Introduction
Diabetic nephropathy (DN)-associated end-stage renal
disease (ESRD) is a growing public health concern worldwide
(1). Over the past three decades,
DN-associated ESRD has been identified as a major cause for
dialysis in several countries, including Saudi Arabia, in which
type 2 diabetes mellitus (T2DM) accounts for >33% of all cases
of ESRD (1,2). Several promising studies have
identified genes associated with the occurrence and progression of
DN, including genes associated with microRNAs (miRNAs/miRs)
(3,4). This family of non-coding RNAs is
highly conserved, tissue-specific and short in length (20-24
nucleotides). miRNAs play key roles in numerous physiological and
pathological conditions in human organs, including the kidneys. For
example, upregulation of miR-192, miR-194, miR-204, miR-215 and
miR-216, and downregulation of miR-133a, miR-133b, miR-1d, miR-296,
miR-1a, miR-122 and miR-124a have been observed in human kidneys
(5). Also, miR-192 has been
reported to be 20-fold higher expressed in cortical kidney tissue
compared with medullary kidney tissue, where it is implicated in
sodium transport regulation; in addition, miR-155 could suppress
the expression of the angiotensin II receptor type 1, which could
influence the blood pressure (5,6). By
binding to mRNA targets, miRNAs have the ability to cause
translational repression, mRNA destabilization and/or degradation
(7). The thermodynamics of
miRNA-mRNA target interactions may be affected by single-nucleotide
polymorphisms (SNPs) occurring in the precursor-miRNA (pre-miRNAs),
resulting in target gene dysregulation and, subsequently, phenotype
variations or disease susceptibility (8). The miRNA-499a gene (MIR499A) is
located in the human myosin heavy chain 7B cardiac muscle β gene on
chromosome 20q11.22 34990376-34990497 (+) and consists of a single
exon that encodes for a single transcript (MI0003183). It is 122
base pairs in length and was reported to be a marker of
cardiomyocyte injury (9). Although
the number of studies focusing on the precise role of miR-499a in
the pathogenesis and development of ESRD due to DN is limited,
miR-499a has been proposed to affect several biological processes,
such as cellular senescence, inflammation, apoptosis and immune
response (10), that may play roles
in ESRD biogenesis (Table SI)
(11-14).
Moreover, it was previously reported that miR-499a upregulation may
regulate insulin resistance, glycogenesis and modulate insulin
signaling by inhibition of phosphatase and tensin homolog (15).
A common variant (rs3746444), which is located
within the miR-499a-3p seed region AC(A/G)UCAC at the position
20:34,990,448 (GRCh38), has been reported to be associated with
several diseases, particularly in the Middle Eastern population,
such as cardiovascular disease (9),
autoimmune diseases (16) and
cancer (17). In our previous
study, significant dysregulation of circulating miR-499a levels was
identified in the current DM-associated ESRD cohort (18). Therefore, the present study aimed to
explore the association of the MIR499A (A/G) seed region variant
(rs3746444) with DN-associated ESRD susceptibility in patients with
T2DM, and to determine whether there is an association between the
different genotypes and the patients' clinic-laboratory data. To
the best of our knowledge, this is the first study to investigate
the potential association of the specified variant with ESRD
susceptibility in a Middle Eastern patient sample.
Materials and methods
Study population
A total of 90 consecutive patients with
T2DM-associated ESRD on regular hemodialysis (three times/week)
were recruited as the present study's patient group. The patients
were attending the Nephrology Center of Mohammed bin Saud Al-Kabeer
for renal dialysis at Arar Central Hospital (Arar, Saudi Arabia).
The inclusion/exclusion criteria for the ESRD patient group
selection have been described in detail in our previous work on the
same ESRD patient cohort (18).
Patients presenting with any chronic disease, autoimmune disorders,
cancers and renal disease other than DM-associated nephropathy
(diagnosed by renal biopsy according to the local standard
protocols adopted from the international standards for diagnosis of
non-diabetic renal disease) (19)
were excluded. As it is highly likely that, eventually, a
significant proportion of the patients diagnosed with diabetes
early will develop ESRD, patients with T2DM with a matching
duration of diabetes and normoalbuminuria (urinary
albumin/creatinine ratio, <30 µg/mg) (20,21)
were considered as the control group. All biomedical research
involving human participants conformed to the guidelines of the
Helsinki Declaration. The protocol of the present study was
approved by the Research Ethics Committees of Northern Border
University (approval no. 6/340/H), and written informed consent was
obtained from all participants.
Laboratory analysis
Fasting venous samples were collected on plain and
ethylenediaminetetraacetic acid-vacuum tubes before the specified
second dialysis session for patients with ESRD. Serum was separated
by centrifugation for 12 min at 1,300 x g at room temperature,
immediately from the former tube for biochemical analysis using
commercially available kits for glucose (cat. no. 44044831190),
kidney function tests [creatinine (cat. no. 4810716190) and urea
(cat. no. 4460715190)] and lipid profiling [total cholesterol (cat.
no. 3039773190), triacylglycerol (cat. no. 20767107322), HDL-c
(cat. no. 7528566190) and LDL-c (cat. no. 7005717190)] on a Cobas
Integra Biochemical analyzer (all Roche Diagnostics GmbH).
