Introduction
Noni fruit (Morinda citrifolia) has been
widely used in traditional medicine across tropical and subtropical
regions and is now being paid more attention in Western medicine
(1). Noni has been used for the
treatment of cuts, bruises, bone fractures and other wounds to
promote connective tissue regenerations, and to decrease infection
and inflammation traditionally (2,3).
Currently, noni has also been applied and studied for the treatment
of flu, pain, diabetes, hypertension and cancer (4). In addition, noni has been applied in
the dental field (5-8).
Aqueous extract of noni fruit has an inhibitory effect on
Streptococcus mutans and Streptococcus mitis, which
are associated with dental caries (5). Furthermore, noni juice has been used
in root canal irrigation due to its antimicrobial effects against
Enterococcus faecalis and Candida albicans (7). In a human intervention study, rinsing
and swallowing of noni juice improved periodontitis as a result of
the anti-inflammatory properties of noni juice (6). Advanced periodontitis results in
damaged alveolar bone (8). For the
treatment of periodontitis, regeneration of alveolar bony defects
is important, as well as inflammation control (9). The mineralization properties of noni
also have been studied. In a previous pilot clinical study, bone
turnover increased in a group of postmenopausal women who had
consumed noni juice (10).
However, the effect of noni on gingival-derived stem cells remains
unknown.
Human mesenchymal stem cells possess the ability to
differentiate into multiple tissues, including the bone (11). Among the mesenchymal stem cells,
gingiva-derived stem cells have comparable multilineage
differentiation potential, including osteogenic potential and may
be favorable harvesting stem cells source (12,13).
Therefore, gingiva-derived stem cells are good sources of studying
the effect of noni on mineralization for the treatment of
periodontal defect. In a previous study, noni was applied for stem
cell spheroids (14). It was shown
that noni extract enhanced osteogenic differentiation of stem cell
spheroids. Considering the promising findings in previous studies
(11,14). The present study aimed to
investigate the effects of noni extract on maintaining morphology,
improving cellular viability and strengthening osteogenic
differentiation of stem cells.
Materials and methods
Preparation of noni extract
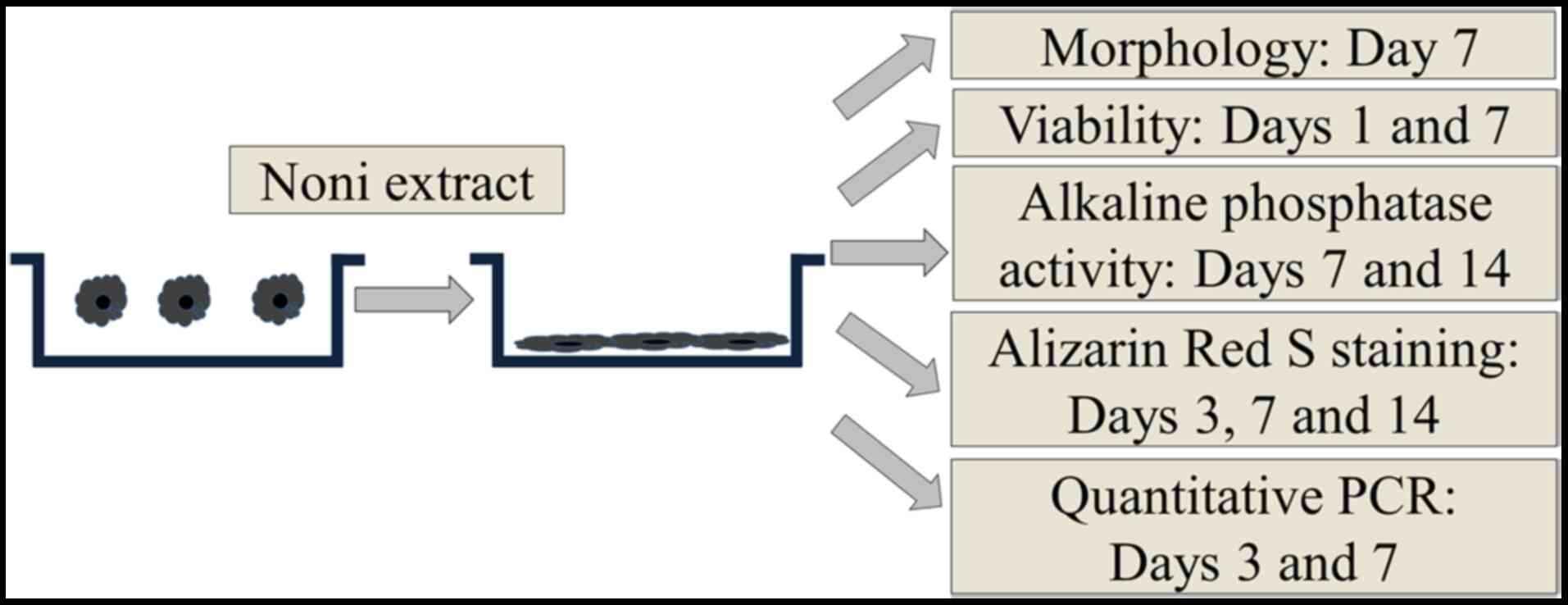
Fig. 1 depicts a
general view of the design of the present study. Noni (Morinda
citrifolia) was obtained from Ngaraard (Ngiwal, Palau) from a
plant >15 years. The noni fruit was ripened at room temperature
(20-28˚C) at a humidity of 60-85% for 11-12 weeks. The noni liquid
was freeze dried using a lyophilizer (Ilshin Lab Co., Ltd.)
(14).
Culturing of gingiva-derived stem
cells and evaluation of the characteristics of stem cell
morphology
Gingival stem cells were acquired from a healthy
patient receiving periodontal treatment, as previously described
(13). The present study was
approved by the Institutional Review Board of The Catholic
University of Korea (approval nos. KC19SESI0234 and KC20SISE0696;
approval date, 07 May 2019) and written informed consent was
provided prior to the study start. The stem cells were plated at
2x103 cells/well into 96-well plates and incubated in
osteogenic media comprising α-MEM (Gibco; Thermo Fisher Scientific,
Inc.), dexamethasone, glycerophosphate disodium salt hydrate,
ascorbic acid 2-phosphate and L-glutamine in the presence of noni,
with final concentrations of 0, 10, 100 and 200 ng/ml (G1, G2, G3
and G4), respectively. The morphological characteristics of the
cells were evaluated using an inverted light microscope at x40
magnification on day 7.
Determining cellular viability
Qualitative analysis of cellular viability was
performed on days 1 and 7 via the commercially available two-color
assay (Live/Dead Kit assay, Molecular Probes; Thermo Fisher
Scientific, Inc.), which determines both the plasma membrane
integrity and esterase activity (15). Quantitative analysis of viable
cells was performed via the Cell Counting Kit-8 assay (Dojindo
Laboratories, Inc.), where absorbance was measured at 450 nm, using
a spectrophotometer (BioTek Instruments, Inc.). Cells were
incubated at room temperature for 1 h.
Evaluation of Alizarin Red S of
alkaline phosphatase activity assays and Alizarin Red S staining
under osteogenic media
Alkaline phosphatase activity was determined on days
7 and 14, after detaching cells using trypsin and Alkaline
Phosphatase Activity Colorimetric Assay kit (cat. no. K412-500;
BioVision Inc.), according to the manufacturer's instructions at
405 nm (16). Cells were washed,
fixed and stained with 2% Alizarin Red S solution (cat. no. 0223;
ScienCell Research Laboratories, Inc.) for 30 min at room
temperature, and quantification of the bound dyes was performed by
applying 10% cetylpyridinium chloride (cat. no. C0732;
Sigma-Aldrich; Merck KGaA) for 15 min at 560 nm (17).
RNA isolation, library preparation,
sequencing and data analysis
Total RNA was extracted from the gingiva-derived
stem cells using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). RNA quality was assessed using Agilent
2100 bioanalyzer (Agilent Technologies, Inc.) and RNA
quantification was performed using a spectrophotometer (ND-2000;
Thermo Fisher Scientific, Inc.). Libraries were prepared from total
RNA using the NEBNext Ultra II Directional RNA-Seq kit (New England
BioLabs, Inc.). The isolated mRNAs were reverse transcribed into
cDNA, according to the manufacturer's instructions (New England
BioLabs, Inc.). Quantification was performed using the library
quantification kit (18) and
library concentration was measured using Tape Station HS D1000
Screen Tape (Agilent Technologies, Inc.). The library loading
volume and loading concentration were 150 µl and 300 pM,
respectively. Sequencing was performed using Illumina Novaseq 6000
(Illumina, Inc.). The NovaSeq 6000 S4 Reagent kit v1.5 (300 cycles;
Illumina, Inc.) was used as the sequencing reagent kit. Running
format was set at PE100bP. Quality control of raw sequencing data
was performed and low quality reads (<Q20) were removed
(19). Estimation of gene
expression levels and normalization of the values were performed
(19,20). Pathway analysis was performed on
differentially expressed genes using the Kyoto Encyclopedia of
Genes and Genomes mapping tool (21). Data mining and graphic
visualization were performed using ExDEGA (Ebiogen, Inc.) (22).
