
Oncol Ther. 1:34–40. 2016.PubMed/NCBI
|
|
59
|
Groves IJ, Drane ELA, Michalski M, Monahan
JM, Scarpini CG, Smith SP, Bussotti G, Várnai C, Schoenfelder S,
Fraser P, et al: Three-dimensional interactions between integrated
HPV genomes and cellular chromatin dysregulate host gene expression
in early cervical carcinogenesis. bioRxiv: Feb. 3, 2021 2021 (Epub
ahead of print). doi: 10.1101/2021.02.03.429496.
|
|
60
|
Sen P, Ganguly P and Ganguly N: Modulation
of DNA methylation by human papillomavirus E6 and E7 oncoproteins
in cervical cancer. Oncol Lett. 15:11–22. 2018.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Mac M and Moody CA: Epigenetic regulation
of the human papillomavirus life cycle. Pathogens.
9(483)2020.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Shadeo A, Chari R, Lonergan KM, Pusic A,
Miller D, Ehlen T, Van Niekerk D, Matisic J, Richards-Kortum R,
Follen M, et al: Up regulation in gene expression of chromatin
remodelling factors in cervical intraepithelial neoplasia. BMC
Genomics. 9(64)2008.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Kanwal R and Gupta S: Epigenetic
modifications in cancer. Clin Genet. 81:303–311. 2012.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Lu Y, Chan YT, Tan HY, Li S, Wang N and
Feng Y: Epigenetic regulation in human cancer: The potential role
of epi-drug in cancer therapy. Mol Cancer. 19(79)2020.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Feng C, Dong J, Chang W, Cui M and Xu T:
The progress of methylation regulation in gene expression of
cervical cancer. Int J Genomics. 2018(8260652)2018.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Rathinasamy B and Velmurugan BK: Role of
lncRNAs in the cancer development and progression and their
regulation by various phytochemicals. Biomed Pharmacother.
102:242–248. 2018.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Wang X, Wang HK, Li Y, Hafner M, Banerjee
NS, Tang S, Briskin D, Meyers C, Chow LT, Xie X, et al: microRNAs
are biomarkers of oncogenic human papillomavirus infections. Proc
Natl Acad Sci USA. 111:4262–4267. 2014.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Jia W, Wu Y, Zhang Q, Gao GE, Zhang C and
Xiang Y: Expression profile of circulating microRNAs as a promising
fingerprint for cervical cancer diagnosis and monitoring. Mol Clin
Oncol. 3:851–858. 2015.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Kong Q, Tang Z, Xiang F, Jiang J, Yue H,
Wu R and Kang X: Diagnostic value of Serum hsa-mir-92a in patients
with cervical cancer. Clin Lab. 63:335–340. 2017.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Laengsri V, Kerdpin U, Plabplueng C,
Treeratanapiboon L and Nuchnoi P: Cervical cancer markers:
Epigenetics and microRNAs. Lab Med. 49:97–111. 2018.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Gibb EA, Becker-Santos DD, Enfield KS,
Guillaud M, Niekerk Dv, Matisic JP, Macaulay CE and Lam WL:
Aberrant expression of long noncoding RNAs in cervical
intraepithelial neoplasia. Int J Gynecol Cancer. 22:1557–1563.
2012.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Aalijahan H and Ghorbian S: Long
non-coding RNAs and cervical cancer. Exp Mol Pathol. 106:7–16.
2019.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Burgers WA, Blanchon L, Pradhan S, de
Launoit Y, Kouzarides T and Fuks F: Viral oncoproteins target the
DNA methyltransferases. Oncogene. 26:1650–1655. 2007.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Laurson J, Khan S, Chung R, Cross K and
Raj K: Epigenetic repression of E-cadherin by human papillomavirus
16 E7 protein. Carcinogenesis. 31:918–926. 2010.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Leonard SM, Wei W, Collins SI, Pereira M,
Diyaf A, Constandinou-Williams C, Young LS, Roberts S and Woodman
CB: Oncogenic human papillomavirus imposes an instructive pattern
of DNA methylation changes which parallel the natural history of
cervical HPV infection in young women. Carcinogenesis.
33:1286–1293. 2012.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Hsu CH, Peng KL, Jhang HC, Lin CH, Wu SY,
Chiang CM, Lee SC, Yu WC and Juan LJ: The HPV E6 oncoprotein
targets histone methyltransferases for modulating specific gene
transcription. Oncogene. 31:2335–2349. 2012.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Patel D, Huang SM, Baglia LA and McCance
DJ: The E6 protein of human papillomavirus type 16 binds to and
inhibits co-activation by CBP and p300. EMBO J. 18:5061–5072.