Genotyping for MIR499A
(rs3746444)
Genomic DNA was extracted from whole blood using a
Wizard Genomic DNA Purification kit (cat. no. A1120; Promega
Corporation) according to the manufacturer's protocol, followed by
DNA concentration and purity assessment using the NanoDrop ND-1000
spectrophotometer (NanoDrop Technologies; Thermo Fisher Scientific,
Inc.) and agarose gel (2%) electrophoresis to assess the DNA
integrity. Samples were subsequently stored at -20˚C until the time
of genotyping. A MIR499A polymorphism assay (rs3746444, assay ID:
C_2142612_30) was run using TaqMan allele discrimination real-time
PCR with the quality measures as described previously (16). In brief, PCR was run in a 25-µl
reaction volume containing gDNA (20 ng) diluted to 11.25 µl with
nucleases-free water, 12.5 µl TaqMan Universal PCR Master Mix (2X)
and 1.25 µl 20 x TaqMan SNP Assay provided from the same supplier.
A no-template and a no-polymerase enzyme samples were run each time
with the study samples as negative controls. Genotyping was
performed in a manner blinded to the case/control status. PCR
amplification was performed on a StepOne Real-Time PCR System
(Applied Biosystems; Thermo Fisher Scientific, Inc.), using the
following conditions: Two-phases initial hold (2 min at 50˚C, and
10 min at 95˚C), followed by a 40-cycle two-step PCR (denaturation
for 15 sec at 95˚C and annealing/extension for 1 min at 60˚C).
Allelic discrimination was determined by analysing the fluorescence
data files from each run using automated allele-calling software
(SDS version 1.3.1; Applied Biosystems; Thermo Fisher Scientific,
Inc.; Fig. S1). The overall
genotype call rate was 100%.
Statistical analysis
Sample size and power calculations using G power-3
software (http://www.gpower.hhu.de/)
demonstrated that, with the specified study design, allowable error
rates, α error=0.05, a medium effect size=0.5 and a sample size of
90/group, may yield 91% power of the study. Data distribution and
variance homogeneity were assessed by the Shapiro-Wilk test and
Levene's test, respectively. Continuous data are expressed as the
mean ± standard deviation, and categorical variables are presented
as frequency counts. Data were compared using the χ2
test, while unpaired Student's t-test, one-way ANOVA or
Kruskal-Wallis tests were applied to compare continuous variables.
The deviation of the observed genotype distribution from the
Hardy-Weinberg equilibrium was analyzed using a χ2
goodness-of-fit using the Online Encyclopedia for Genetic
Epidemiology (http://www.oege.org/).
Genotype-specific adjusted odds ratios (ORs) and 95% confidence
intervals (CIs) were calculated for the different genetic models.
The study variables, including age, sex, number of hemodialysis
sessions, presence of hypertension and duration of the disease,
were adjusted. P<0.05 was considered to indicate a statistically
significant difference. SPSS version 23 (IBM Corp.) was used for
the statistical analyses.
MIR499A (rs3746444) variant in silico
analysis
Gene structural and functional analysis were
performed using Ensembl Genomic database (ensembl.org) and miRBase.org;
target prediction was performed using DIANA-microT-CDS v5.0
(http://diana.imis.athena-innovation.gr/DianaTools/index.php?r=microT_CDS/index),
miRBase (http://www.mirbase.org/) and
DIANA-TarBase v7.0 algorithm (http://diana.imis.athena-innovation.gr/DianaTools/index.php?r=tarbase/index)
databases; and annotation clustering and pathway enrichment
analysis were performed using miRPath v3.0 (http://www.microrna.gr/miRPathv3) and all were
extensively detailed in our previous studies (8,12).
Predicted miR-499a targets were compared with the presence of both
A and G alleles using the miR2Go program (http://compbio.uthsc.edu/miR2GO) at a medium
hierarchical filtering level (P<0.05).
Results
Characteristics of the study
population
The baseline characteristics of the study subjects
are summarized in Table I. The mean
age of the patients with ESRD was 46.6±12.3 years for men and
47.0±14.7 years for women. The mean duration of dialysis was
comparable for both sexes in the study participants. There were no
sex-specific differences in the clinical and biochemical
characteristics of the participants, except those associated with
lipid profile parameters, which were worse in female patients
compared with those in men (Table
I).
 | Table IClinical and biochemical
characteristics of patients with ESRD and the corresponding
controls. |
Table I
Clinical and biochemical
characteristics of patients with ESRD and the corresponding
controls.