Quantification of RUNX2, BSP, OCN and
COL1A1 mRNA expression levels via quantitative (q)PCR
Total RNA was extracted from the gingiva-derived
stem cells using a commercially available kit (Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions
(17). The quality of RNA was
assessed using a bioanalyzer, and RNA quantity was evaluated using
a spectrophotometer (ND-2000; Thermo Fisher Scientific, Inc.). RNA
was reverse transcribed into cDNA using reverse transcriptase
(SuperScript II; Invitrogen; Thermo Fisher Scientific, Inc.).
mRNA expression levels were detected via qPCR
analysis on days 3 and 7. GenBank was used to design the sense and
antisense primers for PCR. The following primer sequences were
used: RUNX2 (accession no. NM_001015051.3; forward,
5'-CAGTTCCCAAGCATTTCATCC-3' and reverse, 5'-AGG
TGGCTGGATAGTGCATT-3'), BSP (accession no. NM_004967.4; forward,
5'-CCTCTCCAAATGGTGGGTTT-3' and reverse,
5'-ATTCAACGGTGGTGGTTTTC-3'), OCN (accession no. NM_199173.6;
forward, 5'-GGTGCAGAGTC CAGCAAAGG-3' and reverse,
5'-GCGCCTGGGTCTCTTCA CTA-3'), COL1A1 (accession no. NM_000088.4;
forward, 5'-TACCCCACTCAGCCCAGTGT-3' and reverse, 5'-CCGAAC
CAGACATGCCTCTT-3'); and β-actin (accession. no. NM 001101; forward,
5'-AATGCTTCTAGGCGGACTATGA-3' and reverse,
5'-TTTCTGCGCAAGTTAGGTTTT-3'). DNA synthesis and PCR amplification
were performed using a qPCR detection system (StepOnePlus Real-Time
PCR System, Applied Biosystems; Thermo Fisher Scientific, Inc.).
qPCR was performed using 2X Power SYBR-Green Master Mix (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The following
thermocycling conditions were used: 95˚C for 15 min, followed by 40
cycles of 95˚C for 15 sec and Tm at 59˚C for 30 sec. Relative
expression levels were calculated using the 2-ΔΔCq
method (23) and normalized to the
internal reference gene β-actin.
Statistical analysis
Statistical analysis was performed using SPSS 12
software (SPSS, Inc.). All experiments were performed in triplicate
and data are presented as the mean ± standard deviation. The
Shapiro-Wilk test was used to assess normality. One-way ANOVA
followed by Tukey's post hoc test was used to compare differences
between multiple groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Evaluation of stem cell morphology and
determination of cellular viability
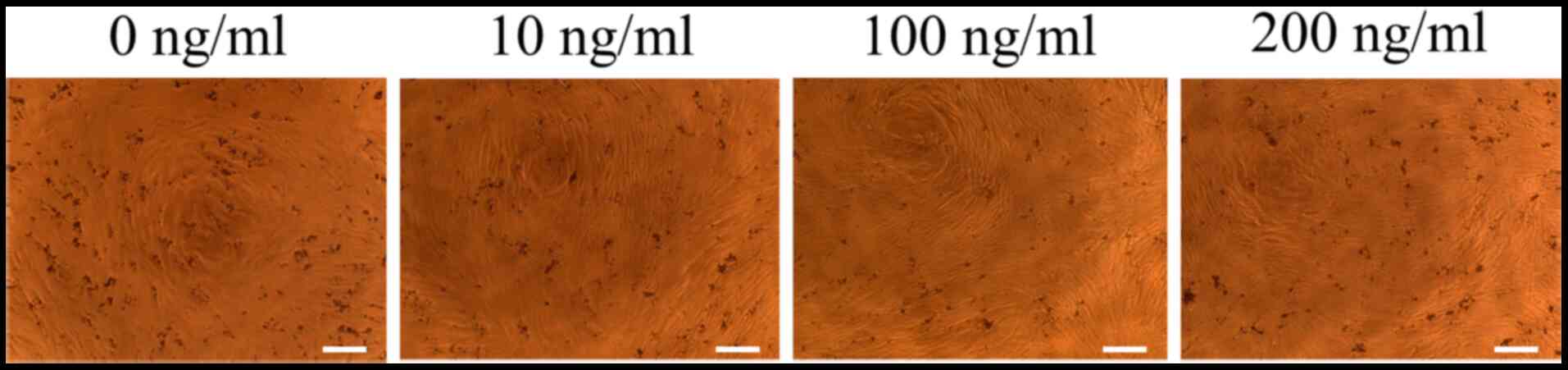
The structures of cells grown in osteogenic media on
day 7 are presented in Fig. 2.
Treatment with noni at 10, 100 and 200 ng/ml did not yield
noticeable morphological changes. The qualitative viability of stem
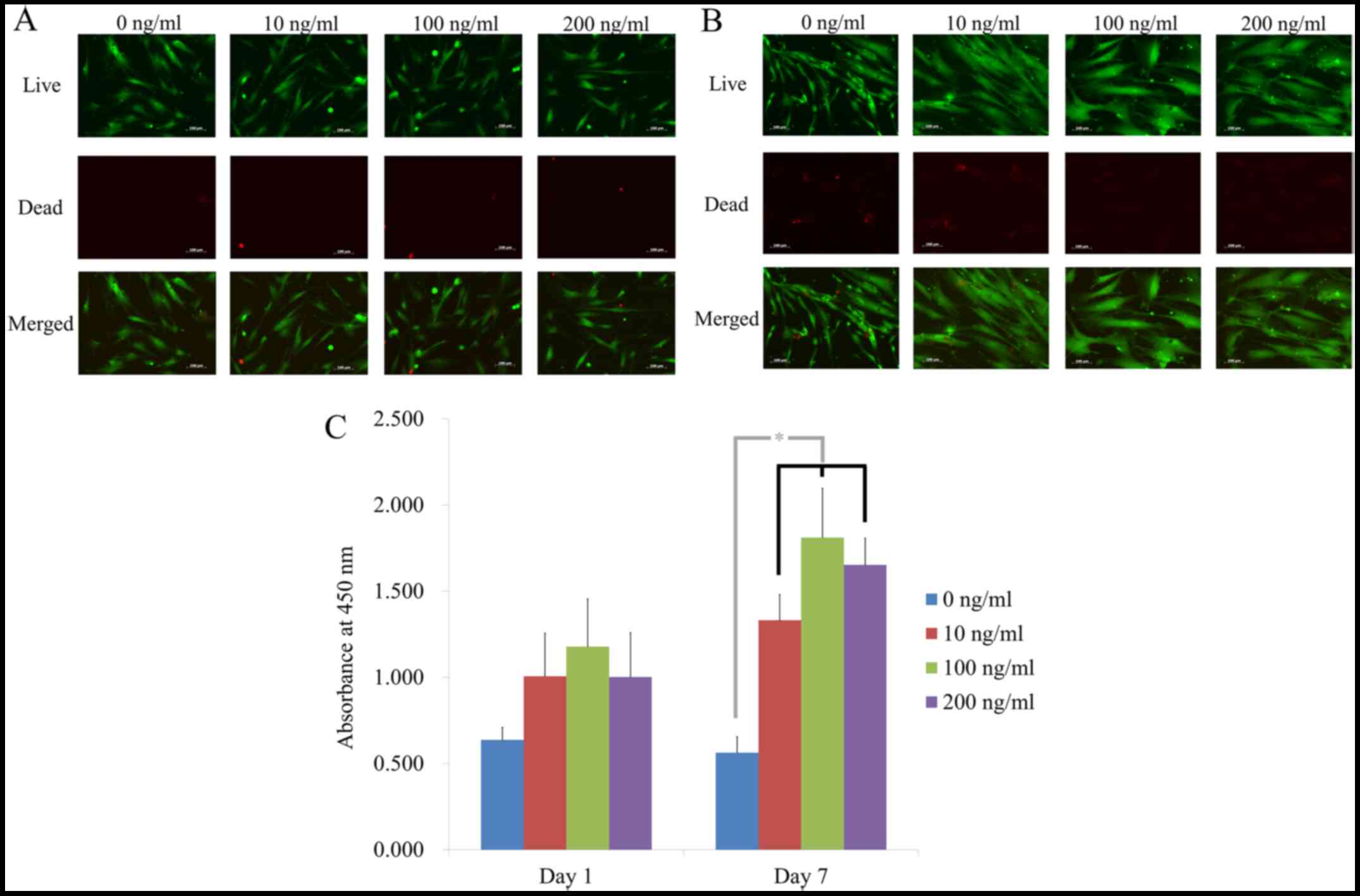
cells was analyzed via a Live/Dead kit assay on days 1 and 7
(Fig. 3A and B). In all cases, most stem cells
exhibited a fibroblast-like shape with intense green fluorescence,
suggesting the presence of live cells on day 1 (Fig. 3A). Longer incubation of cells on
day 7 did not exhibit noticeable decrease in green fluorescence
(Fig. 3B).