1999.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Bernat A, Avvakumov N, Mymryk JS and Banks
L: Interaction between the HPV E7 oncoprotein and the
transcriptional coactivator p300. Oncogene. 22:7871–7881.
2003.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Thomas MC and Chiang CM: E6 oncoprotein
represses p53-dependent gene activation via inhibition of protein
acetylation independently of inducing p53 degradation. Mol Cell.
17:251–264. 2005.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Avvakumov N, Torchia J and Mymryk JS:
Interaction of the HPV E7 proteins with the pCAF acetyltransferase.
Oncogene. 22:3833–3841. 2003.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Huang SM and McCance DJ: Down regulation
of the interleukin-8 promoter by human papillomavirus type 16 E6
and E7 through effects on CREB binding protein/p300 and P/CAF. J
Virol. 76:8710–8721. 2002.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Münger K, Baldwin A, Edwards KM, Hayakawa
H, Nguyen CL, Owens M, Grace M and Huh K: Mechanisms of human
papillomavirus-induced oncogenesis. J Virol. 78:11451–11460.
2004.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Jha S, Vande Pol S, Banerjee NS, Dutta AB,
Chow LT and Dutta A: Destabilization of TIP60 by human
papillomavirus E6 results in attenuation of TIP60-dependent
transcriptional regulation and apoptotic pathway. Mol Cell.
38:700–711. 2010.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Brehm A, Nielsen SJ, Miska EA, McCance DJ,
Reid JL, Bannister AJ and Kouzarides T: The E7 oncoprotein
associates with Mi2 and histone deacetylase activity to promote
cell growth. EMBO J. 18:2449–2458. 1999.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Longworth MS and Laimins LA: The binding
of histone deacetylases and the integrity of zinc finger-like
motifs of the E7 protein are essential for the life cycle of human
papillomavirus type 31. J Virol. 78:3533–3541. 2004.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Longworth MS, Wilson R and Laimins LA:
HPV31 E7 facilitates replication by activating E2F2 transcription
through its interaction with HDACs. EMBO J. 24:1821–1830.
2005.PubMed/NCBI View Article : Google Scholar
|
|
87
|
McLaughlin-Drubin ME, Crum CP and Münger
K: Human papillomavirus E7 oncoprotein induces KDM6A and KDM6B
histone demethylase expression and causes epigenetic reprogramming.
Proc Natl Acad Sci USA. 108:2130–2135. 2011.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Wang Y, Tong J, Lin H, Ma L and Xu Y:
CCHE1 accelerated the initiation of oral squamous cell carcinoma
through enhancing PAK2 expression by sponging miR-922. J Oral
Pathol Med. 49:636–644. 2020.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Sharma S, Mandal P, Sadhukhan T, Roy
Chowdhury R, Ranjan Mondal N, Chakravarty B, Chatterjee T, Roy S
and Sengupta S: Bridging links between long noncoding RNA HOTAIR
and HPV oncoprotein E7 in cervical cancer pathogenesis. Sci Rep.
5(11724)2015.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Zhang J and Gao Y: Long non-coding RNA
MEG3 inhibits cervical cancer cell growth by promoting degradation
of P-STAT3 protein via ubiquitination. Cancer Cell Int.
19(175)2019.PubMed/NCBI View Article : Google Scholar
|
|
91
|
He H, Liu X, Liu Y, Zhang M, Lai Y, Hao Y,
Wang Q, Shi D, Wang N, Luo XG, et al: Human papillomavirus E6/E7
and long noncoding RNA TMPOP2 mutually upregulated gene expression
in cervical cancer cells. J Virol. 93:e01808–18. 2019.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Wang X, Wang HK, McCoy JP, Banerjee NS,
Rader JS, Broker TR, Meyers C, Chow LT and Zheng ZM: Oncogenic HPV
infection interrupts the expression of tumor-suppressive miR-34a
through viral oncoprotein E6. RNA. 15:637–647. 2009.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Melar-New M and Laimins LA: Human
papillomaviruses modulate expression of microRNA 203 upon
epithelial differentiation to control levels of p63 proteins. J
Virol. 84:5212–5221. 2010.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Yao T and Lin Z: MiR-21 is involved in
cervical squamous cell tumorigenesis and regulates CCL20. Biochim
Biophys Acta. 1822:248–260. 2012.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Kottaridi C, Leventakou D, Pouliakis A,
Pergialiotis V, Chrelias G, Patsouri E, Zacharatou A, Panopoulou E,
Damaskou V, Sioulas V, et al: Searching HPV genome for methylation
sites involved in molecular progression to cervical precancer. J
Cancer. 10:4588–4595. 2019.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Hsu YW, Huang RL, Su PH, Chen YC, Wang HC,
Liao CC and Lai HC: Genotype-specific methylation of HPV in
cervical intraepithelial neoplasia. J Gynecol Oncol.