| | Men | | Women |
|---|
| Clinical
characteristic | Controls
(n=48) | Patients with ESRD
(n=48) | P-value | Controls
(n=42) | ESRD (n=42) | P-value |
|---|
| Age, years | 41.7±15.4 | 46.6±12.3 | 0.112 | 50.5±14.3 | 47.0±14.7 | 0.247 |
| Hemodialysis
duration, years | NA | 3.52±1.68 | NA | NA | 3.95±2.06 | NA |
| eGFR, ml/min/1.73
m2 | 86.4±18.8 | 6.33±2.19 |
<0.001a | 76.5±21.4 | 5.29±1.23 | 0.001a |
| Hypertension, n
(%) | 13 (32.5) | 24 (57.1) | 0.029a | 24 (51.1) | 12 (25.0) | 0.011a |
| Biochemical
findings | | | | | | |
|
FBS,
mmol/l | 4.53±0.43 | 5.57±2.45 | 0.009a | 4.62±0.35 | 5.49±1.38 |
<0.001a |
|
BUN,
mmol/l | 3.94±1.08 | 19.6±8.2 |
<0.001a | 3.38±0.89 | 18.3±6.58 |
<0.001a |
|
Creatinine,
µmol/l | 93.7±16.9 | 848.1±366.4 |
<0.001a | 80.0±18.2 | 781.8±149.1 |
<0.001a |
|
UA,
µmol/l | 267.5±78.8 | 376.3±73.0 |
<0.001a | 238.0±78.7 | 383.7±91.8 |
<0.001a |
|
Total
cholesterol, mmol/l | 4.10±0.76 | 3.53±0.77 | 0.001a | 4.92±1.82 | 3.84±0.86 |
<0.001a |
|
Total
triglyceride, mmol/l | 1.26±0.83 | 1.17±0.40 | 0.554 | 1.02±0.66 | 1.59±0.95 | 0.001a |
|
HDL-c,
mmol/l | 1.01±0.39 | 0.97±0.34 | 0.654 | 1.33±0.44 | 0.89±0.26 |
<0.001a |
|
LDL-c,
mmol/l | 2.68±0.61 | 2.89±1.23 | 0.321 | 3.12±1.45 | 2.75±0.84 | 0.139 |
|
TC/HDL
ratio | 4.57±1.77 | 4.10±1.94 | 0.246 | 3.98±1.56 | 4.61±1.48 | 0.046a |
MIR499A variant (rs3746444) in the
study population
In the study population, the genotype frequencies of
MIR499A (rs3746444) SNP followed the Hardy-Weinberg equilibrium
(P=0.149 in controls and P=0.398 in patients). The minor allele
frequencies for rs3746444*G were 0.28 and 0.42 in controls and
patients with ESRD, respectively. A significant difference in
MIR499A genotypes was observed between the study groups. The
frequency of the GG genotype was 11.1% in controls, compared with
20% in patients with ESRD (P=0.030). Carrying the rs3746444*G
allele conferred a nearly 2-fold increase in the susceptibility to
the development of ESRD, with an OR (95% CI) of 1.82 (1.17-2.83)
under the allelic genetic association model. Consistently,
homozygote/heterozygote individuals (rs3746444*GG/AG) were more
likely to develop ESRD under the homozygous/heterozygous comparison
and dominant models [GG vs. AA: OR=2.41, 95% CI: 1.61-6.68; AG vs.
AA: OR=2.49, 95% CI: 1.41-3.89; (AG + GG) vs. AA: OR=2.30, 95% CI:
1.18-3.90; Table II].
| Table IIGenotype and allele frequencies of
MIR499A (rs3746444) polymorphism in patients with ESRD and the
corresponding controls. |
Table II
Genotype and allele frequencies of
MIR499A (rs3746444) polymorphism in patients with ESRD and the
corresponding controls.
| Genetic model | Genotype | Controls, n=9 0
(%) | Patients with ESRD,
n=90 (%) | P-value | OR (95% CI) |
|---|
| Co-dominant
model | AA | 49 (54.4) | 32 (35.6) | 0.030a | 1.0 |
| | AG | 31 (34.5) | 40 (44.4) | | 2.49
(1.41-3.89)a |
| | GG | 10 (11.1) | 18 (20.0) | | 2.41
(1.61-6.68)a |
| HWE P-value | | 0.149 | 0.398 | | |
| Dominant model | AA | 49 (54.4) | 32 (35.6) | 0.011a | 1.0 |
| | AG + GG | 41 (45.6) | 58 (64.4) | | 2.30
(1.18-3.90)a |
| Recessive
model | AA + AG | 80 (88.9) | 72 (80.0) | 0.099 | 1.0 |
| | GG | 10 (11.1) | 18 (20.0) | | 1.9
(0.82-4.62) |
| Allelic model | A | 129 (71.7) | 104 (57.8) | 0.006a | 1.0 |
| | G | 51 (28.3) | 76 (42.2) | | 1.82 (1.17-2.83)
a |
MIR499A variant (rs3746444) and
clinic-laboratory characteristics of patients with ESRD
When investigating the association between different
genotypes of the specified variant and the clinical and biochemical
characteristics of the patient group, no significant associations
were demonstrated, as demonstrated in Table III.