Quantitative cellular viability on days 1 and 7 are
presented in Fig. 3C. The
absorbance values at 450 nm on day 1 were 0.636±0.074, 1.006±0.250,
1.178±0.275 and 1.002±0.257 for noni at 0, 10, 100 and 200 ng/ml,
respectively (P>0.05). On day 7, the absorbance values were
0.562±0.093, 1.331±0.150, 1.811±0.286 and 1.652±0.155, for noni at
0, 10, 100 and 200 ng/ml, respectively (P<0.05). Treatment with
noni produced significantly higher values in cellular viability
compared with the 0 ng/ml group (P<0.05), with the highest value
at 100 ng/l.
Evaluation of alkaline phosphatase
activity and Alizarin Red S staining under osteogenic media
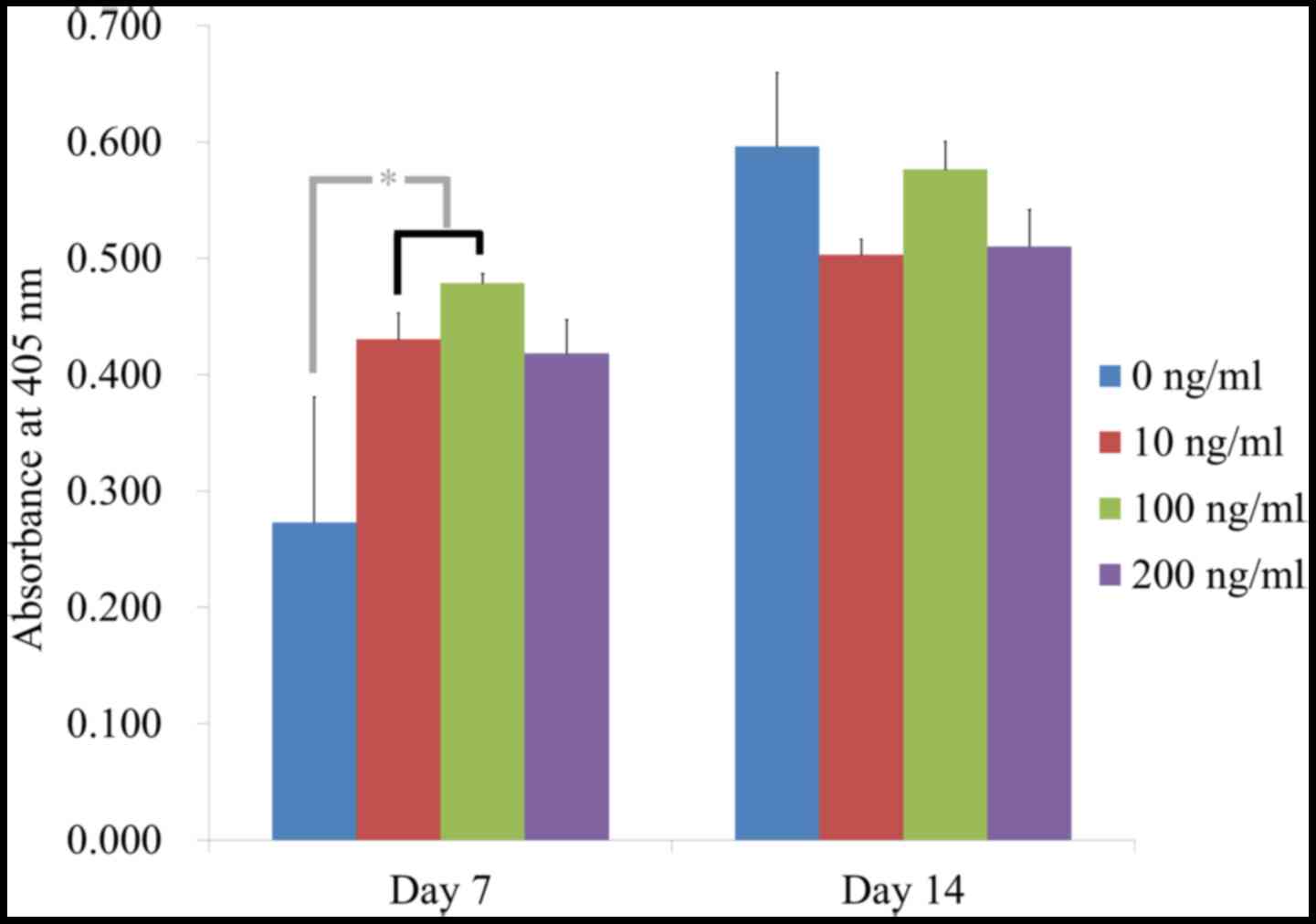
Significantly higher values for alkaline phosphatase
activity were observed in the 10 and 100 ng/ml groups on day 7,
with the highest value at 100 ng/ml (P<0.05; Fig. 4).
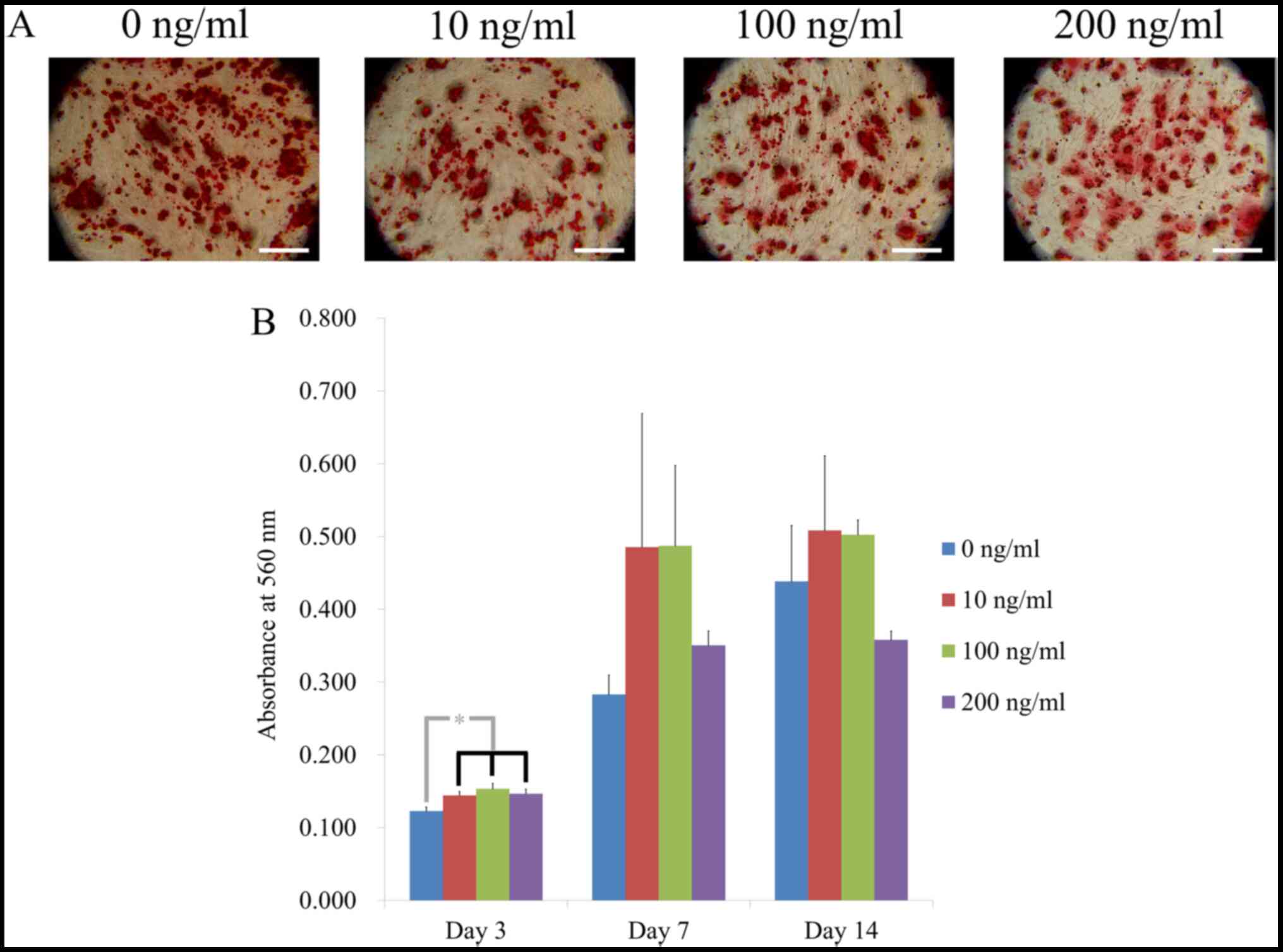
Fig. 5A shows the
results for Alizarin Red S staining. Calcium deposits were clearly
observed in each group on day 7. The absorbance values on day 3
were 0.123±0.006, 0.144±0.006, 0.153±0.007 and 0.147±0.006 for 0,
10, 100 and 200 ng/ml, respectively (Fig. 5B). Treatment with noni produced
significantly higher values in Alizarin Red S staining, with the
highest value from 100 ng/ml (P<0.05).