28(e56)2017.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Lorincz AT, Brentnall AR,
Scibior-Bentkowska D, Reuter C, Banwait R, Cadman L, Austin J,
Cuzick J and Vasiljević N: Validation of a DNA methylation HPV
triage classifier in a screening sample. Int J Cancer.
138:2745–2751. 2016.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Gu YY, Zhou GN, Wang Q, Ding JX and Hua
KQ: Evaluation of a methylation classifier for predicting
pre-cancer lesion among women with abnormal results between
HPV16/18 and cytology. Clin Epigenetics. 12(57)2020.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Sahasrabuddhe VV, Luhn P and Wentzensen N:
Human papillomavirus and cervical cancer: Biomarkers for improved
prevention efforts. Future Microbiol. 6:1083–1098. 2011.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Wilting SM and Steenbergen RDM: Molecular
events leading to HPV-induced high grade neoplasia. Papillomavirus
Res. 2:85–88. 2016.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Au Yeung CL, Tsang WP, Tsang TY, Co NN,
Yau PL and Kwok TT: HPV-16 E6 upregulation of DNMT1 through
repression of tumor suppressor p53. Oncol Rep. 24:1599–1604.
2010.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Yeo-Teh NSL, Ito Y and Jha S: High-risk
human papillomaviral oncogenes E6 and E7 target key cellular
pathways to achieve oncogenesis. Int J Mol Sci.
19(1706)2018.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Leonard SM, Wei W, Collins SI, Pereira M,
Diyaf A, Constandinou-Williams C, Young LS, Roberts S and Woodman
CB: Oncogenic human papillomavirus imposes an instructive pattern
of DNA methylation changes which parallel the natural history of
cervical HPV infection in young women. Carcinogenesis.
33:1286–1293. 2012.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Kalantari M, Osann K, Calleja-Macias IE,
Kim S, Yan B, Jordan S, Chase DM, Tewari KS and Bernard HU:
Methylation of human papillomavirus 16, 18, 31, and 45 L2 and L1
genes and the cellular DAPK gene: Considerations for use as
biomarkers of the progression of cervical neoplasia. Virology.
448:314–321. 2014.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Brentnall AR, Vasiljević N,
Scibior-Bentkowska D, Cadman L, Austin J, Szarewski A, Cuzick J and
Lorincz AT: A DNA methylation classifier of cervical precancer
based on human papillomavirus and human genes. Int J Cancer.
135:1425–1432. 2014.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Brentnall AR, Vasiljevic N,
Scibior-Bentkowska D, Cadman L, Austin J, Cuzick J and Lorincz AT:
DNA methylation assay for HPV33 contributes independent triage
information to HPV16, HPV18, HPV31, and EPB41L3 for detecting
cervical pre-cancer. Cancer Biomark. 15:669–675. 2015.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Qiagen: Products: Cervical Cancer
Screening. https://www.qiagen.com/be/products/diagnostics-and-clinical-research/sexual-reproductive-health/cervical-cancer-screening/qiasure-methylation-test-kit-eu/#productdetails.
Accessed June 16, 2021.
|
|
108
|
Dippmann C, Schmitz M, Wunsch K, Schütze
S, Beer K, Greinke C, Ikenberg H, Hoyer H, Runnebaum IB, Hansel A
and Dürst M: Triage of hrHPV-positive women: Comparison of two
commercial methylation-specific PCR assays. Clin Epigenetics.
12(171)2020.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Schmitz M, Eichelkraut K, Schmidt D,
Zeiser I, Hilal Z, Tettenborn Z, Hansel A and Ikenberg H:
Performance of a DNA methylation marker panel using liquid-based
cervical scrapes to detect cervical cancer and its precancerous
stages. BMC Cancer. 18(1197)2018.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Kocsis A, Takács T, Jeney C, Schaff Z,
Koiss R, Járay B, Sobel G, Pap K, Székely I, Ferenci T, et al:
Performance of a new HPV and biomarker assay in the management of
hrHPV positive women: Subanalysis of the ongoing multicenter TRACE
clinical trial (n > 6,000) to evaluate POU4F3 methylation as a
potential biomarker of cervical precancer and cancer. Int J Cancer.