| Table IIIAssociation of MIR499A variant with
clinical and biochemical characteristics of patients with ESRD. |
Table III
Association of MIR499A variant with
clinical and biochemical characteristics of patients with ESRD.
| | Rs3746444
genotypes | |
|---|
| Clinical
characteristic | AA (n=32) | AG (n=40) | GG (n=18) | P-value | OR (95% CI) AG/GG
vs. AA |
|---|
| Age, years (%) | | | | 0.385 | |
|
<30 | 3 (9.4) | 4 (10.0) | 0 (0.0) | | 1.0 |
|
≥30 | 29 (90.6) | 36 (90.0) | 18(100) | | 1.39
(0.29-6.66) |
| Sex, n (%) | | | | 0.514 | |
|
Female | 15 (46.9) | 24 (60.0) | 9 (50.0) | | 1.0 |
|
Male | 17 (53.1) | 16 (40.0) | 9 (50.0) | | 0.66
(0.28-1.59) |
| Hemodialysis
duration, years | 3.8±1.8 | 3.8±1.9 | 3.3±1.9 | 0.577 | |
| eGFR, ml/min/1.73
m2 | 5.9±1.9 | 5.8±1.8 | 5.4±1.4 | 0.656 | |
| Hypertension, n
(%) | | | | 0.069 | |
|
Negative | 23 (71.9) | 25 (62.5) | 7 (38.9) | | 1.0 |
|
Positive | 9 (28.1) | 15 (37.5) | 11 (61.1) | | 0.48
(0.19-1.21) |
| Biochemical
findings | | | | | |
|
FBS,
mmol/l | 6.0±2.6 | 5.3±1.4 | 5.2±2.1 | 0.219 | |
|
BUN,
mmol/l | 19.2±7.4 | 17.9±7.7 | 20.9±6.2 | 0.349 | |
|
Creatinine,
µmol/l | 829±226 | 763±302 | 894±271 | 0.220 | |
|
UA,
µmol/l | 371±85.5 | 380±83.9 | 395±80.0 | 0.628 | |
|
Total
cholesterol, mmol/l | 3.82±0.8 | 3.6±0.71 | 3.5±1.1 | 0.520 | |
|
Total
triglyceride, mmol/l | 1.4±0.6 | 1.3±0.6 | 1.5±1.1 | 0.500 | |
|
HDL-c,
mmol/l | 0.9±0.3 | 0.9±0.3 | 0.8±.2 | 0.821 | |
|
LDL-c,
mmol/l | 2.7±0.9 | 2.7±0.9 | 3.0±1.4 | 0.676 | |
|
TC/HDL
ratio | 4.6±2.0 | 41±1.3 | 4.2±1.8 | 0.527 | |
Effect of MIR499A (rs3746444) on gene
structure and function
Although miR-499a forms a secondary hairpin loop
with complementary sequences in its structure (Fig. S2), different genes were predicted
to be targeted by both mature forms synthesized from either arm. A
total of 1,890 genes were predicted to be affected by miR-499a (919
genes by 3p, 810 by 5p and 161 genes by both), as detailed
previously (8). The most
significant Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways
associated with miR-499a targets that may be involved in DN
progression into ESRD are summarized in Table IV.
| Table IVKey Kyoto Encyclopedia of Genes and
Genomes pathways involving miR-499a targets. |
Table IV
Key Kyoto Encyclopedia of Genes and
Genomes pathways involving miR-499a targets.
| Gene targets of
miR-499a-3p, n | Gene targets of
miR-499a-5p, n | Biological pathway
or disease |
|---|
| 0 | 35 | Adherence
junction |
| 0 | 91 | Focal adhesion |
| 0 | 41 |
Phosphatidylinositol signaling system |
| 47 | 40 | GABAergic
synapse |
| 74 | 73 | FoxO signaling
pathway |
| 0 | 11 | Biosynthesis of
unsaturated fatty acids |
| 0 | 2 | Biotin
metabolism |
| 41 | 33 | TGF-β signaling
pathway |
| 18 | 17 | Mucin type-O-glycan
biosynthesis |
| 62 | 54 | Thyroid hormone
signaling pathway |
| 15 | 0 | Prion disease |
| 61 | 52 | Sphingolipid
signaling pathway |
| 48 | 50 | ErbB signaling
pathway |
| 59 | 0 | TNF signaling
pathway |
| 74 | 0 | Ubiquitin-mediated
proteolysis |
| 5 | 0 | Phenylalanine,
tyrosine and tryptophan biosynthesis |
| 35 | 0 | mTOR signaling
pathway |
| 13 | 0 | Glycosphingolipid
biosynthesis |
| 39 | 0 | ECM-receptor
interaction |
The presence of the rs3746444 SNP within the seed
region of the passenger miR-499a strand may lead to disruption of
targets and the creation of a different set of genes (8,16).
Also, it may shorten the stem-loop structure; thus, affecting the
processing of the entire miRNA.