Gene Ontology
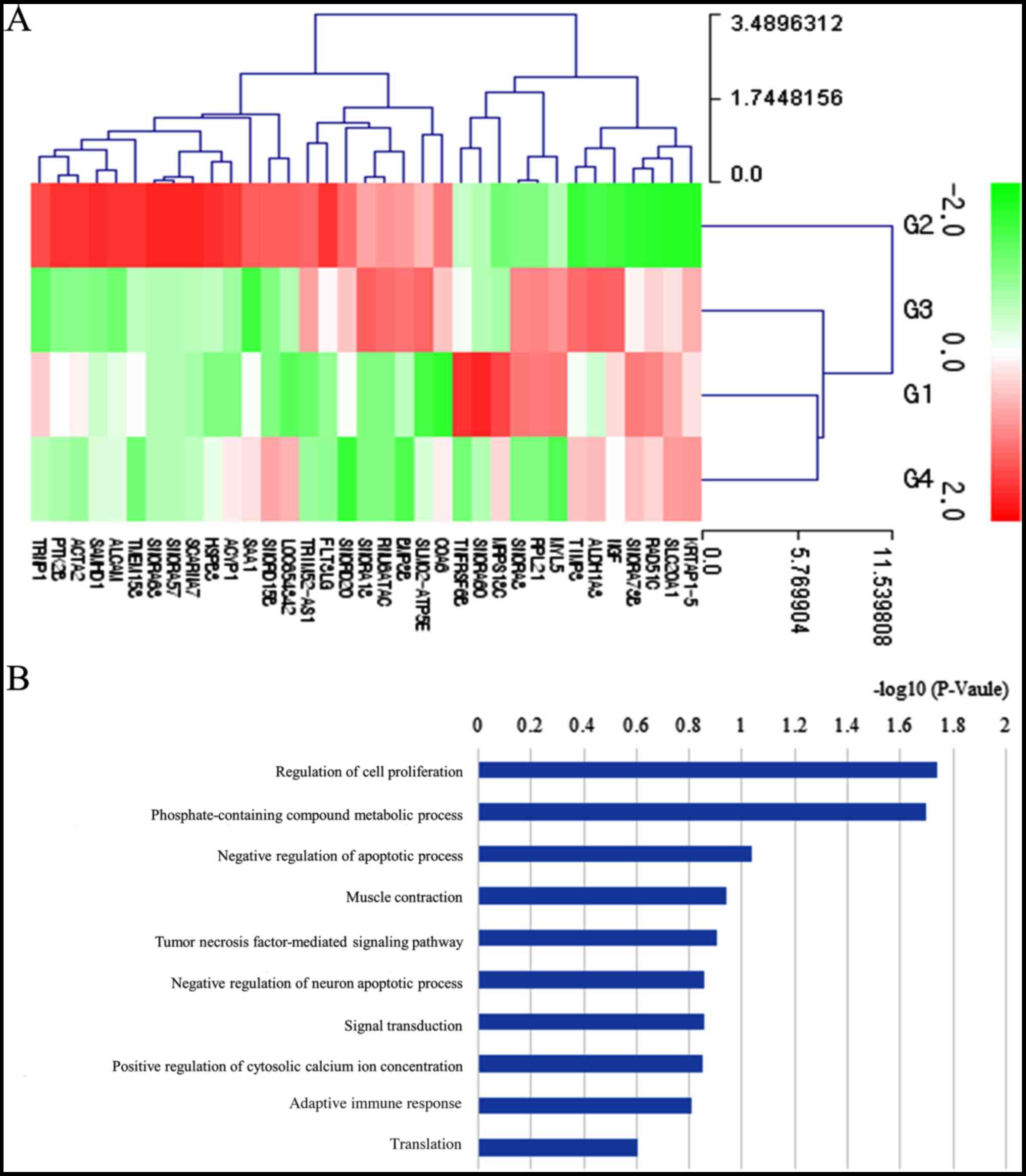
Differentially expressed mRNAs in the 10 ng/ml group
vs. the control group are presented in Table SI (fold change >1.5 and
log3 normalized read counts >3 were selected).
Comparison of the 10 ng/ml group vs. the control group revealed
that 22 mRNAs were upregulated and 13 were downregulated.
Clustering analysis of the differentially expressed mRNAs is
presented in Fig. 6A. Bar charts
depicting the top Gene Ontology terms are presented in Fig. 6B. The most highly associated
pathway was ‘regulation of cell proliferation’.
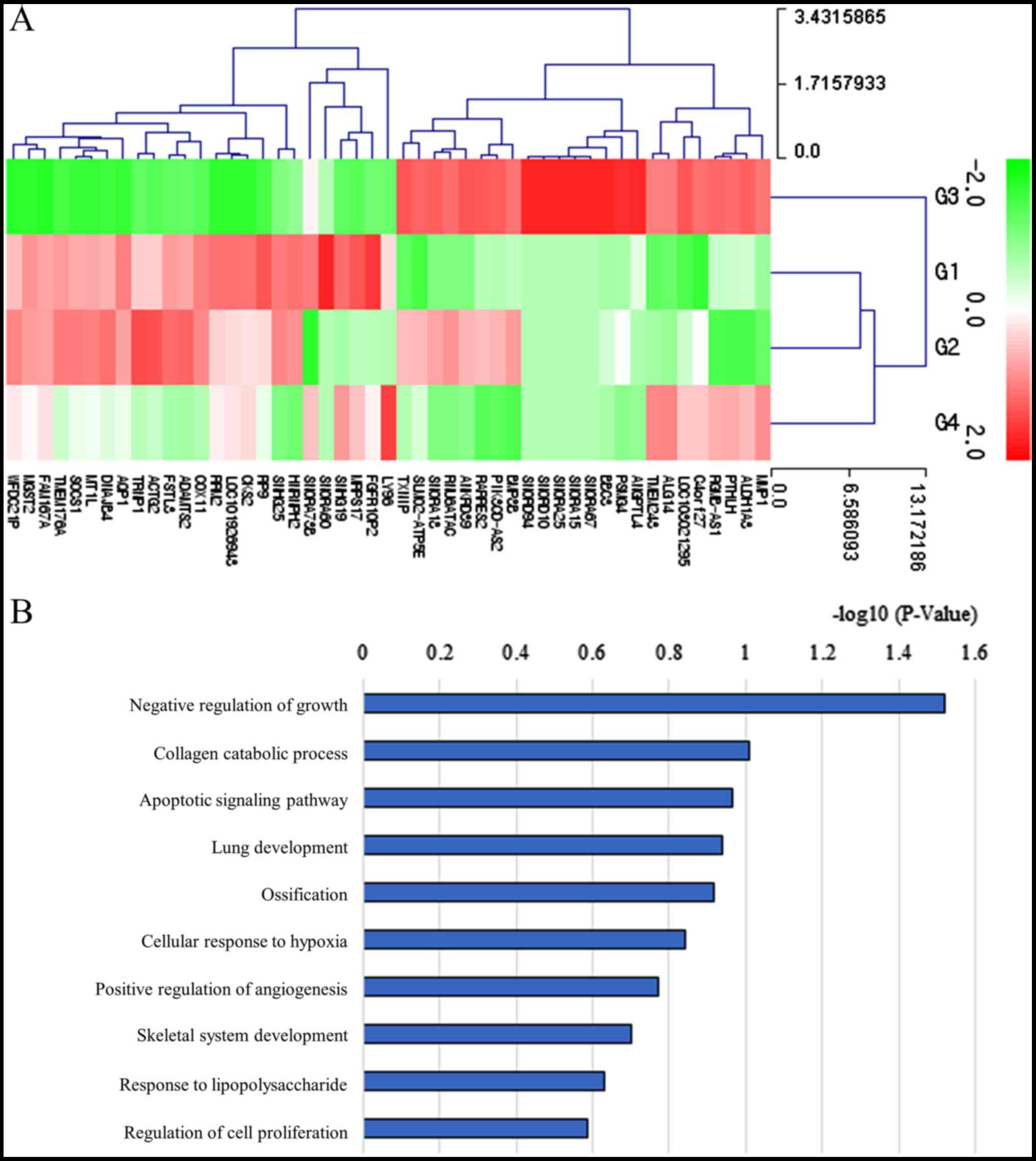
Differentially expressed mRNAs in the 100 ng/ml
group vs. the control group are presented in Table SII (fold change >1.5 and
log2 normalized read counts >3 were selected).
Comparison of the 100 ng/ml group vs. the control group revealed
that 24 mRNAs were upregulated and 25 were downregulated.
Clustering analysis of the differentially expressed mRNAs is
presented in Fig. 7A. Bar charts
depicting the top gene ontology terms are presented in Fig. 7B. ‘Collagen catabolic pathway’ was
associated with mRNA expression.