140:1119–1133. 2017.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Kan YY, Liou YL, Wang HJ, Chen CY, Sung
LC, Chang CF and Liao CI: PAX1 methylation as a potential biomarker
for cervical cancer screening. Int J Gynecol Cancer. 24:928–934.
2014.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Liou YL, Zhang TL, Yan T, Yeh CT, Kang YN,
Cao L, Wu N, Chang CF, Wang HJ, et al: Combined clinical and
genetic testing algorithm for cervical cancer diagnosis. Clin
Epigenetics. 8(66)2016.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Witjes JA, Morote J, Cornel EB, Gakis G,
van Valenberg FJP, Lozano F, Sternberg IA, Willemsen E, Hegemann
ML, Paitan Y and Leibovitch I: Performance of the Bladder EpiCheck™
Methylation Test for Patients Under Surveillance for
Non-muscle-invasive Bladder Cancer: Results of a Multicenter,
Prospective, Blinded Clinical Trial. Eur Urol Oncol. 1:307–313.
2018.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Wasserstrom A, Frumkin D, Dotan Z, Bukin
E, Gadish T, Hanuka S, Knirsh R, Darawsha AE and Leibovitch I:
MP13-15 molecular urine cytology-bladder epicheck is a novel
molecular diagnostic tool for monitoring of bladder cancer
patients. J Urol. 195(e140)2016.
|
|
115
|
D'Andrea D, Soria F, Zehetmayer S, Gust
KM, Korn S, Witjes JA and Shariat SF: Diagnostic accuracy, clinical
utility and influence on decision-making of a methylation urine
biomarker test in the surveillance of non-muscle-invasive bladder
cancer. BJU Int. 123:959–967. 2019.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Su SF, de Castro Abreu AL, Chihara Y, Tsai
Y, Andreu-Vieyra C, Daneshmand S, Skinner EC, Jones PA, Siegmund KD
and Liang G: A panel of three markers hyper- and hypomethylated in
urine sediments accurately predicts bladder cancer recurrence. Clin
Cancer Res. 20:1978–1989. 2014.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Van Kessel KE, Beukers W, Lurkin I,
Ziel-van der Made A, van der Keur KA, Boormans JL, Dyrskjøt L,
Márquez M, Ørntoft TF, Real FX, et al: Validation of a DNA
Methylation-Mutation Urine Assay to Select Patients with Hematuria
for Cystoscopy. J Urol. 197:590–595. 2017.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Feber A, Dhami P, Dong L, de Winter P, Tan
WS, Martínez-Fernández M, Paul DS, Hynes-Allen A, Rezaee S, Gurung
P, et al: UroMark-a urinary biomarker assay for the detection of
bladder cancer. Clin Epigenetics. 9(8)2017.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Maier S, Nimmrich I, Koenig T,
Eppenberger-Castori S, Bohlmann I, Paradiso A, Spyratos F, Thomssen
C, Mueller V, Nährig J, et al: DNA-methylation of the homeodomain
transcription factor PITX2 reliably predicts risk of distant
disease recurrence in tamoxifen-treated, node-negative breast
cancer patients-Technical and clinical validation in a multi-centre
setting in collaboration with the European Organisation for
Research and Treatment of Cancer (EORTC) PathoBiology group. Eur J
Cancer. 43:1679–1686. 2007.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Imperiale TF, Ransohoff DF, Itzkowitz SH,
Levin TR, Lavin P, Lidgard GP, Ahlquist DA and Berger BM:
Multitarget stool DNA testing for colorectal-cancer screening. N
Engl J Med. 370:1287–1297. 2014.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Oussalah A, Rischer S, Bensenane M, Conroy
G, Filhine-Tresarrieu P, Debard R, Forest-Tramoy D, Josse T,
Reinicke D, Garcia M, et al: Plasma mSEPT9: A novel circulating
cell-free DNA-based epigenetic biomarker to diagnose hepatocellular
carcinoma. EBioMedicine. 30:138–147. 2018.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Chalasani NP, Ramasubramanian T, Bruinsma
JJ, Allawi HT, Olson M, Roberts LR, Kisiel J, Reddy KR, Lidgard GP,
et al: Combined methylated DNA and protein markers: An accurate
blood-based test for early-stage detection of Hepatocellular
carcinoma. Hepatology. 70 (Suppl 1)(109)2019.