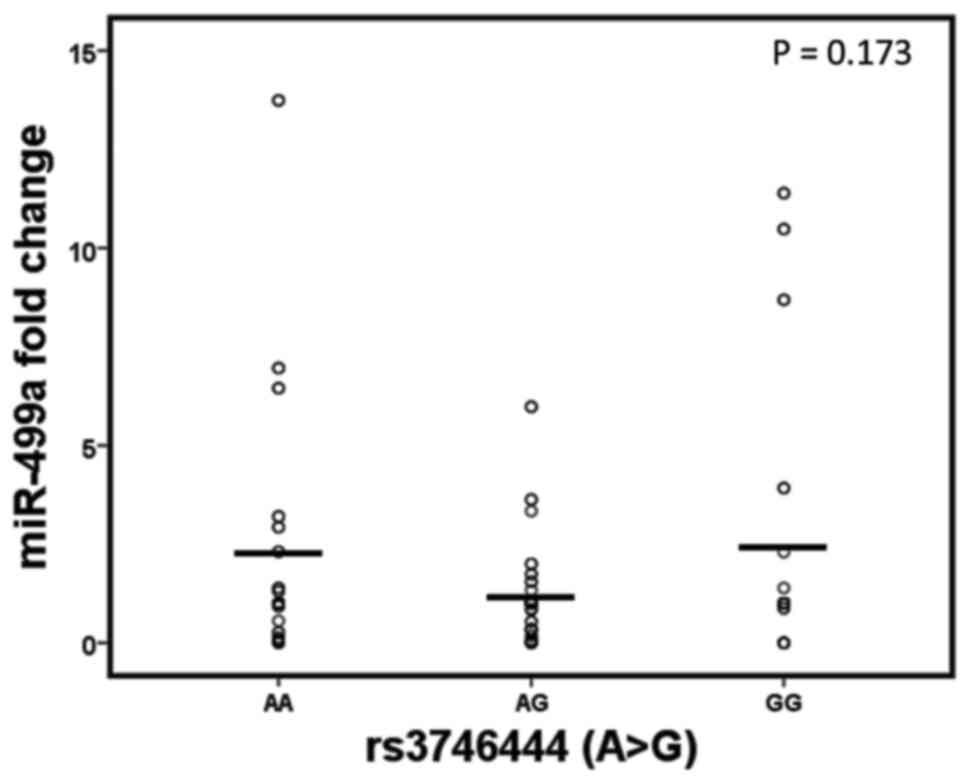
Our previous study reported dysregulation of the
circulatory levels of miR-499a in the same patient cohort (18); therefore, the present study focused
on determining whether these circulatory levels exhibited
significant variations among different genotypes of the same gene.
The findings, however, did not reveal significant associations
between the different genotypes of MIR499A (rs3746444) and the
circulating miR-499a plasma levels detected previously using the
Livak method (22) (P=0.173;
Fig. 1).
Discussion
SNPs have been reported to be the most common type
of genetic variation associated with population diversity, disease
susceptibility and individual therapeutic response (23). SNPs may affect miRNA expression
and/or maturation at the level of transcription of the primary
transcript, post-transcriptional processing of primary transcripts
or pre-miRNAs, or affecting miRNA-mRNA interactions (24).
The present study demonstrated that carrying the
rs3746444*G allele conferred susceptibility to ESRD in patients
with DM compared with non-carriers under the allelic, co-dominant
and dominant genetic association models. Results of our previous
study demonstrated that the rs3746444 polymorphism results in an
A>G mismatch in the miR-499a precursor stem region (located
opposite the mature sequence), affecting the mature miR-499a-3p
seed region implicated in correct miRNA-mRNA binding, with
subsequent disruption and creation of several targets (8). Carriers of the homozygote genotypes
(AA or GG) yield two mature forms, miR-499a-5p and miR-499a-3p*A or
*G, respectively, with each having its specific panel of target
genes. In comparison, the heterozygous (AG) carriers produce three
mature forms, miR-499a-5p, -3p*A and -3p*G, with three subsets of
target genes. The in silico analyses of the present study
revealed that the presence of these different alleles may disrupt
and create 667 and 744 gene targets, respectively. Among validated
gene targets that have been investigated are nucleolar protein 4,
nuclear receptor interacting protein-1, Bcl-2-like protein 14,
chemokine (C-C motif) ligand 8 and sex-determining region Y-box
4(20). To the best of our
knowledge, this is the first study to investigate the MIR499A
rs3746444 variant for ESRD susceptibility in patients with DM in a
sample of the Middle Eastern population. Consistent with the
findings of the present study, Misra et al (25) reported that the heterozygous and the
homozygous genotypes of MIR-499A rs3746444 conferred risk for ESRD
development, which was induced by other disorders apart from DN,
and may be associated with an almost 3-fold increased risk for
acute allograft rejection in a North Indian population (25).