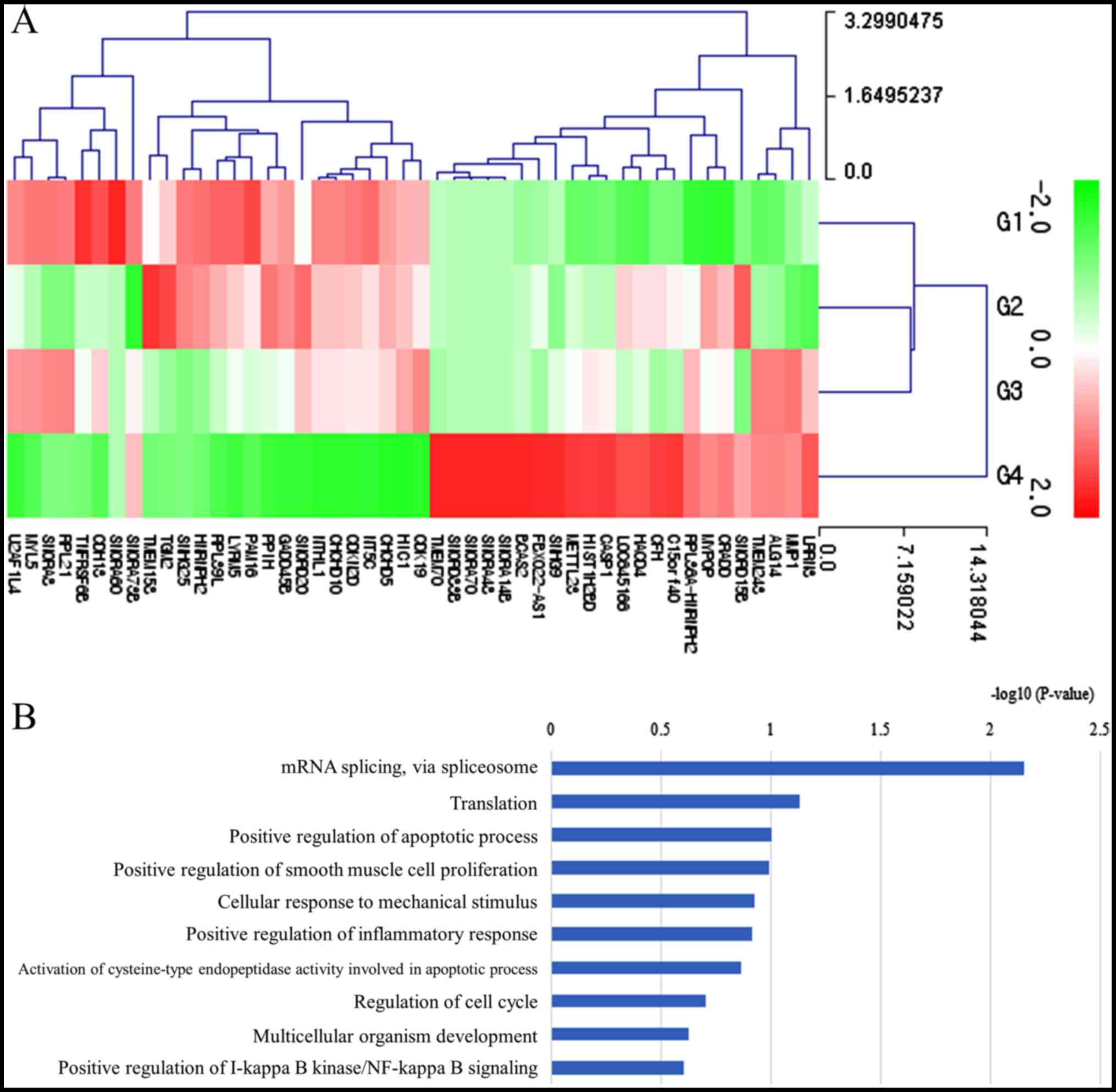
Differentially expressed mRNAs in the 200 ng/ml
group vs. the control group are presented in Table SIII (fold change >1.5 and
log2 normalized read counts >3 were selected).
Comparison of the 200 ng/ml group vs. the control group revealed
that 23 mRNAs were upregulated and 25 were downregulated.
Clustering analysis of the differentially expressed mRNAs is
presented in Fig. 8A. Bar charts
depicting the top gene ontology terms are presented in Fig. 8B. The most highly associated
pathways were ‘mRNA splication’ and ‘translation’. The signaling
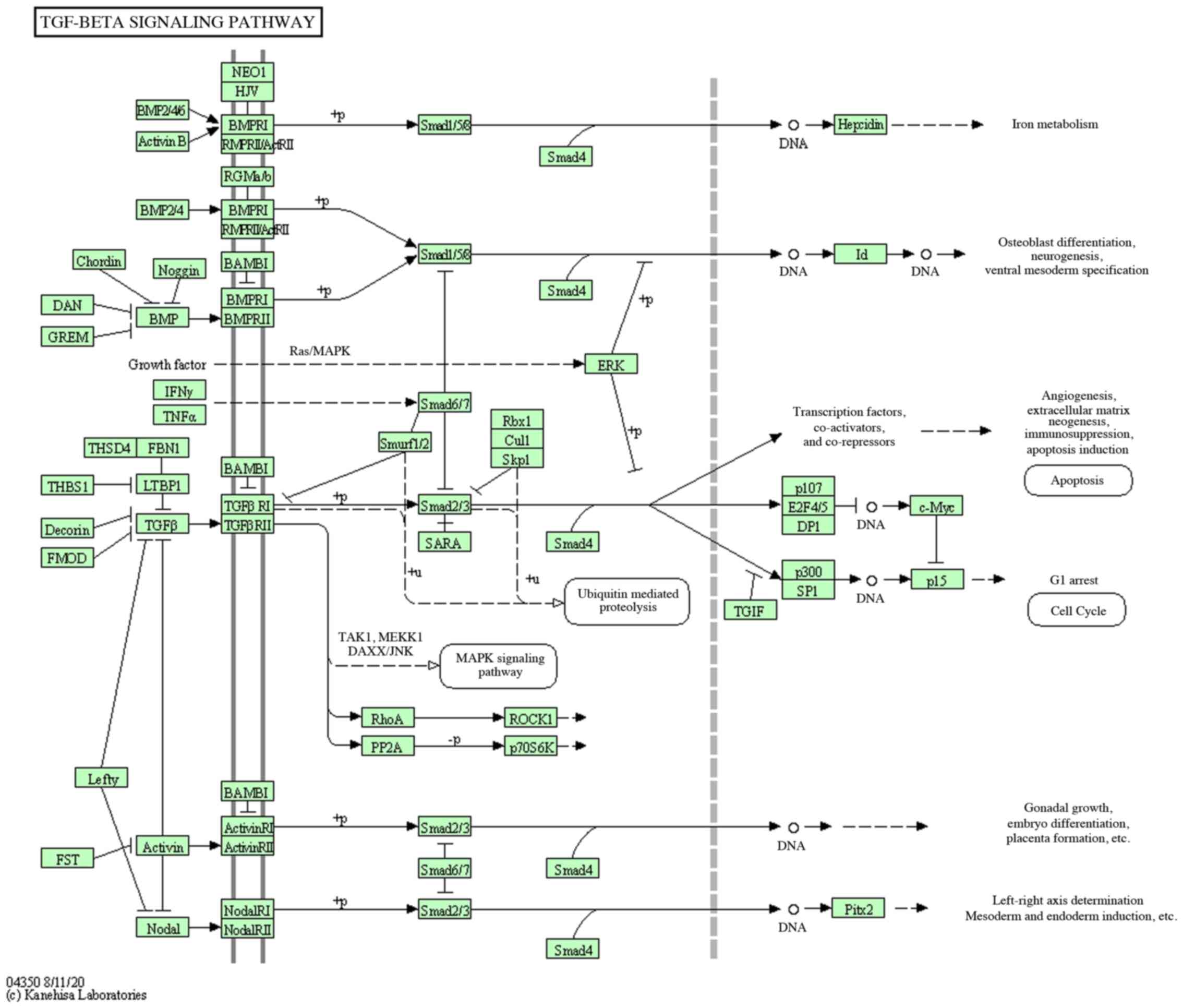
pathway associated with the target genes was transforming growth
factor-β (TGF-β) pathway (Fig.
9).