|
|
123
|
Taggart D, Roy D, Li G, Liu D, Zheng L and
Zhang K: DNA methylation biomarkers for noninvassive detection of
hepatocellular carcinoma [abstract]. SITC 2018 Abstracts 263, 2018.
https://sitc.sitcancer.org/2018/abstracts/titles/poster/.
|
|
124
|
Johannessen LE, Brandal P, Myklebust T,
Heim S, Micci F and Panagopoulos I: MGMT Gene Promoter methylation
status-Assessment of two pyrosequencing kits and three
methylation-specific PCR methods for their predictive capacity in
glioblastomas. Cancer Genomics Proteomics. 15:437–448.
2018.PubMed/NCBI View Article : Google Scholar
|
|
125
|
Gilbert MR, Dignam JJ, Armstrong TS, Wefel
JS, Blumenthal DT, Vogelbaum MA, Colman H, Chakravarti A, Pugh S,
Won M, et al: A randomized trial of bevacizumab for newly diagnosed
glioblastoma. N Engl J Med. 370:699–708. 2014.PubMed/NCBI View Article : Google Scholar
|
|
126
|
Waterhouse RL Jr, Van Neste L, Moses KA,
Barnswell C, Silberstein JL, Jalkut M, Tutrone R, Sylora J, Anglade
R, Murdock M, et al: Evaluation of an epigenetic assay for
predicting repeat prostate biopsy outcome in African American men.
Urology. 128:62–65. 2019.PubMed/NCBI View Article : Google Scholar
|
|
127
|
Weiss G, Schlegel A, Kottwitz D, König T
and Tetzner R: Validation of the SHOX2/PTGER4 DNA Methylation
Marker Panel for Plasma-Based Discrimination between Patients with
Malignant and Nonmalignant Lung Disease. J Thorac Oncol. 12:77–84.
2017.PubMed/NCBI View Article : Google Scholar
|
|
128
|
Hao YX, Fu Q, Guo YY, Ye M, Zhao HX, Wang
Q, Peng XM, Li QW, Wang RL and Xiao WH: Effectiveness of
circulating tumor DNA for detection of KRAS gene mutations in
colorectal cancer patients: A meta-analysis. Onco Targets Ther.
10:945–953. 2017.PubMed/NCBI View Article : Google Scholar
|
|
129
|
Moran S, Martínez-Cardús A, Sayols S,
Musulén E, Balañá C, Estival-Gonzalez A, Moutinho C, Heyn H,
Diaz-Lagares A, de Moura MC, et al: Epigenetic profiling to
classify cancer of unknown primary: A multicentre, retrospective
analysis. Lancet Oncol. 17:1386–1395. 2016.PubMed/NCBI View Article : Google Scholar
|
|
130
|
Snoek BC, Splunter APV, Bleeker MCG,
Ruiten MC, Heideman DAM, Rurup WF, Verlaat W, Schotman H, Gent MV,
Trommel NEV and Steenbergen RDM: Cervical cancer detection by DNA
methylation analysis in urine. Sci Rep. 9(3088)2019.PubMed/NCBI View Article : Google Scholar
|
|
131
|
Darwiche N: Epigenetic mechanisms and the
hallmarks of cancer: An intimate affair. Am J Cancer Res.
10:1954–1978. 2020.PubMed/NCBI
|
|
132
|
Locke WJ, Guanzon D, Ma C, Liew YJ,
Duesing KR, Fung KYC and Ross JP: DNA methylation cancer
biomarkers: Translation to the clinic. Front Genet.
10(1150)2019.PubMed/NCBI View Article : Google Scholar
|
|
133
|
Keller S, Ridinger J, Rupp AK, Janssen JW
and Altevogt P: Body fluid derived exosomes as a novel template for
clinical diagnostics. J Transl Med. 9(86)2011.PubMed/NCBI View Article : Google Scholar
|
|
134
|
Meng Y, Sun J, Wang X, Hu T, Ma Y, Kong C,
Piao H, Yu T and Zhang G: Exosomes: A promising avenue for the
diagnosis of breast cancer. Technol Cancer Res Treat.
18(1533033818821421)2019.PubMed/NCBI View Article : Google Scholar
|
|
135
|
Bolha L, Ravnik-Glavač M and Glavač D:
Long Noncoding RNAs as biomarkers in cancer. Dis Markers.
2017(7243968)2017.PubMed/NCBI View Article : Google Scholar
|