Given that SNPs in pre-miRNAs may alter miRNA
processing and/or expression, on analyzing the association of
MIR499A polymorphism with miR-499a circulating plasma levels and
different clinic-laboratory characteristics of patients, no
significant associations were observed, as previously described. It
is hypothesized that the presence of a miR-499a variant may affect
the thermodynamics of the RNA-RNA interactions and the affinity of
this miRNA by interfering with optimal binding to target mRNAs,
thereby resulting in the dysregulation of target genes that mediate
disease susceptibility to ESRD without affecting the relative
expression and/or disease phenotype (22). The in silico analyses of the
present study revealed some of the top significant KEGG pathways
involving miR-499a targets that may be involved in DN progression
into ESRD (26-32).
For example, the (TGF-β1) pathway was implicated in DN
etiopathology (26,28). The TGF-β1 pathway activates key
signaling pathways, including the PI3K/AKT pathway that mediates
the phosphorylation and inactivation of the transcriptional FOXO,
leading to oxidant stress, mesangial cell expansion and
accumulation of extracellular matrix proteins, key components of
DN-associated chronic kidney disease (26). Upregulation of epidermal growth
factor receptor (ErbB) tyrosine kinase activity is important in
mediating several complications of diabetes, including renal
pathologies, cardiac fibrosis and vascular dysfunction (29). Moreover, the role of the TNF pathway
in the progression of DN in diabetes mice and rat models was
evident. For example, using a soluble TNF receptor-2 fusion protein
has been reported to improve the early stage of DN in DM model of
the KK-Ay mouse (30). Also,
TNFR-Fc TNF inhibitor had been found to attenuate the renal
hypertrophy without affecting the metabolic profile in rats with
induced diabetes (31). mTOR has
been implicated in maintaining the glomerular podocyte function;
increased activity contributed to glomerular hypertrophy and
hyperfiltration associated with progressive DN (32).
Notably, the aforementioned pathways were targeted
in numerous experimental investigations to validate their
therapeutic implication in DN (29,30,33,34).
Ziyadeh et al (33) reported
an improvement in glomerular filtration rates induced by the
monoclonal anti-TGF-β antibody in db/db diabetic mice (33). Furthermore, long-term treatment of
streptozotocin-induced diabetic rats with AG825, a specific
inhibitor of receptor tyrosine-protein kinase ErbB-2, was reported
to significantly correct the hyperreactivity of the perfused
mesenteric vascular bed to norepinephrine and the attenuated
responsiveness to carbachol (29).
Inhibition of TNF-α with the soluble TNF receptor (TNFR)2 fusion
protein, etanercept, improved the progression of the early stage of
DN in a diabetic model established in the KK-A(y) mouse, mainly
through inhibition of the anti-inflammatory action of the renal
TNF-α-TNFR2 pathway (30).
Moreover, rapamycin, the first reported compound to inhibit mTOR,
inhibits glucose-induced phosphorylation of p70S6 kinase and its
substrate, S6 ribosomal protein, in mesangial cells, attenuating
the morphological and functional disorders of diabetic kidneys in a
T2DM mouse model (34).
Consistent with the findings of the present study,
Ciccacci et al (35)
revealed the involvement of miR-499a in DN susceptibility in an
Italian T2DM cohort (35). Results
of the aforementioned study revealed that this association was due
to the impact of miR-499a on two apoptotic players, calcineurin and
dynamin-related protein, by which miR-499a dysregulation caused
mitochondrial dysfunction and cell apoptosis, which may have
contributed to disease susceptibility (33). Notably, the latter processes have
been implicated in DN (36,37), which may in part support the
association of the miR-499a variant with ESRD susceptibility in the
present cohort. However, an increasing number of studies involving
larger independent cohorts with different ethnicities are required
to confirm the findings of the present study.
In conclusion, to the best of our knowledge, the
present study was the first to propose the impact of the MIR499A
A/G seed region variant (rs3746444) on DN-associated ESRD
susceptibility. It may be included in the list of molecular
susceptibility genes that may support patient-risk stratification
and early preventive measurement implementations. However, it is
worth noting that the implication of other potential confounders
could not be conclusively ruled out, such as exposure to different
environmental factors (such as type of treatment or nutrition) and
the additional genetic factors. Further larger-sample size and
long-term follow-up replication studies, particularly in
populations with different ethnicities, are warranted. Functional
studies are also required to validate the role of miR-499a in
DN-associated ESRD etiopathology.
Supplementary Material
Allele discrimination plot
(single-nucleotide polymorphism assay, rs3746444). Alleles as
wild-type homozygous A/A (lower right red cluster), heterozygous
A/G (middle green cluster) and mutant homozygous G/G (upper left
blue cluster) were retrieved using allele-calling software.
Secondary stem-loop structure of the
miR-499a precursor, found in Homo sapiens (length, 90
nucleotides). This structure is annotated by seven databases,
including European Nucleotide Archive, miRBase, Ensembl, NCBI
Reference Sequence, Rfam, GENCODE and HUGO Gene Nomenclature
Committee. Human miRNA hsa-mir-449a precursor sequence is a
product of the miR-449a gene (MIR449A). The black nucleic acid
bases are the same type as those present in the template, green
bases indicate that these bases are not the same as the template,
while red base indicates inserted new nucleotide. Data source,
https://rnacentral.org/rna/URS00006A0D74/9606.
miR/miRNA, microRNA.