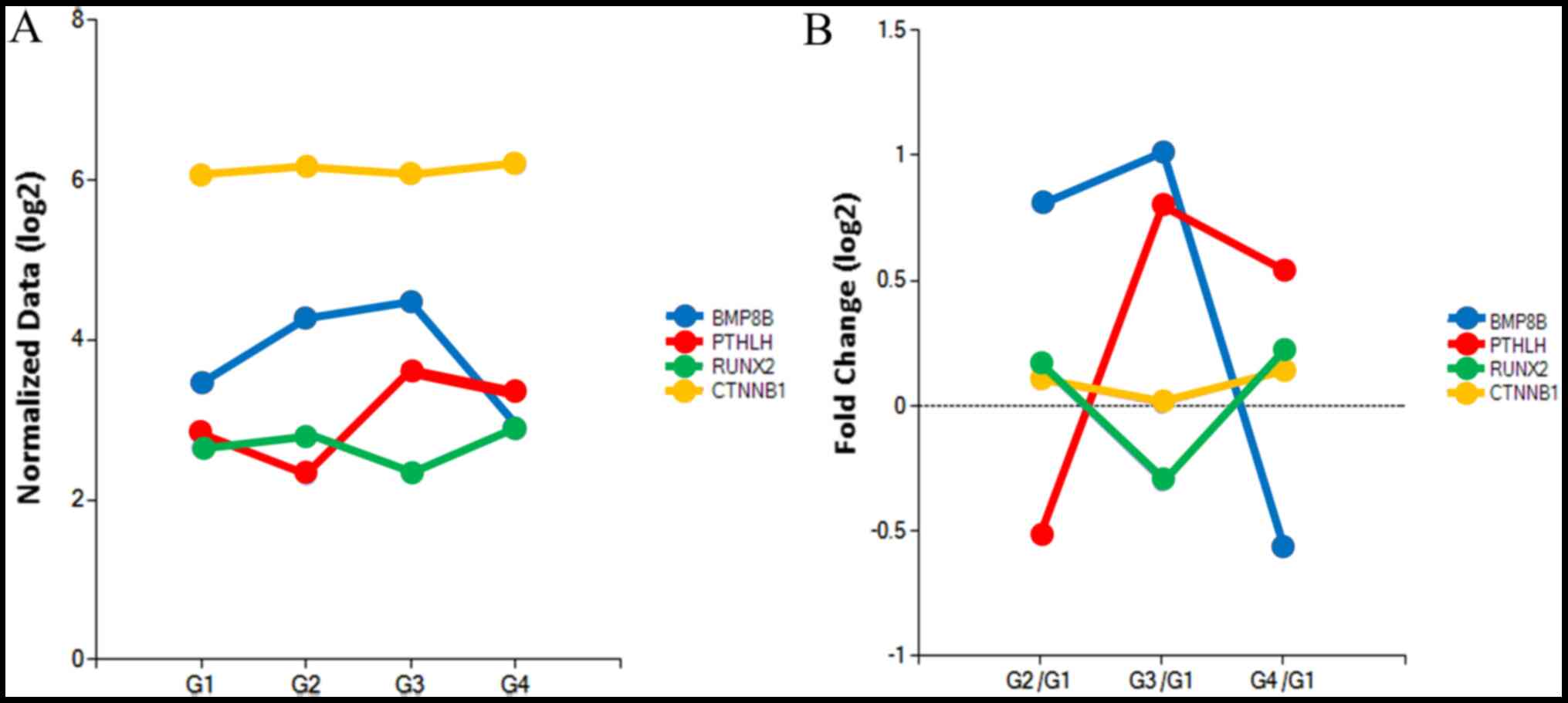
Changes in the expression levels of bone
morphogenetic protein 8b (BMP8B), parathyroid hormone like hormone
(PTHLH), RUNX2 and catenin β-1 (CTNNB1) are presented in Fig. 10A. The results demonstrated that
BMP8B expression was higher in the 10 and 100 ng/ml noni group
compared with the control group (Fig.
10B). Furthermore, RUNX2 expression was higher in the 10 and
200 ng/ml noni groups compared with the control group. Similarly,
CTNNB1 expression was higher in the 10 and 200 ng/ml noni groups
compared with the control group. PTHLH expression was higher in the
100 and 200 ng/ml noni groups compared with the control group.
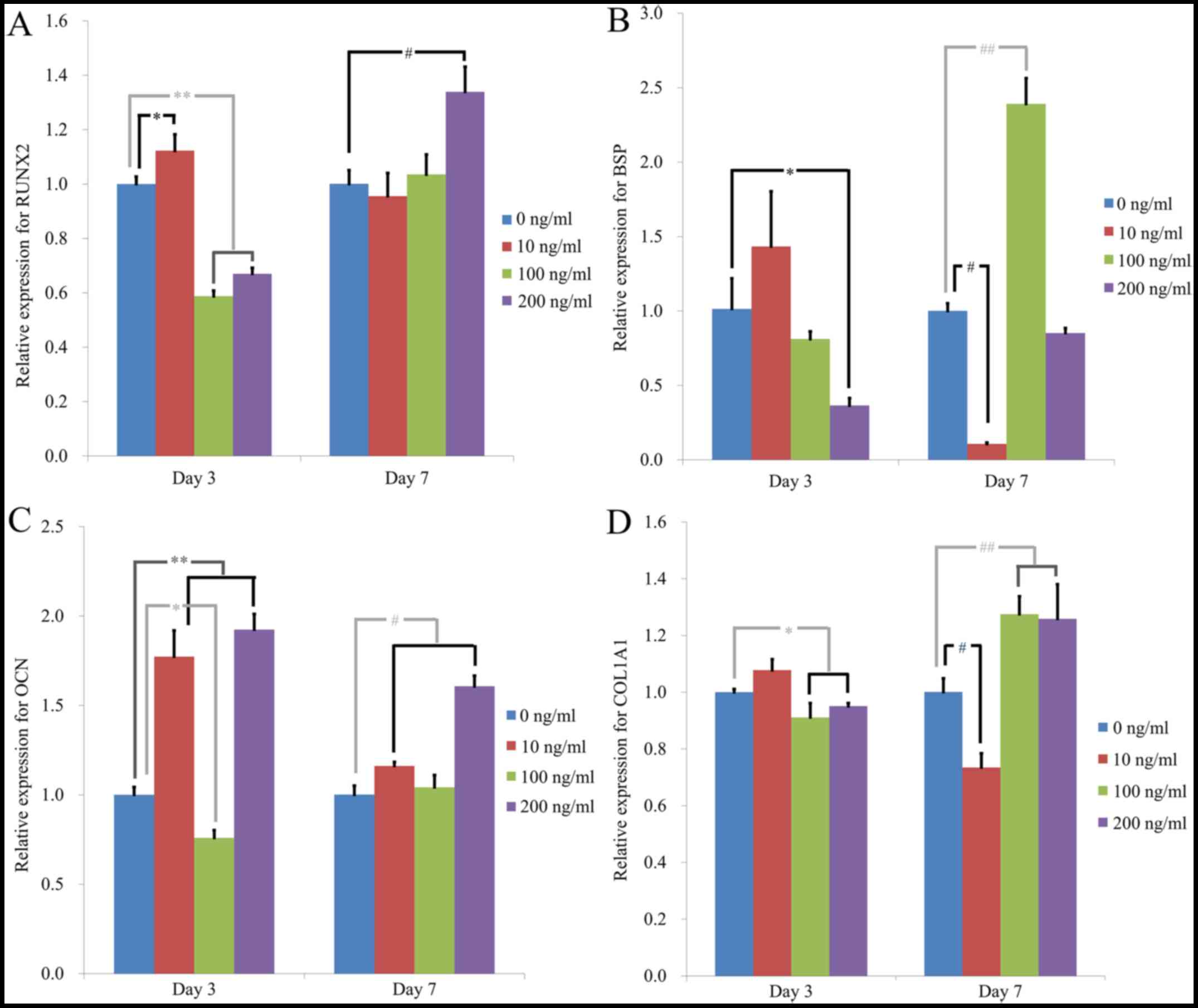
Evaluation of RUNX2, BSP, OCN and
COL1A1 mRNA expression levels via qPCR in osteogenic media
qPCR analysis demonstrated that RUNX2 mRNA
expression levels on day 3 were 1.000±0.027, 1.123±0.060,
0.587±0.021 and 0.670±0.022 for noni at 0, 10, 100 and 200 ng/ml,
respectively (P<0.05; Fig.
11A). RUNX2 mRNA expression levels on day 7 were 1.001±0.050,
0.956±0.085, 1.035±0.073 and 1.339±0.092 for noni at 0, 10, 100 and
200 ng/ml, respectively (P<0.05). Notably, treatment with 200
ng/ml of noni significantly increased RUNX2 expression at day 7.
Furthermore, BSP mRNA expression levels on day 3 were 1.015±0.204,
1.434±0.370, 0.812±0.050 and 0.365±0.049, respectively (P<0.05)
(Fig. 11B). BSP mRNA expression
levels on day 7 were 1.001±0.050, 0.107±0.009, 2.391±0.172 and
0.852±0.033 for noni at 0, 10, 100 and 200 ng/ml, respectively
(P<0.05). Notably, treatment with 100 ng/ml of noni
significantly increased BSP expression at day 7. OCN mRNA
expression levels on day 3 were 1.001±0.043, 1.773±0.146,
0.759±0.043 and 1.924±0.087 for noni at 0, 10, 100 and 200 ng/ml,
respectively (P<0.05; Fig.
11C). OCN mRNA expression levels on day 7 were 1.001±0.051,
1.162±0.022, 1.042±0.069 and 1.607±0.059 for noni at 0, 10, 100 and
200 ng/ml, respectively (P<0.05). Notably, treatment with 10
(day 3) and 200 ng/ml (days 3 and 7) of noni significantly
increased OCN expression. COL1A1 mRNA expression levels on day 3
were 1.000±0.011, 1.078±0.038, 0.911±0.050 and 0.951±0.011 for noni
at 0, 10, 100 and 200 ng/ml, respectively (P<0.05) (Fig. 11D). COL1A1 mRNA expression levels
on day 7 were 1.001±0.048, 0.735±0.050, 1.275±0.063 and 1.259±0.122
at 0, 10, 100 and 200 ng/ml, respectively (P<0.05). Notably,
treatment with 100 and 200 ng/ml of noni significantly increased
OCN expression at day 7.
Discussion
The present study investigated the effects of noni
extract on maintaining morphology, improving cellular viability and
strengthening osteogenic differentiation of dental stem cells.
Several methods have been applied to noni
extraction, including cold press, methanol, ethanol, ethyl acetate
and hexane extract (2,24), and the extraction method may
influence the results. A previous study reported that ethyl acetate
extract has the highest anticancer activity profile (4). In the present study, ripening and
freeze-drying processes were applied for the experiments. It has
been reported that ripening and aging may influence the antioxidant
capacity and the microbiological flora of noni (25).
The dosage may influence the optimal effects, and
adverse effects have been reported (26,27).
In a previous report, noni at a concentration range of 0.015-1.000%
(w/v%) failed to exhibit a cytotoxic effect on human periodontal
ligament (28). Similarly, noni
was applied at concentrations of 0.0001, 0.0100 and 1.000% to
lymphocytes isolated from lymph nodes (29). Regarding animal models, 50-1,000
mg/kg of noni extract were applied to a rat model (30). For a mouse model, 3,000-5,000 mg/kg
of methanol extract was used (31). For human participants, 2 ml/kg of
noni juice was applied (32). In
the present study, the most effective results were obtained in the
100 ng/ml group. This is consistent with a previous study that
assessed the effects of noni using three-dimensional cultures
(14). However, no noticeable
effects of noni on cellular viability were exhibited, which may be
due to the nature of the culture condition and exchange of
nutrients, gases and waste materials (33,34).
The mechanism of noni extract can be explained based
on a previous report. Noni extract has been reported to enhance the
expression of DNA repair genes (35). In addition, it contains various
phytochemicals, including damnacanthal, myricetin and ursolic acid
(36). Damnacanthal is an
anthraquinone known to target p56Ick, PDGFR, erbB2, EGFR and
insulin receptor, c-Met receptor tyrosine kinases and has
anticancer properties (37,38).
Due to its antioxidant effect and its interaction with the immune
system and inflammation pathways, several health benefits are
associated with noni juice, including periodontal health (39). Phytochemicals of noni fruit have
been reported to selectively regulate age-related immunity and
antioxidant enzyme activities via the ERK pathways of splenic
lymphocytes (36).
The improvement of osteogenic differentiation of
noni has also been investigated. BMP8B is a protein-coding gene
that encodes a secreted ligand of the TGF-β superfamily of proteins
(40). The protein encoded by the
PTHLH gene is a member of the parathyroid hormone family (41). CTNNB1 is a protein-coding gene, and
the protein encoded by this gene is associated with regulating cell
proliferation and adhesion between cells (42,43).
Evaluation of several genes, including RUNX2, BSP,
OCN and COL1A1 were evaluated via qPCR analysis in the present
study. RUNX2 is involved in the regulation of osteogenic
differentiation (44), and RUNX2
mutation reduces osteogenic differentiation (45). BSP is involved in the proliferation
and osteogenic differentiation of stem cells in vitro
(46). Similarly, OCN is
considered a differentiation marker for osteogenic differentiation
of stem cells (47). COL1A1 is an
early marker of osteogenic differentiation and its expression
increases during transformation of osteogenic progenitor cells to
osteoblasts (48). In the present
study, qPCR analysis demonstrated that the mRNA expression levels
of RUNX2, BSP, OCN and COL1A1 increased following treatment with
noni. Similarly, a previous study reported that treatment with noni
juice promoted osteoblast differentiation by increasing RUNX2 and
osteocalcin expression in rat bone marrow stem cells (3). In another study, noni leaf extract
enhanced osteogenic differentiation by inducing the PI3K/Akt
signaling pathway, which contributed to activation of Wnt/β-catenin
in C2C12 cells (28). The results
of the present study suggest that noni fruit extract enhanced
osteogenic differentiation via the TGF-β signaling pathway.
Various side effects of noni have been reported in
clinical cases. For example, herb-to-drug interactions have
reported between phenytoin and the commercial product of noni juice
(49). Furthermore, acute liver
failure has been associated with the consumption of noni juice
(50). Long-term consumption of
noni has been reported to change the coagulation profile and
intraoperative hemorrhage (51).
The limitation of the present study is that the protein expression
levels of BMP8B, PTHLH, RUNX2 and CTNNB1 were not verified and
protein expression may not correlate with mRNA expression.
In conclusion, the results of the present study
demonstrated that treatment with noni extract improved cellular
viability and increased osteogenic differentiation. Based on
current literature and the results presented here, noni extract may
have beneficial effects on dental mesenchymal stem cells by
strengthening the cellular viability and osteogenic
differentiation. These findings may lead to the discovery of
potential natural agents for chronic periodontitis, which may
provide clinical benefits for bone regeneration in patients with
periodontitis.
Supplementary Material
Differentially expressed mRNAs in the
10 ng/ml group vs. the control group (fold change >1.5 and
log2 normalized data >3 were selected).
Differentially expressed mRNAs in the
100 ng/ml group vs. the control group (fold change >1.5 and
log2 normalized data >3 were selected).
Differentially expressed mRNAs in the
200 ng/ml group vs. the control group (fold change >1.5 and
log2 normalized data >3 were selected).
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Research
Foundation of Korea grant funded by the Korea government (MSIT,
grant no. 2020R1A2C4001624).
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in the National Center for
Biotechnology Information Sequence Read Archive database
repository, ncbi.nlm.nih.gov/bioproject/PRJNA768946/.
Authors' contributions
YMS, HJL, SKM, YHP, JKO, JYK and JBP conceived the
present study. YMS, HJL, SKM, YHP, JKO, JYK and JBP performed the
experiments. YMS, HJL, SKM, YHP, JKO, JYK and JBP analyzed the
data. YMS, HJL, SKM, YHP, JKO, JYK and JBP drafted the initial
manuscript. All authors have read and approved the final
manuscript. YHP and JBP confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Review Board of The Catholic University of Korea after reviewing
the protocol (approval nos. KC19SESI0234 and KC20SISE0696; approval
date, 07 May 2019) and written informed consent was provided prior
to the study start.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wall MM, Miller S and Siderhurst MS:
Volatile changes in Hawaiian noni fruit, Morinda citrifolia
L., during ripening and fermentation. J Sci Food Agric.
98:3391–3399. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Palu A, Su C, Zhou BN, West B and Jensen
J: Wound healing effects of noni (Morinda citrifolia L.)
leaves: A mechanism involving its PDGF/A2A receptor ligand binding
and promotion of wound closure. Phytother Res. 24:1437–1441.
2010.PubMed/NCBI View
Article : Google Scholar
|
|
3
|
Hussain S, Tamizhselvi R, George L and
Manickam V: Assessment of the role of Noni (Morinda
citrifolia) juice for inducing osteoblast differentiation in
isolated rat bone marrow derived mesenchymal stem cells. Int J Stem
Cells. 9:221–229. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Sharma K, Pachauri SD, Khandelwal K, Ahmad
H, Arya A, Biala P, Agrawal S, Pandey RR, Srivastava A, Srivastav
A, et al: Anticancer effects of extracts from the fruit of
Morinda Citrifolia (Noni) in breast cancer cell lines. Drug
Res (Stuttg). 66:141–147. 2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Kumarasamy B, Manipal S, Duraisamy P,
Ahmed A, Mohanaganesh S and Jeevika C: Role of aqueous extract of
Morinda citrifolia (Indian noni) ripe fruits in inhibiting
dental caries-causing Streptococcus mutans and
Streptococcus mitis. J Dent (Tehran). 11:703–710.
2014.PubMed/NCBI
|
|
6
|
Glang J, Falk W and Westendorf J: Effect
of Morinda citrifolia L. fruit juice on
gingivitis/periodontitis. Mod Res Inflamm. 2:21–27. 2013.
|
|
7
|
Choudhary E, Indushekar KR, Saraf BG,
Sheoran N, Sardana D and Shekhar A: Exploring the role of
Morinda citrifolia and Triphala juice in root canal
irrigation: An ex vivo study. J Conserv Dent. 21:443–449.
2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Needleman I, Worthington H, Giedrys-Leeper
E and Tucker R: Guided tissue regeneration for periodontal
infra-bony defects. Cochrane Database Syst Rev: Apr 19, 2006 (Epub
ahead of print). doi: 10.1002/14651858.CD001724.pub2.
|
|
9
|
Kim YT, Jeong SN and Lee JH: Effectiveness
of porcine-derived xenograft with enamel matrix derivative for
periodontal regenerative treatment of intrabony defects associated
with a fixed dental prosthesis: A 2-year follow-up retrospective
study. J Periodontal Implant Sci. 51:179–188. 2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Langford J, Doughty A, Wang M, Clayton L
and Babich M: Effects of Morinda citrifolia on quality of
life and auditory function in postmenopausal women. J Altern
Complement Med. 10:737–739. 2004.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wang X, Wang Y, Gou W, Lu Q, Peng J and Lu
S: Role of mesenchymal stem cells in bone regeneration and fracture
repair: A review. Int Orthop. 37:2491–2498. 2013.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Lee SI, Ko Y and Park JB: Evaluation of
the maintenance of stemness, viability, and differentiation
potential of gingiva-derived stem-cell spheroids. Exp Ther Med.
13:1757–1764. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Jin SH, Lee JE, Yun JH, Kim I, Ko Y and
Park JB: Isolation and characterization of human mesenchymal stem
cells from gingival connective tissue. J Periodontal Res.
50:461–467. 2015.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Min SK, Oh J and Park JB: The effects of
Morinda citrifolia (Noni) on the cellular viability and
osteogenesis of stem cell spheroids. Medicina (Kaunas).