Summary of the molecular and
biological functions of miR-499a.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MSF, BTAA, EAA, SAAQ, SWK, WA and EAT conceived and
designed the experiments. BTAA and MSF recruited the study samples
and recorded clinical patient data. MSF and BTAA confirm the
authenticity of all the raw data. MSF, BTAA and EAT contributed to
the experiments. EAT contributed to the statistical analysis. All
authors contributed to obtaining the reagents and materials of the
present study. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The Research Ethics Committees of Northern Border
University approved the present study. Patients who participated in
this research exhibited complete clinical data, and written
informed consent was obtained from all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sattar A, Argyropoulos C, Weissfeld L,
Younas N, Fried L, Kellum JA and Unruh M: All-cause and
cause-specific mortality associated with diabetes in prevalent
hemodialysis patients. BMC Nephrol. 13(130)2012.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Al-Sayyari AA and Shaheen FA: End stage
chronic kidney disease in Saudi Arabia. A rapidly changing scene.
Saudi Med J. 32:339–346. 2011.PubMed/NCBI
|
|
3
|
Mooyaart AL: Genetic associations in
diabetic nephropathy. Clin Exp Nephrol. 18:197–200. 2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Lu Z, Liu N and Wang F: Epigenetic
regulations in diabetic nephropathy. J Diabetes Res.
2017(7805058)2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Schena FP, Serino G and Sallustio F:
MicroRNAs in kidney diseases: New promising biomarkers for
diagnosis and monitoring. Nephrol Dial Transplant. 29:755–763.
2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Felekkis K, Touvana E, Stefanou Ch and
Deltas C: MicroRNAs: A newly described class of encoded molecules
that play a role in health and disease. Hippokratia. 14:236–240.
2010.PubMed/NCBI
|
|
7
|
Mashayekhi S, Saeidi Saedi H, Salehi Z,
Soltanipour S and Mirzajani E: Effects of miR-27a, miR-196a2, and
miR-146a polymorphisms on the risk of breast cancer. Br J Biomed
Sci. 75:76–81. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Toraih EA, Hussein MH, Al Ageeli E, Riad
E, AbdAllah NB, Helal GM and Fawzy MS: Structure and functional
impact of seed region variant in MIR-499 gene family in bronchial
asthma. Respir Res. 18(169)2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Fawzy MS, Toraih EA, Hamed EO, Hussein MH
and Ismail HM: Association of MIR-499a expression and seed region
variant (rs3746444) with cardiovascular disease in Egyptian
patients. Acta Cardiol. 73:131–140. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Wang WJ, Cai GY and Chen XM: Cellular
senescence, senescence-associated secretory phenotype, and chronic
kidney disease. Oncotarget. 8:64520–64533. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Sluijter JP, van Mil A, van Vliet P, Metz
CH, Liu J, Doevendans PA and Goumans MJ: MicroRNA-1 and -499
regulate differentiation and proliferation in human-derived
cardiomyocyte progenitor cells. Arterioscler Thromb Vasc Biol.
30:859–868. 2010.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ling TY, Wang XL, Chai Q, Lau TW, Koestler
CM, Park SJ, Daly RC, Greason KL, Jen J, Wu LQ, et al: Regulation
of the SK3 channel by microRNA-499-potential role in atrial
fibrillation. Heart Rhythm. 10:1001–1009. 2013.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Xu Z, Han Y, Liu J, Jiang F, Hu H, Wang Y,
Liu Q, Gong Y and Li X: MiR-135b-5p and MiR-499a-3p promote cell
proliferation and migration in atherosclerosis by directly
targeting MEF2C. Sci Rep. 5(12276)2015.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Gene Ontology Consortium. The Gene
Ontology resource: Enriching a GOld mine. Nucleic Acids Res.
49:D325–D334. 2021.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wang L, Zhang N, Pan HP, Wang Z and Cao
ZY: MiR-499-5p contributes to hepatic insulin resistance by
suppressing PTEN. Cell Physiol Biochem. 36:2357–2365.
2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Toraih EA, Ismail NM, Toraih AA, Hussein
MH and Fawzy MS: Precursor miR-499a Variant but not miR-196a2 is
associated with rheumatoid arthritis susceptibility in an Egyptian
population. Mol Diagn Ther. 20:279–295. 2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Toraih EA, Fawz MS, Elgazzaz MG, Hussein
MH, Shehata RH and Daoud HG: Combined genotype analyses of
precursor miRNA196a2 and 499a variants with hepatic and renal
cancer susceptibility a preliminary study. Asian Pac J Cancer Prev.
17:3369–3375. 2016.PubMed/NCBI
|
|
18
|
Fawzy MS, Abu AlSel BT, Al Ageeli E,
Al-Qahtani SA, Abdel-Daim MM and Toraih EA: Long non-coding RNA
MALAT1 and microRNA-499a expression profiles in diabetic ESRD
patients undergoing dialysis: A preliminary cross-sectional
analysis. Arch Physiol Biochem. 126:172–182. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Ghani AA, Al Waheeb S, Al Sahow A and
Hussain N: Renal biopsy in patients with type 2 diabetes mellitus:
Indications and nature of the lesions. Ann Saudi Med. 29:450–453.
2009.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Nah EH, Cho S, Kim S and Cho HI:
Comparison of urine albumin-to-creatinine ratio (ACR) between ACR
strip test and quantitative test in prediabetes and diabetes. Ann
Lab Med. 37:28–33. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Fawzy MS and Beladi FIA: Association of
circulating vitamin D, VDBP, and vitamin D receptor expression with
severity of diabetic nephropathy in a group of saudi type 2
diabetes mellitus patients. Clin Lab. 64:1623–1633. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Shastry BS: SNPs: Impact on gene function
and phenotype. Methods Mol Biol. 578:3–22. 2009.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Hoffman AE, Zheng T, Yi C, Leaderer D,
Weidhaas J, Slack F, Zhang Y, Paranjape T and Zhu Y: MicroRNA
miR-196a-2 and breast cancer: A genetic and epigenetic association
study and functional analysis. Cancer Res. 69:5970–5977.
2009.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Misra MK, Pandey SK, Kapoor R, Sharma RK
and Agrawal S: Genetic variants of MicroRNA-related genes in
susceptibility and prognosis of end-stage renal disease and renal
allograft outcome among north Indians. Pharmacogenet Genomics.
24:442–450. 2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Kato M, Yuan H, Xu ZG, Lanting L, Li SL,
Wang M, Hu MC, Reddy MA and Natarajan R: Role of the Akt/FoxO3a
Pathway in TGF-beta1-mediated mesangial cell dysfunction: A novel
mechanism related to diabetic kidney disease. J Am Soc Nephrol.
17:3325–3335. 2006.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Qin G, Zhou Y, Guo F, Ren L, Wu L, Zhang
Y, Ma X and Wang Q: Overexpression of the FoxO1 ameliorates
mesangial cell dysfunction in male diabetic rats. Mol Endocrinol.
29:1080–1091. 2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chang AS, Hathaway CK, Smithies O and
Kakoki M: Transforming growth factor-β1 and diabetic nephropathy.
Am J Physiol Renal Physiol. 310:F689–F696. 2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Akhtar S, Yousif MH, Dhaunsi GS, Sarkhouh
F, Chandrasekhar B, Attur S and Benter IF: Activation of ErbB2 and
downstream signalling via Rho kinases and ERK1/2 contributes to
diabetes-induced vascular dysfunction. PLoS One.
8(e67813)2013.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Omote K, Gohda T, Murakoshi M, Sasaki Y,
Kazuno S, Fujimura T, Ishizaka M, Sonoda Y and Tomino Y: Role of
the TNF pathway in the progression of diabetic nephropathy in
KK-A(y) mice. Am J Physiol Renal Physiol. 306:F1335–F1347.
2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Al-Lamki RS and Mayadas TN: TNF receptors:
Signaling pathways and contribution to renal dysfunction. Kidney
Int. 87:281–296. 2015.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Gödel M, Hartleben B, Herbach N, Liu S,
Zschiedrich S, Lu S, Debreczeni-Mór A, Lindenmeyer MT, Rastaldi MP,
Hartleben G, et al: Role of mTOR in podocyte function and diabetic
nephropathy in humans and mice. J Clin Invest. 121:2197–2209.
2011.PubMed/NCBI View
Article : Google Scholar
|
|
33
|
Ziyadeh FN, Hoffman BB, Han DC,
Iglesias-De La Cruz MC, Hong SW, Isono M, Chen S, McGowan TA and
Sharma K: Long-term prevention of renal insufficiency, excess
matrix gene expression, and glomerular mesangial matrix expansion
by treatment with monoclonal anti transforming growth factor-beta
antibody in db/db diabetic mice. Proc Natl Acad Sci USA.
97:8015–8020. 2000.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Mori H, Inoki K, Masutani K, Wakabayashi
Y, Komai K, Nakagawa R, Guan KL and Yoshimura A: The mTOR pathway
is highly activated in diabetic nephropathy and rapamycin has a
strong therapeutic potential. Biochem Biophys Res Commun.
384:471–475. 2009.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ciccacci C, Latini A, Greco C, Politi C,
D'Amato C, Lauro D, Novelli G, Borgiani P and Spallone V:
Association between a MIR499A polymorphism and diabetic neuropathy
in type 2 diabetes. J Diabetes Complications. 32:11–17.
2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Forbes JM and Thorburn DR: Mitochondrial
dysfunction in diabetic kidney disease. Nat Rev Nephrol.
14:291–312. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Flemming NB, Gallo LA and Forbes JM:
Mitochondrial dysfunction and signaling in diabetic kidney disease:
Oxidative stress and beyond. Semin Nephrol. 38:101–110.
2018.PubMed/NCBI View Article : Google Scholar
|