56(56)2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Kang SH, Park JB, Kim I, Lee W and Kim H:
Assessment of stem cell viability in the initial healing period in
rabbits with a cranial bone defect according to the type and form
of scaffold. J Periodontal Implant Sci. 49:258–267. 2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Kim BB, Tae JY, Ko Y and Park JB:
Lovastatin increases the proliferation and osteoblastic
differentiation of human gingiva-derived stem cells in
three-dimensional cultures. Exp Ther Med. 18:3425–3430.
2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Lee H, Lee H, Na CB and Park JB: The
effects of simvastatin on cellular viability, stemness and
osteogenic differentiation using 3-dimensional cultures of stem
cells and osteoblast-like cells. Adv Clin Exp Med. 28:699–706.
2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Tae JY, Park YH, Ko Y and Park JB: The
effects of bone morphogenetic protein-4 on cellular viability,
osteogenic potential, and global gene expression on gingiva-derived
stem cell spheroids. Coatings. 10(1055)2020.
|
|
19
|
Trapnell C, Pachter L and Salzberg SL:
TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics.
25:1105–1111. 2009.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Roberts A, Trapnell C, Donaghey J, Rinn JL
and Pachter L: Improving RNA-Seq expression estimates by correcting
for fragment bias. Genome Biol. 12(R22)2011.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Kanehisa M, Furumichi M, Tanabe M, Sato Y
and Morishima K: KEGG: New perspectives on genomes, pathways,
diseases and drugs. Nucleic Acids Res. 45:D353–D361.
2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Lee HJ, Min SK, Park YH and Park JB:
Application of bone morphogenetic protein 7 enhanced the osteogenic
differentiation and mineralization of bone marrow-derived stem
cells cultured on deproteinized bovine bone. Coatings.
11(642)2021.
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Huang HL, Liu CT, Chou MC, Ko CH and Wang
CK: Noni (Morinda citrifolia L.) fruit extracts improve
colon microflora and exert anti-inflammatory activities in Caco-2
cells. J Med Food. 18:663–676. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Chan-Blanco Y, Vaillant F, Pérez AM,
Belleville M-P, Zúñiga C and Brat P: The ripening and aging of noni
fruits (Morinda citrifolia L.): Microbiological flora and
antioxidant compounds. J Sci Food Agric. 87:1710–1716. 2007.
|
|
26
|
Singh B and Sharma RA: Indian Morinda
species: A review. Phytother Res. 34:924–1007. 2020.PubMed/NCBI View
Article : Google Scholar
|
|
27
|
Wang M, Wang Q, Yang Q, Yan X, Feng S and
Wang Z: Comparison of anthraquinones, iridoid glycosides and
triterpenoids in Morinda officinalis and Morinda
citrifolia using UPLC/Q-TOF-MS and multivariate statistical
analysis. Molecules. 25(25)2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Gu H, Boonanantanasarn K, Kang M, Kim I,
Woo KM, Ryoo HM and Baek JH: Morinda citrifolia Leaf extract
enhances osteogenic differentiation through activation of
Wnt/β-catenin signaling. J Med Food. 21:57–69. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Pratap UP, Priyanka HP, Ramanathan KR,
Raman V, Hima L and Thyagarajan S: Noni (Morinda citrifolia
L.) fruit juice delays immunosenescence in the lymphocytes in lymph
nodes of old F344 rats. J Integr Med. 16:199–207. 2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Osman WNW and Mohamed S: Standardized
Morinda citrifolia L. and Morinda elliptica L. leaf
extracts alleviated fatigue by improving glycogen storage and
lipid/carbohydrate metabolism. Phytother Res. 32:2078–2085.
2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Pandy V, Wai YC, Amira Roslan NF, Sajat A,
Abdulla Jallb AH and Vijeepallam K: Methanolic extract of
Morinda citrifolia Linn. unripe fruit attenuates
methamphetamine-induced conditioned place preferences in mice.
Biomed Pharmacother. 107:368–373. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Algenstaedt P, Stumpenhagen A and
Westendorf J: The effect of Morinda citrifolia L. fruit
juice on the blood sugar level and other serum parameters in
patients with diabetes type 2. Evid Based Complement Alternat Med.
2018(3565427)2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Park JB, Jeong JH, Lee M, Lee DY and Byun
Y: Xenotransplantation of exendin-4 gene transduced pancreatic
islets using multi-component (alginate, poly-L-lysine, and
polyethylene glycol) microcapsules for the treatment of type 1
diabetes mellitus. J Biomater Sci Polym Ed. 24:2045–2057.
2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Lee H, Min SK, Park YH and Park JB: The
role of insulin-like growth factor-2 on the cellular viability and
differentiation to the osteogenic lineage and mineralization of
stem cells cultured on deproteinized bovine bone mineral. Appl Sci
(Basel). 10(5471)2020.
|
|
35
|
Gupta RK, Bajpai D and Singh N: Influence
of Morinda citrifolia (Noni) on expression of DNA repair
genes in cervical cancer cells. Asian Pac J Cancer Prev.
16:3457–3461. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Pratap UP, Anand K, Yasmine F, Hima L,
Priyanka HP and Thyagarajan S: Phytochemicals in Morinda
citrifolia fruit selectively modulate age-associated immunity
and antioxidant enzyme activities through ERK pathway in splenic
lymphocytes of male F344 rats. J Recept Signal Transduct Res.
36:139–151. 2015.PubMed/NCBI View Article : Google Scholar
|
|
37
|
García-Vilas JA, Quesada AR and Medina MA:
Damnacanthal, a noni anthraquinone, inhibits c-Met and is a potent
antitumor compound against Hep G2 human hepatocellular carcinoma
cells. Sci Rep. 5(8021)2015.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Garcia-Vilas JA, Medina MA, Melo FR,
Pejler G and Garcia-Faroldi G: Damnacanthal inhibits IgE
receptor-mediated activation of mast cells. Mol Immunol. 65:86–93.
2015.PubMed/NCBI View Article : Google Scholar
|
|
39
|
West BJ, Deng S, Isami F, Uwaya A and
Jensen CJ: The potential health benefits of noni juice: A review of
human intervention studies. Foods. 7(7)2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Whittle AJ, Carobbio S, Martins L, Slawik
M, Hondares E, Vázquez MJ, Morgan D, Csikasz RI, Gallego R,
Rodriguez-Cuenca S, et al: BMP8B increases brown adipose tissue
thermogenesis through both central and peripheral actions. Cell.
149:871–885. 2012.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Pieles O, Reck A and Morsczeck C: High
endogenous expression of parathyroid hormone-related protein
(PTHrP) supports osteogenic differentiation in human dental
follicle cells. Histochem Cell Biol. 154:397–403. 2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Stuss M, Migdalska-Sek M,
Brzezianska-Lasota E, Michalska-Kasiczak M, Bazela P and Sewerynek
E: Assessment of Wnt pathway selected gene expression levels in
peripheral blood mononuclear cells (PBMCs) of postmenopausal
patients with low bone mass. Bosn J Basic Med Sci. 21:461–470.
2021.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Kim BB, Kim M, Park YH, Ko Y and Park JB:
Short-term application of dexamethasone on stem cells derived from
human gingiva reduces the expression of RUNX2 and β-catenin. J Int
Med Res. 45:993–1006. 2017.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Komori T: Regulation of osteoblast
differentiation by Runx2. Adv Exp Med Biol. 658:43–49.
2010.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Liu Y, Wang Y, Sun X, Zhang X, Wang X,
Zhang C and Zheng S: RUNX2 mutation reduces osteogenic
differentiation of dental follicle cells in cleidocranial
dysplasia. Mutagenesis. 33:203–214. 2018.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Xia B, Wang J, Guo L and Jiang Z: Effect
of bone sialoprotein on proliferation and osteodifferentiation of
human bone marrow-derived mesenchymal stem cells in vitro.
Biologicals. 39:217–223. 2011.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Lee H, Song Y, Park YH, Uddin MS and Park
JB: Evaluation of the effects of Cuminum cyminum on cellular
viability, osteogenic differentiation and mineralization of human
bone marrow-derived stem cells. Medicina (Kaunas).
57(57)2021.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Kannan S, Ghosh J and Dhara SK: Osteogenic
differentiation potential of porcine bone marrow mesenchymal stem
cell subpopulations selected in different basal media. Biol Open.
9(9)2020.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Kang YC, Chen MH and Lai SL: Potentially
unsafe herb-drug interactions between a commercial product of noni
juice and phenytoin - a case report. Acta Neurol Taiwan. 24:43–46.
2015.PubMed/NCBI
|
|
50
|
Garrido-Gallego F, Muñoz-Gómez R,
Muñoz-Codoceo C, Delgado-Álvarez P, Fernández-Vázquez I and
Castellano G: Acute liver failure in a patient consuming Herbalife
products and Noni juice. Rev Esp Enferm Dig. 107:247–248.
2015.PubMed/NCBI
|
|
51
|
Panditrao MM, Panditrao MM, Edghill F and
Lockhart HF: Can Chronic consumption of noni (Morinda
citrifolia) juice lead to changes in the coagulation profile,
deranged liver function and excessive intraoperative haemorrhage?
West Indian Med J. 63:814–815. 2014.PubMed/NCBI View Article : Google Scholar
|